Buxolestes
Buxolestes ist eine heute ausgestorbene Gattung insektenfresserartiger Tiere aus der Gruppe der Pantolestidae. Die Vertreter der Gattung lebten vor allem im Mittleren Eozän vor 48 bis 41 Millionen Jahren und sind von mehreren, zumeist vollständigen Skeletten aus der Grube Messel, aber auch von Funden aus dem Geiseltal und aus Bouxwiller in Frankreich bekannt. Der Skelettbau von Buxolestes ist eher ursprünglich, so dass die Tiere durch einen gebogenen Rückenverlauf und einen sehr langen Schwanz gekennzeichnet sind. Er zeigt aber spezielle Anpassungen an eine teils im Wasser lebende (semiaquatische), teils aber auch unterirdisch grabende (fossoriale) Lebensweise. Ökologisch nahmen die Vertreter von Buxolestes damals die Nische der heutigen Fischotter ein. Anhand von fossilen Magenresten konnten Fische und kleine Wirbeltiere, aber auch pflanzliches Material als Nahrungsreste nachgewiesen werden, so dass Buxolestes wohl analog zu heutigen Mardern und Füchsen eine allesfresserische Ernährungsweise pflegte. Die Erstbeschreibung von Buxolestes erfolgte 1970.
| Buxolestes | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Zeitliches Auftreten | ||||||||||||
| Mittleres Eozän | ||||||||||||
| 47,8 bis 38 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Buxolestes | ||||||||||||
| Jaeger, 1970 | ||||||||||||
Beschreibung
Buxolestes war ein kleines bis mittelgroßes, insektenfresserartiges Tier und besaß eine Kopf-Rumpf-Länge von 36 bis 46 cm. Der für frühe Höhere Säugetiere charakteristische, außerordentlich lange Schwanz maß zwischen 24 und 36 cm und war damit etwa so lang wie der Rumpf. Das Körpergewicht dürfte bei rund 2 kg gelegen haben. Ein weiteres typisches Merkmal stellte die aufgewölbte Rumpfwirbelsäule dar. Der Schädel wies Längen von 7,1 bis 9,3 cm auf. Er war kompakt gebaut und wirkte dadurch eher hoch als lang. Gleiches gilt für das Rostrum, dieses war zudem durch einen großen Naseninnenraum zwischen dem Nasenbein und dem Mittelkieferknochen gekennzeichnet. Das Nasenbein selbst war schlank und deutlich nach hinten verlängert, so dass es bis auf die Höhe der Orbita reichte. Die Augenhöhle war weit nach hinten geöffnet. Das paarige Stirnbein besaß einen kleinen Scheitelkamm, während der Jochbeinbogen entweder nach oben gekrümmt oder relativ gerade verlief. Das weit nach hinten auskragende Hinterhauptsbein wies einen sehr kräftigen Wulst als Ansatzstelle der Nackenmuskulatur sowie kräftige Gelenkenden (Condylen) für die Halswirbelsäule auf.[1][2][3][4]
Der Unterkiefer war massig gebaut und zwischen 4,9 und 6,9 cm lang. Der kräftige Knochenkörper erreichte im Bereich des ersten Molaren etwa 1,1 cm Höhe. Das Gebiss umfasste die vollständige Zahnanzahl der frühen Höheren Säugetiere, die Gebissformel lautete demzufolge: . Die Schneidezähne zeichneten sich durch ungleiche Größen aus. So war im unteren Gebiss der zweite, im oberen der erste und dritte vergrößert, letzterer wirkte durch seine kegelige Form eher wie ein Eckzahn (caniniform). Die Eckzähne waren deutlich entwickelt und konisch ebenfalls spitz gestaltet. Der untere wies auf der Innenseite eine markante Rinne auf, die aber nicht so stark entwickelt war wie bei verwandten Gattungen. Sowohl zu den Schneidezähnen als auch zum hinteren Gebiss bestand jeweils ein kleineres Diastema, so dass der Eckzahn frei stand. Das hintere Gebiss bildete eine geschlossene Zahnreihe. Die Prämolaren zeigten bis auf den hinteren deutliche Abweichungen von den Molaren und waren demnach nicht oder nur wenig molarisiert. Sie wiesen auf der Kauoberfläche aber jeweils mehrere spitze Zahnschmelzhöcker auf. An den unteren Molaren waren es fünf, wobei die drei höheren eine Gruppe bildeten, die durch eine Rinne von den beiden niedrigeren getrennt wurden (tribosphenischer Zahnaufbau). Die oberen Molaren besaßen ebenfalls mehrere spitze Höcker, von denen sich zwei an der Außenseite und ein dritter etwas weiter innwärts gruppierten, die Zahninnenseite war von einem breiten Zahnschmelzgürtel (Cingulum) umrandet. Die Länge der Molaren variierte um die 4,8 mm, die Breite war mit rund 6 mm etwas größer. Der hinterste Backenzahn zeigte teilweise einige Streckungen.[1][3] Die gesamte Zahnreihe vom Eckzahn bis zum letzten Molaren betrug bei großen Arten bis zu 4 cm.[5]

Das Körperskelett ist durch mehrere Fossilfunde gut bekannt. Die Wirbelsäule bestand aus 7 Hals-, 13 Brust-, 5 bis 6 Lenden- und 22 bis 28 Schwanzwirbeln, die Anzahl der Kreuzbeinwirbel ist unbekannt, lag bei nahe verwandten Gattungen wie Pantolestes aber bei 3. Vor allem an den 6 vorderen Schwanzwirbeln befanden sich auffällig breite Querfortsätze. Zudem waren sie mit 1,5 cm Länge jeweils groß. Die dahinter liegenden Wirbel wurden zunehmend kleiner und wiesen nur noch rudimentäre Fortsätze auf. Die vorderen und hinteren Extremitäten besaßen ein ausgewogenes Längenverhältnis und waren insgesamt relativ kurz, jedoch hatten die hinteren eine massigere Ausprägung.[6] Der Oberarmknochen maß bis zu 5,4 cm und war im Schaftverlauf etwas gekrümmt. Die Knochen des Unterarms waren nicht verbunden, die Elle erreichte über 6 cm Länge und zeigte ein großes oberes Gelenk (Olecranon). Der robuste Oberschenkelknochen mit dem kräftigen Gelenkkopf besaß einen gut ausgebildeten Dritten Trochanter und wurde 7 cm lang, das Schienbein etwas länger. Dieses stellte somit den längsten Röhrenknochen im Skelett dar. Schien- und Wadenbein waren fest verwachsen. Vorder- und Hinterbeine endeten in jeweils fünfstrahlige Füße. Dabei besaßen die jeweils mittleren drei Strahlen (II bis IV) die längste Ausprägung und hier wiederum der dritte. Insgesamt wiesen die Vorder- und Hinterfüße keine starken Spezialisierungen auf, wie etwa deutliche Verlängerungen oder Kürzungen der Metapodien oder Phalangen. Allerdings waren die Endglieder der Zehen spitz zusammenlaufend und geben so die Existenz langer Krallen an, zudem bestanden auf der Oberfläche leichte Einkerbungen.[1][2][7][3]
Fossilfunde
Funde von Buxolestes stammen überwiegend aus dem Mittleren Eozän vor etwa 48 bis 43 Millionen Jahren und sind weitgehend nur aus Europa bekannt. Hervorzuheben sind vier nahezu vollständige Skelette aus der Grube Messel bei Darmstadt, die in seitlicher Lage überliefert sind.[2][7][3][4] Ein viertes Skelett aus der gleichen Lokalität wurde bei der Bergung aber beschädigt und ist damit weniger gut erhalten.[1] Alle Funde aus Messel sind dem unteren Abschnitt des Mitteleozäns zuzuweisen.[1] Von einem zerdrückten Schädel wurde weiterhin aus dem Geiseltal bei Halle berichtet. Er entstammt der dortigen Oberen Mittelkohle, die den oberen Abschnitt des Mitteleozäns repräsentiert.[8] Weitere Funde kommen aus Bouxwiller im Elsass in Frankreich. Diese umfassen neben einigen isolierten Zähnen und längeren Zahnreihen auch mehrere vollständige Unterkieferäste. Ihr Alter weist etwa eine Mittelstellung zwischen den Fossilien des Geiseltales und denen der Grube Messel auf.[5]
Paläobiologie
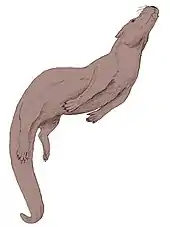
Im allgemeinen Körperbau, vor allem der im Vergleich zum Rumpf kurzen Gliedmaßen, dem langen Schwanz und der auf Fleischnahrung hinweisenden Gebissstruktur, entspricht Buxolestes einem heutigen Fischotter (Lutra lutra) aus der Familie der Marder. Eine dem Fischotter vergleichbare, semiaquatische Lebensweise lässt sich anhand der Skelettmorphologie erkennen. Das nach hinten kragende Hinterhauptsbein, die kräftigen Ansätze der Halswirbelsäule und der kleine Scheitelkamm verweisen auf eine gut entwickelte, kurze Nackenmuskulatur, was für einen sehr beweglichen Kopf spricht. Diese hohe Beweglichkeit war notwendig, um den Kopf beim Schwimmen über Wasser zu halten. Andererseits zeigen die kräftigen Fortsätze am Atlas, dem ersten Halswirbel, dass der Musculus obliquus besonders stark ausgebildet war und für eine erhöhte laterale Beweglichkeit des Kopfes sorgte. Dies ist vor allem bei der Jagd von Vorteil, wenn die Beute seitlich ausschert. An Ober- sowie Unterkiefer sind deutliche Foramina, Öffnungen im Knochen, ausgebildet, etwa das Foramen mentale oder das Foramen infraorbitale. Dadurch ist eine sehr sensible Schnauzenregion anzunehmen und das Vorhandensein von Tasthaaren um das Maul wahrscheinlich. Allerdings erreicht das Foramen infraorbitale nicht die vergleichbare Größe wie beim Fischotter, so dass die Vibrissen bei Buxolestes nicht so markant ausgeprägt waren. Möglicherweise waren die Vertreter von Buxolestes dadurch tagaktiv oder jagten in überwiegend klarem Wasser.[1][7]
Auch am Schwanz zeigen sich bestimmte Anpassungen an eine teils schwimmende Lebensweise. Die sechs großen Wirbel der Schwanzbasis mit ihren breiten Querfortsätzen, die ein Drittel der gesamten Schwanzlänge einnehmen, finden sich entsprechend beim Fischotter wieder, der den Schwanz als Antriebsorgan nutzt. Die dort ansetzenden Muskeln ermöglichen ein kraftvolles Auf- und Abgleiten sowie seitliches Schlängeln des Schwanzes. Aus der Grube Messel sind auch Reste des Weichteilgewebes durch Bakterien nachgezeichnet (Bacteriographie) und somit überliefert. Sie geben einen gerundeten, nicht abgeplatteten Schwanz wieder, wie ihn auch der Fischotter heute zeigt. Die Gliedmaßen zeigen dagegen nur wenige speziellen Anpassungen an das Schwimmen. Der Oberschenkelknochen ist nicht verkürzt, was als besondere Anpassung an das Schwimmen zu werten wäre, da die dabei entstehende verkürzte Hebelfunktion am Fuß das Paddeln erleichtern würde. Eine Verlängerung der Zehenglieder wie beim Seeotter ist nicht feststellbar. Der Hinterfuß war dabei durch die Verwachsung des Wadenbeins mit dem Schienbein in seinen seitlichen Bewegungen eingeschränkt. Die Ausprägung des Fersenbeins, vor allem des Fersenhöckers, ermöglichte aber eine kraftvolle Rückbewegung des Fußes. Die nicht miteinander verbundenen Knochen des Unterarms indizieren einen frei beweglichen Vorderfuß. Das kräftig ausgebildete obere Gelenk der Elle lässt zudem eine deutliche Streckung des Vorderbeins zu. Dies ist typisch für Tiere mit einer grabenden Lebensweise, wodurch Buxolestes demzufolge auch befähigt war, unterirdisch auf Nahrungssuche zu gehen oder Unterschlupfe zu graben. Auf eine grabende Tätigkeit könnte weiterhin der lange Mittelfinger hindeuten, der unter anderem typisch ist für die fossorial lebenden Gürteltiere, ebenso wie die bereits erwähnte gute Rückbewegung des Fußes zum Wegschieben des Aushubs.[1][7]
Häufig wird Buxolestes als räuberisch lebendes Tier mit Spezialisierung auf Fischnahrung angesehen. Hinweise dafür liefern neben dem allgemeinen Gebissaufbau die Backenzähne mit ihren sehr spitzen Höckern. Allerdings fehlt die für Raubtiere und den ausgestorbenen „Creodonten“ (Hyaenodonta und Oxyaenodonta) typische Brechschere im hinteren Gebiss.[3] Ein erster, nahezu vollständiger, aber etwas beschädigter Skelettfund aus der Grube Messel enthielt auch Hinterlassenschaften aus dem Magen-Darm-Bereich. Diese umfassten unzählige, zersplitterte Langknochen und Wirbelreste kleiner Wirbeltiere, aber auch dünne Knochenplättchen, die als Fischschuppen angesehen werden können, meist aber nicht aussagekräftig genug waren. Ein nur 2,2 mm langes Kieferfragment mit Resten der Bezahnung erwies sich als Rest von Atractosteus, einer in Messel häufig vorkommenden Fischform.[1] Ein weiterer Skelettfund der gleichen Lokalität enthielt bisher nur unbestimmbares Knochenmaterial.[7] Aus dem Mageninhalt zweier weiterer Individuen von Messel konnten dagegen nur pflanzliche Reste, so Samen und Früchte ermittelt werden.[3] Diese Befunde aus den Verdauungstrakten scheinen sich demzufolge zu widersprechen. Da aber heutige Raubtiere, vor allem aus der Gruppe der Hunde und Marder zum Teil auch gehäuft Pflanzenkost verspeisen, können die überlieferten Magenreste von Buxolestes auf eine, eventuell saisonabhängig variierende Ernährungsweise hindeuten.[2][9] Insgesamt ergibt sich aufgrund dieser Befunde für Buxolestes eine ähnliche Lebensweise wie beim heutigen Fischotter, wodurch er im Mittleren Eozän dessen ökologische Nische einnahm. Dabei brauchen von Buxolestes, wie Wighart von Koenigswald 1980 betonte, „aber weder die gleiche Eleganz beim Schwimmen noch das Ausmaß der Sinnesleistung wie bei Lutra erreicht worden zu sein.“.[10]
Systematik
Buxolestes ist eine Gattung aus der Familie der Pantolestidae. Diese umfassen urtümliche insektenfresserartige Tiere, die vom Mittleren bis Oberen Paläozän bis zum Unteren Oligozän in Nordamerika und Eurasien nachgewiesen sind und vorübergehend im Übergang vom Paläozän zum Eozän auch im nördlichen Afrika auftraten. Sowohl die inneren als auch die weiteren Verwandtschaftsverhältnisse der Pantolestidae sind nur unzureichend geklärt. Sie gelten häufig als Mitglieder der Unterordnung der Pantolesta. Diese wiederum werden von einigen Autoren zur übergeordneten Gruppe der Cimolesta verwiesen, welche frühe insektenfresserartige Säugetiere repräsentieren, die unter Umständen eine engere Bindung an die Ferae besitzen.[11][12][13] Andere Autoren ordnen die Pantolesta dagegen den Proteutheria zu und sehen sie in der Nahverwandtschaft der Insektenfresser.[14][15] Im Allgemeinen können die Pantolesta als eher basal innerhalb der Höheren Säugetiere eingestuft werden. Einer Studie aus dem Jahr 2013 legt eine engere Beziehungen zu den Leptictida und den Palaeanodonta nahe.[16] Innerhalb der Pantolestidae lässt sich Buxolestes in die Unterfamilie der Pantolestinae eingliedern. Deren Vertreter weisen im Gegensatz zu jenen der Pentacodontinae eindeutige Anpassungen an eine semiaquatische Lebensweise auf.[15] In die nähere Verwandtschaft von Buxolestes können Palaeosinopa und Pantolestes aus dem Unter- und Mitteleozän Nordamerikas und Pagonomus aus dem Paläozän Europas gestellt werden. Letzteres ist auch aus Walbeck in Sachsen-Anhalt überliefert.[1][17] Der Ursprung der Pantolesta wird teilweise bei den Leptictida gesehen, die bereits aus der oberen Kreide bekannt sind.[18]
Heute sind drei Arten bekannt:[5][1][3]
- B. hammeli Jaeger, 1970
- B. minor Pfretzschner, 1999
- B. piscator von Koenigswald, 1980
Zwei der drei Arten (B. minor und B. piscator) wurden in der Grube Messel nachgewiesen. Eventuell handelt es sich bei B. minor um eine noch nicht ausgewachsene Form von B. piscator, bei der letzte Mahlzahn zwar durchgebrochen, aber noch nicht eingerückt war. Auch fehlen ersterer das deutlich auskragende Hinterhauptsbein und die Querfortsätze an den Schwanzwirbeln.[4]
Die Erstbeschreibung von Buxolestes erfolgte durch Jean-Jaques Jaeger im Jahr 1970. Grundlage dafür bildeten mehrere Zahnfunde aus Bouxwiller. Der Holotyp (Exemplarnummer: Bchs 100) umfasst einen linken Unterkieferast mit der Bezahnung vom Eckzahn bis zum letzten Backenzahn, wobei der vorderste Prämolar unvollständig ist. Er befindet sich im Naturhistorischen Museum von Basel. Der Name Buxolestes bezieht sich einerseits auf den Fundort Bouxwiller, andererseits auf die Ähnlichkeit und nahe Verwandtschaft zum nordamerikanischen Pantolestes.[5]
Literatur
- Wighart von Koenigswald, Gregg F. Gunnell, Thomas Lehmann, Kenneth D. Rose und Irina Ruf: Vier ursprüngliche, aber hoch spezialisierte Säugetiere. In: Stephan F. K. Schaal, Krister T. Smith und Jörg Habersetzer (Hrsg.): Messel – ein fossiles Tropenökosystem. Senckenberg-Buch 79, Stuttgart, 2018, S. 223–233
Einzelnachweise
- Wighart von Koenigswald: Das Skelett eines Pantolestiden (Proteutheria, Mammalia) aus dem mittleren Eozän von Messel bei Darmstadt. Paläontologische Zeitschrift 54 (3/4), 1980, S. 267–287
- Wighart von Koenigswald: Das zweite Skelett eines Pantolestiden (Pantolestidae, Proteutheria, Mammalia) aus dem Mitteleozän von Messel bei Darmstadt. Carolines 45, 1987, S. 36–42
- Hans-Ulrich Pfretzschner: Buxolestes minor n. sp. - ein neuer Pantolestide (Mammalia, Proteutheria) aus der eozänen Messelformation. Courier Forschungsinstitut Senckenberg 216, 1999, S. 19–29
- Wighart von Koenigswald, Gregg F. Gunnell, Thomas Lehmann, Kenneth D. Rose und Irina Ruf: Vier ursprüngliche, aber hoch spezialisierte Säugetiere. In: Stephan F. K. Schaal, Krister T. Smith und Jörg Habersetzer (Hrsg.): Messel – ein fossiles Tropenökosystem. Senckenberg-Buch 79, Stuttgart, 2018, S. 223–233
- Jean-Jaques Jaeger: Pantolestidae nouveaux (Mammalia, Insectivora) de L'Éocène Moyen de Bouxwiller (Alsace). Palaeovertebrata 3, 1970, S. 63–82
- Kenneth D. Rose: The importance of Messel for interpreting Eocene Holarctic mammalian faunas. Palaeobiology and Palaeoenvironments 92, 2012, S. 631–647
- Hans-Ulrich Pfretzschner: Muscle reconstruction and aquatic locomotion in the Middle Eocene Buxolestes piscator from Messel near Darmstadt. Kaupia 3, 1993, S. 75–87
- Wighart von Koenigswald: Der erste Pantolestide (Proteutheria, Mammalia) aus dem Eozän des Geiseltales bei Halle. Zeitschrift für Geologische Wissenschaften 11, 1983, S. 781–787
- Gotthard Richter: Untersuchungen zur Ernährung eozäner Säuger aus der Fossilfundstätte Messel bei Darmstadt. Courier Forschungsinstitut Senckenberg 91, 1987, S. 1–33
- Wighart von Koenigswald: Das Skelett eines Pantolestiden (Proteutheria, Mammalia) aus dem mittleren Eozän von Messel bei Darmstadt. Paläontologische Zeitschrift 54 (3/4), 1980, S. 267–287 (S. 286)
- Malcolm C. McKenna: Toward a phylogenetic classification of the Mammalia. In W. Patrick Luckett und Frederick S. Szalay (Hrsg.): Phylogeny of the primates: a multidisciplinary approach. New York, London, 1975, S. 21–46
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 217–219)
- Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, S. 1–431 (S. 94–118)
- Leigh van Valen: New Paleocene insectivores and insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284 (S. 222–230)
- Gregg F. Gunnell, Thomas M. Bown, Jonathan I. Bloch und Douglas M. Boyer: Proteutheria. In: Christine M. Janis, Gregg F. Gunnell und Mark D. Uhlen (Hrsg.): Evolution of Tertiary Mammals of North America: Vol. 2, Small Mammals, Xenarthrans, and Marine Mammals. Cambridge University Press, 2008, S. 63–81
- Jerry J. Hooker: Origin and evolution of the Pseudorhyncocyonidae, a European Paleogene famaly of insectivorous placental mammals. Palaeontology 56 (4), 2013, S. 807–835
- Kenneth D. Rose und Wighart von Koenigswald: An exceptionally complete skeleton of Palaeosinopa (Mammalia, Cimolesta, Pantolestidae) from the Green River Formation, and other postcranial elements of the Pantolestidae from the Eocene of Wyoming (USA). Palaeontographica A 273, 2005 (3-6), S. 55–96
- Doug M. Boyer und Justin A. Georgi: Cranial Morphology of a Pantolestid Eutherian Mammal from the Eocene Bridger Formation, Wyoming, USA: Implications for Relationships and Habitat. Journal of Mammalian Evolution 14, 2007, S. 239–280