Angiotensin-konvertierendes Enzym 2
Angiotensin-konvertierendes Enzym 2 (englisch Angiotensin-converting enzyme 2, kurz ACE2) ist eine Metallocarboxypeptidase und ein Typ-1-Transmembranprotein mit Homologie zum Angiotensin-konvertierenden Enzym (ACE), das hauptsächlich in Eukaryoten, aber auch in Bakterien vorkommt. ACE2 spielt eine wichtige Rolle im Renin-Angiotensin-Aldosteron-System (RAAS), das den Volumenhaushalt des menschlichen Körpers steuert und den Blutdruck reguliert.
| Angiotensin-konvertierendes Enzym 2 | ||
|---|---|---|
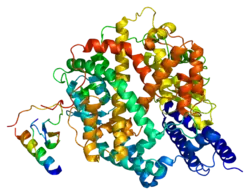
| ||
| Bändermodell des menschlichen Angiotensin-konvertierenden Enzyms 2, nach PDB 1R42 | ||
| Andere Namen |
| |
|
Vorhandene Strukturdaten: 3SCL, 1R42, 1R4L, 2AJF, 3D0G, 3D0H, 3D0I, 3KBH, 3SCL, 3SCJ, 3SCK | ||
| Eigenschaften des menschlichen Proteins | ||
| Masse/Länge Primärstruktur | 805 Aminosäuren | |
| Sekundär- bis Quartärstruktur | Heterodimer | |
| Kofaktor | Zn2+, Cl− | |
| Isoformen | 2 | |
| Bezeichner | ||
| Gen-Namen | ACE2 ; ACEH | |
| Externe IDs | ||
| Enzymklassifikation | ||
| EC, Kategorie | 3.4.17.23, Hydrolase | |
| MEROPS | M02.006 | |
| Reaktionsart | Hydrolyse | |
| Substrat | Angiotensin II, Angiotensin I | |
| Produkte | Angiotensin-(1-7), Angiotensin-(1-9) | |
| Vorkommen | ||
| Homologie-Familie | CLU_014364_3_0 | |
| Übergeordnetes Taxon | Eukaryoten, Bakterien | |
| Orthologe | ||
| Mensch | Hausmaus | |
| Entrez | 59272 | 70008 |
| Ensembl | ENSG00000130234 | ENSMUSG00000015405 |
| UniProt | Q9BYF1 | Q8R0I0 |
| Refseq (mRNA) | NM_021804 | NM_001130513 |
| Refseq (Protein) | NP_068576 | NP_001123985 |
| Genlocus | Chr X: 15.56 – 15.6 Mb | Chr X: 164.14 – 164.19 Mb |
| PubMed-Suche | 59272 | 70008 |
PCR-Analysen ergeben, dass ACE2 im Herzen sowie in der Lunge, Niere, im Endothel und im Magen-Darm-Trakt exprimiert ist.[1][2]
Außerdem ist ACE2 ein Rezeptor für verschiedene Coronaviren, einschließlich SARS-CoV und SARS-CoV-2, um in Zellen zu gelangen.[3][4][5] Dieses Bindungsverhalten stellt einen vielversprechenden Mechanismus für eine mögliche Behandlung dar.[6]
Struktur
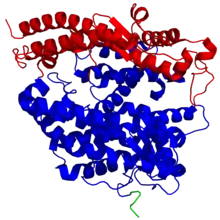
ACE2 ist ein Transmembranprotein Typ I mit einer extrazellulären N-terminalen Domäne, die das aktive Zentrum enthält, und einem kurzen intrazellulären C-terminalen „Schwanz“.[8] Die extrazelluläre Region des menschlichen ACE2-Enzyms besteht aus zwei Proteindomänen, zum einen die Zink-Metallopeptidase-Domäne als katalytische Domäne (Reste 19–611) und zum anderen die Domäne am C-Terminus (Reste 612–740), die zu ungefähr 48 % identisch mit dem menschlichen Protein Collectrin, ein Transmembran- und Glycoprotein sowie ein Homolog von ACE2[9], ist.
Das Zinkion wird im aktiven Zentrum durch die Aminosäurereste His374, His378, Glu402 und ein Wassermolekül (im nativen Zustand) koordiniert. Diese Aminosäurereste bilden zusammen eine Zink-bindende „HEXXH + E“-Konsensussequenz (H = Histidin, E = Glutaminsäure, X = unbekannte Aminosäure; siehe Einbuchstabencode), die bei Metalloproteasen im Clan MA konserviert ist. Ein Chloridion wird im nativen Zustand durch die Reste Arg169, Trp477 und Lys481 koordiniert.[7]
ACE2 unterzieht sich einer proteolytischen Spaltung, die zur Freisetzung der extrazellulären Domäne von der Zellmembran und zu einer enzymatisch aktiven Ektodomäne führt. Dieser Prozess wird als Ectodomain-Shedding bezeichnet. Ectodomain-Shedding kann auf zwei verschiedenen Wegen ablaufen: einem unterschwelligen Grund-Shedding und einem regulierten Shedding, das durch Phorbolester stimuliert wird. Das regulierte Shedding von ACE2 wird von der Metalloprotease bzw. Sheddase ADAM17 umgesetzt.[8]
Funktion
Physiologische Funktionen
ACE2 ist eine Protease, die Angiotensin-Peptide umsetzt. Damit hat es im menschlichen Körper viele physiologische Funktionen.[10] Insbesondere verschiebt es das Gleichgewicht von der Seite der vasokonstriktiven Peptide auf die der vasodilatorischen, welche hauptsächlich durch die Reaktion mit Angiotensin II umgesetzt wird. ACE2 dient somit als konterregulatorische Komponente des Renin-Angiotensin-Aldosteron-Systems.[11] ACE2 ist außerdem in der Lage, andere Peptide im kardiovaskulären System mit vasoaktivem Potential wie Apelin-13, Neurotensin, Kinestensin, Dynorphin und die Bradykinin(Bk)-Fragmente [des-Arg9]-Bk und [Lys-des-Arg9]-Bk zu hydrolysieren. Diese Reaktionen haben mögliche wichtige Implikationen in der Kontrolle des Blutdrucks, Herzfunktion und Reaktivität der Blutgefäße.[12] Eine Modulation der ACE2-Konzentration (und somit auch eine Änderung des Gleichgewichts zwischen zirkulierendem Angiotensin II und Angiotensin-(1-7)) hat mögliche Relevanz für Diabetes, Akutes Lungenversagen, Fibrose und sogar Muskeldystrophien.[11]
Als eine Carboxypeptidase[13] spaltet ACE2 die Aminosäure Phenylalanin am C-terminalen Ende von Angiotensin II ab, was zur Bildung von Angiotensin-(1-7) führt. Ein anderer Weg zur Bildung von Angiotensin-(1-7) ohne Bildung von Angiotensin II, aber kinetisch weniger effizient, ist ein alternierender Rollenwechsel zwischen ACE und ACE2. Dabei wird zunächst Angiotensin I durch ACE2 hydrolysiert und Angiotensin-(1-9) gebildet, ein Peptid mit unbekannter biologischer Aktivität, und anschließend durch ACE zu Angiotensin-(1-7) umgesetzt. Zusätzlich werden bei einer direkten Umwandlung von Angiotensin I zu Angiotensin-(1-7) weitere Enzyme benötigt, z. B. Prolylendopeptidase in vaskulären Endothelzellen, Neprilysin im Blutkreislauf oder in der Niere und Thimet-Oligopeptidase in der vaskulären glatten Muskelzelle. Angiotensin-(1-7) kann entweder vasodilatorische oder -konstriktive Effekte über verschiedene Mechanismen auslösen, abhängig von der Konzentration an Angiotensin-(1-7) und dem Gefäßsystem. Die Rolle von Angiotensin-(1–7) und seines bildenden Enzyms ACE2 könnte bei der Verringerung der Wirkungen von Angiotensin II größer sein, als dass sie selbst positive Wirkungen haben.[14]
Vorgeschlagener Mechanismus
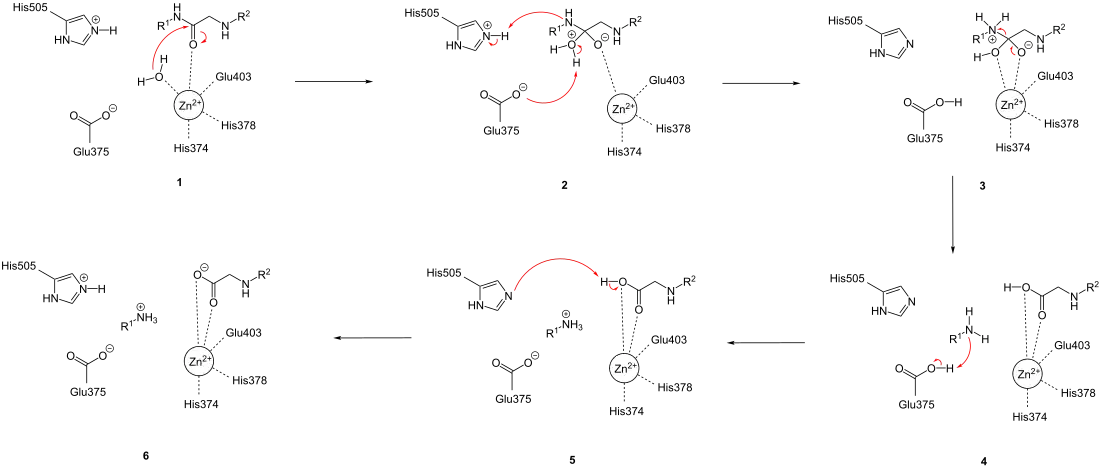
Zur vereinfachten Darstellung des Reaktionsmechanismus wird ein allgemeines Peptid als Substrat eingesetzt. Bei Substratbindung wird eine Scharnierbewegung induziert, um das aktive Zentrum zu schließen und die erforderlichen Aminosäurereste für die Katalyse in Position zu bringen. Bei der Reaktion wird zuerst der Enzym-Substrat-Komplex (in 1) zum tetraedrischen Zwischenprodukt (in 2) umgewandelt. Dafür führt das Zink-gebundene Wassermolekül einen nukleophilen Angriff auf die Carbonylgruppe des Peptids aus (1), was zu einer Protonenübertragung vom Wassermolekül zum Aminosäurerest Glu375 führt. Gleichzeitig wird ein Proton von His505 zum Stickstoffatom der abzuspaltenden Aminosäure übertragen (2). Das sp3-hybridisierte Stickstoffatom der abgespaltenen Aminosäure wird über Wasserstoffbrückenbindungen zu Pro346, His505 und/oder His345 stabilisiert. Danach folgt der Zerfall des tetraedrischen Zwischenproduktes und die Spaltung der Peptidbindung (3), was zur Protonenübertragung von Glu375 zur freien Aminogruppe der abgespaltenen Aminosäure führt (4). Anschließend wird ein Proton von der Carboxygruppe des neu entstandenen Peptids zu His505 direkt (5) oder indirekt durch Protonenaustausch über das Lösungsmittel zurückübertragen.[7]
Rezeptor für Coronaviren
SARS-CoV
Zunächst wurde vermutet, dass der Eintritt von SARS-CoV in die Zellen über eine direkte Fusion an der Zellmembran erfolgte. Später haben einige Studien gezeigt, dass der Eintritt von SARS-CoV pH-abhängig und dass dabei die endosomale Protease Cathepsin L involviert sein könnte, was darauf hinweist, dass dieser Virus über Endocytose in die Zellen gelangt. Schließlich konnte gezeigt werden, dass SARS-CoV über eine rezeptorabhängige und pH-sensible Endocytose in die Zellen gelangt. Die Behandlung von Zellen entweder mit dem SARS-CoV-Spike-Protein oder SARS-CoV-Spike enthaltenen Pseudoviren resultieren in einer Translokation von ACE2, dem funktionalen Rezeptor von SARS-CoV, von der Zelloberfläche zu Endosomen. SARS-CoV kann auch über eine rezeptorvermittelte Clathrin- und Caveolae-unabhängige Endocytose in die Zellen gelangen, die möglicherweise über Lipid Rafts erfolgt.[15]
SARS-CoV-2
Bei einer SARS-CoV-2-Infektion hat das Virion Kontakt mit menschlichen Zellen und wird in den Zellinnenraum aufgenommen. Bei den meisten Zellen wird dieser Vorgang durch die Bindung des Spike-Proteins der Virushülle an ein ACE2-Protein der Zellmembran ausgelöst. Für das Eindringen ist zumeist die Mitwirkung der zellulären Serinprotease TMPRSS2 notwendig.[16] Dringt SARS-CoV-2 in Alveolarepithelzellen (Pneumozyten) ein, kann dies zu respiratorischen Symptomen führen.
Diese Symptome sind bei Patienten mit vorbestehenden Herz-Kreislauf-Erkrankungen schwerwiegender, vermutlich aufgrund einer im Vergleich zu gesunden Personen erhöhten ACE2-Dichte auf der Zellmembran. Durch die Erhöhung des ACE2-Niveaus wird das Gleichgewicht in Richtung der konterregulatorischen Achse verschoben.[17] Die SARS-CoV-2-Infektion über ACE2 führt zur entzündungsfördernden Zytokinfreisetzung über die Angiotensin-II-AT1R-Achse; dies stellt ein mögliches therapeutisches Target über die IL-6-STAT3-Achse dar.[18]
Die Behandlung mit Inhibitoren des Renin-Angiotensin-Aldosteron-Systems (RAAS-Inhibitoren) hat Einfluss auf das Ausmaß der Infektion.[19] Verschiedene RAAS-Inhibitoren haben jeweils unterschiedliche Einflüsse auf das ACE2-Niveau. Bei Lewis-Ratten (in den 1950er Jahren entwickelte Laborratten) ist bei Verabreichung von entweder ACE-Hemmern oder Angiotensin-Rezeptorblockern das Ace2-mRNA-Niveau im Vergleich zu Ratten, die Placebos erhielten, erhöht. Insbesondere im Herzen der Ratte ist das Ace2-mRNA-Niveau bei Behandlung mit Lisinopril um das 4,7-Fache und bei Behandlung mit Losartan um das 2,8-Fache erhöht. Im Vergleich zum Placebo ist die ACE2-Aktivität bei einer Lisinopril-Behandlung, jedoch nicht bei einer Losartan-Behandlung, erhöht.[20] Bei einer Behandlung mit Captopril kann die ACE2-Expression in Ratten mit akutem Lungenversagen deutlich erhöht werden.[21] Bei Rattenmodellen zum akuten Lungenversagen ist die ACE-Aktivität und Angiotensin-II-Expression erhöht, wohingegen die ACE2-Aktivität und Angiotensin-(1-7)-Expression reduziert ist.[22][17]
Während gezeigt wurde, dass Angiotensin-Rezeptorblocker und Mineralocorticoid-Rezeptorblocker die ACE2-Expression und -Aktivität in verschiedenen experimentellen und klinischen Modellen erhöhen,[23][24] wird bei Verabreichung von ACE-Inhibitoren das Ace2-mRNA-Niveau des Herzens erhöht, hatte jedoch in experimentellen Modellen keinen Einfluss auf die ACE2-Aktivität.[25] Darüber hinaus war in einem Tiermodell zur diabetischen Nephropathie die Verabreichung von Aliskiren (einem direkten Renininhibitor) mit einer Reduzierung der ACE2-Expression verbunden.[26] Zur Behandlung von COVID-19 wurde im YouAn-Krankenhaus in Peking die intravenöse Transplantation von ACE2-negativen mesenchymalen Stammzellen (MSC) eingesetzt, insbesondere für Patienten im kritischen Zustand.[27]
Klinische Bedenken gibt es hinsichtlich der ACE2-Regulierung mit RAAS-Inhibitoren und Statinen zur Behandlung von COVID-19.[28][29][30]
Das deutsche Bundesinstitut für Arzneimittel und Medizinprodukte hat eine klinische Prüfung eines rekombinanten ACE2 an schwer erkrankten COVID-19-Patienten genehmigt.[31]
Einzelnachweise
- C. Tikellis, M. C. Thomas: Angiotensin-Converting Enzyme 2 (ACE2) Is a Key Modulator of the Renin Angiotensin System in Health and Disease. In: International journal of peptides. Band 2012, 2012, S. 256294, doi:10.1155/2012/256294, PMID 22536270, PMC 3321295 (freier Volltext).
- ACE-2: The SARS Receptor Identified. In: R&D Systems. Abgerufen am 16. Februar 2020.
- W. Li et al.: Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. In: Nature. Band 426, Nummer 6965, November 2003, S. 450–454, doi:10.1038/nature02145, PMID 14647384.
- Yushun Wan, Jian Shang, Rachel Graham, Ralph S. Baric, Fang Li: Receptor recognition by novel coronavirus from Wuhan: An analysis based on decade-long structural studies of SARS. In: Journal of Virology., doi:10.1128/JVI.00127-20.
- H. Hofmann et al.: Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. In: Proceedings of the National Academy of Sciences. Band 102, Nummer 22, Mai 2005, S. 7988–7993, doi:10.1073/pnas.0409465102, PMID 15897467, PMC 1142358 (freier Volltext).
- Tarek Mohamed et al.: Human recombinant soluble ACE2 (hrsACE2) shows promise for treating severe COVID19. In: Signal Transduction and Targeted Therapy. Band 5, Nr. 1, 3. November 2020, ISSN 2059-3635, S. 1–2, doi:10.1038/s41392-020-00374-6 (nature.com [abgerufen am 7. November 2020]).
- P. Towler, B. Staker, S. G. Prasad, S. Menon, J. Tang, T. Parsons, D. Ryan, M. Fisher, D. Williams, N. A. Dales, M. A. Patane, M. W. Pantoliano: ACE2 X-ray structures reveal a large hinge-bending motion important for inhibitor binding and catalysis. In: Journal of Biological Chemistry. Band 279, Nummer 17, April 2004, S. 17996–18007, doi:10.1074/jbc.M311191200, PMID 14754895.
- Daniel W. Lambert, Mike Yarski, Fiona J. Warner: Tumor Necrosis Factor-α Convertase (ADAM17) Mediates Regulated Ectodomain Shedding of the Severe-acute Respiratory Syndrome-Coronavirus (SARS-CoV) Receptor, Angiotensin-converting Enzyme-2 (ACE2). In: Journal of Biological Chemistry. Band 280, Nr. 34, 2005, S. 30113–30119, doi:10.1074/jbc.M505111200, PMID 15983030, PMC 8062222 (freier Volltext) – (englisch).
- Pei-Lun Chu, Thu H. Le: Role of Collectrin, an ACE2 Homologue, in Blood Pressure Homeostasis. In: Current Hypertension Reports. Band 16, Nr. 11, 2014, S. 490, doi:10.1007/s11906-014-0490-4, PMID 25182162, PMC 4164340 (freier Volltext) – (englisch).
- M. Bader: ACE2 – das missbrauchte Multitalent. In: Nephrologe. 2020, S. 1–4, doi:10.1007/s11560-020-00448-0, PMID 32837572, PMC 7362321 (freier Volltext).
- Anthony J.Turner: The Protective Arm of the Renin Angiotensin System (RAS). Functional Aspects and Therapeutic Implications. Academic Press, 2015, ISBN 978-0-12-801364-9, Chapter 25 - ACE2 Cell Biology, Regulation, and Physiological Functions, S. 185–189, doi:10.1016/B978-0-12-801364-9.00025-0 (englisch).
- M. K. Raizada, A. J. Ferreira: ACE2: a new target for cardiovascular disease therapeutics. In: Journal of cardiovascular pharmacology. Band 50, Nr. 2, August 2007, S. 112–119, doi:10.1097/FJC.0b013e3180986219, PMID 17703127 (englisch, Review).
- Mary Donoghue, Frank Hsieh, Elizabeth Baronas et al.: A Novel Angiotensin-Converting Enzyme–Related Carboxypeptidase (ACE2) Converts Angiotensin I to Angiotensin 1-9. In: Circulation Research. Band 87, Nr. 5, 2000, S. E1–E9, doi:10.1161/01.RES.87.5.e1, PMID 10969042 (englisch).
- Shlomo Keidar, Marielle Kaplan, Aviva Gamliel-Lazarovich: ACE2 of the heart: From angiotensin I to angiotensin (1–7). In: Cardiovascular Research. Band 73, Nr. 3, 2007, S. 463–469, doi:10.1016/j.cardiores.2006.09.006, PMID 17049503 (englisch).
- Hongliang Wang, Peng Yang, Kangtai Liu, Feng Guo, Yanli Zhang, Gongyi Zhang, Chengyu Jiang: SARS coronavirus entry into host cells through a novel clathrin- and caveolae-independent endocytic pathway. In: Cell Research. Band 18, Nr. 2, 2008, S. 290–301, doi:10.1038/cr.2008.15, PMID 18227861, PMC 7091891 (freier Volltext) – (englisch).
- M. Hoffmann et al.: SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. In: Cell. [elektronische Veröffentlichung vor dem Druck] März 2020, doi:10.1016/j.cell.2020.02.052, PMID 32142651, PMC 7102627 (freier Volltext).
- Ying-Ying Zheng et al.: Reply to: ‘Interaction between RAAS inhibitors and ACE2 in the context of COVID-19’. In: Nature Reviews Cardiology. 30. März 2020, doi:10.1038/s41569-020-0369-9.
- T. Hirano, M. Murakami: COVID-19: A New Virus, but a Familiar Receptor and Cytokine Release Syndrome. In: Immunity. [elektronische Veröffentlichung vor dem Druck] April 2020, doi:10.1016/j.immuni.2020.04.003, PMID 32325025, PMC 7175868 (freier Volltext).
- Y. Y. Zheng, Y. T. Ma, J. Y. Zhang, X. Xie: COVID-19 and the cardiovascular system. In: Nature reviews. Cardiology. [elektronische Veröffentlichung vor dem Druck] 5. März 2020, doi:10.1038/s41569-020-0360-5, PMID 32139904.
- C. M. Ferrario et al.: Effect of angiotensin-converting enzyme inhibition and angiotensin II receptor blockers on cardiac angiotensin-converting enzyme 2. In: Circulation. Band 111, Nummer 20, Mai 2005, S. 2605–2610, doi:10.1161/CIRCULATIONAHA.104.510461, PMID 15897343.
- Y. Li et al.: Angiotensin-converting enzyme inhibition attenuates lipopolysaccharide-induced lung injury by regulating the balance between angiotensin-converting enzyme and angiotensin-converting enzyme 2 and inhibiting mitogen-activated protein kinase activation. In: Shock. Band 43, Nummer 4, April 2015, S. 395–404, doi:10.1097/SHK.0000000000000302, PMID 25768373.
- R. M. Wösten-van Asperen et al.: Acute respiratory distress syndrome leads to reduced ratio of ACE/ACE2 activities and is prevented by angiotensin-(1-7) or an angiotensin II receptor antagonist. In: The Journal of pathology. Band 225, Nummer 4, Dezember 2011, S. 618–627, doi:10.1002/path.2987, PMID 22009550.
- S. Keidar et al.: Mineralocorticoid receptor blocker increases angiotensin-converting enzyme 2 activity in congestive heart failure patients. In: Circulation research. Band 97, Nummer 9, Oktober 2005, S. 946–953, doi:10.1161/01.RES.0000187500.24964.7A, PMID 16179584.
- J. C. Zhonget et al.: Telmisartan attenuates aortic hypertrophy in hypertensive rats by the modulation of ACE2 and profilin-1 expression. In: Regulatory peptides. Band 166, Nummer 1–3, Januar 2011, S. 90–97, doi:10.1016/j.regpep.2010.09.005, PMID 20854846.
- S. R. Tipnis et al.: A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidase. In: Journal of Biological Chemistry. Band 275, Nummer 43, Oktober 2000, S. 33238–33243, doi:10.1074/jbc.M002615200, PMID 10924499.
- W. Ding et al.: [Aliskiren inhibits angiotensin II/angiotensin 1-7(Ang II/Ang1-7) signal pathway in rats with diabetic nephropathy]. In: Xi bao yu fen zi mian yi xue za zhi = Chinese journal of cellular and molecular immunology. Band 34, Nummer 10, Oktober 2018, S. 891–895, PMID 30554582 (chinesisch).
- Zikuan Leng et al.: Transplantation of ACE2- Mesenchymal Stem Cells Improves the Outcome of Patients with COVID-19 Pneumonia. In: Aging and disease. Band 11, 9. März 2020, S. 216, doi:10.14336/AD.2020.0228.
- A. M. South, D. Diz, M. C. Chappell: COVID-19, ACE2 and the Cardiovascular Consequences. In: American Journal of Physiology – Heart and Circulatory Physiology. [elektronische Veröffentlichung vor dem Druck] 31. März 2020, doi:10.1152/ajpheart.00217.2020, PMID 32228252.
- ESH UPDATE ON COVID-19. In: eshonline.org. European Society of Hypertension, 19. März 2020, abgerufen am 8. April 2020.
- B. Williams et al.: 2018 ESC/ESH Guidelines for the management of arterial hypertension: The Task Force for the management of arterial hypertension of the European Society of Cardiology and the European Society of Hypertension. In: Journal of hypertension. Band 36, Nummer 10, 10 2018, S. 1953–2041, doi:10.1097/HJH.0000000000001940, PMID 30234752.
- Coronavirus SARS-CoV-2. In: bfarm.de. Bundesinstitut für Arzneimittel und Medizinprodukte, 2020, abgerufen am 10. April 2020 (die Seite wird regelmäßig aktualisiert, siehe auch Versionen vom 25. März und 10. April im Web Archive).