Mylodon
Mylodon ist eine Gattung aus der ausgestorbenen Familie der Mylodontidae, die große, bodenlebende Faultiere umfasst. Sie gehört mit einer Gesamtlänge von 3 bis 4 m zu den bekanntesten und größten Vertretern der Gruppe. Nachgewiesen ist die Gattung vor allem im südlichen Teil Südamerikas. Die ältesten Funde datieren wahrscheinlich in das Unterpleistozän. Der größte Teil der Fossilreste stammt jedoch aus der Zeit des Oberpleistozäns. Eine der wichtigsten Fundstellen dieser Phase findet sich mit der Cueva del Milodón im südlichen Chile. Kurz darauf, vor etwa 10.000 Jahren, starb Mylodon aus. Zu diesem Zeitpunkt trat die Faultiergattung gemeinsam mit den ersten menschlichen Besiedlern Amerikas auf. Es existieren aber kaum Hinweise darauf, dass sie verstärkt vom Menschen bejagt wurde.
| Mylodon | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
.jpg.webp)
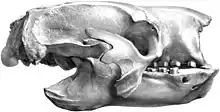
Schädel von Mylodon | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unteres bis Oberes Pleistozän | ||||||||||||
| 1,8? Mio. Jahre bis etwa 10.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Mylodon | ||||||||||||
| Owen, 1840 | ||||||||||||
Von Mylodon sind nicht nur Knochen- und Gebissreste, sondern auch verschiedenes Weichteilgewebe wie Haut und Fell sowie Nahrungsreste in Form von versteinerten Kotballen überliefert. Als herausragendes Kennzeichen kann der bei Mylodon stark verlängerte und gegenüber anderen großen Mylodonten deutlich schmalere Schädel herausgestellt werden, der vorn einen vollständig geschlossenen Nasenbogen aufwies. Weitere Unterscheidungsmerkmale betreffen die Struktur des vorderen Gebisses.
Die Tiere lebten ausschließlich bodenbewohnend. Ein dichtes Fell mit langen Haaren lässt sich als Anpassung an ein Leben unter kalten Klimabedingungen interpretieren, wie sie in der ausgehenden letzten Kaltzeit im südlichen Südamerika vorgeherrscht haben. Damit korrespondiert auch eine überwiegend auf Gräser basierte Ernährung in dieser Region. Die weite Verbreitung von Mylodon bis in die Pamparegion und einige Merkmale am Schädel zeigen jedoch auf, dass die Tiere eine weitaus größere ökologische Bandbreite besaßen und auch mit wärmeren Temperaturbedingungen und möglicherweise auch einer gemischten Pflanzenkost zurechtkamen. Die Tiere fielen teilweise größeren Beutegreifern zum Opfer.
Die Gattung wurde im Jahr 1840 wissenschaftlich eingeführt, in der Regel ist nur eine Art anerkannt. Das Typusmaterial stammt aus dem Gebiet der Pampa, wo es von Charles Darwin während seiner Reise mit der HMS Beagle aufgesammelt worden war. Im Laufe der Forschungsgeschichte kam es zu Verwechslungen und Gleichsetzungen mit anderen großen Bodenfaultieren wie Glossotherium und Paramylodon, was erst in den 1920er Jahren gelöst werden konnte. Zudem gehörte Mylodon zu den ersten ausgestorbenen Faultieren, an denen genetische Untersuchungen zur Klärung der stammesgeschichtlichen Beziehungen vorgenommen wurden.
Merkmale
Körpergröße
Mylodon war ein großer Vertreter der Mylodontidae. Seine Gesamtlänge betrug schätzungsweise rund 3 bis 4 m. Anhand der Schädelausmaße wird ein Gewicht zwischen 1 und 2 t vermutet mit einer annähernden Schätzung von 1,65 t.[1] Damit wies Mylodon etwa die Größe verwandter Formen wie Glossotherium oder Paramylodon auf, war aber deutlich kleiner als das riesenhafte Lestodon. Im Körperbau entsprachen die Tiere weitgehend den anderen großen bodenlebenden Faultieren.[2][3]
Schädel- und Gebissmerkmale
Vor allem im Schädelbau wich Mylodon deutlich von anderen verwandten Formen ab. Seine Länge variierte nach Untersuchung von zehn mehr oder weniger gut überlieferten Exemplaren zwischen 59,0 und 71,5 cm, was deutlich länger ist als bei Glossotherium oder Lestodon.[4] Am Hirnschädel war er zwischen 16,5 und 22,5 cm breit, im vorderen Nasenbereich zwischen 11,3 und 15,5 cm. Die Höhe betrug entsprechend am hinteren Schädel 14,0 bis 19,0 cm und am vorderen 15,0 bis 23,5 cm.[5] Der Schädel war dadurch langgestreckt und schmal, im Gegensatz zu Glossotherium und Lestodon, die einen kurzen und sehr breiten Schädel aufwiesen. Die außerordentliche Länge des Schädels von Mylodon wurde vor allem durch Streckungen im rostralen Abschnitt erreicht. In Aufsicht von oben verschmälerte sich das Rostrum nach vorn. Hier findet sich auch der bedeutendste Unterschied zu den meisten anderen Vertretern der Mylodontidae: Das Nasenbein war lang und schmal sowie im vorderen Bereich nach unten gewölbt. Am vorderen Ende verband es sich mit dem über einen Fortsatz verlängerten Mittelkieferknochen, der wiederum mit dem Oberkiefer fusionierte. Dadurch entstand ein bei ausgewachsenen Individuen vollständig geschlossener Nasenbogen, der bei anderen Faultieren weitgehend unbekannt ist. Im Vergleich dazu zeigten die Schädel von Glossotherium und Lestodon, aber auch von Paramylodon einen von oben gesehen verbreiterten Nasenbereich, der eher kurz war und in Seitensicht deutlich abgeschnitten wirkte, der Mittelkieferknochen hatte hier keinen Kontakt zum Nasenbein. Das Schädeldach verlief bei Mylodon weitgehend gerade, lediglich oberhalb der Orbita konnte eine leichte Eindellung auftreten. Am Scheitelbein setzten deutliche Temporallinien an, die aber keinen Scheitelkamm bildeten. Der Jochbogen war schlank, der vordere Ansatz begann oberhalb des dritten und vierten Backenzahns. Er bildete mit dem hinteren Bogenansatz keinen festen Schluss. Wie üblich bei Faultieren bestand der vordere Bogenansatz aus drei Fortsätzen: einem aufsteigenden, einem horizontalen und einem absteigenden, von denen der erstere am längsten war. Der hintere Bogenansatz formte eine dreieckige Platte. Das Hinterhauptsbein knickte in einem Winkel von 120 ° vom Schädeldach ab. Die Unterseiten der Hinterhauptsgelenke saßen etwa auf Höhe der Kauebene. In der Ansicht von hinten zeigte sich das Hinterhauptsbein nahezu kreisrund und nicht so gedrückt wie bei Glossotherium und Lestodon.[6] Auf der Schädelunterseite wies der Gaumen eine schmale und nach vorn mehr oder weniger dreieckig orientierte Form auf. Charakteristisch waren hier zahlreiche kleine Knochenöffnungen. Die Glenoidgrube, in der das Gelenk des Unterkiefers einrastet, entsprach mit ihrer schwachen Ausprägung der anderer Mylodonten, wodurch aber insgesamt eine freie Rotationsmöglichkeit bestand.[7][5][4]

Der Unterkiefer variierte in seiner Länge zwischen 42,0 und 48,0 cm, gemessen an drei Fossilfunden.[4] Er war langgestreckt, deutlicher als bei Glossotherium und Lestodon, da sich bei Mylodon vor allem der Bereich vor den Zähnen gedehnt hatte. Der horizontale Knochenkörper nahm nach hinten kontinuierlich an Höhe zu, unterhalb des letzten Backenzahns betrug sie etwa 10,5 bis 12,7 cm. Die Symphyse am vorderen Ende zur Gelenkung der beiden Unterkieferhälften wurde rund 12,4 cm lang. Hier stieg die Unterkante des Unterkieferkörpers schräg auf, so dass das vordere Ende der Symphyse oberhalb der Kauebene der Zähne lag. Die Symphyse zog wie bei anderen Faultieren nach vorn aus, sie endete leicht abgerundet. Entsprechend dem Rostrum des Schädels war die Symphyse bei Mylodon schmal und nicht so breit wie bei Glossotherium und Lestodon. Kurz hinter der Symphyse öffnete sich das Foramen mandibulae. Der aufsteigende Ast setzte hinter dem letzten Backenzahn an und bildete einen Winkel von 140 ° zur Kauebene. Der Kronenfortsatz erhob sich teilweise bis zu 20 cm. Dagegen saß der Gelenkfortsatz tiefer, etwa auf Höhe der Kauebene, wodurch sich eine niedrige Schädel-Unterkiefer-Verbindung ergab. Der Winkelfortsatz am hinteren Unterkieferende zeichnete sich deutlich ab. Teils kippte er nach unten ab und lag unterhalb der Unterkante des horizontelen Knochenkörpers. Die Oberseite des Winkelfortsatzes erreicht nicht die Kauebene.[7][5][4]
Das für die Faultiere typische Gebiss weicht stark von dem der anderen Höheren Säugetiere ab und besteht in der Regel oben aus fünf und unten aus vier Zähnen je Kieferbogen, insgesamt also aus 18 Zähnen. Bei den Mylodonten zeigte sich der jeweils erste Zahn häufig als eckzahnförmig (caniniform) in seiner Gestalt, während die hinteren Zähne eher molarenartig (molariform) wirkten. Innerhalb der Faultiere kann diese Gebissstruktur als ursprünglich bezeichnet werden. Mylodon wies dahingehend als Besonderheit auf, dass der obere eckzahnartige Zahn einer jeden Reihe vollständig zurückgebildet war und hier nur die molarenartigen vier hinteren Zähne vorkamen. In der unteren Zahnreihe war der vordere caniniforme Zahn in einen molariformen umgestaltet. Das Gebiss setzte sich somit aus insgesamt 16 Zähnen zusammen. Dies erinnert ein wenig an Paramylodon, bei dem die oberen eckzahnförmigen Zähne ebenfalls fehlten, die unteren aber ihre auffallend spitze Form beibehalten hatten. Abweichend davon wiesen Glossotherium und Lestodon das ursprüngliche Faultiergebiss auf. Als Charakteristikum der Mylodonten kann die flache, lappenartige und weitgehend eingedellte Struktur der molariformen Zähne hervorgehoben werden, die sich dadurch deutlich von der der Megatheriidae und Megalonychidae mit ihren zwei quergestellten erhabenen Leisten je Zahn unterscheidet. Die Form der Zähne war bei Mylodon insgesamt einfacher gestaltet. Sie hatten in der Oberkieferzahnreihe einen eher runden bis ovalen, in der Unterkieferzahnreihe einen eher rautenförmigen Umriss. Die typisch komplexere zweilappige Gestaltung der molarenartigen Zähne von Glossotherium und Lestodon, hervorgerufen durch eine zentrale Einschnürung, kam bei Mylodon nur am unteren hintersten Zahn vor. Allgemein standen die Zahnreihen nach vorn divergierend zueinander, zudem waren die Zähne sehr hochkronig (hypsodont). Die obere Zahnreihe erstreckte sich über eine Länge von 10,9 bis 13,3 cm, die untere war zwischen 12,0 und 15,0 cm lang.[8][7][5][4]
Körperskelett

Funde des postcranialen Skeletts sind bei Mylodon weitaus seltener als bei den anderen großen mylodonten Faultieren Glossotherium, Lestodon und Paramylodon. Dadurch ist das Körperskelett weniger gut dokumentiert. Von der Wirbelsäule sind nur einzelne Elemente wie der Atlas und verschiedenen Brustwirbel beschrieben worden. Der Oberarmknochen war massiv und mit 46,0 bis 48,0 cm ausgesprochen lang. Der Gelenkkopf, dessen Durchmesser bei über 10 cm lag, hob sich durch seine halbkugelige, jedoch seitlich etwas abgeflachte Form ab. Am Schaft zog sich eine deutliche deltopectorale Leiste herab, die als Ankerpunkt für die Schultermuskulatur fungierte. Wie bei vielen Bodenfaultieren lud das untere Gelenkende weit aus und brachte es hier auf eine Breite von fast 26 cm. Teilweise wurde dies durch eine massive innere Epicondyle hervorgerufen. Die Gelenkflächen (Capitulum und Trochlea) standen nahezu senkrecht zueinander und bildeten keinen so stumpfen Winkel wie bei Glossotherium. Die Elle war grazil gebaut. Ihre Länge betrug rund 37 cm. Das Olecranon, also der obere Gelenkfortsatz, nahm davon rund 8,1 cm ein, was etwa 22 % der Gesamtlänge entspricht und deutlich weniger ist als vergleichsweise bei Glossotherium und Lestodon. Es war seitlich verschmälert, was sich unter anderem auch bei Paramylodon wiederfindet. Die Speiche ähnelte weitgehend der von Glossotherium und war kompakt und gerade gebaut mit einer Länge von etwa 30 cm. Der Kopf hatte einen ovalen Umriss mit einer auffälligen Lippe. Das Becken war äußerst ausladend und zwischen den beiden Darmbeinen 114 cm breit. Der Oberschenkelknochen maß zwischen 55 und 59 cm in der Länge. Er war typisch für Bodenfaultiere brettartig flach. Am Schaft zog seine Breite deutlich ein, der niedrigste Wert wurde kurz unterhalb des Mittelpunktes erreicht. Hier betrug die Breite rund 18 cm, die Dicke etwa 7,5 cm. Die Gelenkenden waren hingegen markant breiter, am Knieende rund 30 cm und am Fußende etwa 26 cm. Der halbkugelige, gut 14 cm durchmessende Gelenkkopf überragte den Großen Rollhügel auffallend. Der dritte Rollhügel zeichnete sich an der Außenkante des Schaftes unterhalb des Großen Rollhügels nur als kleine Erhebung ab. Gegenüber dem Oberschenkelknochen erreichte das Schienbein mit etwa 27 cm nur etwa die Hälfte der Länge, ein für Mylodonten charakteristisches Merkmal. Auch dieser Knochen war deutlich flach mit einer Dicke, die nur die Hälfte des Wertes der Breite am Schaft einnahm. Das Wadenbein liegt bisher nur fragmentiert vor. Es war am Schaft eingezogen und an den Gelenkenden verbreitert, wobei das obere Gelenkende deutlichere Rundungen zeigte als bei Glossotherium.[9][10][11]
Die Hand umfasste insgesamt fünf Strahlen (I bis V), wobei am ersten Strahl der Mittelhandknochen mit dem Großen Vieleckbein verwachsen war. Dadurch entstand der für viele Bodenfaultiere typische sogenannte Metacrapal Carpal Complex (kurz MCC genannt). Als Besonderheit an der Handwurzel zeigte sich das Erbsenbein deutlich flach, in seiner Form glich es dem von Glossotherium, wich aber von dem entsprechenden Knochen anderer Mylodonten mit kugeliger, walnussartiger oder pyramidaler Gestalt ab. Den längsten Mittelhandknochen hatte der vierte Strahl ausgebildet, während der des fünften nur wenig kürzer wurde. Die jeweiligen Knochen maßen dort rund 12,5 und 10,7 cm in der Länge. Wahrscheinlich waren wie bei Glossotherium und bei Paramylodon nur die drei inneren Strahlen krallentragend, lediglich vom zweiten Strahl sind aber bisher alle Knochenelemente dokumentiert. Der Mittelhandknochen wies an diesem eine Länge von 7,8 cm auf und war sehr grazil gebaut. Die erste Phalanx war äußerst kurz und nur rund 2,5 cm lang, die zweite wurde rund 4,2 cm lang und die dritte wenigstens 11,5 cm. Sie war röhrenförmig gestaltet und ging nach vorn in einen Fortsatz über, auf dem die Kralle ruhte. Die ersten Fingerglieder der beiden äußeren Strahlen waren in ihrer Länge deutlich reduziert. Vom Fuß liegen nur einzelne Wurzelknochen wie das Sprungbein vor.[12][10]
Haut, Fell und Osteoderme

Mylodon gehört zu den wenigen ausgestorbenen Säugetierformen, von denen mumifizierte Hautreste vorliegen. Der bedeutendste Fundpunkt für derartige Funde ist die Cueva del Milodón in der chilenischen Provinz Última Esperanza, wo die ersten Fellteile bereits Ende des 19. Jahrhunderts zu Tage gefördert wurden.[13][14] Einzelne Stücke weisen Längen von bis zu 150 cm auf, sind aber durch Trocknungsprozesse geschrumpft. Ihre Dicke beträgt an manchen Stellen bis zu 1,5 cm, meist liegt sie aber bei rund 1 cm. Die Haut ist dicht mit steifen, wenig gewellten Haaren besetzt, wobei in der Regel nur das Deckhaar ausgebildet ist, während die Unterwolle fehlt. In diesem Merkmal bestehen Gemeinsamkeiten zu den Zweifinger-Faultieren (Choloepus), jedoch weniger zu den Dreifinger-Faultieren (Bradypus) die über eine Unterwolle verfügen. Die Länge der einzelnen Haare variiert zwischen 5 bis teilweise über 20 cm mit den kürzesten im Bereich des Hinterkopfes, mittellangen am Rücken und sehr langen Haaren an den Gliedmaßen. Ihre gegenwärtige Färbung reicht von gelblich bis rötlichbraun. Die Haarschäfte sind uniform röhrenförmig, am oberen Ende formen sie stumpfe Spitzen aus. Wie bei den heutigen Faultieren wiesen die Haare keine Markröhre (Medulla) auf. Im Unterschied zu den Haaren der Zweifinger-Faultiere fehlt ihnen deren charakteristische Längsriffelung.[13][14][15][16][17]


Als einzige Vertreter der Faultiere haben die Mylodonten knöcherne Plättchen in der Haut eingelagert. Solche Osteoderme genannten Bildungen sind im größeren Maße heute nur bei den Gürteltieren bekannt. Im Gegensatz zu dem äußeren Panzer der Gürteltiere waren bei den Mylodonten die Knochenplättchen eher locker gestreut. Die ersten Funde von einzelnen Osteodermen von Mylodon veröffentlichte Hermann Burmeister bereits in den 1860er Jahren.[18][19] Die in den Höhlen von Última Esperanza aufgefundenen Hautreste vermitteln einen Eindruck, wie diese in der Haut eingebettet und über den Körper verteilt waren. Die Knochenplättchen liegen alle im unteren Abschnitt der Haut, während in den oberen Abschnitten die Haare ihren Ursprung haben. Die Verteilung erwies sich als sehr uneinheitlich. Einige Bereiche mit einer dichten Anordnung an Osteodermen enthalten auf 10 cm² zwischen 83 und 95 Knochenplättchen. Bei anderen dünnt die Anzahl hingegen sehr stark aus. Jedoch selbst bei dichter Anordnung vereinigen sich die Osteoderme nie zu einem geschlossenen Panzer, sondern sind immer durch einzelne Hautfalten voneinander getrennt. In Übereinstimmung mit den Panzern der Gürteltiere bilden die Knochenplättchen eine einzelne Schicht und kommen nicht gestapelt vor. Da alle Hautreste isoliert von den Körperskeletten aufgefunden wurden, ist es teilweise schwierig, die Hautbereiche mit dichter und dünner Anordnung von Knochenplättchen einer bestimmtem Körperpartie zuzuordnen. Es ist aber anzunehmen, dass der Rücken weitgehend gepanzert und der Bauch frei war. In den Abschnitten mit dichter Osteodermausprägung waren diese größer gestaltet als in den lichten Bereichen. Die Knochenplättchen von Mylodon waren zumeist von unregelmäßiger ovaler Gestalt mit Ausmaßen von 0,5 bis 2,5 cm in der Länge, 0,3 bis 1,8 cm in der Breite sowie 0,2 bis 1,1 cm in der Dicke bei Gewichten von maximal 2 g. Oberflächig wiesen sie einzelne Grübchen auf.[20] Im Querschnitt bestanden sie aus zahlreichen Faserbündeln vermischt mit harten Knochenlamellen (Osteome). Ihr Aufbau war dadurch wesentlich einfacher als bei den Gürteltieren, auch fehlte ihnen wohl die von den Gürteltieren bekannte Keratinauflage. Prinzipiell ähnelten die Osteoderme von Mylodon denen anderer großer Mylodonten.[13][14][21][22][23]
Verbreitung und wichtige Fossilfunde
Überblick und Ursprünge
Mylodon war vor allem im südlichen Bereich von Südamerika verbreitet. Fossilfunde liegen aus Argentinien, Chile, Bolivien, Uruguay und Brasilien vor. Die besiedelten Regionen umfassen somit sehr weit südliche Fundpunkte auf der Insel Feuerland sowie den Großteil Patagoniens bis nach Norden zur Pamparegion hin. Seine südliche Grenze erreichte das Verbreitungsgebiet bei etwa 53 ° südlicher Breite. Die Fundstelle Tres Arroyos auf Feuerland sowie die Region um die Cueva del Milodón im südwestlichen Patagonien gehören zu den südlichsten bekannten Nachweisen eines Faultiervertreters im Pleistozän.[24][25] In der Pamparegion fand sich die nördliche Grenze etwa am Fluss Chuí im südostbrasilianischen Bundesstaat Rio Grande do Sul um den 30. südlichen Breitengrad. Noch weiter nördlich liegende Fundpunkte wie Ñuapua in Bolivien tangieren den 20. südlichen Breitengrad. Aus Paraguay berichtete Funde gelten allerdings als eher unsicher.[26][27][28]
Ihr erstes Auftreten hatte die Faultiergattung möglicherweise bereits im Unterpleistozän, jedoch sind Funde eher rar.[29][30] In diesem Zeitraum trat im Pampasgebiet zusätzlich noch die möglicherweise nahe verwandte Form Archaeomylodon auf, deren vorderste eckzahnartige Zähne der oberen Zahnreihe zwar stark verkleinert, aber noch nicht vollständig reduziert waren.[31] Zu den frühen und weiter nördlich gelegenen Funden von Mylodon gehört beispielsweise ein Schädel aus der El-Palmar-Formation in der argentinischen Provinz Entre Ríos, der noch in die ausgehende letzte Warmzeit vor rund 80.000 Jahren datiert.[5] Ebenfalls aus den nördlichen Verbreitungsarealen sind zwei Teilskelette erwähnenswert, von denen das eine am Río Anisacate in der argentinischen Provinz Córdoba und das andere in Arroyo Quequén Salado bei Oriente in der argentinischen Provinz Buenos Aires zu Tage gefördert wurden.[10][11] Hauptsächlich in der Pampa kam es im Oberpleistozän zu einer Überschneidung des Vorkommens von Mylodon mit den beiden anderen großen mylodonten Faultiervertretern Glossotherium und Lestodon. Ein tatsächliches gemeinsames Auftreten ist aber nur selten belegt. Hierzu gehören die bedeutende archäologische Fundstelle Paso Otero[32][33][34] in der Provinz Buenos Aires, die Lokalität Arroyo de Vizcaíno[35] im südlichen Uruguay und der Fluss Chuí.[36][37][38]
Bedeutende Funde des Oberpleistozäns
Wie bei einem Großteil der anderen großen Bodenfaultiere auch stammt das meiste Fundmaterial von Mylodon aus dem Oberpleistozän mit einem Schwerpunkt zum Ende der letzten Kaltzeit hin. Es ist gleichzeitig die Phase, in der Mylodon wieder aus dem Fossilbericht verschwand. Aus globaler Sicht starben im Übergang vom Pleistozän zum Holozän zahlreiche größere Tiere aus, weswegen dieses Ereignis als Quartäre Aussterbewelle angesehen wird. In Südamerika fällt diese mit dem ersten Auftreten des Menschen zusammen. Ob beides in einem ursächlichen Zusammenhang steht, wird vielfach und kontrovers diskutiert. Neben der potentiellen Jagd und möglichen Landschaftsüberprägungen seitens der frühen menschlichen Jäger-Sammler-Gruppen können hier auch klimatische Veränderungen einen Einfluss gehabt haben.[39][40] Zahlreiche archäologische Fundstellen, vor allem in der Pamparegion und im patagonischen Raum, sind zwischen 13.500 und 10.000 Jahre alt. Der überwiegende Teil davon bezeugt zumindest eine Koexistenz von Menschen und Bodenfaultieren über längere Zeit. Direkte Assoziationen von menschlichen Kulturerzeugnissen und Fossilresten von Mylodon finden sich unter anderem in der Gruta del Indio am Ostfuß der Anden, in Piedra Museo oder in Las Buitreras, alle Argentinien, beziehungsweise in Tres Arroyos auf Feuerland. Mylodon wird häufig durch einzelne Osteoderme, Knochen oder in Form von Koprolithen repräsentiert, während die menschlichen Hinterlassenschaften sich auf Steinartefakte und/oder Feuerstellen beschränken. Ob es dabei auch zu einer mehr oder weniger intensiven Rohmaterialnutzung von Faultierknochen seitens des Menschen kam, ist vielfach unbelegt. Zahlreiche Knochenmarken, die ursprünglich als anthropogen verursacht interpretiert wurden, sind neueren Untersuchungen zufolge auf Raubtierfraß zurückzuführen. Noch schwieriger ist der Nachweis einer direkten Jagd des Menschen auf die großen Bodenfaultiere. Als ein Beleg wird häufig Quebrada de Quereo betrachtet, einer Fundstelle an einer alten Küstenlinie im nördlichen Chile. Von hier stammen unter anderem Skelettreste zweier Individuen von Mylodon, verteilt auf einer jeweils eng begrenzten Fläche, aber in zwei unterschiedlichen stratigraphischen Einheiten und in einem räumlichen Abstand von 21 m zueinander. Eines der Individuen war mit rund 70 Steinobjekten vergesellschaftet, deren anthropogener Ursprung in Diskussion ist. Auf den Knochen finden sich keine Schnittmarken als Hinweise auf eine etwaige menschliche Manipulation. Das Alter der Fundstelle wird mit 11.600 bis 10.900 Jahren vor heute angegeben.[41][42][43]


Eine der bedeutendsten Fundstellen bildet die Cueva del Milodón nahe dem Lago Sofía in der chilenischen Provinz Última Esperanza, die vor allem wegen der überlieferten Hautreste bekannt ist. Sie gehört zu einem ganzen System an Höhlen in der Region, wie der Cueva del Medio[44][45] oder der Cueva Chica,[46] die sich perlenschnurartig an der Südflanke des 556 m hohen Cerro Benitez entlangreihen. Cueva del Milodón ist eine große Höhle von 250 m Länge, 140 m Breite und 30 m Höhe am Eingang beziehungsweise 10 m im hinteren Bereich. Entdeckt wurde sie 1895 vom deutschen Kapitän Hermann Eberhard, der auch die ersten Hautreste fand. Die hohe Bedeutung dieser Funde führte dazu, dass die Höhle, anfänglich unter „Cueva Eberhardt“ bekannt, in der Folgezeit von zahlreichen Wissenschaftlern aufgesucht und erforscht wurde. Dadurch sammelte sich im Laufe der Zeit eine hohe Fundanzahl an, unter der Mylodon mit Knochenresten, und zahlreichen Koprolithen einen großen Anteil hat. Weitere Funde gehören zu Kamelen wie Lama, Pferden wie Hippidion oder Südamerikanischen Huftieren wie Macrauchenia, zusätzlich sind mehrere Raubtiere vertreten, darunter der Jaguar, Smilodon als Angehöriger der Säbelzahnkatzen und die riesige Bärenform Arctotherium. Einige der Säugetierknochen weisen Marken auf, die ursprünglich mit menschlicher Aktivität in Verbindung gebracht wurden, nach heutiger Ansicht gehen sie aber eher auf Raubtierverbiss zurück. Neben den faunistischen Resten barg die Höhle auch eine Unzahl an botanischem Material. Sie lieferte auch eine der umfangreichsten Datensequenzen aus dem Oberen Pleistozän. Mehrere Radiocarbondaten, gemessen an den unterschiedlichsten Funden von Mylodon, reichen über einen Zeitraum von vor rund 16.700 bis vor 10.200 Jahren zurück. Die oberen Angaben gehören zu den jüngsten, die direkt an Funden des Faultierverteters gewonnen wurden.[47][42][48][43][49]
Mit rund 11.480 bis 11.250 Jahren vor heute ähnlich junge Daten lieferte unter anderem auch die Fundstelle Baño Nuevo im zentral-südlichen Chile. Die Osteoderme, von denen diese Werte stammen, lagerten aber in stratigraphisch jüngeren Schichten mit archäologischem Kontext. Cueva Lago Sofía, eine Gruppe mehrerer kleiner Höhlen unweit nördlich der Cueva del Milodón, weist vergleichbare Alterswerte auf. Die kleine, dunkle Felskammer Cueva Lago Sofía 4 enthielt zahlreiche Knochen, darunter vier Jungtiere von Mylodon und mehr als 4200 Knochenplättchen, deren Alter zwischen 13.400 bis 11.050 Jahren liegt. Noch jünger sind Datierungen einer Mylodon-Rippe, die assoziiert mit drei Dutzend Knochenplättchen an der benachbarten Cueva Lago Sofía 1 zu Tage kam und mit einem Alter von 9700 Jahren bereits dem Unteren Holozän angehört. Der hier bedeutende archäologische Fundbereich mit Steinartefakten, Feuerstellen und Knochen mit Schnittmarken ist dagegen älter und kann in einen Zeitraum um 11.000 Jahre eingestuft werden, während sich weitere Funde von Mylodon noch einmal knapp 2000 Jahre zuvor abgelagert hatten. Einige Fossilien von Mylodon stammen aus Schichten, die noch deutlicher in das Holozän hineinstreuen. Hier fehlen zumeist direkte Altersbestimmungen der Knochen, ihr Fundzusammenhang wird daher in der Regel als problematisch angesehen. Grund dafür ist, dass in vielen Höhlen-, aber auch auf Freilandfundstellen zahlreiche natürliche und anthropogene Prozesse einwirken, die zur Verlagerung von Knochen oder Osteodermen führen können, seien es die Wühltätigkeiten unterirdisch lebender Tiere oder unterschiedlichste menschliche Aktivitäten.[20] Damit starb Mylodon wie zahlreiche andere Formen der großen Bodenfaultiere aller Wahrscheinlichkeit nach im Übergang vom Pleistozän zum Holozän aus.[50][51][52][42][43]
Paläobiologie
Ernährungsweise

Die Mylodonten gelten häufig aufgrund ihres Zahnbaus mit ebenen Kauflächen auf den molarenartigen Zähnen als ausgesprochene Grasfresser. Dies findet bedingt auch durch die hohen (hypsodonten) Zahnkronen und das breite Maul bei zahlreichen Formen Unterstützung. Als analoge Beispiele werden zumeist die Huftiere herangezogen, bei denen Formen mit hohen Zahnkronen und breitlippigen Mäulern in der Regel grasfressend sind, etwa verschiedene Rinder, die Pferde oder das Breitmaulnashorn. Demgegenüber ernähren sich solche mit niedrigen Zahnkronen und schmalen Schnauzen wie die Ducker oder das Spitzmaulnashorn weitgehend selektiv von verschiedenen Blättern und weiterer weicher Pflanzenkost. Im Gegensatz zu anderen großen mylodonten Faultieren wie Glossotherium, Paramylodon oder Lestodon ist das Maul bei Mylodon relativ schmal gestaltet. Als Besonderheit findet sich der geschlossene Nasenbogen, der in seinem vorderen Bereich stark aufgeraut ist und so Muskelansatzstellen für eine bewegliche Oberlippe bietet. Ähnliches lässt sich zu einzelnen Vertiefungen in der Umgebung des Foramen infraorbitale sagen, die ebenfalls als Ansatzpunkte einzelner Muskelstränge des Nasen-Lippen-Bereiches fungierten. Möglicherweise war Mylodon daher stärker an gemischte Pflanzenkost angepasst, die mit Hilfe einer beweglichen Oberlippe aufgenommen wurde. Der Verlust der vorderen Zähne in der oberen Zahnreihe führt darüber hinaus zu der Annahme, dass vergleichbar den Rindern eine hornartige Struktur am Mittelkieferknochen ausgeprägt war, die zum Abzupfen der Nahrung eingesetzt werden konnte.[53][7]
Die gesamte vordere Schädelstruktur ist bei Mylodon relativ massiv gebaut, verbunden mit einer teils verknöcherten Nasenscheidewand lässt sich annehmen, das beim Zerkleinern der Nahrung relativ hohe Kaukräfte wirkten. Abweichend von den teils riesigen Vertretern der Megatheriidae saß bei den Mylodonten die Gelenkung zwischen dem Unterkiefer und dem Schädel relativ niedrig, etwa auf Kauhöhe der Zähne. Der dadurch sich verringernde Hebelarm des Massetermuskels erfährt durch die Struktur des Jochbogens, hauptsächlich des absteigenden Fortsatzes, eine gewisse Kompensation, so dass bezüglich der Beißkraft nur geringe Unterschiede zu den Megatherien bestanden haben dürften. Das ausgedehnte Unterkiefergelenk ermöglicht eine weite Bewegungsfreiheit beim Kauen. Dem steht aber wiederum der Jochbogen gegenüber, der nicht geschlossen ist und dadurch den entgegenwirkenden Kräften des Masseter- und des Flügelmuskels (Musculus pterygoideus) nur wenig standhalten konnte. Es ist daher anzunehmen, dass bei Mylodon vor- und rückwärtsgerichtete Kaubewegungen dominierten.[53][7] Die flachen Zahnkronen führen zu einer vergleichsweise geringen Größe der insgesamt verfügbaren Kaufläche. Bei Mylodon beträgt diese gut 1320 mm² entsprechend zu etwa gleich großen anderen Mylodonten. Das in seinen Ausmaßen vergleichbare Panzernashorn besitzt demgegenüber mit 2660 bis 5190 mm² gut den doppelten bis vierfachen Wert. Ähnlich verhält es sich beim Flusspferd, dessen Gesamtkaufläche sich auf insgesamt zwischen 3290 und 5410 mm² beläuft. Die geringe Gesamtkaufläche der Zähne bei Mylodon hatte wohl eine eher niedrige Verarbeitungskapazität für die Nahrung im Maul zur Folge. Daraus kann entweder eine hohe Fermentationsrate im Magen-Darm-Trakt und/oder ein sehr langsamer Stoffwechsel geschlussfolgert werden. Bei heutigen Faultieren ist letzteres der Fall. Verantwortlich dafür ist eine lange Durchlaufzeit der Nahrung von bis zu einer Woche durch den großen, mehrfach gekammerten Magen. Es ist anzunehmen, dass dies auch auf die ausgestorbenen Faultiere zutrifft. Eventuell war dadurch der Magen der Mylodonten ein funktionales Äquivalent zu dem komplexen Magen der Wiederkäuer, wodurch eine lange Durchlaufzeit der Nahrung eine effiziente Verdauung ermöglichte, bei der auch schwerer zugängliche Nährstoffe bereitgestellt werden konnten, beispielsweise aus stärker faserhaltiger Nahrung. Ein derartiges Verdauungssystem könnte die niedrige Verarbeitungsmenge im Maul und somit letztendlich auch die geringe Gesamtkaufläche bei Mylodon ausgeglichen haben.[54][55]
Eine direkte Analyse der genutzten Nahrungsressourcen ist unter anderem durch die zahlreichen Dungreste in Form von Koprolithen möglich, die unter anderem aus der Cueva del Milodón im chilenischen Teil Patagoniens, aber auch aus anderen Höhlen vorliegen. Die Koprolithen weisen bei Mylodon Durchmesser bis zu 18 cm Durchmesser auf.[49] Untersuchungen der Pflanzenreste ergaben zu 80 bis 95 % Süßgräser und zu 5 bis 20 % Sauergräser. Krautige Pflanzen ließen sich dagegen nur in Spuren nachweisen. Demnach ernährte sich Mylodon zumindest im südwestlichen Patagonien fast ausschließlich von Gräsern. Die Nahrung spiegelt sich im Paläohabitat wider, da Pollenanalysen zeigen, dass die Landschaft zu jener Zeit einen tundrenartigen Charakter trug und daher nahezu baumfrei war mit nur vereinzelten niedrigen Gebüschen. Gelegentlich auftretende Nachweise von Scheinbuchen werden als durch Wind angetragene Pollen interpretiert.[56][57][58] Abweichend zu den Befunden aus den Koprolithen weisen Isotopenanalysen an fossilen Haaren von Mylodon durch auffällige Stickstoffwerte auch auf eine gewisse tierische Nahrungskomponente hin. Dadurch könnten die Tiere eher als Allesfresser eingestuft werden. Da Knochenteile bisher in den Dungresten fehlen, nutzten sie vermutlich weichere und weniger verdauungsbeständige Ressourcen wie Fleisch und Eier. Ein tierischer Anteil in der Ernährung begründet sich möglicherweise auch in der im Vergleich zu anderen Mylodonten recht schmalen Schnauze und geringeren gesamten Kapazität an Kaufläche. Ebenso begünstigte sie die weit südliche Verbreitung der Gattung außerhalb der warmen tropisch- bis subtropisch geprägten Landschaften. Der Körperbau von Mylodon spricht aber gegen einen aktiven Jäger oder Beutegreifer, wodurch wohl eine aasfressende Lebensweise in Betracht kommt.[59]
Fortbewegung
Allgemein handelt es sich bei den großen Mylodonten um bodenbewohnende Tiere. Der gegenüber dem oberen sehr kurze untere Abschnitt des Hinterbeins findet sich auch bei Mylodon wieder, dessen Schienbein mit 27 cm Länge nur halb so lang ist wie der Oberschenkelknochen mit 59 cm Länge. Im Vergleich dazu besitzen die Megatheriidae deutlich längere untere Beinabschnitte, was etwa beim nahezu gleich großen Pyramiodontherium zu einem 47 cm langen Schienbein bezogen auf einen 49 cm langen Oberschenkelknochen führt. Möglicherweise resultieren diese Differenzen in der Hinterbeinstruktur zu einer deutlich agileren Fortbewegung bei den Megatherien im Verhältnis zu den Mylodonten.[60] Ähnlich wie bei anderen großen Bodenfaultieren kontaktierte die Hand von Mylodon den Untergrund mit der äußeren Seitenkante und saß somit gedreht auf. Indiziert wird dies durch die langen Mittelhandknochen der äußeren Strahlen und die abnehmende Anzahl der Fingerglieder an diesen. Die spezielle Handposition schützte damit die langen Krallen der Innenstrahlen, die so beim Gehen nicht in den Untergrund eindrangen. Eine funktional ähnliche, nur grundsätzlich andere Handposition findet sich im Knöchelgang des entfernt verwandten heutigen Großen Ameisenbären. Das Ellenbogengelenk war im vierfüßigen Stand leicht nach außen gerichtet und die Arme somit etwas nach innen gewinkelt, was sich aus der Lage des Olecranons ergibt. Die Hände kamen damit leicht innerhalb der Ellenbogenbreite zur Ruheposition. Eine derartige Ausrichtung der Arme kann die große Masse von Mylodon effektiv unterstützen. Die Hände würden dadurch auch in einer Linie zu den Füßen stehen, was unter anderem auch Trittsiegel von Paramylodon vermitteln. Die seitlich begrenzte Gelenkfläche des Orberamkopfes schränkte die Beweglichkeit des Arms stark ein. Gleiches gilt für den Unterarm, dessen gerade Speiche mit seitlich verlängertem Kopf keine größeren Drehbewegungen zuließ. Diese Merkmale können als Anpassungen an eine rein landbewohnende Lebensweise gedeutet werden. Zuletzt seien noch auf die Muskelansatzstellen am ersten Halswirbel verwiesen, die stärker ausgebildet sind als etwa bei Paramylodon. Korrespondierend dazu stehen auch die Hinterhauptsgelenkflächen etwas weiter auseinander. Beides kann dahingehend gedeutet werden, dass der massivere Schädel von Mylodon, hervorgerufen durch die Verlängerung der Schnauzenregion, eine größere Muskelunterstützung benötigte.[11][10]
Für einige der Mylodonten Südamerikas wie Glossotherium wird eine teils grabende Lebensweise rekonstruiert, was sich unter anderem aus dem Bau des Vorderbeins ergibt. Ein Indikator dafür stellt der obere Gelenkfortsatz (Olecranon) der Elle dar. Je länger das Olecranon ist, um so höher wird dabei die Hebelwirkung des Unterarms, da mehr Ansatzfläche für die Unterarmmuskulatur zur Verfügung steht. Bei Glossotherium nimmt das Olecranon bis zu 35 % der Gesamtlänge der Elle ein, die daraus resultierende Befähigung zum Graben wäre vergleichbar mit den Kugelgürteltieren, die zwar selten eigene Baue anlegen, dazu aber in der Lage sind.[61] Die bisherigen Analysen für Mylodon ergaben ein wesentlich kürzeres Olecranon, dessen Anteil an der Gesamtlänge der Elle nur rund 22 % beträgt. Problematisch ist allerdings der Umstand, dass sich die Werte bei Mylodon auf ein nicht vollständig ausgewachsenes Exemplar beziehen.[11] Andere Hinweise lassen sich aus dem Bau der Hand ableiten. So sind bei Mylodon die Mittelhandknochen des zweiten und dritten Strahls sehr grazil, abweichend von Glossotherium. Gerade ein schwach ausgeprägter Mittelstrahl scheint eine grabende Tätigkeit nicht zu unterstützen, da dieser bei unterirdisch lebenden Säugetieren zumeist am kräftigsten ausgebildet ist. Allerdings zeigt die körperferne Gelenkfazette des dritten Mittelhandknochens eine flache Ausprägung, wodurch der mittlere Finger generell eher steif und stabil war. Die gleiche Artikulationsfläche am zweiten Mittelhandknochen ist deutlich gerundeter und unterstützt somit eine höhere Mobilität des Fingers beim Greifen. Dadurch bestanden offensichtlich Funktionsunterschiede zwischen den einzelnen Strahlen der Hand. Als zusätzliches Indiz gegen grabende Tätigkeiten können die selten auftretenden Abnutzungsspuren an den letzten Fingergliedern herhalten, die unter anderem mehrfach isoliert aus der Cueva del Milodón vorliegen. Die aufgezeigten Befunde negieren jedoch nicht das gelegentliche Kratzen im Untergrund, etwa bei der Suche nach Nahrung oder ähnlichem, wozu die Struktur des Vorderarms mit einzelnen markanten Muskelmarken am Unterarm offensichtlich kräftig genug ausgebildet ist.[12][10]
Ökologische Anpassungen

Mylodon kam von Feuerland an der Südspitze Südamerikas über Patagonien bis in die Pamparegion vor und besaß so ein Verbreitungsgebiet, das die kalt- und kühl-gemäßigten bis teils subtropischen Klimazonen Südamerikas einschließt. Vor allem die zahlreichen Nachweise in Patagonien und auf Feuerland aus der ausgehenden letzten Kaltzeit, die auf Pollenanalysen basierte Rekonstruktion der damaligen Umwelt als tundrenartige offene Landschaften, die sich daraus ergebende überwiegend grashaltige Ernährungsweise und die Überlieferung von Fellresten weisen Mylodon als kälteangepasstes Tier aus. Dies unterstützen auch Berechnungen der Wärmeleitfähigkeit des Fells, die sehr niedrig war und somit die Thermoregulation unterstützte. Der Wert von rund 1,9 ist etwa nur ein Fünftel dessen, was bei einem Tier von rund 1 t Lebendgewicht zu erwarten wäre. Teilweise wird dies auf die geringe Stoffwechselrate zurückgeführt. Sofern letzteres für Mylodon auch zutrifft, könnte dies eine Thermoneutralität von etwa 28,5 °C bewirkt haben, was noch weit höher ist als bei den rezenten Faultieren. Da die Lufttemperaturen im ausgehenden Pleistozän aber häufig darunter gelegen haben dürften, ist ausgehend von diesen Berechnungen ein phasenweise erhöhter Metabolismus anzunehmen um die Körpertemperatur weitgehend konstant zu halten.[62]
Andererseits sind Funde von Mylodon auch aus deutlich weiter nördlich gelegenen Gebieten gut belegt. Für die Reste aus der El-Palmar-Formation in der argentinischen Provinz Entre Ríos wird ein Alter in die ausgehende letzte Warmzeit, genauer gesagt in das Ende des Wärme-Isotopenstadiums (OIS oder MIS) 5 vor gut 80.000 Jahren angenommen. Die damalige Umwelt bestand dort aus sich abwechselnden geschlossenen Wäldern und offeneren Grasgebieten in einer Landschaft, die von zahlreichen Wasseradern im Einzugsgebiet des heutigen Río Uruguay durchzogen war und die unter warm-feuchten Klimabedingungen bestand.[5] Da Mylodon hier aber noch im ausgehenden Pleistozän unter vergleichbaren Klimabedingungen anwesend war,[28] kann dadurch von einer deutlich größeren ökologischen Toleranz bei den Tieren ausgegangen werden. Es ist zu vermuten, dass dies auch auf die Ernährung zutrifft. Die waldreicheren Landschaften in den nördlicheren Refugien des Faultierbertreters stellten weniger Grasnahrung zur Verfügung, wie auch die bereits erwähnten Schädeladaptionen eine eher auf gemischte Pflanzenkost basierte Ernährung befürworten.[5]
Anhand einzelner Funde lassen sich gewisse Größendiffernzen bei Mylodon feststellen. So verweisen Fossilreste von nördlicheren Fundpunkten auf durchschnittlich kleinere Individuen hin als solche aus südlicheren. Prinzipiell könnte dies die Bergmannsche Regel widerspiegeln, möglich ist aber auch, dass sich hier ein bisher nicht festgestellter Sexualdimorphismus ausdrückt. Allgemein wird das verfügbare Fossilmaterial als bisher zu wenig umfangreich angesehen, um eine genauere Geschlechtsbestimmung eines Individuums zuzulassen. Bei anderen großen Bodenfaultieren wie etwa Paramylodon gibt es aber durchaus Hinweise auf einen Sexualdimorphismus, ebenso bei den rezenten Baumfaultieren, der sich hier aber nicht auf die Körpergröße, jedoch auf das Körpergewicht auswirkt. Andererseits bezeugen Mylodon-Funde aus sehr weit südlichen Bereichen auch recht kleine Individuen, was teilweise mit deren Auftreten in gebirgigen Lagen in Verbindung gebracht wird.[5]
Beutegreifer und Parasiten
Vor allem im südlichen und südwestlichen Patagonien lassen sich zahlreiche Knochenveränderungen an Funden von Mylodon als durch Raubtiere verursacht belegen. Hierzu gehören vor allem die Reste aus der Cueva del Milodón im südwestlichen Chile. Einige Höhlen in deren direkter Umgebung wie die Cueva Lago Sofía 4 und die Cueva Chica werden als Horste von Raubtieren gedeutet.[63][46] Gleiches gilt für die Cueva del Puma oder die Cueva Fell im Pali-Aike-Gebiet des südlichen Chile. Einige der Höhlen enthalten vornehmlich kleinere Skelettelemente wie Hand- und Fußknochen oder Knochenplättchen, die darauf hinweisen, dass nur ein Teil des Kadavers in den Unterschlupf verschleppt wurde. Ob dies ein Resultat eines direkten Beutezugs oder auf Aasfresserei zurückzuführen ist, lässt sich vielfach nicht direkt klären. Andere Felskammern bargen wiederum einen größeren Anteil an Jungtieren von Mylodon.[64][42] Die größten damals auftretenden Raubtiere stellen der Puma und der Jaguar sowie die ausgestorbene Formen Smilodon als Vertreter der Säbelzahnkatzen und Arctotherium als Angehöriger der Bären dar. Letztere beiden konnten rekonstruiert durchaus Körpergewichte von über 400 kg erreichen, wobei für die Säbelzahnkatze Beutegrößen zwischen 1 und 2 t angenommen werden, wodurch Smilodon durchaus als großer Predator von Mylodon in Betracht kommt.[65][64][42][66]
An verschiedenen der untersuchten Koprolithen von Mylodon konnten noch Eier von Fadenwürmern dokumentiert werden. Die Eier waren ovaloid geformt mit Längen von knapp 50 µm Länge und Dicken von 29 µm.[67] Außerdem ließen sich einzelne Käfer nachweisen.[49]
Systematik
Klassische äußere Systematik
Innere Systematik der Mylodontidae nach Boscaini et al. 2019 (basierend auf skelettanatomischen Analysen)[68]
|
Mylodon ist eine ausgestorbene Gattung aus der ebenfalls erloschenen Familie der Mylodontidae. Die Mylodontidae bilden einen Teil der Unterordnung der Faultiere (Folivora) und formen innerhalb dieser zusammen mit den Orophodontidae und den Scelidotheriidae die Überfamilie der Mylodontoidea (teilweise werden die Scelidotheriidae und die Orophodontidae aber auch nur als Unterfamilie innerhalb der Mylodontidae geführt[69]). In einer klassischen Auffassung, die weitgehend die skelettanatomischen Merkmale berücksichtigt, repräsentieren die Mylodontoidea wiederum neben den Megatherioidea die zweite große und bedeutende Faultierlinie. Die Megatherioidea schließen die Megatheriidae mit den größten bekannten Vertretern der Faultiere, darüber hinaus auch die Nothrotheriidae und die Megalonychidae ein. Letzteren werden die Zweifinger-Faultiere (Choloepus) als eine der beiden heute noch bestehenden Linien zugewiesen. Die Mylodontidae innerhalb der Mylodontoidea stellen eine der vielfältigsten Gruppen der Faultiere dar. Zu ihren wichtigsten definierenden Merkmalen gehören die hochkronigen Zähne, die abweichend von den Megatherioidea eine flache (lobate) Kaufläche aufweisen. Diese Zahngestaltung wird zumeist als eine Anpassung an stärker grashaltige Nahrung aufgefasst. Die hinteren Zähne sind im Querschnitt häufig rund oder oval gestaltet, der jeweils vorderste einer Zahnreihe ist in der Regel eckzahnartig ausgeprägt. Der Hinterfuß ist zudem deutlich gedreht, so dass die Sohle nach innen zeigt.[25][70] Die Mylodonten lassen sich mit Paroctodontotherium aus Salla-Luribay in Bolivien bereits im Oligozän nachweisen.[71]
Die innere Gliederung der Mylodontidae ist komplex und momentan ungenügend ausgearbeitet. Zumeist anerkannt sind die stammesgeschichtlich entwickelten Linien der Mylodontinae mit Mylodon als Typusform und der Lestodontinae, deren Charakterform Lestodon darstellt (auf tribaler Ebene als Mylodontini und Lestodontini bezeichnet). Zu letzterer werden teilweise auch Paramylodon und Glossotherium gerechnet.[72] Neben diesen beiden Gruppen wurden in der Vergangenheit zahlreiche weitere Unterfamilien aufgestellt. Genannt werden sollen etwa die Nematheriinae, die die Vertreter aus dem Unteren Miozän zusammenfassen oder die Octomylodontinae für alle Basalformen. Diese sind aber nicht allgemein anerkannt.[73] Eine weitere Linie wurde mit den Urumacotheriinae im Jahr 2004 herausgearbeitet, welche die spätmiozänen Vertreter des nördlichen Südamerikas beinhalten.[74] Prinzipiell mahnen einige Forscher eine Revision für die gesamte Familie an, da zahlreiche der höheren taxonomischen Einheiten keine formale Diagnose besitzen.[75]
Für die Untergliederung der terminalen Vertreter der Mylodonten in die Unterfamilien der Lestodontinae und Mylodontinae fand sich allerdings in einer der bisher umfangreichsten Studien zur Stammesgeschichte der Faultiere Bestätigung. Die Untersuchung wurde 2004 von Timothy J. Gaudin publiziert, beruht auf schädelanatomischen Merkmalen und legt eine nahe Beziehung von Mylodon mit Paramylodon nahe, während Glossotherium enger mit Lestodon verbunden ist. Letztere beiden zeigen aber starke anatomische Ähnlichkeiten zu Paramylodon.[76] Zwar konnte diese Verwandtschaftsbeziehung in der Folgezeit mehrfach reproduziert werden,[77][31] doch andere Autoren sahen dies mitunter auch kritischer. Hier sei beispielhaft eine Untersuchung von Luciano Varela und Kollegen aus dem Jahr 2019 unter Einbeziehung zahlreicher Taxa aus der gesamten Unterordnung der Faultiere erwähnt, in der Mylodon eine basale Stellung gegenüber allen späten Mylodonten einnimmt.[69] Eine höherauflösende phylogenetische Analyse der Mylodonten von einem Arbeitsteam um Alberto Boscaini aus dem gleichen Jahr untermauert dem gegenüber die Zweigliederung der terminalen Vertreter mit dem Unterschied aber zu Gaudins Studie aus dem Jahr 2004, dass Mylodon, Paramylodon und Glossotherium eine engere Einheit formen und den Mylodontinae zuzurechnen sind, Lestodon hingegen enger an Thinobadistes gebunden ist. Als ein fundamentaler Unterschied zwischen den Mylodontinae und Lestodontinae kann die Ausprägung der eckzahnartigen vorderen Zähne hervorgehoben werden. Bei letzteren sind sie groß und durch ein langes Diastema von den hinteren Zähnen getrennt, bei ersteren haben sie nur kleine Ausmaße oder sind teilweise reduziert und stehen dichter an den molarenartigen Zähnen an.[68] Eine bereits 2009 von Robert K. McAfee publizierte Studie widmete sich detailliert den schädel- und zahnmorphologischen Unterschieden zwischen Glossotherium sowie Paramylodon und bezog auch Mylodon mit ein. Demnach bestehen zwischen ersteren beiden größere Gemeinsamkeiten als zu letzterem. Die Besonderheit von Mylodon drückt sich in dem reduzierten Gebiss aus, bei dem die oberen eckzahnartigen Zähne fehlen, die unteren aber in molarenartige umgestaltet sind. Zusätzlich hebt sich die Gattung durch einfacherer gestaltete Mahlzähne und eine vorverlagerte Knochennaht zwischen dem Gaumenbein und dem Oberkiefer auf Höhe des dritten molarenartigen Zahns hervor. Glossotherium und Paramylodon auf der anderen Seite verfügen über caniniforme Zähne, wenn auch in unterschiedlicher Ausprägung, komplexere molariforme Zähne und eine Gaumenbein-Oberkiefer-Verbindung auf Höhe des letzten Backenzahns.[4]
Genetische Verwandtschaft
Innere Systematik der Faultiere nach Presslee et al. 2019 (basierend auf Proteinanalysen)[78]
|
Aufgrund der guten Fossilerhaltung mit Resten des Weichteilgewebes gehörte Mylodon neben Nothrotheriops zu den wenigen ausgestorbenen Faultieren, die bereits sehr früh in molekulargenetische Untersuchungen einbezogen wurden. Erste Analysen fanden in der Wende vom 20. zum 21. Jahrhundert statt. In deren übereinstimmenden Ergebnissen kristallisierte sich ein engeres Verwandtschaftsverhältnis zwischen Mylodon und den Zweifinger-Faultieren (Choloepus) heraus, womit letztere der übergeordneten Gruppe der Mylodontoidea zuzuweisen wären. Die Dreifinger-Faultiere (Bradypus) hingegen standen Nothrotheriops aus der Gruppe der Nothrotheriidae näher und gehörten damit den Megatherioidea an.[79][80][81][82][83][84] Dies erwies sich als konträr zu der klassischen, anatomiebasierten Gliederung der Faultiere, in der die Zweifinger-Faultiere eine Gemeinschaft mit den Megalonychidae innerhalb der Megatherioidea bilden, die Dreifinger-Faultiere hingegen die Schwestergruppe zu allen anderen Faultierlinien darstellen.[76] Etwas widersprüchlich dazu zeigten sich Kollagenuntersuchungen aus dem Jahr 2015, die neben den heutigen Faultieren auch weitere ausgestorbene Formen berücksichtigten. Hier ergab sich vorläufig eine Nahbeziehung der rezenten Baumfaultiere zueinander. Allerdings war Mylodon kein Bestandteil dieser Analyse.[85] Als problematisch bei allen Arbeiten kann herausgestellt werden, dass zu wenige fossile Arten einbezogen wurden und die ermittelten Daten so uneindeutig blieben. Eine separate Studie zur genetischen Verwandtschaft von Mylodon zu den heutigen Faultieren unter Berücksichtigung sowohl von mitochondrialer als auch Kern-DNA aus dem Jahr 2018 bestätigte das zuvor mehrfach ermittelte Ergebnis einer engen Bindung an die Zweifinger-Faultiere.[86]
Im Jahr 2019 fanden zwei umfangreiche Untersuchungen zur Molekulargenetik und zur Proteinstruktur ihre Veröffentlichung, in der erstmals auch eine nennenswerte Anzahl an Vertretern fast aller fossilen Familien integriert waren. Erstere fand unter Federführung von Frédéric Delsuc statt, der bereits zuvor über Jahre die genetische Beziehung der heutigen Nebengelenktiere erforscht hatte, letztere stammt maßgeblich von Samantha Presslee. In beiden Studien ließen sich die auch anatomisch belegten Großgruppen der Mylodontoidea und Megatherioidea weitgehend bestätigen. Allerdings konnten ihnen mit den Megalocnoidea eine dritte große Linie zur Seite gestellt werden, die die Faultiere der karibischen Inseln repräsentiert. Die rezenten Faultiere verteilen sich entsprechend der vorigen Studien mit den Dreifinger-Faultieren auf die Megatherioidea und mit den Zweifinger-Faultieren auf die Mylodontoidea. Das in diesem Punkt von den anatomischen begründeten Gliederungsschemata der Faultiere abweichende Bild hatte zur Folge, dass einerseits die Dreifinger-Faultiere trotz einiger besonderer Charakteristika keine Außengruppe bilden und die Zweifinger-Faultiere nicht näher mit den karibischen Faultieren verwandt sind. Letztere beiden – ursprünglich in den Megalonychidae zusammengefasst – waren deutlich getrennt. Die Megalonychidae wurden daher auf die festländischen großen Bodenfaultiere beschränkt, die Zweifinger-Faultiere ausgewiesen und die karibischen Faultiere einer eigenen Großgruppe zugeordnet. Innerhalb der Mylodontidae bildet Mylodon mit Paramylodon und Glossotherium eine enger verwandte Gruppe. Lestodon dagegen steht in einer äußeren Position zu diesen drei Gattungen. Die Konstellation findet sich ähnlich auch bei einigen der bereits erwähnten anatomiebasierten stammesgeschichtlichen Verwandtschaftsmodellen wieder.[87][78]
Innere Systematik
Häufig wird nur eine Art von Mylodon anerkannt:[5]
- M. darwinii Owen, 1840
Daneben wurden zahlreiche weitere Arten innerhalb der Gattung beschrieben. Die bekanntesten sind das im Jahr 1898 von Florentino Ameghino eingeführte M. listai und das auf Lucas Kraglievich aus dem Jahr 1928 zurückgehende M. insigne.[88] Beide Formen werden gelegentlich von einzelnen Autoren als gültig anerkannt, teils aufgrund der geringeren Körpergröße und/oder einzelner abweichender Zahnmerkmale,[29][30] andere sehen sie hingegen als synonym zu M. darwinii an. Gleiches gilt für M. zeballozi, welches im Jahr 1880 von Henri Frédéric Paul Gervais und Ameghino wissenschaftlich benannt worden war.[89] Prinzipiell mahnen aber die Wissenschaftler eine Revision der Gattung an. Eine Untersuchung mehrere Schädel von Mylodon im Jahr 2010 kommt zu dem vorläufigen Schluss, das etwaige Größenunterschiede möglicherweise einen Sexualdimorphismus oder ökologische Differenzen im entsprechenden Lebensraum beziehungsweise zeitliche Unterschiede widerspiegeln könnten.[5] Abseits davon wurde im Jahr 2018 mit Castrocopros hauthali ein Ichnotaxon für die zahlreichen Koprolithen aus der Cueva del Milodón in Chile eingeführt.[90]
Forschungsgeschichte
Erstbeschreibung

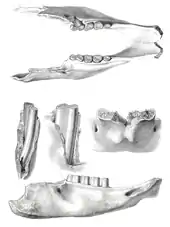
Die taxonomische Geschichte von Mylodon ist komplex. Es kam dabei über einen langen Zeitraum zu Verwechslungen und Gleichsetzungen mit anderen mylodonten Formen wie Glossotherium und Paramylodon. Ein Teil dieser Komplexität lässt sich auf den Erstbeschreiber der Gattung selbst zurückführen, bei dem es sich um Richard Owen (1804–1892) handelt. Owen, einer der bedeutendsten Forscher des Victorianischen Zeitalters, beschäftigte sich von 1836 an mit Fossilfunden, die Charles Darwin von seiner wegweisenden Reise mit der HMS Beagle nach Südamerika mitgebracht hatte. Die Kollektion beinhaltete auch einen Unterkiefer aus Punta Alta bei Bahía Blanca im Süden der argentinischen Provinz Buenos Aires (Exemplarnummer NHM 16617). Das nahezu vollständige Stück zeichnete sich durch eine Zahnreihe aus insgesamt vier molarenartigen Zähnen aus. In einer umfangreichen Schrift aus dem Jahr 1840 verwies Owen den Unterkiefer zu der von ihm neu geschaffenen Gattung Mylodon und benannte die Art mit M. darwinii („darwinii“ ist die von Owen genutzte Schreibweise, in heutiger Zeit wird jedoch häufig auch „darwini“ verwendet; den Regularien der zoologischen Nomenklatur zufolge ist erstere Version korrekt[91]). Den Gattungsnamen bezog er auf die molarenartigen Zähne (von griechisch μυλη (myle) für „Molar“ und ὀδούς (odoús) für „Zahn“, somit also so viel wie „Molarenzahn“ übersetzt), mit dem Artepitheton darwinii ehrte er Darwin als Finder des Belegexemplars. Als zweite Art neben M. darwinii verwies Owen in seiner Schrift auf M. harlani. Diese Form basierte auf einem Unterkiefer und einem Schlüsselbein, welche beide vom Big Bone Lick im Boone County im US-Bundesstaat Kentucky stammten und die Richard Harlan bereits im Jahr 1831 unter der Artzuweisung Megalonyx laqueatus beschrieben hatte.[92] Owen allerdings erkannte im Bau des Unterkiefers Übereinstimmungen mit seinem M. darwinii und benannte Harlans Form um.[93] Harlan nutzte zwei Jahre später einen Aufsatz, um sich über Owens Namenswahl bezüglich Mylodon zu äußern, die er als wenig beschreibend empfand. Demnach könnte seiner Auffassung nach der Name auf nahezu jede ausgestorbene Säugetierform bezogen werden, weil fast alle über die hinteren Backenzähne verfügten. Zudem wäre die Bezeichnung unpassend, da ausgehend von der lateinischen Version dens molaris für „Mahlzahn“ und dem zweiten Wortteil don (von dens für „Zahn“) eine Doppelung entsteht, die mit „Mahlzahn-Zahn“ zu übersetzen wäre.[94] Ungeachtet dessen hatte Owens Etablierung der Gattung Mylodon mit zwei Arten zur Folge, dass der ausgestorbene Faultiervertreter sowohl in Süd- als auch in Nordamerika verbreitet war.[95][96]
Von Glossotherium, Grypotherium und Paramylodon
Owen etablierte in seiner Arbeit von 1840 neben Mylodon auch die Gattung Glossotherium, allerdings ohne Ausweisung einer bestimmten Art. Grundlage bildete hier ein hinteres Schädelfragment, welches vom Flussbett des Arroyo Sarandi im uruguayischen Department Soriano stammt. Im Gegensatz zu Mylodon, für das Owen eine verwandtschaftliche Nähe zu den anderen damals bekannten großen Bodenfaultieren wie Megatherium oder Megalonyx annahm, stellte er Glossotherium in eine Reihe mit den Ameisenbären beziehungsweise mit den Schuppentieren und postulierte eine insektenfressende Lebensweise für die Tiere.[93] Zwei Jahre später aber verwarf Owen den Namen Glossotherium wieder. Dies geschah im Zuge der Bearbeitung eines nahezu vollständigen Skelettes, welches im Jahr zuvor in den Überschwemmungsebenen des Río de la Plata nördlich von Buenos Aires entdeckt worden war. Der weitgehend unversehrte Schädel charakterisierte sich durch eine kurze und breite Schnauze und durch ein Gebiss bestehend aus insgesamt 18 Zähnen, von denen der jeweils vorderste Zahn eine eckzahnartige Gestaltung zeigte. Aufgrund der Ähnlichkeiten im Zahnbau mit den flachen, molarenartigen Zähnen ordnete Owen das Skelett zur Gattung Mylodon und führte die neue Art M. robustus ein. Das Schädelfragment, das er ursprünglich zu Glossotherium gestellt hatte, brachte er nun mit M. darwinii in Verbindung.[97] Im Ergebnis dieser Studie bestanden in den 1840er Jahren bereits drei Arten der Gattung Mylodon.[95][4]
Ein bei Pergamino in der Provinz Buenos Aires aufgefundener Schädel einschließlich Unterkiefer diente dem dänischen Zoologen Johannes Theodor Reinhardt (1816–1882) im Jahr 1879 als Grundlage für eine umfassende Beschreibung. Kennzeichnend für den Schädel war die schmale Schnauze und ein geschlossener Nasenbogen, der durch die feste Verwachsung des Nasenbeins mit dem Mittelkieferknochen entstand. Des Weiteren bestand das Gebiss aus insgesamt 16 Zähnen, der jeweils obere vordere eckzahnähnliche Zahn war reduziert, im Unterkiefer saßen dagegen je vier molarenartige Zähne. Reinhardt bemerkte im Bau des Unterkiefers Ähnlichkeiten zu M. darwinii, in der Schädelgestaltung wich sein Fund durch die schmale Schnauze jedoch deutlich vom breitschnauzigen M. robustus ab. Allerdings ergaben sich nach Reinhardt Übereinstimmungen bei Owens Glossotherium-Schädelfragment mit entsprechenden Schädelabschnitten bei M. robustus. Aufgrund der deutlichen Parallelen zwischen M. darwinii und seinem schmalschnauzigen Schädelfund propagierte Reinhardt die neue Gattung Grypotherium mit Grypotherium darwinii als Typusart.[98] Einen anderen Weg ging Florentino Ameghino (1854–1911) rund zehn Jahre später. Er bestätigte 1889, die Trennung von sowohl von M. darwinii als auch von M. robustus nicht nur auf Art-, sondern auch auf Gattungsebene. Abweichend von Reinhardt, aber übereinstimmend mit Owen sah er den Unterkiefer von M. darwinii und das Schädelfragment von Glossotherium als zusammengehörig an. Da in diesem Szenario Glossotherium Priorität vor Mylodon (Owen erwähnte 1840 ersteres vor letzterem) und Grypotherium besaß, führte Ameghino die Art Glossotherium darwinii ein. Den Status von M. robustus beließ er dagegen unangetastet.[99] Arthur Smith Woodward (1864–1944) wiederum folgte der Argumentation von Reinhardt. In einem im Jahr 1900 erschienenen Aufsatz präsentierte er Funde von Bodenfaultieren des südlichen Patagonien und revidierte gleichzeitig die Sammlung von Charles Darwin. Hierbei setzte er den Unterkiefer von M. darwinii mit Reinhardts Grypotherium gleich und stellte darauf folgend Grypotherium darwinii wieder her. Das Schädelfragment von Glossotherium ordnete Smith Woodward analog zu Reinhardt zu M. robustus.[16][100][95][4]

Im Zeitraum in der Wende vom 19. zum 20. Jahrhundert hatte sich in Nordamerika die Anzahl an fossilen Faultierfunden stark erhöht. Unter anderem entdeckte eine Expedition des American Museum of Natural History im Jahr 1897 ein Teilskelett eines großen mylodonten Faultiers bei Hay Spring im US-Bundesstaat Nebraska. Kennzeichnend am Schädel erwies sich das breite und vorn offene Rostrum und ein Gebiss bestehend aus 16 Zähnen. Hierbei fehlten in der oberen Gebissreihe die vorderen eckzahnartigen Zähne, in der unteren waren sie hingegen ausgebildet. Dadurch ähnelte der Schädel in seiner Gestaltung dem von Glossotherium, das Gebiss wiederum ließ sich eher mit Mylodon vergleichen. Barnum Brown (1873–1963) verwendete den Fund im Jahr 1903 zur Aufstellung der neuen Gattung Paramylodon mit Paramylodon nebrascensis als Typusart. Zu den bereits bekannten Funden von M. harlani aus Nordamerika zeigte sich vor allem die Reduktion der oberen caniniformen Zähne als entscheidend für Brown.[101] Die nordamerikanischen Mylodonten erwiesen sich aber bezüglich dieses Merkmals als recht variabel, was nicht zuletzt dem immensen Fundanstieg in den Asphaltgruben von Rancho La Brea im Süden von Kalifornien und dadurch möglichen Reihenuntersuchungen zu verdanken war. Chester Stock (1892–1950), der sich über einen langen Zeitraum mit den Fossilresten von Rancho La Brea beschäftigt hatte, vereinte daher 14 Jahre später wieder Paramylodon nebrascensis mit M. harlani.[102]
Der zunehmenden Komplexität in Gliederung und Benennung der großen mylodonten Formen Süd- und Nordamerikas widmete sich in den 1920er Jahren besonders Lucas Kraglievich (1886–1932). Unter Zuhilfenahme der Schädelmerkmale arbeitete Kraglievich heraus, dass sich Mylodon, Glossotherium und Grypotherium eindeutig auf zwei unterschiedliche Formen beziehen, die sich auf Gattungsebene trennen lassen. Owens Beschreibung von M. darwinii verband den Gattungsnamen Mylodon fest mit dem Artepitheton. Kraglievich ordnete Gattung und Art unter anderem den vollständigen, schmalschnauzigen Schädel und Unterkiefer von Reinhardts Grypotherium aus Pergamino zu, ebenso Owens Unterkiefer aus Punta Alta aus der Erstbeschreibung der Art als Typusexemplar. Owens breitschnauziges M. robustus verwies er dagegen in die Gattung Glossotherium, welche gleichzeitig auch das Schädelfragment einschloss, das Owen einst zur Beschreibung der Gattung genutzt hatte. Alle nordamerikanischen Funde hingegen beschränkte Kraglievich auf Paramylodon. Dadurch verblieb Mylodon einzig in Südamerika, ließ sich aber hier eindeutig von Glossotherium in Schädel- und Gebissstruktur abgrenzen.[88] Dieser vorgeschlagenen Gliederung schlossen sich in der Folgezeit einzelne Autoren wie etwa Ángel Cabrera an. Im Verlauf des 20. Jahrhunderts kam es dann zu einer kontroversen Diskussion um die Synonymität von Glossotherium und Paramylodon,[103] was sich aber Mitte der 1990er Jahre zugunsten einer Trennung der beiden Formen auflöste.[100][4][5]
Diskurs zur Typusart
Als etwas problematisch gestaltete sich die Bestimmung der Typusart von Mylodon. Owen selbst blieb bei seiner Erstbeschreibung von Mylodon 1840 ungenau in der Definition der Typusart. Zwar basierte er die Gattung auf den von Darwin gefundenen Unterkiefer und somit auf M. darwinii, doch gab er an, dass dies nach M. harlani die zweite beschriebene Art sei.[93] Zwei Jahre später bei seiner Beschreibung von M. robustus wies Owen M. darwinii als primäre Art aus.[93] Im Laufe der Forschungsgeschichte wechselte die Nominatform von Mylodon je nach Bearbeiter. Joseph Leidy übernahm 1855 Owens Sichtweise und listete M. darwinii als erste Art innerhalb der Gattung.[104] Für Reinhardt stellte 1879 M. robustus die Typusform dar, was dem Umstand zuzuschreiben ist, dass er das eigentliche Fundmaterial von Mylodon in die Gattung Grypotherium ausgelagert hatte.[98] Richard Lydekker sah dagegen 1887 in M. harlani die Nominatform,[105] was im Übrigen später auch Barnum Brown befürwortete.[101] Aus heutiger Sicht ist die Frage, ob M. darwinii oder M. harlani als Typusform anzusehen ist, nicht vollständig trivial, da beide Arten unterschiedlichen Gattungen zugewiesen werden und letztere Art durch Erstnennung bei Owen durchaus den Vorrang haben könnte. Kraglievich führte jedoch 1928 in seiner Revision mehrere Gründe auf, warum M. darwinii die eigentliche Nominatform darstellt. Dies sind vor allem die bereits genannten Fakten der Beschreibung von Mylodon auf Basis von Darwins Unterkieferfund und der Listung durch Owen als erste Art. Die Vorgehensweise Kraglievichs wurde in der nachfolgenden Zeit weitgehend anerkannt.[88][4][95]
Domestizierte Faultiere und moderne Mythen
.jpg.webp)
Ein besonderes Kapitel in der Forschungsgeschichte der Gattung Mylodon betrifft die Cueva del Milodón im südwestlichen Patagonien. Kurz nach ihrer Entdeckung durch den deutschen Kapitän Hermann Eberhard im Jahr 1895 und der damit verbundenen Auffindung eines Fellrestes besuchten verschiedenste Wissenschaftler die Höhle, so etwa der schwedische Forscher Otto Nordenskjöld im Rahmen seiner Südamerika-Expedition 1896. Das dabei ebenfalls geborgene Fellstück nebst Fingergliedern wurde später von Einar Lönnberg untersucht.[14] Nur kurz darauf erreichte eine argentinische Gruppe an Naturforschern um Perito Moreno die Region, mit dabei unter anderem Rudolph Hauthal und Emil Racoviță. Moreno selbst betrat die Höhle zwar nicht, seine Begleiter führten dort aber einzelne Untersuchungen durch. Außerdem konnte die Expedition Eberhards originales Fellstück (als Kuriosum in einem Baum hängend) sichern. Dieses bildete später die Grundlage für eine Beschreibung durch Arthur Smith Woodward.[13] Hauthal kehrte 1899 zur Cueva del Milodón zurück und führte umfangreiche Grabungen durch. Hierbei kamen zahlreiche Reste vom Mylodon zum Vorschein. Besondere Aufmerksamkeit erhielt eine Ansammlung von Geröllen im hinteren Teil der Höhle, die scheinbar linear angeordnet lagen. Außerdem fanden sich Konzentrationen von Gräsern und Koprolithen. In einer noch im gleichen Jahr erschienenen Publikation deuteten Hauthal und seine Kollegen die Steinstruktur als eine Art Gatter oder Pferch, in dem die riesigen Faultiere von den damaligen Menschen gehalten wurden, und die Gräser als Vorratshaltung von Nahrung für die Tiere. Darüber hinaus präsentierten sie ein weiteres Fellstück, das einen vorgeblich intentionell durch Menschen verursachten Schnitt aufwies und die Bearbeiter an abgezogene Haut denken ließ. Santiago Roth, der die Reste von Mylodon untersuchte, etablierte darauf basierend im Jahr 1899 die Art Grypotherium domesticum. Der Artzusatz verweist auf die angenommene Domestizierung von Mylodon durch den Menschen.[106] In Teilen widersprach nur wenig später Otto Nordenskjölds Cousin Erland, der im gleichen Zeitraum Untersuchungen in der Cueva del Milodón durchführte, der Interpretation, da er unter anderem eine deutlich weiterreichende Verbreitung der Dungreste beobachtete. Außerdem vermutete er, dass Mylodon in der Höhle nicht durch Menschen gehalten wurde, vielmehr interpretierte er die Knochen als Beutereste von Raubtieren.[107][108] Aus heutiger Sicht ließ sich die Vorstellung eines gezähmten Faultiers nicht bestätigen. Die Steine, die den vermeintlichen Pferch bildeten, werden als natürliche Anhäufung gesehen, entstanden durch das fortwährende Abbrechen von der Höhlendecke im Laufe der letzten 12.000 Jahre. Zudem ist der Einfluss des frühen Menschen auf die Höhle und ihre Nutzung weitaus geringer als anfänglich angenommen.[50][49][44]
Eng mit der Erforschung der Cueva del Milodón ist auch die Vorstellung verbunden, Mylodon habe bis in historischer Zeit überlebt. Verantwortlich dafür ist Florentino Ameghino, ein argentinischer Naturforscher, der Ende des 19. Jahrhunderts den von Moreno geborgenen Fellrest aus der Cueva del Milodón aufgrund der ausgezeichneten Erhaltung als nur wenig alt einstufte. Dessen rötliche Fellfarbe erinnerte Ameghino an einen Bericht von Ramón Lista, ein Entdecker und der Gouverneur der argentinischen Provinz Santa Cruz, der ihm zu Ohren gekommen war. Dem zufolge habe Lista Ende der 1880er Jahre des Nachts in Santa Cruz auf ein Tier geschossen, welches aber entkam. Das Tier wies Listas Beschreibung zufolge ein rotes Fell auf, ähnelte aber ansonsten äußerlich einem Schuppentier. Ameghino war davon überzeugt, dass das Tier aus Listas Erzählung und der Fellrest der gleichen Art angehörten. Bezogen auf die in der Haut eingebetteten Knochenplättchen identifizierte er dieses als einen kleinen Vertreter der Mylodonten. Außerdem verband er Listas Geschichte mit Überlieferungen indigener Einwohner wie den Tehuelche über ebenfalls rothaarige Tiere, die mit ihren Krallen Erdhöhlen aushoben und angeblich eine undurchdringliche Haut aufwiesen. Für dieses Tier und zu Ehren Listas kreierte Ameghino im Jahr 1898 die Artbezeichnung Neomylodon listai. Gleichzeitig regte er in einzelnen Veröffentlichungen zur Suche nach dem Tier an.[109][110] Moreno zweifelte die Existenz eines modernen Bodenfaultiers an und führte die gute Konservierung der Fellreste auf die besonderen Bedingungen der Cueva del Milodón zurück, ebenso wie er die Berichte der Tehuelche über ein „Ellengassen“ genanntes Tier als fiktional einstufte.[111][13] Die Ausführungen Ameghinos veranlassten jedoch wiederum Hesketh Vernon Hesketh-Prichard im Jahr 1900 zu einer Expedition nach Patagonien, um das vermeintlich existierende Mylodon aufzuspüren. Seine Reisebeschreibungen veröffentlichte Hesketh-Prichard im Jahr 1902 unter dem Titel Through the heart of Patagonia, musste aber feststellen, dass weder dies- noch jenseits der Anden Spuren von Mylodon zu finden waren.[112][2] In dem 1977 erschienenen Reiseroman In Patagonia (deutsch In Patagonien) verarbeitete Bruce Chatwin die Geschichten um Mylodon und die Cueva del Milodón.[113]
Literatur
- Diego Brandoni, Brenda S. Ferrero, Ernesto Brunetto: Mylodon darwini Owen (Xenarthra, Mylodontinae) from the Late Pleistocene of Mesopotamia, Argentina, with Remarks on Individual Variability, Paleobiology, Paleobiogeography, and Paleoenvironment. In: Journal of Vertebrate Paleontology. Band 30, Nr. 5, 2010, S. 1547–1558.
- Richard A. Fariña, Sergio F. Vizcaíno, Gerardo De Iuliis: Megafauna. Giant beasts of Pleistocene South America. Indiana University Press, Bloomington 2013, ISBN 978-0-253-00230-3, S. 206–209.
Einzelnachweise
- Per Christiansen, Richard A. Fariña: Mass estimation of two fossil ground sloths (Mammalia, Xenarthra, Mylodontidae). In: Senckenbergiana biologica. Band 83, Nr. 1, 2003, S. 95–101.
- C. M. Bell: Did elephants hang from trees? The giant sloths from South America. In: Geology Today. Band 18, Nr. 2, 2002, S. 63–66.
- Analia M. Forasiepi, Agustin Martinelli, Jorge Blanco: Bestiario fósil: Mamíferos del Pleistoceno de la Argentina. Buenos Aires, 2007, S. 1–181 (S. 60–61).
- Robert K. McAfee: Reassessment of the cranial characters of Glossotherium and Paramylodon (Mammalia: Xenarthra: Mylodontidae). In: Zoological Journal of the Linnean Society. Band 155, 2009, S. 885–903.
- Diego Brandoni, Brenda S. Ferrero, Ernesto Brunetto: Mylodon darwini Owen (Xenarthra, Mylodontinae) from the Late Pleistocene of Mesopotamia, Argentina, with Remarks on Individual Variability, Paleobiology, Paleobiogeography, and Paleoenvironment. In: Journal of Vertebrate Paleontology. Band 30, Nr. 5, 2010, S. 1547–1558.
- Luciano Brambilla, Damián A. Ibarra: The occipital region of late Pleistocene Mylodontidae of Argentina. In: Boletín del Instituto de Fisiografía y Geología. Band 88, 2018, S. 1–9.
- M. Susana Bargo, Sergio F. Vizcaíno: Paleobiology of Pleistocene ground sloths (Xenarthra, Tardigrada): biomechanics, morphogeometry and ecomorphology applied to the masticatory apparatus. In: Ameghiniana. Band 45, Nr. 1, 2008, S. 175–196.
- M. Susana Bargo, Gerardo De Iuliis, Sergio F. Vizcaíno: Hypsodonty in Pleistocene ground sloths. In: Acta Palaeontologica Polonica. Band 51, Nr. 1, 2006, S. 53–61.
- Lucas Kraglievich: Contribución al conocimiento de “Mylodon darwini” Owen y especies afines. In: Revista del Museo de La Plata. Band 34, 1934, S. 255–292.
- Robert K. McAfee: Description of New Postcranial Elements of Mylodon darwinii Owen 1839 (Mammalia: Pilosa: Mylodontinae), and Functional Morphology of the Forelimb. In: Ameghiniana. Band 53, Nr. 4, 2016, S. 418–443.
- José A. Haro, Adan A. Tauber, Jerónimo M. Krapovickas: Thoracic member (pectoral girdle and forelimb) bones of Mylodon darwinii Owen (Xenarthra, Mylodontidae) from the Late Pleistocene of Central Argentina and their phylogenetic implications. In: Paläontologische Zeitschrift. Band 91, 2017, S. 439–457, doi:10.1007/s12542-017-0350-z.
- José A. Haro, Adan A. Tauber, Jerónimo M. Krapovickas: The Manus of Mylodon darwinii Owen (Tardigrada, Mylodontidae) and Ist Phylogenetic Implications. In: Journal of Vertebrate Paleontology. Band 36, Nr. 5, 2016, S. e1188824, doi:10.1080/02724634.2016.1188824.
- Francesco P. Moreno, Arthur Smith Woodward: On a Portion of Mammalian Skin, named Neomylodon listai, from a Cave near Consuelo Cove, Last Hope Inlet, Patagonia. In: Proceedings of the Zoological Society. 1899, S. 144–156 (biodiversitylibrary.org).
- Otto Nordenskjöld (unter Beteiligung weiterer Autoren): Wissenschaftliche Ergebnisse der Schwedischen Expedition nach den Magellansländern 1895–1897, unter Leitung von Dr. Otto Nordenskjöld.Band II: Zoologie. Stockholm, 1899, S. 1–170 (besonders S. 149–170) (biodiversitylibrary.org).
- Einar Lönnberg: On a remarkable piece of skin from Cueva Eberhardt, Last Hope Inlet, Patagonia. In: Proceedings of the Zoological Society. 1900, S. 379–383 (biodiversitylibrary.org).
- Arthur Smith Woodward: On some remains of Grypotherium (Neomylodon) listai and associated mammals from a cavern near Consuelo Cove, Patagonia. In: Proceedings of the Zoological Society. 1900, S. 64–79 (biodiversitylibrary.org).
- W. O. Ridewood: On the Structure of the Hairs of Mylodon Listai and other South American Edentata. In: Quaternary Review of Microscopic Science. Band 44, 1901, S. 393–411.
- Hermann Burmeister: Hautpanzer bei Mylodon. In: Archiv für Anatomie, Physiologie und wissenschaftliche Medicin. 1865, S. 334–336. (biodiversitylibrary.org).
- Hermann Burmeister: Fauna Argentina. Primera Parte. Mamiferos fósiles. Lista de los mamiferos fósiles del terreno diluviano. In: Anales del Museo Público de Buenos Aires. Band 1, 1867, S. 87–300 (S. 173) (biodiversitylibrary.org).
- Patricio López-Mendoza, Francisco Mena-Larraín: Extinct ground sloth dermal bones and their role in the taphonomic research of caves: the case of Baño Nuevo-1 (Andean Central Patagonia, Chile). In: Revista Mexicana de Ciencias Geológicas. Band 28, 2011, S. 519–532.
- Wilhelm Branco: Die Anwendung der Röntgenstrahlen in der Paläontologie. Abhandlungen der Königlich Preussischen Akademie der Wissenschaften Berlin 1906, S. 1–55 (biodiversitylibrary.org).
- Robert V. Hill: Comparative Anatomy and Histology of Xenarthran Osteoderms. In: Journal of Morphology. Band 267, 2005, S. 1441–1460.
- H. Gregory McDonald: An Overview of the Presence of Osteoderms in Sloths: Implications for Osteoderms as a Plesiomorphic Character of the Xenarthra. In: Journal of Mammalogy. Band 25, 2018, S. 485–493, doi:10.1007/s10914-017-9415-8.
- Claudio Latorre: Paleontología de mamíferos del alero Tres Arroyos 1, Tierra del Fuego. In: Anales del Instituto de la Patagonia. Band 26, 1998, S. 77–90.
- H. Gregory McDonald, Gerardo de Iuliis: Fossil history of sloths. In: Sergio F. Vizcaíno, W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 39–55.
- Larry G. Marshall, Annalisa Berta, Robert Hoffstetter, Rosendo Pascual', Osvaldo A. Reig, Miguel Bombin, Alvaro Mones: Mammals and stratigraphy: geochronology of the continental mammal-bearing Quaternary of South America. In: Palaeovertebrata Mém Ext. 1984, S. 1–76.
- Larry G. Marshall, Thierry Sempere: The Eocene to Pleistocene vertebrates of Bolivia and their stratigraphic context: a review. In: Revista Técnica de Yacimientos Petrolíferos Fiscales Bolivianos. Band 12, 1991, S. 631–652.
- Emmanuel Favotti, Brenda S. Ferrero, Diego Brandoni: Primer registro de Mylodon darwini Owen (Xenarthra, Tardigrada, Mylodontidae) en la Formación Arroyo Feliciano (Pleistoceno Tardío), Entre Ríos, Argentina. In: Revista Brasileira de Paleontologia. Band 18, Nr. 3, 2015, S. 547–554.
- Gustavo J. Scillato-Yané, Alfredo A. Carlini, Sergio F. Vizcaíno, Edgardo Ortiz Jaureguizar: Los Xenartros. María Teresa Alberdo, Gabriello Leone und Eduardo P. Tonni (Hrsg.): Evolución Biológica y Climática de la Región Pampeana durante los últimos cincos milliones de años. Madrid 1995, S. 183–175.
- Alfredo A. Carlini, Gustavo J. Scillato-Yané: Evolution of Quaternary Xenarthrans (Mammalia) of Argentina. In: Jorge Rabassa, Mónica Salemme (Hrsg.): Quaternary of South America and Antarctic Peninsula. Rotterdam 1999, S. 149–175.
- Luciano Brambilla, Damián Alberto Ibarra: Archaeomylodon sampedrinensis, gen. et sp. nov., a new mylodontine from the middle Pleistocene of Pampean Region, Argentina. In: Journal of Vertebrate Paleontology. Band 38, Nr. 6, 2018, S. e1542308, doi:10.1080/02724634.2018.1542308.
- Gustavo Martínez, María Gutiérrez, José Luis Prado: New Archaeological Evidences from the Late-Pleistocene/Early-Holocene Paso Otero 5 Site (Pampean Region, Argentina). In: Current Research in the Pleistocene. Band 21, 2004, S. 16–18.
- Gustavo Martínez: Arqueología del curso medio del Río Quequén Grande: Estado actual y aportes a la arqueología de la región Pampeana. In: Relaciones de la Sociedad Argentina de Antropología. Band 31, 2006, S. 249–275.
- Gustavo Martínez, María A. Gutiérrez: Paso Otero 5: a summary of the interdisciplinary lines of evidence for reconstructing early human occupation and paleoenvironment in the Pampean region, Argentina. In: D. Vialou (Hrsg.): Peuplements et Préhistoire de l’Amérique. Muséum National d’Histoire Naturelle. Département de Préhistoire, Paris 2011, S. 271–284.
- Richard A. Fariña, P. Sebastián Tambusso, Luciano Varela, Ada Czerwonogora, Mariana Di Giacomo, Marcos Musso, Roberto Bracco, Andrés Gascue: Arroyo del Vizcaíno, Uruguay: a fossil-rich 30-ka-old megafaunal locality with cut-marked bones. In: Proceedings of the Royal Society B. Band 281, 2014, S. 20132211, doi:10.1098/rspb.2013.2211.
- Édison Vicente Oliveira: Mammiferos Xenarthra (Edentata) do Quaternario do estado do Rio Grande do Sul, Brasil. In: Ameghiniana. Band 33, Nr. 1, 1996, S. 65–75.
- Jamil Corrêa Pereira, Renato Pereira Lopes, Leonardo Kerber: New remains of Late Pleistocene mammals from the Chuí Creek, Southern Brazil. In: Revista Brasileira de Paleontologia. Band 15, Nr. 2, 2012, S. 228–239.
- Luciano Varela, Richard A. Fariña: Co-occurrence of mylodontid sloths and insights on their potential distributions during the late Pleistocene. In: Quaternary Research. Band 85, 2016, S. 66–74.
- Natalia A. Villavicencio, Emily L. Lindsey, Fabiana M. Martin, Luis A. Borrero, Patricio I. Moreno, Charles R. Marshall, Anthony D. Barnosky: Combination of humans, climate, and vegetation change triggered Late Quaternary megafauna extinction in the Última Esperanza region, southern Patagonia, Chile. In: Ecography. Band 39, 2016, S. 125–140, doi:10.1111/ecog.01606.
- Jessica L. Metcalf, Chris Turney, Ross Barnett, Fabiana Martin, Sarah C. Bray, Julia T. Vilstrup, Ludovic Orlando, Rodolfo Salas-Gismondi, Daniel Loponte, Matías Medina, Mariana De Nigris, Teresa Civalero, Pablo Marcelo Fernández, Alejandra Gasco, Victor Duran, Kevin L. Seymour, Clara Otaola, Adolfo Gil, Rafael Paunero, Francisco J. Prevosti, Corey J. A. Bradshaw, Jane C. Wheeler, Luis Borrero, Jeremy J. Austin, Alan Cooper: Synergistic roles of climate warming and human occupation in Patagonian megafaunal extinctions during the Last Deglaciation. In: Science Advances. Band 2, 2016, S. e1501682, doi:10.1126/sciadv.1501682.
- Donald Jackson S.: Evaluating evidence of cultural associations of Mylodon in the semiarid region of Chile. In: L. Miotti, M. Salemme, M. Flegenheimer (Hrsg.): Where the south winds blow: ancient evidence of Paleo South Americans. Texas A&M University, 2003, S. 77–81.
- Luis Alberto Borrero: The Elusive Evidence: The Archeological Record of the South American Extinct Megafauna. In: G. Haynes (Hrsg.): American Megafaunal Extinctions at the End of the Pleistocene. Springer Science + Business Media B V, 2009, S. 145–168.
- Luis Alberto Borrero and Fabiana María Martin: Ground sloths and humans in southern Fuego-Patagonia: taphonomy and archaeology. In: World Archaeology. Band 44, Nr. 1, 2012, S. 102–117, doi:10.1080/00438243.2012.646145.
- Fabiana M. Martin, Dominique Todisco, Joel Rodet, Manuel San Román, Flavia Morello, Francisco Prevosti, Charles Stern, Luis A. Borrero: Nuevas excavationes en Cueva des Medio. Process de formación de la cuevay avances en los estudios de interacción entre cazadores recolectores y fauna extinta (Pleistoceno Final, Patagonia Meridional). In: Magellania (Chile). Band 43, Nr. 1, 2015, S. 165–189.
- Hugo G. Nami, Calvin J. Heusser: Cueva del Medio: A Paleoindian Site and Its Environmental Setting in Southern South America. In: Archaeological Discovery. Band 3, 2015, S. 62–71.
- Fabiana Martin, Manuel San Román, Flavia Morello, Dominique Todisco, Francisco J. Prevosti, Luis A. Borrero: Land of the ground sloths: Recent research at Cueva Chica, Ultima Esperanza, Chile. In: Quaternary International. Band 305, 2013, S. 56–66.
- Eduardo P. Tonni, Alfredo A. Carlini, Gustavo J. Scillato-Yané, Anibal J. Figini: Cronología radiocarbónica y condiciones climáticas en la “Cueva del Milodón” (sur de Chile) durante el Pleistoceno Tardío. In: Ameghiniana. Band 40, Nr. 4, 2003, S. 609–615.
- Anthony D. Barnosky, Emily L. Lindsey: Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. In: Quaternary International. Band 217, 2010, S. 10–29.
- Luis Alberto Borrero, Fabiana María Martin: Taphonomic observations on ground sloth bone and dung from Cueva del Milodón, Ultima Esperanza, Chile: 100 years of research history. In: Quaternary International. Band 278, 2012, S. 3–11.
- Mateo Martinic: La Cueva del Milodon (Ultima Esperanza, Patagonia chilena). Un siglo de descubrimientos y estudios referidos a la vida primitiva en el sur de America. In: Journal de la Société des Américanistes. Band 82, 1996, S. 311–323.
- Luis Alberto Borrero: The extinction of the megafauna: An supra-regional approach. In: Anthropozoologica. Band 25/26, 1997, S. 209–216.
- David W. Steadman, Paul S. Martin, Ross D. E. MacPhee, A. J. T. Jull, H. Gregory McDonald, Charles A. Woods, Manuel Iturralde-Vinent, Gregory W. L. Hodgins: Asynchronous extinction of late Quaternary sloths on continents and islands. In: PNAS. Band 102, 2005, S. 1763–11768.
- M. Susana Bargo, Néstor Toledo, Sergio F. Vizcaíno: Muzzle of South American Pleistocene Ground Sloths (Xenarthra, Tardigrada). In: Journal of Morphology. Band 267, 2006, S. 248–263.
- Sergio F. Vizcaíno, M. Susana Bargo, Guillermo H. Cassini: Dental occlusal surface area in relation to body mass, food habits and other biological features in fossil xenarthrans. In: Ameghiniana. Band 43, Nr. 1, 2006, S. 11–26.
- Sergio F. Vizcaíno: The teeth of the “toothless”: novelties and key innovations in the evolution of xenarthrans (Mammalia, Xenarthra). In: Paleobiology. Band 35, Nr. 3, 2009, S. 343–366.
- Vera Markgraf: Late Pleistocene Faunal Extinctions in Southern Patagonia. In: Science. Band 228, 1985, S. 1110–409.
- Calvin J. Heusser, Luis A. Borrero, José A. Lanata: Late Glacial vegetation at Cueva del Mylodon. In: Anales del Instituto de la Patagonia (Serie Ciencias Naturales). Band 21, 1992, S. 97–102.
- Rodrigo Villa-Martínez, Patricio I. Moreno: Pollen evidence for variations in the southern margin of the westerly winds in SW Patagonia over the last 12,600 years. In: Quaternary Research. Band 68, 2007, S. 400–409.
- Julia V. Tejada, John J. Flynn, Ross MacPhee, Tamsin C. O’Connell, Thure E. Cerling, Lizette Bermudez, Carmen Capuñay, Natalie Wallsgrove, Brian N. Popp: Isotope data from amino acids indicate Darwin’s ground sloth was not an herbivore. In: Scientific Reports. Band 11, 2021, S. 18944, doi:10.1038/s41598-021-97996-9.
- Gerardo De Iuliis, Guillermo H. Ré, Sergio F. Vizcaíno: The Toro Negro megatheriine (Mammalia, Xenarthra): A new species of Pyramiodontherium and a review of Plesiomegatherium. In: Journal of Vertebrate Paleontology. Band 24, Nr. 1, 2004, S. 214–227.
- M. Susana Bargo, Sergio F. Vizcaíno, Fernando M. Archuby, R. Ernesto Blanco: Limb bone proportions, strength and digging in some Lujanian (Late Pleistocene-Early Holocene) mylodontid ground sloths (Mammalia, Xenarthra). In: Journal of Vertebrate Paleontology. Band 20, Nr. 3, 2000, S. 601–610.
- Brian K. McNab: Energetics, population biology, and distribution of Xenarthrans, living and extinct. In: G. Gene Montgomery (Hrsg.): The evolution and ecology of armadillos, sloths, and vermilinguas. Smithonian Institution Press, 1985, S. 219–232.
- Luis A. Borrero, Fabiana M. Martin, Alfredo Prieto: La Cueva Lago Sofía 4, Ultima Esperanza, Chile: U madriguera felina del Pleistoceno tardío. In: Anales del Instituto de la Patagonia. Band 25, 1997, S. 103–122.
- Fabiana M. Martin: Bone crunching felids at the end of the Pleistocene in Fuego-Patagonia, Chile. In: Journal of Taphonomy. Band 6, Nr. 3/4, 2008, S. 337–372.
- Aldo Manzuetti, Daniel Perea, Washington Jones, Martín Ubilla, Andrés Rinderknecht: An extremely large saber-tooth cat skull from Uruguay (late Pleistocene–early Holocene, Dolores Formation): body size and paleobiological implications. In: An Australasian Journal of Palaeontology, Alcheringa. Band 44, Nr. 2, 2020, S. 332–339, doi:10.1080/03115518.2019.1701080.
- Francisco J. Prevosti, Fabiana M. Martin: Paleoecology of the mammalian predator guild of Southern Patagonia during the latest Pleistocene: Ecomorphology, stable isotopes, and taphonomy. In: Quaternary International. Band 305, 2013, S. 74–84.
- Raúl A. Ringuelet: Restos de probables huevos de nematodes en el estiercol del edentado extinguido Mylodon listai (Ameghino). In: Ameghiniana. 1 (1/2); 1957, S. 15–16.
- Alberto Boscaini, François Pujos, Timothy J. Gaudin: A reappraisal of the phylogeny of Mylodontidae (Mammalia, Xenarthra) and the divergence of mylodontine and lestodontine sloths. In: Zoologica Scripta. Band 48, Nr. 6, 2019, S. 691–710, doi:10.1111/zsc.12376.
- Luciano Varela, P. Sebastián Tambusso, H. Gregory McDonald, Richard A. Fariña: Phylogeny, Macroevolutionary Trends and Historical Biogeography of Sloths: Insights From a Bayesian Morphological Clock Analysis. In: Systematic Biology. Band 68, Nr. 2, 2019, S. 204–218.
- H. Gregory McDonald: Evolution of the Pedolateral Foot in Ground Sloths: Patterns of Change in the Astragalus. In: Journal of Mammalian Evolution. Band 19, 2012, S. 209–215.
- Bruce J. Shockey, Federico Anaya: Grazing in a New Late Oligocene Mylodontid Sloth and a Mylodontid Radiation as a Component of the Eocene-Oligocene Faunal Turnover and the Early Spread of Grasslands/Savannas in South America. In: Journal of Mammalian Evolution. Band 18, 2011, S. 101–115.
- Malcolm C. McKenna, Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York 1997, S. 1–631 (S. 94–96).
- Andrés Rinderknecht, Enrique Bostelmann T., Daniel Perea, Gustavo Lecuona: A New Genus and Species of Mylodontidae (Mammalia: Xenarthra) from the Late Miocene of Southern Uruguay, with Comments on the Systematics of the Mylodontinae. In: Journal of Vertebrate Paleontology. Band 30, Nr. 3, 2010, S. 899–910.
- Francisco Ricardo Negri, Jorge Ferigolo: Urumacotheriinae, nova subfamília de Mylodontidae (Mammalia, Tardigrada) do Mioceno Superior-Plioceno, América do Sul. In: Revista Brasileira de Paleontologia. Band 7, Nr. 2, 2004, S. 281–288.
- Ascanio D. Rincón, H. Gregory McDonald, Andrés Solórzano, Mónica Núñez Flores, Damián Ruiz-Ramoni: A new enigmatic Late Miocene mylodontoid sloth from northern South America. In: Royal Society Open Science. Band 2, 2015, S. 140256, doi:10.1098/rsos.140256.
- Timothy J. Gaudrin: Phylogenetic relationships among sloths (Mammalia, Xenarthra, Tardigrada): the craniodental evidence. In: Zoological Journal of the Linnean Society. Band 140, 2004, S. 255–305.
- Ascanio D. Rincón, Andrés Solórzano, H. Gregory McDonald, Mónica Núñez Flores: Baraguatherium takumara, Gen. et Sp. Nov., the Earliest Mylodontoid Sloth (Early Miocene) from Northern South America. In: Journal of Mammalian Evolution. Band 24, Nr. 2, 2017, S. 179–191.
- Samantha Presslee, Graham J. Slater, François Pujos, Analía M. Forasiepi, Roman Fischer, Kelly Molloy, Meaghan Mackie, Jesper V. Olsen, Alejandro Kramarz, Matías Taglioretti, Fernando Scaglia, Maximiliano Lezcano, José Luis Lanata, John Southon, Robert Feranec, Jonathan Bloch, Adam Hajduk, Fabiana M. Martin, Rodolfo Salas Gismondi, Marcelo Reguero, Christian de Muizon, Alex Greenwood, Brian T. Chait, Kirsty Penkman, Matthew Collins, Ross D. E. MacPhee: Palaeoproteomics resolves sloth relationships. In: Nature Ecology & Evolution. Band 3, 2019, S. 1121–1130, doi:10.1038/s41559-019-0909-z.
- Matthias Höss, Amrei Dilling, Andrew Currant, Svante Pääbo: Molecular phylogeny of the extinct ground sloth Mylodon darwinii. In: Proceedings of the National Academy of Sciences. Band 93, 1996, S. 181–185.
- Alex D. Greenwood, Jose Castresana, Gertraud Feldmaier-Fuchs, Svante Pääbo: A Molecular Phylogeny of Two Extinct Sloths. In: Molecular Phylogenetics and Evolution. Band 18, Nr. 1, 2001, S. 94–103.
- Michael Hofreiter, Julio L. Betancourt, Alicia Pelliza Sbriller, Vera Markgraf, H. Gregory McDonald: Phylogeny, diet, and habitat of an extinct ground sloth from Cuchillo Curá, Neuquén Province, southwest Argentina. In: Quaternary Research. Band 59, 2003, S. 364–378.
- Hendrik Poinar, Melanie Kuch, Gregory McDonald, Paul Martin, Svante Pääbo: Nuclear Gene Sequences from a Late Pleistocene Sloth Coprolite. In: Current Biology. Band 13, 2003, S. 1150–1152.
- Andrew A. Clack, Ross D. E. MacPhee, Hendrik N. Poinar: Mylodon darwinii DNA sequences from ancient fecal hair shafts. In: Annals of Anatomy. Band 194, 2012, S. 26–30.
- Graham J. Slater, Pin Cui, Analía M. Forasiepi, Dorina Lenz, Kyriakos Tsangaras, Bryson Voirin, Nadia de Moraes-Barros, Ross D. E. MacPhee, Alex D. Greenwood: Evolutionary Relationships among Extinct and Extant Sloths: The Evidence of Mitogenomes and Retroviruses. In: Genome Biology and Evolution. Band 8, Nr. 3, 2016, S. 607–621.
- Michael Buckley, Richard A. Fariña, Craig Lawless, P. Sebastián Tambusso, Luciano Varela, Alfredo A. Carlini, Jaime E. Powell, Jorge G. Martinez: Collagen Sequence Analysis of the Extinct Giant Ground Sloths Lestodon and Megatherium. In: PLoS ONE. Band 10, Nr. 11, 2015, S. e0139611, doi:10.1371/journal.pone.0139611.
- Frédéric Delsuc, Melanie Kuch, Gillian C. Gibb, Jonathan Hughes, Paul Szpak, John Southon, Jacob Enk, Ana T. Duggan, Hendrik N. Poinar: Resolving the phylogenetic position of Darwin’s extinct ground sloth (Mylodon darwinii) using mitogenomic and nuclear exon data. In: Proceedings of the Royal Society B. Band 285, 2018, S. 20180214, doi:10.1098/rspb.2018.0214.
- Frédéric Delsuc, Melanie Kuch, Gillian C. Gibb, Emil Karpinski, Dirk Hackenberger, Paul Szpak, Jorge G. Martínez, Jim I. Mead, H. Gregory McDonald, Ross D. E. MacPhee, Guillaume Billet, Lionel Hautier, Hendrik N. Poinar: Ancient mitogenomes reveal the evolutionary history and biogeography of sloths. In: Current Biology. Band 29, Nr. 12, 2019, S. 2031–2042, doi:10.1016/j.cub.2019.05.043.
- Lucas Kraglievich: “Mylodon darwini” Owen es la especie genotipo de “Mylodon” Ow. Rectificacíon de la nomenclatura genérica de los Milodontes. In: Physis. Band 9, 1928, S. 169–185.
- Henri Gervais, Florentino Ameghino: Les mammifères fossiles de l´Ámérique du Sud. Paris-Buenos Aires 1880, S. 1–225 (S. 160–161) (archive.org).
- Adrian P. Hunt, Spencer George Lucas: The record of sloth coprolites in north and South America: Implications for terminal Pleistocene extinctions. In: New Mexico Museum of Natural History and Science Bulletin. Band 79, 2018, S. 277–298.
- Gerardo De Iuliis, Cástor Cartelle, H. Gregory McDonald, François Pujos: The mylodontine ground sloth Glossotherium tropicorum from the late Pleistocene of Ecuador and Peru. In: Papers in Palaeontology. Band 3, Nr. 4, 2017, S. 613–636.
- Richard Harlan: Description of the jaws, teeth, and clavicle of the Megalonyx laqueatus. In: The Monthly American journal of geology and natural science. Band 1, 1831, S. 74–76 (biodiversitylibrary.org).
- Richard Owen: Fossil Mammalia. In: Charles Darwin (Hrsg.): Zoology of the Voyage of H.M.S Beagle, under the command of Captain Fitzroy, during the years 1832 to 1836. Part I. Fossil Mammals. London, 1840, S. 12–111 (S. 63–73) (biodiversitylibrary.org).
- Richard Harlan: Notice of two new mammals from Brunswick Cana, Georgia; with observations on the fossil quadrupeds of the United States. In: The American journal of science and arts. Band 43, 1842, S. 141–144 (biodiversitylibrary.org).
- Juan Carlos Fernicola, Sergio F. Vizcaíno, Gerardo De Iuliis: The fossil mammals collected by Charles Darwin in South America during his travels on board the HMS Beagle. In: Revista de la Asociación Geológica Argentina. Band 64, Nr. 1, 2009, S. 147–159.
- Sergio F. Vizcaíno, Richard A. Fariña, Juan Carlos Fernicola: Young Darwin and the ecology and extinction of Pleistocene South American fossil mammals. In: Revista de la Asociación Geológica Argentina. Band 64, Nr. 1, 2009, S. 160–169.
- Richard Owen: Description of the skeleton of an extinct gigantic Sloth, Mylodon robustus, Owen, with observations on the osteology, natural affinities, and probable habitats of the Megatherioid quadrupeds in general. London, 1842, S. 1–176 (speziell S. 154) (archive.org).
- Johannes Theodor Reinhardt: Beskrivelse af Hovedskallen af et Kæmpedovendyr, Grypotherium darwini. In: Det Kongelige Danske Videnskabernes Selskabs Skrifter. 5 Række. Naturvidenskabelig og Mathematisk Afdeling 12 (5), 1879, S. 351–381 (archive.org).
- Florentino Ameghino: Contribución al conocimiento de los mamíferos fósiles de la República Argentina. In: Actas de la Academia Nacional de Ciencias. Band 6, 1889, S. 1–1027 (S. 734–744).
- H. Gregory McDonald: Gravigrade xenarthrans from the early Pleistocene Leisey Shell Pit 1A, Hillsborough County, Florida. In: Bulletin of the Florida Museum of Natural History. Band 37, 1995, S. 345–373 (ufdcweb1.uflib.ufl.edu).
- Barnum Brown: A new genus of ground sloth from the Pleistocene of Nebraska. In: Bulletin of the American Museum of Natural History. Band 29, 1903, S. 569–583.
- Chester Stock: Further observation on the skull structure of the mylodont sloths from Rancho La Brea. In: University of California Publications, Bulletin of the Department of Geology. Band 10, 1917, S. 165–178.
- George Gaylord Simpson: The Principles of Classification and a Classification of Mammals. In: Bulletin of the American Museum of Natural History. Band 85, 1945, S. 1–350 (S. 71).
- Joseph Leidy: A memoir on the extinct Sloth Tribe of North America. In: Smithsonian Contribution to Knowledge. Band 7, 1855, S. 1–68 (S. 58) (reader.digitale-sammlungen.de).
- Richard Lydekker: Catalogue of the fossil Mammalia in the British Museum of Natural History. Part V. London 1887, S. 1–345 (S. 106) (biodiversitylibrary.org).
- Rodolfo Hauthal, Santiago Roth, Robert Lehmann-Nitsche: El Mamifero Misterioso de la Patagonia "Grypotherium domesticum". In: Revista del Museo de La Plata. Band 9, 1899, S. 409–473 (biodiversitylibrary.org).
- Erland Nordenskjöld: Neue Untersuchungen über Neomylodon listai. In: Zoologischer Anzeiger. Band 22, 1899, S. 335–336 (biodiversitylibrary.org).
- Erland Nordenskjöld: Jakttagelser och fynd i Grottor vid Ultima Esperanza i sydvestra Patagonien. In: Kongliga Svenska Vetenskaps Akademiens Handligar. Band 33, Nr. 3, 1900, S. 1–24 (biodiversitylibrary.org).
- Florentino Ameghino: An existing ground-sloth in Patagonia. In: Natural Science. Band 13, 1898, S. 324–326 (biodiversitylibrary.org).
- Anonymous: A living representative of the old ground sloths. In: Nature. Band 58, 1898, S. 547–548 (nature.com).
- Francesco P. Moreno: Note on the discovery of Miolania and of Glossotherium (Neomylodon) in Patagonia. In: Geological Magazine. Band 6, Nr. 9, 1899, S. 385–388 (biodiversitylibrary.org).
- Hesketh Vernon Hesketh-Prichard: Through the heart of Patagonia. New York 1902, S. 1–346 (S. 332) (biodiversitylibrary.org).
- Bruce Chatwin: In Patagonia. Vintage, London 2005, S. 1–260 (speziell S. 242–245).
Weblinks
- Natural History Museum: Mylodon darwinii: Darwins ground sloth, zuletzt abgerufen am 27. März 2020