Großer Ameisenbär
Der Große Ameisenbär (Myrmecophaga tridactyla) ist ein Säugetier aus der Ordnung der Zahnarmen. Gemeinsam mit den Tamanduas und den Zwergameisenbären bildet er die Unterordnung der Ameisenbären (Vermilingua). Er ist deren größter Vertreter und kommt in Süd- und Mittelamerika vor, sein ursprüngliches Verbreitungsgebiet war aber wesentlich größer. Hauptsächlich ist der Große Ameisenbär bodenbewohnend und einzelgängerisch, wobei er recht große Territorien unterhält. Die Hauptnahrung besteht aus Ameisen und Termiten. Weibliche Tiere bringen pro Wurf ein Jungtier zur Welt, das bis zu einem Jahr gesäugt wird. Der Lebensraum umfasst verschiedene Landschaftstypen von Gras- und Savannenländern bis Wäldern. Aufgrund von Landschaftszerstörung und Bejagung gilt der Bestand des Großen Ameisenbären als gefährdet.
| Großer Ameisenbär | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
.jpg.webp)
Großer Ameisenbär (Myrmecophaga tridactyla) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Myrmecophaga | ||||||||||||
| Linnaeus, 1758 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Myrmecophaga tridactyla | ||||||||||||
| Linnaeus, 1758 |
Beschreibung
Habitus

Der Große Ameisenbär ist der weitaus größte Vertreter der Ameisenbären. Er erreicht eine Kopf-Rumpf-Länge von 100 bis 140 cm, der buschige Schwanz wird nochmals 60 bis 90 Zentimeter lang. Das Gewicht wildlebender Tiere beträgt 22 bis 39 kg,[1] es liegen allerdings auch Angaben bis 52 kg vor.[2] Männliche Tiere sind durchschnittlich etwas größer als weibliche. Auffälligstes Kennzeichen ist die lange, dünne und rund geformte Schnauze, die eine stark verlängerte, bis zu 60 cm messende und mit klebrigem Speichel bedeckte Zunge beherbergt, und zudem typisch für Ameisenbären zahnlos ist. Die Augen sind braun und sehr klein, ebenso die Ohren, die eine rundliche Gestalt aufweisen und 3,5 bis 6 cm lang werden, wobei sie nicht über den Kopf hinausragen. An der Basis des Nackens erhebt sich ein leichter Buckel, über den Halsrücken hinweg zieht sich eine Reihe borstiger, langer Haare, die eine Art Mähne bilden. Die Grundfärbung des langhaarigen Fells ist graubraun, an Brust und Schultern hat der Große Ameisenbär eine schwarze Zeichnung mit weißer streifenartiger Einrahmung. Die Vorderbeine sind weiß mit einem schwarzen Knöchelband, die Hinterbeine und der buschige Schwanz, der bis zu 30 cm lange Haare besitzt, weisen eine dunklere Tönung als der Körper auf. Die einzelnen Tiere besitzen eine jeweils charakteristische Fellzeichnung, die stärker bis schwächer ausgebildete Farbbänder oder deutliche beziehungsweise weniger markante Flecken einschließt, wodurch eine individuelle Unterscheidung möglich ist.[3] Die Vorderbeine sind äußerst kräftig ausgebildet, die Vorderfüße haben fünf Zehen, von denen die drei mittleren stark verlängert und mit großen, sichelförmig gestalteten, 10 bis 15 cm langen Krallen versehen sind (daher der Artzusatz tridactylus für „dreizehig“). Die Hinterbeine enden in fünf Zehen mit jeweils kurzen Krallen, die Hinterfußlänge beträgt 15 bis 18 cm. Weibchen besitzen ein Paar Zitzen im Brustbereich.[2][1][4]
Schädel- und Skelettmerkmale
.jpg.webp)
Der Schädel wird insgesamt 21 cm lang und ist allgemein sehr schmal und langgestreckt, in Seitenansicht verläuft die Stirnlinie deutlich gewölbt. Markant ist das röhrenförmige Rostrum, welches sehr ausgedehnt ist und rund 65 % der gesamten Schädellänge einnimmt. Dadurch erreicht das Nasenbein in etwa die gleiche Länge wie das Stirnbein. Der Hirnschädel wiederum ist vergleichsweise breit. Des Weiteren zeigt sich der Jochbogen nur rudimentär entwickelt. Der Unterkiefer ist sehr niedrig, die beiden Hälften sind an der Symphyse nur lose miteinander verbunden. Der niedrige Unterkieferkörper und der reduzierte Jochbogen ermöglichen es dem Großen Ameisenbären, seitliche und rotierende Kaubewegungen auszuüben und zudem die Zunge zu kontrollieren, wenn diese ausgestreckt oder zurückgezogen wird. Zudem sind diese Bewegungen des Unterkiefers, die hauptsächlich durch den Musculus pterygoideus medialis und den Musculus temporalis hervorgerufen werden, eine Anpassung an die Ernährungsweise.[5] Typischerweise treten 16 Brust- und nur zwei Lendenwirbel auf. Die xenarthrischen Gelenke (Nebengelenke) treten meist an den drei bis vier hintersten Brust- und den beiden Lendenwirbeln auf. Dabei sind beim Großen Ameisenbären zwei zusätzliche Nebengelenke ausgebildet, was die Wirbelsäule im hinteren Bereich verstärkt und möglicherweise zum Halten des Gewichtes beim Aufrichten des Körpers während des Fressens oder der Verteidigung dient.[6] Die Rippen besitzen an ihren hinteren Enden starke Verbreiterungen. Zusätzlich weisen vier oder fünf der mittleren Rippen deutliche Verlängerungen auf.[2][1]
Sinnesleistungen und Lautäußerungen
Der Sehsinn des Großen Ameisenbären ist nur rudimentär ausgebildet, der Geruchssinn dagegen sehr gut. Dieser wird vor allem bei der Nahrungssuche eingesetzt. Ein Tier ist generell leise, manchmal gibt es ein leises Grunzen von sich, vor allem wenn es gestört wird. Einen langanhaltenden harrrr-Laut äußern manche Tiere bei einem Angriff auf einen Artgenossen.[2]
Verbreitung
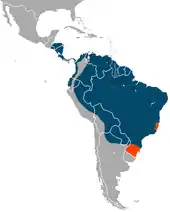
Der Große Ameisenbär ist in Mittel- und Südamerika beheimatet, sein Verbreitungsgebiet erstreckt sich vom südlichen Belize und Guatemala bis ins nördliche Argentinien. Möglicherweise ist er aber schon aus seinen nördlichsten Lebensräumen verschwunden, neuere Hinweise gibt es jedoch aus Honduras.[7][8][9] Auch im südlichen Teil Mittelamerikas, so im zentralen Panama, ist er teilweise schon verschwunden. In Südamerika tritt die Art überwiegend östlich der Anden auf, nur in Kolumbien und möglicherweise in Ecuador bewohnt sie auch die Tiefländer westlich der Anden, die Daten für Ecuador müssen aber erst noch bestätigt werden. Auch im Süden des Ausbreitungsgebietes gab es bereits deutliche Habitatverluste. In historischer Zeit war der Große Ameisenbär bis weit südlich des 31. Breitengrades der Südhalbkugel anzutreffen. Im nördlichen Uruguay, dem östlichen Paraguay und dem südöstlichen Brasilien ist er bereits ausgestorben. Die südlichste Verbreitungsgrenze stellt heute der nördliche Teil Argentiniens dar, wo sich die Ameisenbärenart in den letzten Jahren auch weiter nach Norden zurückgezogen hat.[10] Das gesamte heutige Verbreitungsgebiet wird mit 12,5 Millionen Quadratkilometern angegeben, das tatsächlich bewohnte Gebiet ist aber unbekannt.[2][11][4]
Der Große Ameisenbär bewohnt zahlreiche unterschiedliche Landschaften und kommt in Sümpfen, tropischen Regenwäldern, Trockenwäldern und am häufigsten in Savannen vor. So bewohnt er die Gran-Chaco-Regionen ebenso wie das Pantanal, die Cerrado-Gebiete[12] und das Amazonasbecken. In Zentralamerika gibt es mehrere Sichtungen aus Hochlandgebieten. Voraussetzung für die Anwesenheit des Großen Ameisenbären ist ein genügend großes Vorkommen von Ameisen und Termiten.[2][11] Bei Untersuchungen im nordöstlichen Argentinien zeigte sich, dass die Tiere in Landschaften mit gemischten Vegetationstypen häufig dichte Wälder oder Grasländer mit hohem Bewuchs gegenüber kurzgrasigen Savannen bevorzugen. Dies könnte einerseits mit dem Dämmeffekt bei extremen Temperaturen, andererseits mit besseren Versteckmöglichkeiten bei potentiellen Gefahren zusammenhängen. Generell meiden sie aber intensiv genutzte Weideflächen.[13] Die Dichte der Populationen ist abhängig von der Beschaffenheit der Landschaften. So liegt sie im Pantanal bei 0,15 Individuen je Quadratkilometer. Deutlich höher ist sie in Wäldern mit 0,41 Individuen, eher gering im Llanos mit 0,18 und in Überflutungsebenen mit 0,12 Individuen auf der gleichen Fläche. Für den Emas-Nationalpark wird eine Populationsdichte von 0,2 Individuen je Quadratkilometer angegeben.[14][4]
Lebensweise
Territorialverhalten

Als einziges Mitglied seiner Familie ist der Große Ameisenbär ein reiner Bodenbewohner. Normalerweise ist er tagaktiv, dies variiert aber mit der Jahreszeit, so dass ein Tier an heißen Tagen überwiegend in den kühlen Morgen- und Abendstunden auf Nahrungssuche geht, an kühlen Tagen aber eher tagsüber.[15] Regional besteht teilweise eine Abhängigkeit von der Aktivität der Beutetiere, dadurch ist der Große Ameisenbär etwa in den Llanos-Gebieten Venezuelas häufiger nachts unterwegs. Generell verschiebt sich seine Aktivität in stark von Menschen bewohnten Gebieten zur Nacht hin. Zudem ist der Große Ameisenbär einzelgängerisch und territorial. Die Reviere sind in der Regel zwischen 5 und 25 km² groß, können aber im Extremfall bis zu 90 km² erreichen. Die Größe ist dabei abhängig von der Beschaffenheit des Habitats. So wurde im Pantanal eine Größe von 5,7 km² für männliche und bis zu 11,9 km² für weibliche Tiere festgestellt. Ob Weibchen generell größere Territorien besitzen, ist unklar, da im Emas-Nationalpark im Cerrado eine durchschnittliche Reviergröße von 9,8 km² für beide Geschlechter nachgewiesen wurde.[16] Die Grenzen der einzelnen Territorien können sich dabei überschneiden, allerdings halten die Tiere eher Abstand zueinander. Überschneidungen der Reviergrenzen finden sich häufiger bei Weibchen als bei Männchen.[2][17][1][4]
Die Reviere werden häufig markiert. Diese Markierungen erfolgen in der Regel an den Stämmen größerer Bäume durch Kratzen mit den langen Krallen der Vorderfüße. Dies dient gleichzeitig auch dem Schärfen der Krallen; da die Kratzspuren aber weit unterhalb der Verästelung des Stammes angebracht werden und deutlich sichtbar sind, dienen diese Experten zufolge auch zur visuellen Kommunikation. Eher selten markiert der Große Ameisenbär mit Urin.[18] Innerhalb des Reviers bewegt sich die Ameisenbärenart am Boden auf allen vieren fort, wobei die Krallen der Vorderfüße nach unten geklappt sind, so dass die Zehen stark gebeugt sind und das Gewicht von den Unterflächen der drei größeren Zehen getragen wird.[19] Meist läuft sie dabei eher langsam, kann aber auch in einen schnellen Sprint übergehen. Täglich legt ein Tier zwischen 1,3 und 11 km zurück, durchschnittlich sind es 3,7 km. Zudem ist der Große Ameisenbär ein guter Schwimmer, im Amazonasgebiet wurde ein Tier beim Überqueren eines 150 m breiten Flusses beobachtet, wobei es abwechselnd schwamm und über die herausragenden Felsen kletterte.[20] Gelegentlich wird ein Tier beim Besteigen eines bis zu 2 m hohen Termitenhügels beobachtet, Jungtiere vermögen auch bis zu 20 m hoch in Bäumen zu klettern.[21] Begegnungen zwischen zwei oder mehreren Tieren verlaufen häufig sehr aggressiv, vor allem unter Männchen. Die Kämpfe, die teils ritualisiert sind, aber auch sehr blutig ausgehen können, werden dabei mit den Vorderfußkrallen ausgeführt.[22][23] Sehr selten wurden bisher Zusammentreffen verschiedener Individuen ohne Konfrontation beobachtet, etwa von mehreren Mutter- mit Jungtieren bei der Nahrungsaufnahme.[24] Zur Ruhe zieht sich ein Tier in flache, überwiegend selbst angelegte Erdkuhlen in dichter Vegetation zurück, tiefere Erdbauten oder Höhlen gräbt der Große Ameisenbär trotz seiner kräftigen Krallen und seines körperlichen Vermögens nicht. Die Schlafposition ist dabei seitlich liegend mit der Schnauze vor den Vorderfüßen, wobei der dichte Schwanz den Körper bedeckt. Die Position des Schwanzes dient dabei einerseits dazu, nicht zu viel Körperwärme zu verlieren, andererseits aber auch der Tarnung.[25][17][1][4]
Ernährung

Der Große Ameisenbär ist ein spezialisierter Insektenfresser, seine Hauptnahrung besteht aus Ameisen und Termiten, von denen er täglich bis zu 35.000 Tiere frisst, die er mittels seines guten Geruchssinnes aufspürt. Mit seinen starken Krallen bricht er Nester der Insekten, darunter auch sehr feste Termitenbaue. Dabei benutzt er eine „Haken-Zug-Methode“, indem er die Krallen in kleine Öffnungen steckt und die Vorderbeine kräftig zurückzieht. Die Insekten leckt der Große Ameisenbär mit seiner langen, bis zu 40 cm aus dem Maul ragenden Zunge auf, die Beute bleibt aufgrund des schleimhautartigen Belages kleben. Allerdings kann der Große Ameisenbär nur kurze Zeit am jeweiligen Bau fressen, bevor die Termiten und Ameisen ihre Körpergifte gezielt und organisiert einsetzen, so dass die durchschnittliche Fressdauer rund 40 Sekunden beträgt, bevor er ein neues Nest sucht und später zurückkehrt, um die Kolonie erneut auszubeuten. Durch dieses Verhalten werden die einzelnen Nester nie vollständig ausgeräumt. Die röhrenförmig ausgebildete Schnauze begrenzt darüber hinaus das weitere Nahrungsspektrum, so dass die Ameisenbärenart stark von koloniebildenden Insekten abhängig ist. Dabei treten regional deutliche Unterschiede auf, wobei die Ernährung im Extrem laut Untersuchungen an Kotresten im brasilianischen Bundesstaat Amapá zu 100 % auf Termiten, im Bundesstaat Ceará aber zu 100 % auf Ameisen basiert.[26] Im Pantanal werden hauptsächlich Ameisen konsumiert, so unter anderem Vertreter der Gattung Solenopsis, die rund 46 % der Gesamtmenge ausmachen, zu geringeren Anteilen aber auch Camponotus (12 %), Labidus (2 %) und Odontomachus (2 %). Termiten dagegen sind nur mit zwei Gattungen vertreten: Nasutitermes und Armitermes.[27] Weitere Analysen in Argentinien zeigen, dass der Große Ameisenbär dort häufig Termiten der Gattungen Nasutitermes und Cornitermes sowie Ameisengattungen wie Camponotus, Iridomyrmex oder Solenopsis bevorzugt, wobei ihr jeweiliger Anteil in der Nahrungsmenge jahreszeitlich schwankt.[2] Allein für die Region des Gran Chaco sind Vertreter von insgesamt 14 Ameisengattungen im Nahrungsspektrum belegt, darunter auch einige, die bevorzugt Bäume bewohnen.[28] Darüber hinaus erbrachten Untersuchungen an 23 Kotproben aus dem südbrasilianischen Bundesstaat Paraná in den Jahren 2007 und 2008 in fast allen Fällen auch den Nachweis von Resten von Blattschneideameisen der Gattungen Atta und Acromyrmex. Der regelmäßige Verzehr dieser Ameisen war aufgrund ihrer Aggressivität nicht angenommen worden, zumal Tiere in Gefangenschaft meist dieses Nahrungsangebot mieden.[29] Zuvor war der Verzehr von Blattschneideameisen nur aus Kolumbien bekannt gewesen.[30] Daneben wurde aber auch beobachtet, wie der Große Ameisenbär Nester von Honigbienen plündert, unter anderem in einem 1,5 m hohen, verlassenen Termitenbau im Emas-Nationalpark.[31] Der Große Ameisenbär trinkt regelmäßig Wasser und gräbt in trockenen Jahreszeiten auch in kleinen Löchern danach.[2][17][1][4]
Fortpflanzung

Die Fortpflanzung erfolgt ganzjährig, es gibt aber Hinweise auf einen enger begrenzten Zeitraum zwischen Mai und Juli. Es ist die einzige Phase, in der mehrere Tiere zusammenkommen. Der Sexualzyklus der Weibchen währt 47 bis 62 Tage und setzt etwa alle sieben Wochen ein.[32] Das Balzverhalten ist stark ritualisiert und umfasst gegenseitiges Umkreisen, das Zeigen der Krallen und Schläge mit den Vorderarmen, um den potentiellen Partner zu Boden zu bringen. Mitunter kann es auch zu Aggressionen kommen, vor allem wenn das Weibchen noch ein Jungtier von der vorhergehenden Geburt bei sich hat.[33][34] Nach etwa 183- bis 190-tägiger Tragzeit bringt das Muttertier ein einzelnes Junges zur Welt. In der Regel werden etwa doppelt so viele männliche wie weibliche Jungtiere geboren. Ein Neugeborenes wiegt 1,1 bis 1,6 kg, ist mit Schwanz etwa 53 cm lang und hat die Augen geschlossen; diese öffnen sich erst nach rund sechs Tagen. Häufig trägt das Muttertier das Junge, wenn es auf Nahrungssuche geht, was bis zu einem Alter von einem Jahr geschehen kann. Dabei liegt das Junge längs auf dem Rücken und ist so teils vom dichten Fell der Mutter verdeckt, andererseits verschmilzt seine Fellzeichnung mit der des Muttertieres. Am Boden ist das Jungtier in den ersten sechs Wochen nie weiter als einen Meter von der Mutter entfernt, der Abstand erhöht sich bis zum vierten Lebensmonat auf 20 Meter. In den ersten Monaten spielen Mutter und Junges häufig miteinander, was sich in Springen, Spielen mit Objekten oder den Krallen zeigt; auch gegenseitiges Lecken kommt häufig vor und kann bis zu eine Stunde andauern. Aufgrund der langen Schnauze saugt das Jungtier mit der Zunge, wobei es anfangs bis zu fast 20 % seiner aktiven Zeit mit der Aufnahme der Muttermilch verbringt. Ab dem dritten Lebensmonat nimmt es erstmals feste Nahrung zu sich, ab dem neunten Monat überwiegt dieser Nahrungsbestandteil. Die Mutter bringt bei der Nahrungsumstellung dem Nachwuchs bei, welche Nahrung verwertbar und welcher Geruchsspur zu folgen ist, was eines der wenigen Beispiele eines aktiven Anlernens innerhalb der Nebengelenktiere darstellt.[35] Die vollständige Entwöhnung erfolgt mit zehn bis zwölf Monaten, wobei der junge Ameisenbär dann rund 6,0 bis 6,5 kg wiegt und 105 cm lang ist. Die Geschlechtsreife erreichen die Tiere mit zwei bis vier Jahren. In menschlicher Obhut wurde ein Großer Ameisenbär knapp 26 Jahre alt, das dokumentierte Durchschnittsalter liegt jedoch bei etwa 16 Jahren. Die Lebenserwartung in freier Wildbahn ist nicht bekannt.[36][2][1][4]
Beutegreifer und Feindverhalten

Die größten Fressfeinde stellen Jaguar und Puma dar, allerdings wird der Große Ameisenbär nicht allzu häufig von diesen großen Katzen erbeutet. Untersuchungen im Gran Chaco an 106 Kotresten des Jaguars und 95 des Pumas ergaben lediglich zwei beziehungsweise drei Hinweise auf das Erlegen des Großen Ameisenbären. Während einer Beobachtung von 10 Jaguaren, je fünf männliche und weibliche Tiere, von Oktober 2001 bis April 2004 im südlichen Pantanal konnten insgesamt 14 erbeutete Große Ameisenbären festgestellt werden, was lediglich 3,2 % aller erlegten Tiere in diesem Zeitraum ausmachte.[37] Weiterhin ist die Ameisenbärenart dort seltener, wo häufiger Hunde vorkommen. Ein bedrohtes Tier richtet sich auf seinen Hinterbeinen auf und verteidigt sich durch Schläge mit den Krallen der Vorderfüße.[2] Prinzipiell ortet der Große Ameisenbär Beutegreifer über die Witterung, wobei er zwischen potentiellen Fressfeinden und sonstigen Raubtieren zu unterscheiden vermag.[38]
Parasiten
Zu den äußeren Parasiten zählen vor allem verschiedene Arten der Zeckengattung Amblyomma, auch Milben wie Otodectes oder Edentalges kommen vor, selten dagegen Flöhe.[39] Als innere Parasiten sind hauptsächlich Fadenwürmer bekannt, darunter Aspidodera, Brevigraphidium oder Physaloptera. Weiterhin wurden Bandwürmer nachgewiesen, unter anderem Mathevotaenia. Mit Eimeria konnten auch Kokzidien als Parasiten festgestellt werden.[2]
Systematik
Innere Systematik der rezenten Ameisenbären nach Delsuc et al. 2012[40]
|
Der Große Ameisenbär ist die rezent einzige Art aus der monotypischen Gattung Myrmecophaga. Sie gehört wiederum zur Familie der Myrmecophagidae, denen auch die Tamanduas (Tamandua) zugerechnet werden, innerhalb der Unterordnung der Ameisenbären (Vermilingua). Neben den Myrmecophagidae werden nur die Cycopedidae mit den Zwergameisenbären (Cyclopes) als deren einziges heute lebendes Mitglied zu dem Vermilingua gestellt. Die Myrmecophagidae und die Cyclopedidae trennten sich nach molekulargenetischen Untersuchungen im Mittleren Eozän vor rund 38 Millionen Jahren. Die Aufspaltung der Myrmecophagidae in Tamandua und Myrmecophaga begann laut denselben Untersuchungen zu Beginn des Oberen Miozäns vor 13 Millionen Jahren.[41][40][42]
Fossil ist Myrmecophaga erstmals im Pliozän fassbar,[19] zu den ältesten Funden gehören jene von Monte Hermoso aus der argentinischen Provinz Buenos Aires. Der aufgefundene dritte Mittelhandknochen gehört der ausgestorbenen Art Myrmecophaga caroloameghinoi an, welche ursprünglich in die eigene Gattung Nunezia verwiesen worden war.[43] Weitere alte Funde kamen aus El Breal de Orocual im Nordosten Venezuelas zum Vorschein, welche in den Übergang vom Pliozän zum Unteren Pleistozän zu stellen sind.[44] Funde aus Sonora im Nordwesten Mexikos zeigen, dass die Ameisenbärenart im Unteren Pleistozän im Zuge des Großen Amerikanischen Faunenaustausches bis weit nach Norden vordrang und später durch klimatische Veränderungen wieder auf ihr heutiges Verbreitungsgebiet zurückgedrängt wurde. Die südlichsten Funde stammen aus Uruguay. Allgemein ist der Große Ameisenbär recht häufig fossil aus dem Pleistozän überliefert.[45]
Heute werden drei Unterarten vom Großen Ameisenbär unterschieden:[2]
- M. t. artata Osgood, 1912; nordöstliches Kolumbien und nordwestliches Venezuela
- M. t. centralis Lyon, 1906; Mittelamerika bis nordwestliches Kolumbien
- M. t. tridactyla Linnaeus, 1758; Südamerika
Benannt wurde die Art Myrmecophaga tridactyla im Jahr 1758 von Linnaeus, welcher unter den gleichen Gattungsnamen weiterhin den Gemeinen Zwergameisenbären (Cyclopes didactylus) und den Südlichen Tamandua (Tamandua tetradactyla) einordnete und diese anhand der Anzahl der krallenbewehrten Vorderfußzehen unterschied. Als Typuslokalität für den Großen Ameisenbären gab Linnaeus ‘‘America Meridionali’’ an. Der Gattungsname Myrmecophaga setzt sich aus den griechischen Wörtern μύρμηξ (mýrmēx, Genitiv: μύρμηκος mýrmēkos „Ameise“) und φαγεῖν (phageín „fressen“) zusammen und bezieht sich auf die hauptsächliche Ernährungsweise der Tiere.[2]
Bedrohung und Schutz

Aufgrund seiner Größe ist der Große Ameisenbär ein häufiges Jagdziel, und sein Fleisch wird teilweise als Nahrungsressource genutzt. Allerdings ist diese Tradition unter indigenen Völkern sehr unterschiedlich. Laut einer Untersuchung erlegte die damals rund 800 Personen umfassende Volksgruppe der Waimiri Atroari im zentralen Amazonastiefland innerhalb eines Jahres (1993–1994) insgesamt drei Große Ameisenbären. Das Gesamtgewicht umfasste 90 Kilogramm, was rund 0,2 Prozent der gesamten über das Jahr erlegten Biomasse der Waimiri Atroari ausmachte.[46] Die Chamacoco aus Paraguay wiederum verzehren aus religiösen Gründen kein Fleisch des Großen Ameisenbären, während die Bewohner Nicaraguas das Fleisch aufgrund des Geschmacks meiden. Manchmal wird jedoch ein Großer Ameisenbär auch von Jägern oder deren Hunden getötet.[47] In der Regel meiden die Tiere Begegnungen mit Menschen, was aufgrund des schlechten Sehsinns und Gehörs aber nicht immer möglich ist. Ein bedrängtes Tier verteidigt sich mit seinen langen, scharfen Krallen, was in Einzelfällen auch für Menschen tödlich endete.[48] Der illegale Wildtierhandel und die Haltung als Haustier stellen weitere anthropogene Bedrohungen dar.[49]
Häufig ist die Ameisenbärenart allerdings Opfer von Verkehrsunfällen, vor allem in Offenland, da die Augen in der Nacht keinen Widerschein geben. Bedeutend ist auch der Einfluss von Feuer, das vor allem in Grasländern die einzelnen Populationen deutlich variieren lässt. Hauptfaktor für den Rückgang des Bestands ist aber die Zerstörung der Landschaften durch die Ausbreitung menschlicher Siedlungen und landwirtschaftlich genutzter Flächen, was vor allem für die mittelamerikanischen Populationen eine große Bedrohung ist.
Experten gehen für den Gesamtbestand von einem Rückgang von rund 30 Prozent seit 1996 aus. Die IUCN listet den Großen Ameisenbären als „gefährdet“ (vulnerable), regional bestehen aber noch höhere Warnstufen.[49] Die Tiere kommen in zahlreichen geschützten Gebieten vor, so unter anderem im Emas-Nationalpark, im Serra-da-Canastra-Nationalpark und im Iguaçu-Nationalpark in Brasilien, im letzteren konnte die Art erst 2014 nachgewiesen werden.[50] Im selben Zeitraum wurde der Große Ameisenbär mittels Kamerafallen im Baritú-Nationalpark im nördlichen Argentinien belegt.[51] Zudem gibt es ein internationales Zuchtprogramm in zoologischen Einrichtungen. Der Zoo Dortmund, wo über 50 Geburten verzeichnet werden konnten, führt das internationale Zuchtbuch.[52] Im Iberá Nature Reserve im Nordosten Argentiniens wurden zwischen 2007 and 2012 im Rahmen eines Wiederansiedlungsprojekts insgesamt 18 Große Ameisenbären frei gelassen und in ihrer Habitatwahl überwacht.[13]
Zu den weiteren Schutzmaßnahmen gehören auch Untersuchungen zur Einflussnahme von Wald- und Buschbränden auf die Bestandsentwicklung des Großen Ameisenbären und die Erarbeitung wirksamer Maßnahmen zum Brandschutz und zur Kontrolle natürlicher oder vom Menschen verursachter Buschbrände.[2][11]
Literatur
- Alessandra Bertassoni: Myrmecophagidae (Anteaters). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 74–90 (S. 88–89) ISBN 978-84-16728-08-4.
- Timothy J. Gaudin, Patrick Hicks und Yamil diBlanco: Myrmecophaga tridactyla (Pilosa: Myrmecophagidae). Mammalian Species 50 (956), 2018, S. 1–13.
- Ronald M. Nowak: Walker’s mammals of the world. 6. Auflage. Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9.
Einzelnachweise
- Timothy J. Gaudin, Patrick Hicks und Yamil diBlanco: Myrmecophaga tridactyla (Pilosa: Myrmecophagidae). Mammalian Species 50 (956), 2018, S. 1–13.
- Paul Smith: Giant Anteater Myrmecophaga tridactyla Linnaeus, 1758. Fauna of Paraguay 2, 2007, S. 1–18.
- Lydia Möcklinghoff, Karl-L. Schuchmann und Marinêz I. Marques: New non-invasive photo-identification technique for free-ranging giant anteaters (Myrmecophaga tridactyla) facilitates urgently needed field studies. Journal of Natural History 52 (37–38), 2018, S. 2397–2411.
- Alessandra Bertassoni: Myrmecophagidae (Anteaters). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 74–90 (S. 89–90) ISBN 978-84-16728-08-4.
- Hideki Endo, Nobuharu Niizawa, Teruyuki Komiya, Shinichiro Kawada, Junpei Kimura, Takuya Itou, Hiroshi Koie und Takeo Sakai: Three-Dimensional CT Examination of the Mastication System in the Giant Anteater. Zoological Science 24, 2007, S. 1005–1011.
- Hideki Endo, Teruyuki Komiya, Shinichiro Kawada, Akiko Hayashida, Junpei Kimura, Takuya Itou, Hiroshi Koie und Takeo Sakai: Three-Dimensional Reconstruction of the Xenarthrous Process of the Thoracic and Lumber Vertebrae in the Giant Anteater. Mammal Study 34 (1), 2009, S. 1–6.
- Christy M. McCain: First Evidence of the Giant Anteater (Myrmecophaga tridactyla) in Honduras. The Southwestern Naturalist 46 (2), 2001, S. 252–254.
- Hector Orlando Portillo Reyes, Wilfredo Antonio Matamoros und Sheri L. Glowinski: Distribution and Conservation Status of the Giant Anteater (Myrmecophaga tridactyla) in Honduras. The Southwestern Naturalist 55 (1), 2010, S. 118–120.
- Marcio A. Martinez, Manfredo Alejandro Turcios-Casco und Shasling Pacheco Amador: On the conservation of Myrmecophaga tridactyla (Pilosa: Myrmecophagidae) in the core of Río Plátano Biosphere Reserve, Honduras. Mammalia, 2020, doi:10.1515/mammalia-2019-0152.
- Guillermo Pérez Jimeno und Lucía Llarín Amaya: Contribución al Conocimiento de la Distribución del oso Hormiguero Gigante (Myrmecophaga tridactyla) en Argentina. Edentata 8–10, 2009, S. 8–12.
- Flávia Miranda und I. M. Medri: Myremecophaga tridactyla. Edentata 11 (2), 2010, S. 103–104.
- James Sanderson und Leandro Silveira: Observations of Xenarthra in the Brazilian Cerrado and Guyana. Edentata 5, 2003, S. 40–44.
- Yamil E. Di Blanco,Ignacio Jiménez Pérez und Mario S. Di Bitetti: Habitat Selection in Reintroduced Giant Anteaters: The Critical Role of Conservation Areas. Journal of Mammalogy 96 (5), 2015, S. 1024–1035.
- Arnaud Léonard Jean Desbiez undÍsis Meri Medri: Density and Habitat Use by Giant Anteaters (Myrmecophaga tridactyla) and Southern Tamanduas (Tamandua tetradactyla) in the Pantanal Wetland, Brazil. Edentata 11 (1), 2010, S. 4–10.
- Constança de Sampaio, Paiva Camilo-Alves und Guilherme de Miranda Mourão: Responses of a Specialized Insectivorous Mammal (Myrmecophaga tridactyla) to Variation in Ambient Temperature. Biotropica 38 (1), 2006, S. 52–56.
- Rosane G. Collevatti, Kelly C. E. Leite, Guilherme H. B. de Miranda und Flavio H. G. Rodrigues: Evidence of high inbreeding in a population of the endangered giant anteater, Myrmecophaga tridactyla (Myrmecophagidae), from Emas National Park, Brazil. Genetics and Molecular Biology 30 (1), 2007, S. 112–120.
- Flávio H. G. Rodrigues, Ísis. M. Medri, Guilherme. H. B. de Miranda, Constança. Camilo-Alves und Guilherme Mourão: Anteater behavior and ecology. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The biology of the Xenarthra. University Press of Florida, 2008, S. 257–268
- Fernanda G. Braga, Raphael E. F. Santos und Antonio C. Batista: Marking behavior of the giant anteater Myrmecophaga tridactyla (Mammalia: Myrmecophagidae) in Southern Brazil. Zoologia 27 (1), 2010, S. 7–12.
- Sue D. Hirschfeld: A new fossil anteater (Edentata, Mammalia) from Colombia, S.A. and evolution of the Vermilingua. Journal of Paleontology 50 (3), 1976, S. 419–432.
- Louisa Mamalis, Suenny Sousa da Silva, Andressa Bezerra Valerio, Darren Norris und Fernanda Michalski: Stepping stones facilitate river crossings by Myrmecophaga tridactyla in the north-eastern Brazilian Amazon. Edentata 19, 2018, S. 30–34.
- Robert J. Young, Carlyle M. Coelho und Dalía R. Wieloch: A note on the climbing abilities of giant anteater, Myrmecophaga tridactyla (Xenarthra, Myrmecophagidae). Boletim do Museu de Biologia Mello Leitão, Nova Série 15, 2003, S. 41–46.
- Fabiana L. Rocha und Guilherme Mourão: An Agonistic Encounter Between Two Giant Anteaters (Myrmecophaga tridactyla). Edentata 7, 2006, S. 50–51.
- Kolja Kreutz, Frauke Fischer und K. Eduard Linsenmair: Observations of Intraspecific Aggression in Giant Anteaters (Myrmecophaga tridactyla). Edentata 8–10, 2009, S. 6–7.
- Mariana Labão Catapani, Karina Theodoro Molina, Alexandre Martins Costa Lopes und Flávia Miranda: Report of three non-agonistic encounters of free-living giant anteaters (Myrmecophaga tridactyla). Edentata 20, 2019, S. 31–34.
- Ísis Meri Medri und Guilherme Mourão: A Brief Note on the Sleeping Habits of the Giant Anteater Myrmecophaga tridactyla (Xenarthra, Myrmecophagidae). Revista Brasileira de Zoologia 22, 2005, S. 1213–1215.
- Kent H. Redford: Dietary specialization and variation in two mammalian myrmecophages (variation in mammalian myrmecophagy). Revista Chilena de Historia Natural 59, 1986, S. 201–208.
- Ísis Meri Medri, Guilherme de Miranda Mourão und Ana Yoshi Harada: Dieta de Tamanduá-Bandeira (Myrmecophaga tridactyla) no Pantanal da Nhecolândia, Brasil. Edentata 5, 2003, S. 29–34.
- Jorge Alberto Gallo, Agustín Manuel Abba, Luciana Elizalde, Dante Di Nucci, Tatiana Agustina Ríos und María Cecilia Ezquiaga: First study on food habits of anteaters, Myrmecophaga tridactylaand Tamandua tetradactyla, at the southern limit of their distribution. Mammalia 81 (6), 2017, S. 601–604.
- Fernanda Góss Braga, Nilton José Souza, Antonio Carlos Batista und Pedro Pacheco dos Santos Lima: Consumo de formigas cortadeiras por tamanduá-bandeira Myrmecophaga tridactyla (Linnaeus, 1758) em plantios de Pinus spp. no Paraná, Brasil. Edentata 15, 2014, S. 1–8.
- Vivian E. Sandoval-Gómez, Héctor E. Ramírez-Chaves und David Marín: Registros de hormigas y termitas presentes en la dieta de osos hormigueros (Mammalia: Myrmecophagidae) en tres localidades de Colombia. Edentata 13, 2012, S. 1–9.
- Guilherme Henrique Braga de Miranda, Flávio Henrique Guimarães Rodrigues, Ísis Meri Medri und Fernanda Vinci dos Santos: Giant Anteater Mymecophaga tridactyla Beehive Foraging at Emas National Park, Brazil. Edentata 5, 2003, S. 55.
- Katrina K Knott, Beth M Roberts, Morgan A Maly, Carrie K Vance, Jennifer DeBeachaump, Jackie Majors, Peter Riger, Heather DeCaluwe und Andrew J Kouba: Fecal estrogen, progestagen and glucocorticoid metabolites during the estrous cycle and pregnancy in the giant anteater (Myrmecophaga tridactyla): evidence for delayed implantation. Reproductive Biology and Endocrinology 11, 2013, S. 83 doi:10.1186/1477-7827-11-83.
- Jorge Anthony Astwood Romero, Patrícia Carolina Casas Martínez, Sol Ángela Ojeda Holguín und Ricardo Murillo Pacheco: Notas sobre el Comportamiento de Cortejo y Apareamiento de Myrmecophaga tridactyla Bajo Condiciones Ex Situ. Edentata 11 (1), 2010, S. 34–43.
- José Fernando Miranda Júnior und Alessandra Bertassoni: Potential agonistic courtship and mating behavior between two adult giant anteaters (Myrmecophaga tridactyla). Edentata 15, 2014, S. 69–72.
- Danielle D. Brown und Miranda A. Moore: Giant anteater (Myrmecophaga tridactyla) mothers may teach their calves what “not to eat”. Edentata 19, 2018, S. 61–66.
- Sara de Valle Jerez und Monique Halloy: El Oso Hormiguero Myrmecophaga tridactyla: Crecimiento e Independización de una Cria. Mastozoologia Neotropical 10, 2003, S. 323–330.
- Sandra M. C. Cavalcanti und Eric M. Gese: Kill rates and predation patterns of jaguars (Panthera onca) in the southern Pantanal, Brazil. Journal of Mammalogy 91 (3), 2010, S. 722–736.
- Cristian Gabriel Orlando und Gustavo Javier Fernández: Respuesta antidepredatoria de osos hormigueros (Myrmecophaga tridactyla) mantenidos en cautividad. Edentata 15, 2014, S. 52–59.
- Adrian G. Mutlow, Michael W. Dryden und Patricia A. Payne: Flea (Pulex simulans) infestation in captive Giant anteater (Myrmecophaga tridactyla). Journal of Zoo and Wildlife Medicine 37 (3), 2006, S. 427–429.
- Frédéric Delsuc, Mariella Superina, Marie-Ka Tilak, Emmanuel J. P. Douzery und Alexandre Hassanin: Molecular phylogenetics unveils the ancient evolutionary origins of the enigmatic fairy armadillos. Molecular Phylogenetics and Evolution 62, 2012, 673–680.
- Frédéric Delsuc, Sergio F Vizcaíno und Emmanuel JP Douzery: Influence of Tertiary paleoenvironmental changes on the diversification of South American mammals: a relaxed molecular clock study within xenarthrans. BMC Evolutionary Biology 4 (11), 2004, S. 1–13.
- Gillian C. Gibb, Fabien L. Condamine, Melanie Kuch, Jacob Enk, Nadia Moraes-Barros, Mariella Superina, Hendrik N. Poinar und Frédéric Delsuc: Shotgun Mitogenomics Provides a Reference Phylogenetic Framework and Timescale for Living Xenarthrans. Molecular Biology and Evolution 33 (3), 2015, S. 621–642.
- H. Gregory McDonald, Sergio F. Vizcaíno und M. Susana Bargo: Skeletal anatomy and the fossil history of the Vermilingua. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 64–78.
- Ascanio D. Rincón, Gilberto E. Parra, Francisco J. Prevosti, Maria teresa Alberdi und Christopher J. Bell: A preliminary assessment of the Mammalian fauna from the Pliocene-Pleistocene El Breal de Orocual locality, Monagas state, Venezuela. In: L. B. Albright III (Hrsg.): Papers on Geology, Vertebrate Paleontology, and Biostratigraphy in Honor of Michael O. Woodburne. Museum of Northern Arizona Bulletin 65, 2009, S. 593–620.
- Christopher A. Shaw und H. Gregory McDonald: First Record of Giant Anteater (Xenarthra, Myrmecophagidae) in North America. Science 236, 1987, S. 186–188.
- Roselis Remor de Souza-Mazurek, Temehe Pedrinho, Xinymy Feliciano, Waraié Hilário, Sanapyty Gerôncio und Ewepe Marcelo: Subsistence hunting among the Waimiri Atroari Indians in central Amazonia, Brazil. Biodiversity and Conservation 9:, 2000, S. 579–596.
- Jeremy M. Koster: Giant Anteaters (Myrmecophaga tridactyla) Killed By Hunters With Dogs In the Bosawas Biosphere Reserve, Nicaragua. The Southwestern Naturalist 53 (3), 2008, S. 414–416.
- Vidal Haddad Jr., Guilherme C. Reckziegel, Domingos G. Neto und Fábio L. Pimentel: Human Death Caused by a Giant Anteater (Myrmecophaga trydactila) in Brazil. Wilderness & Environmental Medicine 2014 (sciencedirect.com).
- Flávia Miranda, A. Bertassoni und A. M. Abba: Myrmecophaga tridactyla. In: IUCN 2014. IUCN Red List of Threatened Species. Version 2012.2. e.T14224A47441961 (), zuletzt abgerufen am 29. Juni 2013.
- Anne-Sophie Bertrand und Amadeu M. V. M. Soares: First reports of Giant anteater (Myrmecophaga tridactyla) and Greater naked-tailed armadillo (Cabassous tatouay) for the Iguaçu National Park, Paraná, Brazil, with notes on all xenarthran occurrences. Edentata 19, 2018, S. 35–41.
- Norberto Ángel Nigro, Nicolás Lodeiro Ocampo und Luz Estefanía Paronzini: Primeros registros mediante cámaras trampa de oso hormiguero gigante (Myrmecophaga tridactyla) en el Parque Nacional Baritú, provincia de Salta, Argentina. Edentata 21, 2020, S. 34–37.
- Zoo Dortmund: Großer Ameisenbär. (), zuletzt abgerufen am 12. August 2013.
Weblinks
- Myrmecophaga tridactyla in der Roten Liste gefährdeter Arten der IUCN 2008. Eingestellt von: G. Porini u. a., 2008. Abgerufen am 29. Juni 2013.
- Yvonne Maier: Der Ameisenbär - Minigehirn, wehender Schweif, klebrige Zunge Bayern 2 Radiowissen. Ausstrahlung am 13. November 2020 (Podcast)