Lestodon
Lestodon ist eine Gattung aus der heute ausgestorbenen Familie der Mylodontidae, die große, bodenlebende Faultiere umfasste. Es gehört neben Megatherium und Eremotherium zu den größten bekannten Faultieren überhaupt, war mit diesen beiden aber nicht näher verwandt. Funde von Lestodon sind vor allem aus der Pamparegion und nördlich anschließenden Gebieten in Südamerika überliefert und datieren vom Pliozän vor rund 5 Millionen Jahren bis in das ausgehende Pleistozän vor etwa 10.000 Jahren, wobei Fossilreste aus dem Oberen Pleistozän weitaus häufiger beobachtet werden. Sie lassen annehmen, dass der Faultiervertreter hauptsächlich offene Landschaften bewohnte.
| Lestodon | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Lestodon | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Pliozän bis frühes Holozän | ||||||||||||
| 5,33 Mio. Jahre bis 10.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Lestodon | ||||||||||||
| Gervais, 1855 | ||||||||||||
Höchstwahrscheinlich ernährte sich Lestodon überwiegend von Gräsern, was durch den Bau des Gebisses und der allgemeinen Schädelgestaltung belegt werden kann. Es stellte aufgrund der Körperproportionen einen eher langsamen Wanderer dar, der sich aber bei Bedarf auf den Hinterbeinen aufrichten konnte. Möglicherweise war der Faultiervertreter befähigt, im Boden zu wühlen, wie es von anderen Mitgliedern der Mylodonten belegt ist, doch war er für intensive Grabungsaktivitäten weniger gut ausgestattet. Der Aufbau des Gehörs spricht dafür, dass Lestodon Frequenzen im Infraschall wahrnehmen konnte und wahrscheinlich auch darüber kommunizierte.
Die wissenschaftliche Erstbeschreibung der Gattung geht auf das Jahr 1855 zurück und wurde von Paul Gervais durchgeführt. Bereits 1841 hatte aber ein dänischer Zoologe Funde beschrieben, die heute zu Lestodon gestellt werden. Sein verwendeter Name Platygnathus ist aber nicht anerkannt. Im Laufe der Forschungsgeschichte wurden zahlreiche Arten der Gattung beschrieben. Untersuchungen lassen jedoch vermuten, dass höchstwahrscheinlich nur eine Art im Pleistozän Südamerikas präsent war.
Merkmale
Allgemein und Körpergröße
Lestodon war ein sehr großer Vertreter der bodenbewohnenden Faultiere und der mit Abstand größte bekannte Vertreter der Mylodontidae. Seine Körperlänge wird mit 4,6 m angegeben.[1] Einigen Berechnungen zufolge erreichten einzelne Individuen ein Körpergewicht von 2,7 bis 3,6 t,[2][3] andere Annahmen gehen von bis zu 4,1 t[4] aus. Er gehörte somit neben Megatherium und Eremotherium zu den größten bekannten Bodenfaultieren überhaupt, war aber mit beiden nicht näher verwandt.[5] Im Bezug auf die Körperproportionen wich Lestodon von den beiden anderen genannten riesigen Bodenfaultieren ab, da es als Vertreter der Mylodonten sehr kurze untere Gliedmaßen besaß. Wie alle bodenlebenden Faultiere war es insgesamt durch gegenüber den Hinterbeinen kürzere Vorderbeine und durch einen – abweichend von den heutigen, baumlebenden Arten – langen Schwanz gekennzeichnet.[4]
Schädel- und Gebissmerkmale

Der Schädel von Lestodon ähnelte dem seines nahen Verwandten Glossotherium, war aber deutlich größer. Er zeichnete sich allgemein durch eine in der Aufsicht nahezu prismatisch-rechteckige Form aus. Seine Länge variierte von 52 bis 65 cm, die Gesamthöhe lag bei maximal 19 cm. Der größte Teil des Rostrums wurde durch den Oberkiefer gebildet, der in Seitenansicht rechteckig und sehr hoch gestaltet war. In der Aufsicht erweiterte er sich im vorderen Bereich stark, sodass die maximale Breite am vorderen Ende des Oberkiefers bis zu 25 cm betrug.[6] Dadurch entstand eine sehr weite und hohe Nasenöffnung, was ein deutlicher Unterschied zu der eher schmalen Schnauze von Mylodon ist. Das Nasenbein war seitlich vollständig mit dem Oberkiefer verwachsen, die Knochennähte zwischen den beiden Schädelknochen zeigten einen konvexen Verlauf, sodass das Nasenbein in der Mitte deutlich eingeschnürt war. In der Regel übertraf die hintere Breite in Kontakt mit dem Stirnbein die vordere erheblich. Der Mittelkieferknochen war, wie bei allen Faultieren typisch, sehr kurz. Er hatte eine Bogenform und besaß nur eine lockere Verbindung mit dem Oberkiefer. Ebenso zeichnete sich der Jochbogen entsprechend zahlreichen anderen Faultieren – mit Ausnahme der großen Megatherien – als nicht geschlossen aus. Der vordere, nach hinten orientierte Bogenteil, der vom Jochbein ausgeht, besaß hinten eine massive, plattenartige Verbreiterung und endete in drei Fortsätzen: einem aufsteigenden oberen, einem abwärtsführenden unteren und einem weitgehend horizontal orientierten mittleren. Während der aufsteigende und absteigende Fortsatz jeweils gut entwickelt waren, wies der mittlere in der Regel nur eine geringe Länge auf. Der Bogenabschnitt des Schläfenbeins zeigte nach vorn und besaß eine schlanke, fingerartige Form. Die beiden Enden des vorderen und hinteren Bogenabschnitts lagen wie bei allen Mylodonten sehr eng beieinander. Das Hinterhauptsbein war breiter als hoch und wies einen kräftigen Querwulst auf. In seiner Form unterschied es sich eindeutig von dem deutlich flacheren Hinterhauptsbein bei Glossotherium und dem eher kreisförmigen bei Mylodon.[7][8][2]
Der Unterkiefers besaß eine Länge zwischen 37 und 51 cm. Die Unterkante des horizontalen Knochenkörpers verlief weitgehend gerade. Er wies seine größte Höhe am hintersten Backenzahn auf, wo er bis zu 12,5 cm maß.[9] Seine Höhe nahm nach vorn kontinuierlich ab, im Bereich der Symphyse wurde er wieder mächtiger. Die Symphyse selbst war extrem kräftig und breit, analog dem sehr weiten Oberkiefer, und stieg am Vorderrand des Unterkiefers in einem Winkel von 45° auf. Der verlängerte vordere Bereich erreichte die halbe Länge der Zahnreihe, die Gesamtlänge der Symphyse betrug etwa 10 cm. Sie wies vorn eine gerade oder nur leicht gewölbte Abschlusskante auf. Der Winkelfortsatz am hinteren Ende des Unterkiefers war prominent ausgebildet und lag deutlich unterhalb der Zahnebene. Das Gebiss wies die für Faultiere typische Struktur auf. Es setzte sich aus 5 Zähnen je Kieferast im Oberkiefer und 4 je Kieferast im Unterkiefer zusammen, bestand also aus insgesamt 18 Zähnen. Der jeweils vorderste Zahn war in Form eines Eckzahns gestaltet (caniniform) und hatte teilweise recht große Ausmaße. Durch den breiten Bau des Maules standen diese eckzahnartigen Zähne im Unterkiefer bis zu 17 cm auseinander.[10] Zwischen dem vordersten, caniniformen Zahn und den hinteren Zähnen bestand ein sehr ausgedehntes Diasterma, das im Oberkiefer durchschnittlich 6,8 cm, im Unterkiefer rund 5,5 cm erreichte. Die hinteren Backenzähne zeigten einen molarenartigen Bau. Charakteristisch war die für späte Mylodonten flache bis leicht eingesenkte Kaufläche, die so eine lappenartige (lobate) Gestaltung erhielt. Die Zähne bestanden im Innern aus einer harten Variante des Zahnbeins (Orthodentin) und wurden außen von einer Hülle aus Zahnzement umschlossen. Die Länge der gesamten Zahnreihe im Unterkiefer betrug bis zu 19 cm,[9] wovon die molarenartigen Zähne etwa 11 cm beanspruchten. Die Länge der einzelnen hinteren Backenzähne variierte hier zwischen 3 und 4 cm, der hinterste war in der Regel am größten.[8][11][2]
Körperskelett

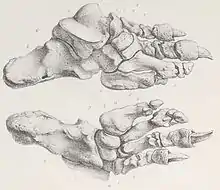
Das Körperskelett ist relativ gut überliefert. Der Oberarmknochen wurde bis zu 51 cm lang und besaß ein für Bodenfaultiere typisches massives unteres Gelenk mit einem Durchmesser von gut 26 cm. Im Vergleich zu anderen großen Mylodonten war der Oberarmknochen aber relativ grazil gebaut. Dem gegenüber betrug die Länge der Elle bis zu 46 cm. Sie verfügte entsprechend anderen nahe verwandten Formen über einen recht ausgedehnten körpernahen Gelenkfortsatz, das Olecranon, welches bis zu 16 cm in der Länge maß.[4] Der charakteristisch gestaltete Oberschenkelknochen, der eine Länge von bis zu 74 cm aufwies und am oberen Gelenkende bis zu 34 cm breit wurde, war extrem abgeplattet und wirkte dadurch brettartig flach. Er ähnelte dem von Mylodon, hatte aber größere Ausmaße. Von den Megatherien unterschied sich der Oberschenkelknochen durch die weniger deutlich rechteckig-breite Gestaltung in der Ansicht von vorn. Am Schaft war ein dritter Trochanter als Muskelansatzstelle ausgebildet, der aber nicht sehr erhaben wirkte. Schien- und Wadenbein waren teilweise am oberen Ende miteinander verwachsen, was aber innerhalb der Gattung eher variabel auftrat.[12] Beide zeichneten sich durch ihren wiederum eher flachen und sehr kurzen Bau aus, das Schienbein erreichte gegenüber dem Oberschenkelknochen nur knapp die Hälfte an Länge. Dies ist typisch für zahlreiche Mylodonten und unterscheidet diese von den Megatherien, bei denen die unteren Beinabschnitte verhältnismäßig länger waren.[13] Der Hinterfuß von Lestodon stimmte weitgehend mit dem anderer Mylodonten überein. Er besaß insgesamt vier Strahlen (II bis V), wobei nur die beiden inneren (II und III) Krallen trugen. Die ersten beiden Zehenglieder des zweiten Strahls waren üblicherweise miteinander verwachsen, abweichend von Paramylodon und übereinstimmend mit Glossotherium bestand der dritte Strahl aber aus drei Phalangen. Die jeweiligen Endglieder konnten hier mit 14 bis 17 cm sehr lang werden. Die beiden äußeren Strahlen (IV und V) wiesen nur jeweils zwei Zehenglieder auf, demzufolge waren hier keine Krallen ausgebildet. Dafür besaßen die Mittelfußknochen hier einen deutlich massiveren Bau.[10][14][3] Wie bei zahlreichen großen Bodenfaultieren war der Fuß nach innen gedreht und stützte mit dem äußersten Strahl und dem Fersenbein auf den Boden auf (pedolateral).[15]
Verbreitung und bedeutende Fossilfunde

Lestodon lebte vom Pliozän bis zum ausgehenden Pleistozän vor rund 5 Millionen bis vor etwa 10.000 Jahren, sein hauptsächliches Verbreitungsgebiet umfasste die heutigen Pampagebiete in Südamerika. Dabei überschnitt sich das Vorkommen mit dem des riesigen Megatheriums (speziell Megatherium americanum). Möglicherweise bestanden in diesen damals teils offenen Landschaften gewisse ökologische Unterschiede, die eine direkte Konkurrenz der beiden Faultiervertreter minimierten, was aber bisher noch nicht genau untersucht ist. Auffällig ist eine häufige Konzentration von Fundstellen mit Resten von Lestodon im heutigen Uruguay östlich des Río de la Plata, wo Megatherium allgemein nur selten nachgewiesen wurde. Westlich des Flusses im heutigen Argentinien dagegen ist Lestodon weitaus weniger häufig überliefert. Nach Norden hin reichen Fossilfunde der Faultiergattung bis zum brasilianischen Bundesstaat São Paulo, wo es teilweise auch zu Überschneidungen mit Eremotherium kommt.[16] Als nördliche Verbreitungsgrenze wird der 23. südliche Breitengrad angegeben.[17] Des Weiteren haben die Schelfgebiete als wichtige Fossillager in jüngerer Zeit an Bedeutung zugenommen.[18] Insgesamt sind Fossilfunde von Lestodon aber deutlich seltener als solche von Megatherium oder Eremotherium.[5]
Einer der frühesten Nachweise der Gattung Lestodon stammt mit einem nur 33 cm langen Schädel eines Jungtiers aus Farolo Monte Hermoso in der argentinischen Provinz Buenos Aires. Der Fund wurde in der Monte-Hermoso-Formation geborgen und datiert in das Untere Pliozän.[19] Bedeutende Fossilreste des Unteren und Mittleren Pleistozäns sind vor allem aus der Pamparegion Argentiniens und aus dem Tarija-Tal im südlichen Bolivien überliefert.[20][2] Ein möglicherweise aus dieser Zeit stammendes schädelloses Teilskelett kam in Flussablagerungen des „Arroio do Lestodon“ bei Caçapava do Sul im brasilianischen Bundesstaat Rio Grande do Sul zu Tage.[12] Der Großteil der bekannten Funde ist aber dem Oberpleistozän zuzuweisen. Als außerordentlich fundreich erwies sich die Küstenregion von Uruguay. Hervorzuheben sind unter anderem die Küstenklippen von Arroyo El Caño im uruguayischen Department Colonia, wo in fluviatilen Ablagerungen unzählige Fossilreste lagern, die in das Mittlere und Obere Pleistozän gehören. Etwa 70 % des geborgenen Materials kann Lestodon zugeordnet werden, es umfasst aber zumeist Einzelknochen in unterschiedlichem Erhaltungszustand, zusammengehöriges Skelettmaterial ist kaum überliefert. Aufgrund des gemeinsamen Vorkommens mit dem großen grasfressenden Huftier Toxodon oder des Kamelartigen Hemiauchenia sowie zahlreicher Vertreter der gepanzerten Glyptodontidae lassen sich hier offene und trockene Landschaften rekonstruieren. Aus der nahe gelegenen Bucht von Colonia de Sacramento kam ein nahezu vollständiges Skelett von Lestodon zu Tage.[21] Von herausragender Bedeutung ist die Fundstelle Arroyo del Vizcaíno nahe der Ortschaft Sauce im südöstlichen Uruguay, von der eine umfangreiche Fossilfauna stammt. In den dort auf nur 30 m² aufgeschlossenen sandig-kiesigen Ablagerungen wurden bisher mehr als 1000 Knochenreste geborgen, deren Alter laut radiometrischen Datierungen etwa 32.300 bis 31.200 Jahre B.P. beträgt. Mehr als 90 % des identifizierbaren Materials gehört zu Lestodon, die bisher rund 730 Stücke entstammen allen Bereichen des Skelettes. Sie repräsentieren wenigstens ein Dutzend ausgewachsene Individuen und zwei Jungtiere. Einige besondere Oberflächenmodifikationen der Knochen, etwa mögliche Schnittspuren,[22][23][24][25] ließen die Bearbeiter der Fundstelle an menschliche Manipulationen denken, womit Arroyo del Vizcaíno einer der frühsten Nachweise der Besiedlung Südamerikas durch frühe Jäger-Sammler-Gruppen wäre (in der Regel gilt Monte Verde in Chile mit einem Alter von rund 14.800 Jahren als eine der ältesten Fundstellen Südamerikas[26]).[27] Die Interpretation wird aber nicht vollständig geteilt.[28] Wiederum in Playa del Barco im Südwesten der argentinischen Provinz Buenos Aires kamen auf rund 500 m² über 280 Knochen von Lestodon zu Tage. Diese können insgesamt 13 Individuen zugeordnet werden, darunter Jungtiere, ausgewachsene und extrem alte Tiere. Das Alter des Fundplatzes beträgt rund 19.849 Jahre vor heute. Die Reste fanden sich in Gemeinschaft mit anderen großen Säugetieren wie Bodenfaultieren, etwa Megatherium, Glyptodonten oder Rüsseltieren, hier besonders Notiomastodon, aber auch verschiedenen Raubtieren.[29]
Die jüngsten Funde von Lestodon kamen aus dem ausgehenden Oberpleistozän zu Tage. Zu erwähnen sind unter anderem ein 74 cm langer Oberschenkelknochen und ein nahezu vollständiger Schädel aus der La Paz local fauna aus dem südlichen Uruguay, die auf ein Alter von 17.620 Jahre B.P. datieren.[30] Bereits in den Übergang zum Holozän gehören Reste, die auf dem bedeutenden archäologischen Fundplatz Paso Otero 5 in der argentinischen Provinz Buenos Aires gefunden wurden. Hier lagen auf knapp 100 m² gut 80.000 Knochenreste, die teilweise stark zerschlagen oder gebrannt waren und so nur zu einem geringen Umfang bestimmbar sind. Darunter konnten lediglich zwei Fragmente von Lestodon identifiziert werden. Die Anwesenheit des Menschen hier bezeugen weiterhin über 80 Steinartefakte aus Quarzit, darunter auch Fischschwanz-Spitzen (fish tail points).[31][32] Das Alter der Funde wurde mit Hilfe der Radiokarbonmethode auf 10.440 bis 10.190 Jahre BP bestimmt, einige jüngere Daten sprechen auch für einen Zeitraum um 9400 bis 9560 Jahre BP. Damit starb Lestodon wie zahlreiche andere Bodenfaultiere zum Ende der letzten Kaltzeit im Zuge der Quartären Aussterbewelle aus.[33]
Paläobiologie
Fortbewegung
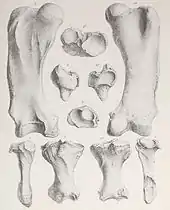
Das Längenverhältnis der unteren zu den oberen Beinabschnitten (Crural Index) gibt Hinweise auf die Agilität eines Tieres. Dabei sind kurze untere Abschnitte typisch für schwerfällige Tiere, etwa Nashörner, während lange untere Beinknochen wie bei den Pferden mit schnellen Tieren in Verbindung stehen. Mylodonten zeigen allgemein sehr kurze untere Gliedmaßenabschnitte. Bei Lestodon erreichen sie mit einer Schienbeinlänge von rund 34 bis 37 cm gegenüber einer Oberschenkelknochenlänge von 68 bis 73 cm nur etwa die Hälfte der oberen.[13][3] Dagegen waren bei Megatherium die unteren Beinabschnitte deutlich länger. Seine Tibialänge variiert zwischen 42 und 62 cm, die Femurlänge zwischen 57 und 77 cm. Somit dürfte Lestodon wie zahlreiche andere Vertreter der Mylodonten an eine deutlich schwerfälligere Gangart angepasst gewesen sein als etwa die Megatherien.[34] Beim Laufen bewegte sich Lestodon auf nach innen gedrehten Füßen vorwärts, sodass diese den Untergrund mit dem äußeren Strahl (Strahl V) und dem Fersenbein berührten. Dies wird als pedolateraler Gang bezeichnet und ist mit Ausnahme der Megalonychidae für alle bodenlebenden Faultiere charakteristisch. Bei den Mylodonten war der Fuß aber nicht so stark gewölbt wie bei den verwandten Scelidotheriidae, sodass das Fersenbein mit der gesamten Länge aufsetzte und nicht nur mit dem hinteren Ende. Dadurch ähnelte der Fuß eher dem der großen Megatheriiden.[15][4]
Für einige Vertreter der Mylodonten, etwa Glossotherium, vermuten Forscher, dass sie gute Bodengräber waren. Indiziert wird dies neben der allgemein kräftigen Ausprägung der vorderen Gliedmaßen durch das extrem ausgedehnte Olecranon, den oberen Gelenkfortsatz der Elle. Eine deutliche Verlängerung des Olecranons verstärkt enorm die Hebelwirkung des Unterarms, da dort der Trizeps ansetzt. Dies führt dazu, dass der Arm eher für hohe Kraftentfaltung eingesetzt werden kann, als für die Erreichung großer Geschwindigkeiten bei der Fortbewegung. Bei Lestodon nimmt das Olecranon rund 35 % der Gesamtlänge des Knochens ein, was etwa dem Wert von Glossotherium entspricht und im unteren Wertebereich der heutigen Gürteltiere liegt, die mitunter als gute Gräber gelten. Ein mögliches Bodengraben von Lestodon könnte auch durch die weit nach hinten versetzte Lage des Körperschwerpunktes im Bereich des Bauchs kurz vor dem Ansatz der Hintergliedmaßen unterstützt worden sein. Dadurch wurden in einer vierfüßigen Position rund 68 bis 71 % des Körpergewichts von den Hinterbeinen getragen, was im Verhältnis umgekehrt ist zu zahlreichen heutigen Huftieren, bei denen sich die Hauptlast auf die Vorderbeine verteilt. Dies befähigte die Faultiergattung, sich auf den Hinterbeinen aufzurichten und so in einen zweifüßigen Stand überwechseln, was die Vorderbeine für andere Tätigkeiten frei werden ließ. Beispielsweise wird beim heutigen Riesengürteltier (Priodontes maximus) als ausgezeichnetem Gräber die Hauptlast des Körpers beim Graben von den Hinterbeinen getragen. Auch die symmetrischen und deutlich gebogenen Krallen der Vorderfüße von Lestodon konnten Belastungsanalysen zufolge den beim Graben auftretenden Zug- und Kompressionskräften gut widerstehen.[35][36] Es ist aber anzunehmen, dass das riesige Faultier mit seinem deutlich grazil gestalteten Oberarmknochen im Vergleich zu seinem Verwandten Glossotherium schlechter für Grabtätigkeiten ausgestattet war. Wahrscheinlich war es auch weniger notwendig als bei kleineren Vertretern, da ausgewachsene Individuen von Lestodon wohl kaum Fressfeinde zu fürchten brauchten.[4] allerdings verweisen Isotopenuntersuchungen an Funden von Smilodon, dass Lestodon durchaus zum Beutespektrum der Säbelzahnkatze gehört haben könnte.[37] Einige extrem große begrabene Baue aus dem südöstlichen Brasilien, die ein schätzungsweise pliozänes bis pleistozänes Alter haben, werden dennoch teilweise mit Lestodon in Verbindung gebracht (unter der Bezeichnung als Ichnospezies Megaichnus major). Diese sind mit rund 2 m Höhe, 4 m Breite und mitunter bis zu 50 m Länge deutlich größer als vergleichbare Strukturen in Argentinien, für die zumeist Glossotherium verantwortlich gezeichnet wird.[38][17]
Ernährung
Die Vertreter der Mylodontidae werden häufig als auf Grasnahrung spezialisierte Faultiere angesehen. Dafür sprechen unter anderem die hohen (hypsodonten) Zahnkronen der Backenzähne, die auch bei grasfressenden Huftieren wie den Pferden auftreten. Da den Faultierzähnen aber der harte Zahnschmelz fehlt, sind Vergleiche zwischen diesen beiden Gruppen nur bedingt möglich. Lestodon besitzt innerhalb der Mylodonten vergleichsweise niedrigere Backenzähne als Mylodon oder Paramylodon. Sie sind auch deutlich niedriger als bei Megatherium und Eremotherium aus der Familie der Megatheriidae, die aber eher weiche Pflanzenkost verzehrten.[9] Außen bestehen die Zähne aus einer Schicht sehr hartem Orthodentin, das auf der Kaufläche der hinteren Backenzähne eine jeweils etwas erhöhte Leiste oder einen Rand ausbildet. Im Zahninneren befindet sich das weichere Vasodentin, welches auf der Kaufläche eine kleine Senke bildet, sodass diese insgesamt eine lappen- oder zungenartige (lobate) Ausprägung besitzt. Dieses für zahlreiche Mylodonten charakteristische Kauflächenmuster spricht eher dafür, dass die Nahrung zermahlen wurde und daher aus hartem bis faserigem Pflanzenmaterial bestand. Der Kauapparat zeigt ebenfalls einzelne Besonderheiten. Die eher seitwärtsorientierten und leicht gewölbten Gelenkflächen des Unterkiefers lassen sowohl seitliche als auch vor- und rückwärts gerichtete Kaubewegungen zu. Gegen deutlich seitliche Kieferbewegungen spricht aber der nicht vollständig ausgebildete Jochbogen, da er den dadurch entgegengesetzt wirkenden Kräften des Musculus masseter und Musculus pterygoideus nicht standhalten konnte. Deswegen scheint eine Dominanz vor- und rückwärts gerichteter Kaubewegungen wahrscheinlich, was auch mit entsprechende Abnutzungsspuren an den Zähnen übereinstimmt. Durch die niedrige Lage des Unterkiefergelenkes nur wenig oberhalb der Kauebene sind zudem vertikale Kaubewegungen nur eingeschränkt möglich gewesen, ebenso wie die Beißkraft durch die Anordnung der Kaumuskulatur eher gering war. Aufgrund dieser anatomischen Merkmale war Lestodon wohl an grashaltige Nahrung angepasst und konnte größere Mengen auf einmal aufnehmen. Im Gegensatz zu pflanzenfressenden Huftieren besitzen die Mylodonten aber eine extrem niedrige Gesamtkaufläche aller molarenartiger Backenzähne zusammengenommen, die bei Lestodon etwa 1000 bis 1100 mm² betrug (ein vergleichbar großes Flusspferd besitzt etwa drei- bis fünfmal so viel Kaufläche, heutige Elefanten von vergleichbarer Größe etwa acht- bis elfmal so viel und mehr). Daraus wird geschlossen, dass Lestodon die Nahrung im Mundraum nur schlecht zerkleinern konnte und dadurch entweder einen großen Teil dieser im Magen-Darm-Trakt verarbeitete oder wie heutige Faultiere eine eher geringe Stoffwechselrate aufwies.[39][8][11]
Indirekt wird auch durch die Form des Mauls auf eine grasfresserische Ernährungsweise geschlossen. So zeigt Lestodon ähnlich wie Glossotherium eine sehr breite Maulpartie mit weit auseinander stehenden Rändern des Oberkiefers, was deutlich abweichend ist von der schmaleren Schnauze bei Mylodon oder bei Megatherium. Rekonstruktionen zufolge war es mit breiten, wulstigen Lippen ausgestattet, die vergleichbar dem heutigen Breitmaulnashorn gut zur Aufnahme großer Mengen an Gräsern in relativ kurzer Zeit geeignet sind.[6] Zudem wird bei den Mylodonten mit breiten Schnauzen aufgrund der Gestaltung des Zungenbeins und seiner weit nach hinten geschobenen Position auf eine lange und bewegliche Zunge geschlossen.[40] In der Regel ist dieser Knochen aber nur selten erhalten, was mit seiner Fragilität zusammenhängt. Von Lestodon wurde bisher lediglich ein Exemplar eines Jungtiers aufgefunden.[25] Im Gegensatz zu Mylodon liegen von Lestodon keine Dungreste in Form von Koprolithen vor, so dass die konsumierte Nahrung nicht direkt analysiert werden kann. Mit Hilfe von Isotopenuntersuchungen an Fossilresten aus Arroyo del Vizcaíno in Uruguay konnte jedoch die Landschaft ermittelt werden, in der die Tiere lebten, was Rückschlüsse auf deren prinzipielle Ernährungsweise erlaubt. Sie ergaben Hinweise auf kühle und trockene Klimaverhältnisse während des ausgehenden Pleistozäns und dementsprechend das Überwiegen offener Landschaften. Die Bedingungen ähneln denen, die heute im nördlichen Patagonien vorherrschen. Die dadurch hauptsächlich verbreiteten Gräser stehen somit auch im Einklang mit den anatomisch ermittelten Hinweisen zur Ernährungsweise von Lestodon.[41] Weitere Isotopenuntersuchungen an Resten mehrerer Individuen aus Playa del Barco geben für Lestodon eine eher gemischte Pflanzenkost an mit einer stärkeren Bevorzugung von Grasnahrung. Da aber auch eine hohe individuelle Variabilität in den Ergebnissen vorliegt, kann eine größere Flexibilität bei der Nahrungsaufnahme nicht ausgeschlossen werden.[29]
Sozialverhalten und Sexualdimorphismus
Die bei Playa del Barco aufgefundenen rund ein Dutzend Individuen von Lestodon umfassen Jungtiere und ausgewachsene Individuen unterschiedlicher Altersstufen. Ihre enge räumliche Distanz zueinander und ihr vergleichbarer Verwitterungsgrad lässt eine etwa zeitgleiche Einbettung der ehemaligen Kadaver annehmen. Wissenschaftler gehen daher davon aus, dass Lestodon zumindest zeitweise auch größere Gruppen oder Familienverbände formte. Unter welchen Umständen dies geschah, ist nicht eindeutig, die heutigen Baumfaultiere sind strikt einzelgängerisch. Vermutlich hatten Konkurrenz mit anderen großen Pflanzenfressern oder Schutz vor Beutegreifern einen gewissen Einfluss.[29] Eine vergleichbare Lebensweise wird auch bei Massenfunden von Eremotherium angenommen.[42]
Als ein weiteres Indiz für zumindest temporäre Gruppenbildungen kann ein markanter Sexualdimorphismus gewertet werden, der sich bei Lestodon in der Ausprägung der unteren vorderen caniniformen Zähne abzeichnet. Es lassen sich hierbei zwei Größentypen feststellen, von denen Individuen mit kleineren vorderen Zähnen eventuell auf weibliche, solche mit größeren auf männliche Tiere verweisen. Ähnliches ist auch von Paramylodon bekannt, wo sich der Sexualdimorphismus lediglich in der Robustizität des Schädels äußert. Bei Glossotherium und bei einigen Vertretern der großen Megatherien sind hingegen Unterschiede in den Körperausmaßen bei den beiden Geschlechtern ersichtlich. Ein starker Sexualdimorphismus bei heutigen Säugetieren, wie etwa bei einigen Primaten und Huftieren, ist in der Regel mit einer polygynen Fortpflanzungsstrategie verbunden, was teilweise Rangordnungskämpfe unter den männlichen Tieren einschließt. Auffällige Zahndimorphismen zwischen den Geschlechtern finden sich heute unter anderem neben den Primaten auch bei Elefanten und beim Flusspferd. Ein weiteres hervorzuhebendes Kennzeichen bei Individuen mit kleineren caniniformen Zähnen ist ein vergrößerter hinterer molarenartiger Zahn. Ähnliche Größenzunahmen von den vorderen zu den hinteren Mahlzähne lassen sich bei einigen Nagetieren belegen, die sich strikt pflanzenfressend ernähren. Eventuell stand dies bei Lestodon mit höheren energetischen Kosten in Verbindung, die während der Trächtigkeit und Laktation aufkamen und eine effizientere Nahrungsaufnahme und -verwertung erforderten.[43][44]
Sinnesleistungen, Weichteile und Pathologien
Heutige Faultiere haben allgemein ein schlechtes Sehleistungsvermögen, ebenso ist das Gehör nur wenig entwickelt und auf Frequenzen von 0,3 bis 30 kHz mit einem Optimum bei 2 bis 8 kHz ausgelegt.[45] Von Lestodon sind einzelne Gehörknöchelchen des Mittelohrs überliefert, namentlich der Amboss, der Hammer und der Steigbügel. Alle drei Knöchelchen leiten die akustischen Signale vom Außenohr zur Hörschnecke im Innenohr weiter. Größe und Gewicht der Gehörknöchelchen sind nicht direkt abhängig von der Körpergröße eines Individuums, sondern lassen in Verbindung mit der Ausdehnung des Trommelfells und der Viskosität des Mittelohrs auf die Befähigung schließen, einen bestimmten Frequenzbereich wahrzunehmen. Amboss und Hammer wiegen bei Lestodon zusammen rund 500 mg. Dies entspricht etwa den Werten, die für die gleichen Knöchelchen von Glossotherium bekannt sind und für den anhand einiger gut erhaltener Schädel eine Trommelfellgröße von 180 bis 208 mm² angenommen werden kann. Im ähnlichen Größenbereich liegen auch heutige Elefanten und Hundsrobben. Für beide Säugetiervertreter ist eine Befähigung zur Wahrnehmung sehr niedriger Frequenzen nachgewiesen. Aufgrund dessen wird für Lestodon ein Hörvermögen im Frequenzbereich von 0,05 bis 16,5 kHz mit einem Optimum von 1,7 bis 2,4 kHz angenommen. Der untere Wert liegt dabei deutlich im Infraschallbereich. Laute mit niedrigen Frequenzen werden über weite Strecken getragen und kaum von Vegetation abgelenkt, sie können demnach auch noch aus großer Entfernung geortet werden. Möglicherweise wurden sie von Lestodon zur Kommunikation mit Artgenossen über weite Entfernungen in den offenen Landschaften der Pampa eingesetzt, beispielsweise für Brunftrufe, ähnlich wie es bei heutigen Elefanten der Fall ist. Andererseits können mit einem derartig ausgelegten Gehör auch Fressfeinde rechtzeitig erkannt werden. Nicht zuletzt stehen sie eventuell auch mit der grabenden Lebensweise in Verbindung, da einige heutige aktiv im Untergrund lebende Tiere, beispielsweise die Taschenratten, auf diese Weise auch seismische Wellen wahrnehmen können.[46][47]
Ebenso wie bei den anderen riesigen bodenlebenden Faultieren, etwa Megatherium und Eremotherium, wird aufgrund der Verbreitung in Landschaften unter gemäßigten bis zum Teil subtropischen Klimabedingungen im Gegensatz zu zahlreichen Lebendrekonstruktionen keine dichte Fellbedeckung für Lestodon angenommen. Gegen die Ausprägung eines dichten Körperfells sprechen hauptsächlich die enorme Körpergröße und die damit verbundene, notwendige Thermoregulation. Da ein dichtes Fellkleid die Körperwärme schlecht ableitet, würde dessen Vorhandensein extrem große Mengen an Trinkwasser voraussetzen, um den Energiehaushalt auszugleichen. Demnach besaß Lestodon ähnlich wie andere große, heutige Pflanzenfresser wie Elefanten, Nashörner oder Flusspferde eine mehr oder weniger nackte Haut.[48] Das bereits erwähnte Zungenbein liegt fragmentarisch vor und weist eine Länge von 10,5 cm auf. Es stammt von einem Jungtier, im Vergleich zu dem gleichen Knochen von Glossotherium könnte das Zungenbein bei Lestodon vollständig rund 17,5 cm Länge erreicht haben. Unter Annahme einer proportionalen Entwicklung wie bei heutigen Hausrindern lässt sich für die Zunge des Jungtiers ein Gewicht von etwa 10 kg annehmen.[25]
Mehrere der bei Playa del Barco aufgefundenen Lestodon-Reste wiesen pathologische Veränderungen wie deformierte Knochen auf, besonders an den Wirbeln. Zumeist sind diese auf Osteoporose und Osteoarthrose zurückzuführen, was häufig bei großen Säugetieren belegt werden kann. Gelegentlich traten Osteophyten auf, welche möglicherweise die Bewegungen der Tiere einschränkten. Auch ist Osteochondritis an einzelnen Schwanzwirbeln belegt, was eventuell auf die Stützfunktion beim Aufrichten auf die Hinterbeine zurückzuführen ist.[29]
Systematik
Innere Systematik der Mylodontidae nach Boscaini et al. 2019 (basierend auf skelettanatomischen Analysen)[49]
|
Innere Systematik der Faultiere nach Presslee et al. 2019 (basierend auf Proteinanalysen)[50]
|
Lestodon ist eine heute ausgestorbene Gattung aus der ebenfalls erloschenen Familie der Mylodontidae. Die Mylodontidae stellen wiederum einen Teil der Unterordnung der Faultiere (Folivora) dar. Zusammen mit den Orophodontidae und den Scelidotheriidae formen die Mylodontidae gemeinsam die Überfamilie der Mylodontoidea (teilweise werden die Scelidotheriidae und die Orophodontidae aber auch nur als Unterfamilie innerhalb der Mylodontidae geführt[51]). Die Mylodontoidea bilden in einer auf skelettanatomischen Merkmalen beruhenden Gliederung neben den Megatherioidea die zweite große und bedeutende Faultierlinie. Nach molekulargenetischen und proteinbasierten Analysen können diesen beiden Großgruppen noch die Megalocnoidea zur Seite gestellt werden. Gemäß diesen Studien beinhalten die Mylodontoidea mit den Zweifinger-Faultieren (Choloepus) auch eine der zwei heute noch lebenden Faultiergattungen.[52][50] Die Mylodontidae stellen eine der formenreichsten Gruppen innerhalb der Faultiere dar. Generell werden sie durch ihre hochkronigen Zähne mit abweichend von den Megatherioidea flacher (lobater) Kaufläche charakterisiert, die eine Anpassung an stärker grashaltige Nahrung darstellen. Die hinteren Zähne zeigen einen runden oder ovalen Querschnitt, die vordersten sind eckzahnartig gestaltet. Der Hinterfuß besitzt zudem eine deutlich seitlich ausgedrehte Gestalt.[53][15] Die frühesten Nachweise der Mylodonten stammen unter anderem mit Paroctodontotherium bereits aus dem Oligozän, aufgefunden in den Salla Beds im Salla-Luribay-Becken in Bolivien.[54]
Häufig wird Lestodon innerhalb der Mylodontidae in die Unterfamilie der Lestodontinae gestellt (diese war 1889 von Florentino Ameghino als eigenständige Familie eingeführt, von Lucas Kraglievich aber 1931 auf den Status der Unterfamilie innerhalb der Mylodontidae verschoben worden; teilweise wird sie heute auch auf tribaler Ebene als Lestodontini geführt). Nahe Verwandte bilden dadurch Formen wie Thinobadistes, möglicherweise auch Glossotherium, während das bekannte und für die Familie namengebende Mylodon in einer eigenen Unterfamilie, den Mylodontinae steht. Diese Untergliederung ist zumeist akzeptiert und konnte auch in einer der bisher umfangreichsten Studien zur Stammesgeschichte der Faultiere durch Timothy J. Gaudin aus dem Jahr 2004 bestätigt[55] und durch nachfolgende Untersuchungen untermauert werden.[56][57] Eine Analyse aus dem Jahr 2019, erstellt durch ein Arbeitsteam um Luciano Varela, verweist Lestodon wiederum in eine Gruppe aus Mylodonten des nördlichen Südamerikas und Mittelamerikas wie Bolivartherium und Thinobadistes, sieht aber zu Glossotherium eine größere verwandtschaftliche Distanz, welches eng an Paramylodon gebunden ist.[51] Ein in Bezug auf Lestodon nahezu gleichwertiges Ergebnis lieferte eine im selben Jahr vorgestellte Arbeit von Alberto Boscaini und Forscherkollegen, die eine höherauflösende stammesgeschichtliche Untersuchung der Mylodonten beinhaltete. In ihr findet auch die deutliche Zweiteilung der terminalen Mylodonten in die Mylodontinae und Lestodontinae, die bereits in der erwähnten Untersuchung von Gaudin aus dem Jahr 2004 herausgearbeitet worden war, ihre Bestätigung. Als deutlicher Unterschied zwischen den beiden Gruppen lässt sich die Ausprägung der vorderen eckzahnartigen Zähne heranziehen. Die Lestodontinae weisen große caniniforme Zähne in deutlichem Abstand zu den hinteren molarenartigen Zähnen auf, bei den Mylodontinae sind diese klein oder teilweise zurückgebildet und stehen enger an den molariformen Zähnen an.[49] Die gegenüber Glossotherium, Mylodon und zusätzlich Paramylodon bestehende Außenposition von Lestodon ergaben auch die bereits angeführten biochemischen Studien des Jahres 2019.[50] Allgemein ist die innere Gliederung der Mylodontidae komplex und variiert je nach Bearbeiter. Teilweise wurden dadurch zahlreiche weitere Unterfamilien aufgestellt, etwa die Nematheriinae für Vertreter aus dem Unteren Miozän oder die Octomylodontinae für alle Basalformen, die aber nicht allgemein anerkannt sind.[58] Mit der Etablierung der Urumacotheriinae für spätmiozäne Gattungen des nördlichen Südamerikas wurde im Jahr 2004 eine weitere Linie innerhalb der Mylodonten herausgearbeitet,[59] die aber nach den Untersuchungen von 2019 keinen Bestand hat.[51] Prinzipiell wird für die gesamte Familie eine Revision angemahnt, da zahlreiche der höheren taxonomischen Einheiten keine formale Diagnose besitzen.[13]
Im Laufe der Forschungsgeschichte wurden über ein Dutzend Arten innerhalb der Gattung Lestodon beschrieben. In der Regel werden aber meist nur drei in der wissenschaftlichen Literatur berücksichtigt:
- Lestodon armatus Gervais, 1855
- Lestodon australis Kraglievich, 1934
- Lestodon trigonidens Gervais, 1873
Die Eigenständigkeit von L. trigonidens ist dabei häufig Gegenstand von Diskussion. Die Art geht auf Paul Gervais zurück, der sie im Jahr 1873 anhand eines vorderen Unterkieferfragments mit im Vergleich zu L. armatus noch kräftigeren eckzahnartigen vorderen Zähnen aufstellte.[10] Die meisten Wissenschaftler sehen heute diese unterschiedliche Ausprägung der vorderen eckzahnartigen Zähne als Ausdruck eines Sexualdimorphismusses an, womit L. trigonidens identisch mit L. armatus wäre. Demnach könnte die erstere Art mit männlichen Tieren in Verbindung gebracht werden, die letztere mit weiblichen.[60] Auch für L. australis ist der Status als eigene Art problematisch. Diese wurde bisher nur in Argentinien nachgewiesen und 1934 von Lucas Kraglievich mit Hilfe eines nahezu vollständigen, 56 cm langen Schädels mit lediglich fehlenden vorderen Jochbögen eingerichtet. Er stammte aus Playa del Barco bei Monte Hermoso in der argentinischen Provinz Buenos Aires. Anatomische Untersuchungen ergaben aber, dass der Schädel in der Variationsbreite der Merkmale von L. armatus liegt und somit keine abweichenden oder definierenden Charakteristika aufweist. Demnach lebte während des Pleistozäns mit L. armatus nur eine Art von Lestodon.[2][61] Für den bisher ältesten bekannten Fund aus dem Pliozän gibt es aufgrund der Zugehörigkeit des Schädels zu einem nicht ausgewachsenen Individuum bisher keine eindeutige Artbestimmung.[19] Im Jahr 2004 wurden aus der Urumaco-Formation des Oberen Miozäns im nordwestlichen Venezuela zwei Arten von Lestodon anhand von Schädel- und Skelettresten beschrieben, die nicht nur deutlich kleiner und älter waren als die bisherigen Funde der Gattung, sondern auch weit außerhalb des bekannten Verbreitungsgebietes lagen.[62] Zwei Jahre später erhielten sie dann eine Zuweisung zu der mit Lestodon nahe verwandten Gattung Bolivartherium.[63] Das Fundmaterial erwies sich bei nachfolgenden Arbeiten aber als zum Teil gemischt, so dass die postcranialen Elemente ausgesondert und in die Gattung Magdalenabradys überführt wurden.[64]
Forschungsgeschichte
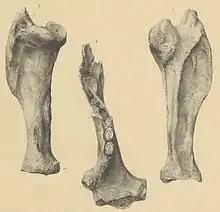
Zu den frühesten Funden, die höchstwahrscheinlich Lestodon zugewiesen werden können, gehören jene, die der dänische Zoologe Henrik Nikolai Krøyer (1799–1870) von seiner Reise nach Südamerika in den Jahren 1840 und 1841 mitgebracht hatte. Während dieser Reise fand er zahlreiche Knochen ausgestorbener Tiere am Río de la Plata auf der Buenos Aires gegenüberliegenden Uferseite nordwestlich der uruguayischen Stadt Colonia de Sacramento. Er stellte diese in einem kurzen Vorbericht 1841 vor, beschrieb aber nur einen Unterkiefer genauer, dem er wegen seiner breiten Form die wissenschaftliche Bezeichnung Platygnathus platensis gab. Aufgrund der Ausprägung von seiner Meinung nach insgesamt fünf Zähnen, darunter einen vorderen spitz gestalteten, verwies er ihn in die Nähe der damals bekannten Faultiere wie Choloepus (Zweifinger-Faultiere) und zog auch Vergleiche zu Megatherium, dem er korrekterweise nur vier Unterkieferzähne zusprach.[65] Erst mehr als 30 Jahre später wurden die Funde erneut vom ebenfalls dänischen Zoologen Johannes Theodor Reinhardt (1816–1882) untersucht. Sie waren in der Zwischenzeit teils stark beschädigt worden, allerdings erkannte Reinhardt, dass sie aufgrund unterschiedlicher Erhaltungszustände kein einheitliches Material darstellen. Darunter befanden sich unter anderem auch Reste weiterer ausgestorbener Tiere, etwa von Glyptodon, einem großen gepanzerten Vertreter der Glyptodontidae, oder von Toxodon aus der Gruppe der Südamerikanischen Huftiere. Der Großteil umfasste aber Knochen von riesigen Faultieren, so Teile der Wirbelsäule und der Gliedmaßen. An dem von Krøyer beschriebenen Unterkiefer bemerkte Reinhardt die fehlerhafte Angabe von fünf Zähnen und korrigierte die Anzahl auf die tatsächlich vorhandenen vier. Die allgemeinen Merkmale der Funde verleiteten Reinhardt dazu, sie der zwei Dekaden zuvor beschriebenen Faultiergattung Lestodon zuzuweisen.[66] Die Bezeichnung Platygnathus platensis wird heute nicht mehr verwendet, der Gattungsname Platygnathus ist zudem seit 1832 durch einen Vertreter der Bockkäfer präokkupiert.[2]

Bei seiner Zuweisung berief sich Reinhardt auf die Arbeiten des französischen Naturwissenschaftlers Paul Gervais (1816–1879). Dieser hatte die wissenschaftliche Erstbeschreibung von Lestodon im Jahr 1855 vorgelegt. Für die Einführung der neuen Gattung stand Gervais Fundmaterial aus der argentinischen Provinz Buenos Aires zur Verfügung, die ins Muséum national d’histoire naturelle in Paris verbracht worden waren. Gervais bemerkte zwar Ähnlichkeiten zu Mylodon, hob aber die enormen caniniformen vorderen Zähne im Unter- und Oberkiefer hervor. Aufgrund dessen benannte er die Art Lestodon armatus. Die Holotypen umfassen einen fragmentierten Oberkiefer sowie einen ebenfalls nur teilweise überlieferten Unterkiefer (Exemplarnummern MNHNP-PAM 90 und 91). Bei beiden Stücken sind der jeweils vordere caniniforme Zahn, im Oberkiefer zudem der erste und im Unterkiefer die ersten beiden molarenartigen Zähne erhalten, ebenso wie oben und unten die Alveole des jeweils nachfolgenden Zahnes.[2] Gervais führte gleichzeitig mit Lestodon myloides eine zweite Art ein, in deren kürzeren, vorderen Zähnen er Anklänge zu Mylodon sah. (L. myloides wurde 1880 von Henri Frédéric Paul Gervais (1845–1915) und Florentino Ameghino (1854–1911) zu Pseudolestodon verwiesen,[67] ein heutiges Synonym für Glossotherium.) Der Gattungsname Lestodon leitet sich von den griechischen Wörtern λῃστής (lēstḗs „Räuber“ „Dieb“) sowie ὀδούς (odoús „Zahn“) ab und bezieht sich auf die eckzahnartige Gestaltung des vordersten Zahns, der Gervais zufolge dem Tier einen raubtierartigen Charakter verlieh.[68][5]
In dem Aufsatz der Erstbeschreibung erwähnt Gervais Lestodon nur kurz, eine weit ausführlichere Beschreibung legte er 1873 vor, in der er auch zahlreiche Elemente des Körperskelettes mit einbezog.[10] Bereits 1880 führten Gervais und Ameghino in einem Überblickswerk zur Fauna Südamerikas insgesamt acht Arten von Lestodon (eine davon unbenannt), das aber keine differenzierte Beschreibung beinhaltete.[67] Sie erwähnen dabei nicht Krøyers Platygnathus platensis, da der Aufsatz von Reinhardt zwar auf das Jahr 1875 datiert, aber erst im selben Jahr wie Gervais’ und Ameghinos Überblick erschien. Zwei Jahre darauf erkennen Mitglieder der Deutschen Geologischen Gesellschaft L. platensis unter Berufung auf Knochenfunde aus Uruguay, die der Gesellschaft von Rudolf Virchow geschenkt worden waren, an und verbinden die Art mit der von Gervais und Ameghino nicht eindeutig benannten Form.[69] Weitere Arten wurden vor allem von Lucas Kraglievich (1886–1932) in den frühen 1930er Jahren eingeführt. Aber bereits rund 20 Jahre später zweifelte Carlos de Paula Couto (1910–1982) die zahlreichen unterschiedlichen Arten von Lestodon an. So sprach er sich auch 1979 dafür aus, mit L. armatus nur eine gültige Art anzuerkennen.[2]
Literatur
- Ada Czerwonogora und Richard A. Fariña: How many Pleistocene species of Lestodon (Mammalia, Xenarthra, Tardigrada)? Journal of Systematic Palaeontology 11 (1-2), 2013, S. 249–261
- Richard A. Fariña, Sergio F. Vizcaíno und Gerardo De Iuliis: Megafauna. Giant beasts of Pleistocene South America. Indiana University Press, 2013, S. 1–436 (S. 212–213) ISBN 978-0-253-00230-3
Einzelnachweise
- American Museum of Natural History: Lestodon armatus. ()
- Ada Czerwonogora und Richard A. Fariña: How many Pleistocene species of Lestodon (Mammalia, Xenarthra, Tardigrada)? Journal of Systematic Palaeontology 11 (1-2), 2013, S. 249–261
- Richard A. Fariña, Sergio F. Vizcaíno und María S. Bargo: Body mass estimations in Lujanian (Late Pleistocene-Early Holocene of South America) mammal megafauna. Mastozoología Neotropical 5 (2), 1998, S. 87–108
- M. Susana Bargo, Sergio F. Vizcaíno, Fernando M. Archuby und R. Ernesto Blanco: Limb bone proportions, strength and digging in some Lujanian (Late Pleistocene-Early Holocene) mylodontid ground sloths (Mammalia, Xenarthra). Journal of Vertebrate Paleontology 20 (3), 2000, S. 601–610
- Richard A. Fariña, Sergio F. Vizcaíno und Gerardo De Iuliis: Megafauna. Giant beasts of Pleistocene South America. Indiana University Press, 2013, S. 1–436 (S. 212–213) ISBN 978-0-253-00230-3
- M. Susana Bargo, Néstor Toledo und Sergio F. Vizcaíno: Muzzle of South American Pleistocene Ground Sloths (Xenarthra, Tardigrada). Journal of Morphology 267, 2006, S. 248–263
- Luciano Brambilla und Damián A. Ibarra: The occipital region of late Pleistocene Mylodontidae of Argentina. Boletín del Instituto de Fisiografía y Geología 88, 2018, S. 1–9
- M. Susana Bargo und Sergio F. Vizcaíno: Paleobiology of Pleistocene ground sloths (Xenarthra, Tardigrada): biomechanics, morphogeometry and ecomorphology applied to the masticatory apparatus. Ameghiniana 45 (1), 2008, S. 175–196
- M. Susana Bargo, Gerardo De Iuliis und Sergio F. Vizcaíno: Hypsodonty in Pleistocene ground sloths. Acta Palaeontologica Polonica 51 (1), 2006, S. 53–61
- Paul Gervais: Mémoire sur plusieurs espècies de mammifères fossils propres a l’Amerique Méridionale. Mémoires de la Société géologique de France, Série 2, 9, 1873, S. 1–44 ()
- Sergio F. Vizcaíno: The teeth of the “toothless”: novelties and key innovations in the evolution of xenarthrans (Mammalia, Xenarthra). Paleobiology 35 (3), 2009; S. 343–366
- Dilson Vargas-Peixoto, Cícero Schneider Colusso, Átila Augusto Stock Da-Rosa und Leonardo Kerber: A new record of Lestodon armatus Gervais 1855 (Xenarthra, Mylodontidae) from the Quaternary of southern Brazil and remarks on its postcranial anatomy. Historical Biology, 2019, doi:10.1080/08912963.2019.1597075
- Ascanio D. Rincón, H. GregoryMcDonald, Andrés Solórzano, Mónica Núñez Flores und Damián Ruiz-Ramoni: A new enigmatic Late Miocene mylodontoid sloth from northern South America. Royal Society Open Science 2, 2015, S. 140256, doi:10.1098/rsos.140256
- Chester Stock: Structure of the pes in Mylodon harlani. University of California Publications. Bulletin of the Department of Geology 10 (16), 1917, S. 267–286
- H. Gregory McDonald: Evolution of the Pedolateral Foot in Ground Sloths: Patterns of Change in the Astragalus. Journal of Mammalian Evolution 19, 2012, S. 209–215
- Carlos Castor Cartelle: Preguiças terrícolas, essas desconhecidas. Anales Ciência Hoje 27, 2000, S. 19–25
- Renato Pereira Lopez, Heinrich Theodor Frank, Francisco Sekiguchi de Carvalho Buchmann und Felipe Caron: Megaichnus igen. nov.: Giant paleoburrows attributed to extinct Cenozoic mammals from South America. Ichnos 24 (2), 2017, S. 133–145, doi:10.1080/10420940.2016.1223654
- Alex Sandro Schiller Aires und Renato Pereira Lopes: Representativity of Quaternary mammals from the Southern Brazilian continental shelf.Revista Brasileira de Paleontologia 15(1), 2012, S. 57–66
- Cecilia M. Deschamps, Gracielia I. Esteban und M. Susana Bargo: El registro más Antigua del género Lestodon Gervais 1855 (Xenarthra, Tardigrada, Mylodontidae) (Montehermosense, Plioceno Temprano). Ameghiniana 38 (2), 2001, S. 151–156
- Esteban Soibelzon, Ángel Ramón Miño-Boilini, Alfredo Eduardo Zurita und Cecilia Mariana Krmpotic: Los Xenarthra (Mammalia) del Ensenadense (Pleistoceno inferior a medio) de la Región Pampeana (Argentina). Revista Mexicana de Ciencias Geológicas 27 (3), 2010, S. 449–469
- Andrés Rinderknecht: Vertebrados fósiles de la costa uruguaya. In: R. Menafra, L. Rodríguez-Gallego, F. Scarabino und D. Conde (Hrsg.): Bases para la Conservación y el Manejo de la Costa Uruguaya. Montevideo, 2006, S. 343–349
- Richard A. Fariña und Reynaldo Castilla: Earliest evidence for human-megafauna interaction in the Americas. In: E. Corona-M und J Arroyo-Cabrales (Hrsg.): Human and faunal relationships reviewed: an archaeozoological approach. Oxford, 2007, S. 31–34
- Richard A. Fariña, P. Sebastián Tambusso, Luciano Varela, Mariana Di Giacomo, Marcos Musso, Andrés Gascue und Roberto Bracco: Among others, cut-marks are archaeological evidence: reply to ‘Archaeological evidences are still missing: a comment on Fariña et al. Arroyo del Vizcaíno Site, Uruguay’ by Suárez et al. Proceedings of the Royal Society B 281, 2014, doi:10.1098/rspb.2014.1637
- Richard A. Fariña: Bone surface modifications, reasonable certainty and human antiquity in the Americas: the case of the Arroyo del Vizcaíno. American Antiquity 80 (1), 2015, S. 193–200
- P. Sebastián Tambusso, H. Gregory McDonald und Richard A. Fariña: Description of the stylohyal bone of a giant sloth (Lestodon armatus). Palaeontologia Electronica 18 (1), 2015, S. 19A ()
- Anthony D. Barnosky und Emily L. Lindsey: Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. Quaternary International 217, 2010, S. 10–29
- Richard A. Fariña, P. Sebastián Tambusso, Luciano Varela, Ada Czerwonogora, Mariana Di Giacomo, Marcos Musso, Roberto Bracco und Andrés Gascue: Arroyo del Vizcaíno, Uruguay: a fossil-rich 30-ka-old megafaunal locality with cut-marked bones. Proceedings of the Royal Society B 281, 2014, S. 20132211, doi:10.1098/rspb.2013.2211
- Rafael Suárez, Luis A. Borrero, Karen Borrazzo, Martín Ubilla, Sergio Martínez und Daniel Perea: Archaeological evidences are still missing: a comment on Fariña et al. Arroyo del Vizcaíno site, Uruguay. Proceedings of the Royal Society B 281, 2014, doi:10.1098/rspb.2014.0449
- Rodrigo L. Tomassini, Claudia I. Montalvo, Mariana C. Garrone, Laura Domingo, Jorge Ferigolo, Laura E. Cruz, Dánae Sanz‑Pérez, Yolanda Fernández‑Jalvo und Ignacio A. Cerda: Gregariousness in the giant sloth Lestodon (Xenarthra): multi‑proxy approach of a bonebed from the Last Maximum Glacial of Argentine pampas. Scientific Reports 10, 2020, S. 10955, doi:10.1038/s41598-020-67863-0
- Andrea Corona, Daniel Perea, Pablo Toriño und César Goso: Taphonomy, sedimentology and chronology of a fossiliferous outcrop from the continental Pleistocene of Uruguay. Revista Mexicana de Ciencias Geológicas 29 (2), 2012, S. 514–525
- Gustavo Martínez, María Gutiérrez und José Luis Prado: New Archaeological Evidences from the Late-Pleistocene/Early-Holocene Paso Otero 5 Site (Pampean Region, Argentina). Current Research in the Pleistocene 21, 2004, S. 16–18
- Gustavo Martínez: Arqueología del curso medio del Río Quequén Grande: Estado actual y aportes a la arqueología de la región Pampeana. Relaciones de la Sociedad Argentina de Antropología 31, 2006, S. 249–275
- Marína A. Gutiérrez und Gustavo A. Martínez: Trends in the faunal human exploitation during the Late Pleistocene and Early Holocene in the Pampean region (Argentina). Quaternary International 191, 2008, S. 53–68
- Gerardo De Iuliis, Guillermo H. Ré und Sergio F. Vizcaíno: The Toro Negro megatheriine (Mammalia, Xenarthra): A new species of Pyramiodontherium and a review of Plesiomegatherium. Journal of Vertebrate Paleontology 24 (1), 2004, S. 214–227
- Santiago Pantiño und Richard A. Fariña: Ungual phalanges analysis in Pleistocene ground sloths (Xenarthra, Folivora). Historical Biology 29 (8), 2017, S. 1065–1075, doi:10.1080/08912963.2017.1286653
- Santiago Pantiño, Jorge Peréz Zerpa und Richard A. Fariña: Finite element and morphological analysis in extant mammal's claws and Quaternary sloth' ungual phalanges. Historical Biology, 2019, doi:10.1080/08912963.2019.1664504
- Hervé Bocherens, Martin Cotte, Ricardo Bonini, Daniel Scian, Pablo Straccia, Leopoldo Soibelzon und Francisco J. Prevosti: Paleobiology of sabretooth catSmilodon populatorin the Pampean Region (Buenos Aires Province, Argentina) around the Last Glacial Maximum: Insights from carbon and nitrogen stable isotopes in bone collagen. Palaeogeography, Palaeoclimatology, Palaeoecology 449, 2016, S. 463–474, doi:10.1016/j.palaeo.2016.02.017
- Francisco Segikurch de Carvalho Buchmann, Heinrich Theodor Frank, Victor Moreira Sandim Ferreira und Erick Antal Cruz: Evidência de vida gregária em paleotocas attribuítas a Mylodontidae (Preguiças-gigantes). Revista Brasileira de Paleontologia 19 (2), 2016, S. 259–270
- Sergio F. Vizcaíno, M. Susana Bargo und Guillermo H. Cassini: Dental occlusal surface area in relation to body mass, food habits and other biological features in fossil xenarthrans. Ameghiniana 43 (1), 2006, S. 11–26
- Leandro M. Pérez, Néstor Toledo, Gerardo De Iuliis, M. Susana Bargo und Sergio F. Vizcaíno: Morphology and Function of the Hyoid Apparatus of Fossil Xenarthrans (Mammalia). Journal of Morphology 271, 2010, S. 1119–1133
- Ada Czerwonogora, Richard A. Fariña und Eduardo Pedro Tonni: Diet and isotopes of Late Pleistocene ground sloths: first results for Lestodon and Glossotherium (Xenarthra, Tardigrada). Neues Jahrbuch für Geologie und Paläontologie, Abhandlunge 262, 2011, S. 257–266
- Emily L. Lindsey, Erick X. Lopez Reyes, Gordon E. Matzke, Karin A. Rice und H. Gregory McDonald: A monodominant late-Pleistocene megafauna locality from Santa Elena, Ecuador: Insight on the biology and behavior of giant ground sloths. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, S. 109599, doi:10.1016/j.palaeo.2020.109599
- Luciano Varela, Sebastián Tambusso und Richard Fariña: Unexpected inhibitory cascade in the molariforms of sloths (Folivora, Xenarthra): A case study in xenarthrans honouring Gerhard Storch’s open mindedness. Fossil Imprint 76 (1), 2020, S. 1–16
- Luciano Varela, H. Gregory McDonald und Richard Fariña: Sexual dimorphism in the fossil ground sloth Lestodon armatus (Xenarthra, Folivora). Historical Biology: An International Journal of Paleobiology, 2021, doi:10.1080/08912963.2021.1933470
- Desmond Gilmore, Denia Fittipaldi Duarte und Carlos Peres da Costa: The physiology of two- and three-toed sloth. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 130–142
- R. Ernesto Blanco und Andrés Rinderknecht: Estimation of Hearing Capabilities of Pleistocene Ground Sloths (Mammalia, Xenarthra) from Middle-Ear Anatomy. Journal of Vertebrate Paleontology 28 (1), 2008, S. 274–276
- R. Ernesto Blanco und Andrés Rinderknecht: Fossil evidence of frequency range of hearing independent of body size in South American Pleistocene ground sloths (Mammalia, Xenarthra). Comptes Rendus Palevol 11, 2012, S. 549–554
- Richard M. Fariña, Sergio F. Vizcaíno und Gerardo de Iuliis: Megafauna. Giant beasts of Pleistocene South America. Indiana University Press, Bloomington und Indianapolis, 2013, S. 1–436 (S. 259–260)
- Alberto Boscaini, François Pujos und Timothy J. Gaudin: A reappraisal of the phylogeny of Mylodontidae (Mammalia, Xenarthra) and the divergence of mylodontine and lestodontine sloths. Zoologica Scripta 48 (6), 2019, S. 691–710, doi:10.1111/zsc.12376
- Samantha Presslee, Graham J. Slater, François Pujos, Analía M. Forasiepi, Roman Fischer, Kelly Molloy, Meaghan Mackie, Jesper V. Olsen, Alejandro Kramarz, Matías Taglioretti, Fernando Scaglia, Maximiliano Lezcano, José Luis Lanata, John Southon, Robert Feranec, Jonathan Bloch, Adam Hajduk, Fabiana M. Martin, Rodolfo Salas Gismondi, Marcelo Reguero, Christian de Muizon, Alex Greenwood, Brian T. Chait, Kirsty Penkman, Matthew Collins und Ross D. E. MacPhee: Palaeoproteomics resolves sloth relationships. Nature Ecology & Evolution 3, 2019, S. 1121–1130, doi:10.1038/s41559-019-0909-z
- Luciano Varela, P. Sebastián Tambusso, H. Gregory McDonald und Richard A. Fariña: Phylogeny, Macroevolutionary Trends and Historical Biogeography of Sloths: Insights From a Bayesian Morphological Clock Analysis. Systematic Biology 68 (2), 2019, S. 204–218
- Frédéric Delsuc, Melanie Kuch, Gillian C. Gibb, Emil Karpinski, Dirk Hackenberger, Paul Szpak, Jorge G. Martínez, Jim I. Mead, H. Gregory McDonald, Ross D. E. MacPhee, Guillaume Billet, Lionel Hautier und Hendrik N. Poinar: Ancient mitogenomes reveal the evolutionary history and biogeography of sloths. Current Biology 29 (12), 2019, S. 2031–2042, doi:10.1016/j.cub.2019.05.043
- H. Gregory McDonald und Gerardo de Iuliis: Fossil history of sloths. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 39–55.
- Bruce J. Shockey und Federico Anaya: Grazing in a New Late Oligocene Mylodontid Sloth and a Mylodontid Radiation as a Component of the Eocene-Oligocene Faunal Turnover and the Early Spread of Grasslands/Savannas in South America. Journal of Mammalian Evolution 18, 2011, S. 101–115
- Timothy J. Gaudin: Phylogenetic relationships among sloths (Mammalia, Xenarthra, Tardigrada): the craniodental evidence. Zoological Journal of the Linnean Society 140, 2004, S. 255–305
- Ascanio D. Rincón, Andrés Solórzano, H. Gregory McDonald und Mónica Núñez Flores: Baraguatherium takumara, Gen. et Sp. Nov., the Earliest Mylodontoid Sloth (Early Miocene) from Northern South America. Journal of Mammalian Evolution 24 (2), 2017, S. 179–191
- Luciano Brambilla und Damián Alberto Ibarra: Archaeomylodon sampedrinensis, gen. et sp. nov., a new mylodontine from the middle Pleistocene of Pampean Region, Argentina. Journal of Vertebrate Paleontology 38 (6), 2018, S. e1542308, doi:10.1080/02724634.2018.1542308
- Andrés Rinderknecht, Enrique Bostelmann T., Daniel Perea und Gustavo Lecuona: A New Genus and Species of Mylodontidae (Mammalia: Xenarthra) from the Late Miocene of Southern Uruguay, with Comments on the Systematics of the Mylodontinae. Journal of Vertebrate Paleontology 30 (3), 2010, S. 899–910
- Francisco Ricardo Negri und Jorge Ferigolo: Urumacotheriinae, nova subfamília de Mylodontidae (Mammalia, Tardigrada) do Mioceno Superior-Plioceno, América do Sul. Revista Brasileira de Paleontologia 7 (2), 2004, S. 281–288
- Sergio F. Vizcaíno, Richard A. Fariña und Juan Carlos Fernicola: Young Darwin and the ecology and extinction of Pleistocene South American fossil mammals. Revista de la Asociación Geológica Argentina 64 (1), 2009, S. 160–169
- V. Pitana, A. M. Ribeiro und J. Ferigolo: New remains of Lestodon armatus Gervais 1855 (Tardigrada, Mylodontidae) from the Pleistocene of Rio Grande do Sul State, Brazil. Ameghiniana 48 (4; suppl.), 2011, S. R193–R194
- Omar J. Linares; Nuevos restos del genero Lestodon Gervais, 1855 (Xenarthra, Tardigrada, Mylodontidae), del Mioceno Tardío y Plioceno Temprano de Urumaco (Venezuela), con descripción de dos nuevas especies. Paleobiologia Neotropical 2, 2004, S. 1–14
- Alfredo A. Carlini, Gustavo J. Scillato-Yané und Rodolfo Sánchez; New Mylodontoidea (Xenarthra, Phyllophaga) from the middle Miocene–Pliocene of Venezuela. Journal of Systematic Palaeontology 4, 2006, S. 255–267
- Ascanio D. Rincón und H. Gregory McDonald: Reexamination of the Relationship of Pseudoprepotherium Hoffstetter, 1961, to the Mylodont Ground Sloths (Xenarthra) from the Miocene of Northern South America. Revista Geológica de América Central 63, 2020, S. 1–20, doi:10.15517/rgac.v62i0.41278
- Henrik Nikolai Krøyer: Forelobig notice om en ny Pattedyrslægt Platygnathus af Ordenen Edentata Cuv. Naturhistorisk Tidskrift 3, 1841, S. 589–594 ()
- Johannes Theodor Reinhardt: Bidrag til kundskab om kjaempedovendyret Lestodon armatus. Kongelige Danske Videnskabernes Selskabs Skrifter. Femte Række. Naturvidenskabelig og Mattematisk Afdeling 11, 1880, S. 1–39 ()
- Henri Gervais und Florentino Ameghino: Les mammifères fossiles de l´Ámérique du Sud. Paris-Buenos Aires, 1880, S. 1–225 (S. 160–161) ()
- Paul Gervais: Recherches sur les mammifères de l’Amérique méridionale. Annales des Sciences Naturelles, Zoologie 3 (6), 1855, S. 330–338 ()
- Wilhelm Dames: Skelettheile der Gattung Lestodon Gervais. Zeitschrift der Deutschen Geologischen Gesellschaft 34 (2), 1882, S. 816–817 ()
Weblinks
- American Museum of Natural History: Lestodon armatus.