Genetik
Die Genetik (moderne Wortschöpfung zu altgriechisch γενεά geneá „Abstammung“ und γένεσις génesis, deutsch ‚Ursprung‘)[1][2] oder Vererbungslehre (früher auch Erbbiologie) ist die Wissenschaft von der Vererbung und ein Teilgebiet der Biologie. Sie befasst sich mit den Gesetzmäßigkeiten und materiellen Grundlagen der Ausbildung von erblichen Merkmalen und der Weitergabe von Erbanlagen (Genen) an die nächste Generation.

Das Wissen, dass individuelle Merkmale über mehrere Generationen hinweg weitergegeben werden, ist relativ jung; Vorstellungen von solchen natürlichen Vererbungsprozessen prägten sich erst im 18. und frühen 19. Jahrhundert aus. Als Begründer der Genetik in diesem Sinn gilt der Augustinermönch Gregor Mendel, der in den Jahren 1856 bis 1865 im Garten seines Klosters systematisch Kreuzungsexperimente mit Erbsen durchführte und diese statistisch auswertete. So entdeckte er die später nach ihm benannten Mendelschen Regeln, die in der Wissenschaft allerdings erst im Jahr 1900 rezipiert und bestätigt wurden. Der heute weitaus bedeutendste Teilbereich der Genetik ist die Molekulargenetik, die sich mit den molekularen Grundlagen der Vererbung befasst. Aus ihr ging die Gentechnik hervor, in der die Erkenntnisse der Molekulargenetik praktisch angewendet werden.
Etymologie
Das Adjektiv „genetisch“ wurde schon um 1800 von Johann Wolfgang von Goethe in dessen Arbeiten zur Morphologie der Pflanzen und in der Folgezeit häufig in der romantischen Naturphilosophie sowie in der deskriptiven Embryologie verwendet.[3] Anders als heute meinte man damit eine Methode („genetische Methode“) der Untersuchung und Beschreibung der Individualentwicklung (Ontogenese) von Organismen. Das Substantiv „Genetik“ gebrauchte erstmals William Bateson 1905 zur Bezeichnung der neuen Forschungsdisziplin.
In Deutschland wurde bis in die zweite Hälfte des 20. Jahrhunderts der Ausdruck „Erbbiologie“ bedeutungsgleich gebraucht, zumeist zur Unterscheidung der „Erbbiologie des Menschen“ (Humangenetik) von der allgemeinen Genetik. Die Bezeichnung „Humangenetik“ war dabei in Deutschland bereits um 1940 etabliert. Damit wurde ein Rückzug auf wissenschaftlich gebotene Grundlagenforschung angezeigt, während „Rassenhygiene“ angewandte Wissenschaft darstellte.[4] Nach 1945 verschwanden die Bezeichnungen „Erbbiologie“ sowie „Rassenhygiene“ allmählich aus dem wissenschaftlichen Sprachgebrauch.
Teilbereiche
- Die von Mendel begründete klassische Genetik untersucht, in welchen Kombinationen die Gene bei Kreuzungsexperimenten bei den Nachkommen auftreten (Mendelsche Regeln) und wie das die Ausprägung bestimmter phänotypischer Merkmale beeinflusst. Zur klassischen Genetik gehört darüber hinaus die Zytogenetik, die im lichtmikroskopischen Größenbereich die Anzahl, Gestalt und Struktur der Chromosomen als Träger der genetischen Information untersucht.
- Die Molekulargenetik – ein Teilgebiet der Molekularbiologie – untersucht die molekularen Grundlagen der Vererbung: die Struktur der molekularen Träger der Erbinformation (gewöhnlich DNA, bei manchen Viren RNA), die Vervielfältigung dieser Makromoleküle (Replikation) und die dabei auftretenden Veränderungen des Informationsgehalts (Mutationen, Rekombination) sowie die Realisierung der Erbinformation im Zuge der Genexpression (Transkription und Translation). Zur Molekulargenetik gehört des Weiteren als angewandter Bereich die Gentechnik.
- Die Populationsgenetik und die Ökologische Genetik untersuchen genetische Strukturen und Prozesse auf der Ebene von Populationen und von anderen ökologischen Einheiten (z. B. ganzen Lebensgemeinschaften).
- Die Epigenetik beschäftigt sich mit der Weitergabe von Eigenschaften auf die Nachkommen, welche nicht auf Abweichungen in der DNA-Sequenz zurückgehen, sondern auf vererbbare Änderungen der Genregulation.
Geschichte
Zeittafel
- 1866 – Gregor Mendel veröffentlichte seine Versuche über Pflanzen-Hybriden, die aber kaum beachtet wurden.
- 1869 – Friedrich Miescher isolierte aus Zellkernen das „Nuclein“, dessen Aufbau und Funktion zunächst rätselhaft blieb.
- 1889 – Richard Altmann identifizierte die „Nucleinsäure“ und eine basische Proteinfraktion als Bestandteile des Nucleins.
- 1900 – Hugo de Vries, Carl Correns und Erich Tschermak bestätigten Mendels Entdeckungen.
- 1903 – Chromosomen wurden als Träger der Erbinformation erkannt (Walter Sutton).
- 1906 – William Bateson prägte den Begriff Genetik.
- 1907 – Thomas Hunt Morgan wählte die Taufliege Drosophila melanogaster als Versuchstier.
- 1909 – Wilhelm Johannsen prägte den Begriff Gen.
- 1927 – Auslösung künstlicher Mutationen durch Röntgenstrahlung (Hermann Joseph Muller)
- 1928 – Erste Beschreibung der Transformation durch Frederick Griffith (Griffiths Experiment)
- 1931 – Zytologische Aufklärung des Crossing-over (Barbara McClintock, Harriet B. Creighton, Curt Stern)
- 1940 – George Beadle und Edward Tatum formulierten die Ein-Gen-ein-Enzym-Hypothese.
- 1943 – Das Luria-Delbrück-Experiment belegte, dass Mutationen in dem Sinn zufällig sind, dass sie keine Umweltreaktionen darstellen.
- 1944 – Oswald Avery, Colin MacLeod und Maclyn McCarty: Transformation von Bakterien durch DNA
- 1950 – Erwin Chargaff zeigte mit den Chargaff-Regeln, dass die vier Nukleotide in paarweise gleicher Häufigkeit in der DNA vorkommen: [A] = [T] und [C] = [G].
- 1951 – McClintock berichtete erstmals über springende Gene, stieß aber auf komplettes Unverständnis.
- 1952 – Das Hershey-Chase-Experiment zeigte, dass die genetische Information von Bakteriophagen in der DNA gespeichert ist.
- 1953 – James Watson und Francis Crick postulierten die Doppelhelix-Struktur der DNA.
- 1957 – Nachweis der semikonservativen Replikation der DNA und des Crossing-over durch James Herbert Taylor (Taylor-Experiment)
- 1958 – Nachweis der semikonservativen Replikation der DNA durch Meselson und Stahl
- 1958 – Crick postulierte das „Zentrale Dogma“ der Molekulargenetik.
- 1961 – François Jacob und Jacques Monod stellten das Operon-Konzept vor
- 1961 bis 1965 – Dechiffrierung des genetischen Codes (Marshall Warren Nirenberg und Heinrich Matthaei)
- 1969 – Jonathan Beckwith gelang als erstem die Isolierung eines einzelnen Gens (aus E. coli).
- 1969 – Werner Arber, Daniel Nathans und Hamilton Othanel Smith entdeckten die Restriktionsenzyme.
- 1975 – DNA-Sequenzierung (Frederick Sanger, Allan Maxam, Walter Gilbert)
- 1977 – Intron-Exon-Struktur eukaryotischer Gene
- 1983 – Kary Mullis entwickelte die Polymerase-Kettenreaktion (PCR) zur Vervielfältigung von DNA.
- 1995 – Das erste prokaryotische Genom (von Haemophilus influenzae) wurde sequenziert.
- 1997 – Das erste eukaryotische Genom, das der Bäckerhefe Saccharomyces cerevisiae, ist sequenziert
- 2003 – Als Resultat des Humangenomprojektes steht die Referenzsequenz des menschlichen Genoms zum Download im Internet bereit
Vorgeschichte
Schon in der Antike versuchten Menschen die Gesetzmäßigkeiten der Zeugung und die Ähnlichkeiten zwischen Verwandten zu erklären, und einige der im antiken Griechenland entwickelten Konzepte blieben bis in die Neuzeit gültig oder wurden in der Neuzeit wieder aufgegriffen.[5][6] So lehrte der griechische Philosoph Alkmaion um 500 v. Chr., dass die Zeugung der Nachkommen durch die Zusammenwirkung des männlichen und des weiblichen „Samens“ geschehe. Sein Postulat eines weiblichen Samens fand in der damaligen Naturphilosophie und später auch in der hippokratischen Medizin allgemeine Anerkennung. Davon abweichend behaupteten Hippon und Anaxagoras, dass nur der Mann zeugungsfähigen Samen bilde und dass der weibliche Organismus den Keim nur ernähre. Die Bildung des Samens erfolgte laut Alkmaion im Gehirn, von wo aus er durch die Adern in den Hoden gelange. Demgegenüber erklärten Anaxagoras und Demokrit, dass der gesamte Organismus zur Bildung des Samens beitrage – eine Ansicht, die als Pangenesistheorie über 2000 Jahre später von Charles Darwin erneut vertreten wurde. Auch die Überlegungen des Anaxagoras, wonach alle Körperteile des Kindes bereits im Samen (Sperma) vorgebildet seien, traten als Präformationslehre in der Neuzeit wieder auf. In der Antike wurden diese frühen Lehren weitgehend abgelöst durch die Ansichten des Aristoteles (De generatione animalium), wonach das Sperma aus dem Blut entsteht und bei der Zeugung nur immateriell wirkt, indem es Form und Bewegung auf die durch den weiblichen Organismus bereitgestellte flüssige Materie überträgt.[7] Die Entwicklung des Keims beschrieb Aristoteles als Epigenese, wonach im Gegensatz zur Präformation die verschiedenen Organe nacheinander durch die Einwirkung des väterlichen Formprinzips ausgebildet werden. Neben der geschlechtlichen Zeugung kannte Aristoteles auch die Parthenogenese (Jungfernzeugung) sowie die (vermeintliche) Urzeugung von Insekten aus faulenden Stoffen.
Der Aristoteles-Schüler Theophrastus postulierte eine transmutatio frumentorum und nahm an, dass sich Getreidearten zu ihrer Wildform zurückverwandeln können. Zudem unterschied er männliche und weibliche Pflanzen bei der Dattelpalme.[8]
Vererbung war bis in das 18. Jahrhundert ein juristischer Begriff und fand für natürliche Vorgänge keine Anwendung. Denn Ähnlichkeiten zwischen Verwandten wurden ausreichend über jeweils spezifische lokale Faktoren und die Lebensweise des Individuums erklärt: über das Klima, die Ernährung, die Art der Betätigungen usw. Wie gewisse Merkmale unter Nachkommen blieben auch diese Faktoren für die Nachkommen in der Regel konstant. Irreguläre Merkmale konnten dann entsprechend auf irreguläre Einflüsse bei der Zeugung oder der Entwicklung des Individuums zurückgeführt werden. Erst mit dem zunehmenden internationalen Verkehr und zum Beispiel der Anlage von exotischen Gärten wurde ein Wahrnehmungsraum dafür geschaffen, dass es vom Individuum und seinem jeweiligen Ort ablösbare, natürliche Gesetze geben müsse, die sowohl die Weitergabe von regulären als auch zuweilen eine Weitergabe von neu erworbenen Eigenschaften regeln.[9]
Der Begriff der Fortpflanzung oder Reproduktion, in dessen Kontext von Vererbung im biologischen Sinn gesprochen werden kann, kam erst gegen Ende des 18. Jahrhunderts auf.[10] In früheren Jahrhunderten galt die „Zeugung“ eines Lebewesens als ein Schöpfungsakt, der grundsätzlich eines göttlichen Eingriffs bedurfte und im Rahmen des Präformismus vielfach als Teilaspekt der Erschaffung der Welt betrachtet wurde. Dabei unterschied man die Zeugung durch den Samen (Sperma) im Mutterleib von der Urzeugung, durch welche niedere Tiere (etwa Würmer, Insekten, Schlangen und Mäuse) aus toter Materie hervorzugehen schienen.[11] Die „Samenzeugung“ betrachtete man als Eigenheit des Menschen und der höheren Tiere, welche zu ihrer Ausbildung eines Mutterleibs bedürfen. Erst gegen Ende des 17. Jahrhunderts setzte sich, vor allem aufgrund der Experimente Francesco Redis, die Einsicht durch, dass Würmer, Insekten und andere niedere Tiere nicht aus toter Materie entstehen, sondern von gleichartigen Tieren gezeugt werden. Nun betrachtete man die Zeugung nicht mehr als Schöpfungsakt, sondern verlegte diesen in die Zeit der Erschaffung der Welt, bei der, wie man annahm, alle zukünftigen Generationen von Lebewesen zugleich ineinandergeschachtelt erschaffen wurden. Die Zeugung war somit nur noch eine Aktivierung des längst vorhandenen Keims, der sich dann zu einem voll ausgebildeten Organismus entfaltete. Strittig war dabei, ob die Keime durch das weibliche oder durch das männliche Geschlecht weitergegeben werden, ob sie also im Ei oder im „Samentierchen“ eingeschachtelt sind. Beide Ansichten hatten ihre Anhänger (Ovisten und Animalkulisten), bis die Entdeckung der Jungfernzeugung bei der Blattlaus durch Charles Bonnet 1740 den Streit zugunsten der Ovisten entschied.[12]
Neben der sehr populären Präformationslehre, die 1625 durch Giuseppe degli Aromatari (1587–1660) ins Spiel gebracht worden war, gab es im 17. Jahrhundert auch renommierte Anhänger der an Aristoteles anknüpfenden Epigenesislehre, namentlich William Harvey und René Descartes. Deren Ansichten galten jedoch als antiquiert und wurden als unwissenschaftlich verworfen, da sie immaterielle Wirkprinzipien voraussetzten, während der Präformismus rein mechanistisch gedacht werden konnte und zudem durch die Einführung des Mikroskops einen starken Auftrieb erfuhr.[13]
Die Vorstellung der Präformation herrschte bis in das 19. Jahrhundert hinein vor, obwohl es durchaus Forschungsergebnisse gab, die nicht mit ihr in Einklang gebracht werden konnten. Großes Erstaunen riefen die Versuche zur Regeneration bei Salamandern, Süßwasserpolypen und anderen Tieren hervor. Polypen kann man fein zerhacken, und jedes Teilstück entwickelt sich, wie Abraham Trembley 1744 beschrieb, innerhalb von zwei bis drei Wochen zu einem kompletten Tier. In den Jahren 1744 bis 1754 veröffentlichte Pierre-Louis Moreau de Maupertuis mehrere Schriften, in denen er aufgrund von Beobachtungen bei Tieren und Menschen, wonach beide Eltern Merkmale an ihre Nachkommen weitergeben können, die Präformationslehre kritisierte und ablehnte. Entsprechende Beobachtungen publizierte auch Joseph Gottlieb Kölreuter (1761), der als Erster Kreuzungen verschiedener Pflanzenarten studierte. Und Caspar Friedrich Wolff beschrieb 1759 minutiös die Entwicklung des Embryos im Hühnerei aus völlig undifferenzierter Materie. Trotz der Probleme, die derartige Forschungen aufwarfen, geriet die Präformationslehre jedoch erst im frühen 19. Jahrhundert durch die embryologischen Untersuchungen von Christian Heinrich Pander (1817) und Karl Ernst von Baer (1828) ins Wanken, bei denen diese die Bedeutung der Keimblätter aufklärten und allgemein gültige Gesetzmäßigkeiten der Embryogenese der Tiere aufzeigten.[14]
Mit der Etablierung der von Matthias Jacob Schleiden (1838), Theodor Schwann (1839) und Rudolf Virchow (1858) entwickelten Allgemeinen Zelltheorie wurde deutlich, dass die Gründe für die Ähnlichkeit von Eltern und Nachkommen in der Zelle lokalisiert sein müssen. Alle Organismen bestehen aus Zellen, Wachstum beruht auf der Vermehrung der Zellen durch Teilung, und bei der geschlechtlichen Fortpflanzung, die bei Vielzellern der Normalfall ist, vereinigen sich je eine Keimzelle beiderlei Geschlechts zu einer Zygote, aus welcher durch fortwährende Teilung und Differenzierung der neue Organismus hervorgeht.[15]
Klassische Genetik
Die Gesetzmäßigkeiten der Vererbung blieben lange im Unklaren. Schon in den Jahren 1799 bis 1823 führte Thomas Andrew Knight – wie einige Jahrzehnte später Gregor Mendel – Kreuzungsexperimente mit Erbsen durch, bei denen er bereits die Erscheinungen der Dominanz und der Aufspaltung von Merkmalen beobachtete.[16] 1863 publizierte Charles Victor Naudin (1815–1899) die Ergebnisse seiner Kreuzungsexperimente mit zahlreichen Pflanzengattungen, wobei er das sehr gleichartige Aussehen aller Pflanzen der ersten Tochtergeneration und die „extreme Verschiedenartigkeit der Formen“ in den folgenden Generationen konstatierte und damit weitere bedeutende Aspekte der fast zeitgleichen Erkenntnisse Mendels vorwegnahm, aber im Unterschied zu Mendel keine statistische Auswertung durchführte.[17]

Der entscheidende Durchbruch gelang dann Mendel mit seinen 1856 begonnenen Kreuzungsversuchen, bei denen er sich auf einzelne Merkmale konzentrierte und die erhaltenen Daten statistisch auswertete. So konnte er die grundlegenden Gesetzmäßigkeiten bei der Verteilung von Erbanlagen auf die Nachkommen ermitteln, die heute als Mendelsche Regeln bezeichnet werden. Diese Entdeckungen, die er 1866 publizierte, blieben jedoch zunächst in der Fachwelt fast unbeachtet und wurden erst im Jahr 1900 von Hugo de Vries, Carl Correns und Erich Tschermak wiederentdeckt und aufgrund eigener Versuche bestätigt.
Einen radikalen Umbruch der Vorstellungen von der Vererbung brachte die Keimbahn- oder Keimplasmatheorie mit sich, die August Weismann in den 1880er Jahren entwickelte.[18] Schon seit dem Altertum galt es als selbstverständlich, dass Merkmale, welche die Eltern während ihres Lebens erworben haben, auf die Nachkommen übertragen werden können. Nach Jean-Baptiste de Lamarck, in dessen Evolutionstheorie sie eine bedeutende Rolle spielte, wird diese Ansicht heute als Lamarckismus bezeichnet. Doch auch Charles Darwin postulierte in seiner Pangenesistheorie, dass der ganze elterliche Organismus auf die Keimzellen einwirke – unter anderem sogar indirekt durch Telegonie. Weismann unterschied nun zwischen der Keimbahn, auf der die Keimzellen eines Organismus sich von der Zygote herleiten, und dem Soma als der Gesamtheit aller übrigen Zellen, aus denen keine Keimzellen hervorgehen können und von denen auch keine Einwirkungen auf die Keimbahn ausgehen. Diese Theorie war allerdings anfangs sehr umstritten.[19]

Mit seinem zweibändigen Werk Die Mutationstheorie (1901/03) führte de Vries den bis dahin in der Paläontologie gebräuchlichen Begriff „Mutation“ in die Vererbungslehre ein. Nach seiner Auffassung handelte es sich bei Mutationen um umfassende, sprunghafte Veränderungen, durch welche eine neue Art entstehe. Dabei stützte er sich auf seine Studien an Nachtkerzen, bei denen eine „in allen ihren Organen“ stark veränderte Pflanze aufgetreten war, deren Merkmale sich als erbkonstant erwiesen und die er daher als neue Art (Oenothera gigas) beschrieb. (Später stellte sich heraus, dass Oe. gigas im Unterschied zu den diploiden Ausgangspflanzen tetraploid war und somit – aus heutiger Sicht – der Sonderfall einer Genommutation (Autopolyploidie) vorlag.) Dieser Befund stand im Widerspruch zu der an Charles Darwin anschließenden Evolutionstheorie, die das Auftreten geringfügiger Veränderungen voraussetzte, und das war einer der Gründe, warum der „Mendelismus“ sich zeitweilig im Widerstreit mit dem damals noch nicht allgemein akzeptierten Darwinismus befand.
In den Jahren um die Jahrhundertwende untersuchten etliche Forscher die unterschiedlichen Formen der Chromosomen und deren Verhalten bei Zellteilungen. Aufgrund der Beobachtung, dass gleich aussehende Chromosomen paarweise auftreten, äußerte Walter Sutton 1902 als erster die Vermutung, dass dies etwas mit den ebenfalls gepaarten Merkmalen und deren „Spaltung“ in den Untersuchungen von Mendel und seinen Wiederentdeckern zu tun haben könne.[20] Im Anschluss daran formulierte Theodor Boveri 1904 die Chromosomentheorie der Vererbung, wonach die Erbanlagen an die Chromosomen gebunden sind und deren Verhalten bei der Meiose und Befruchtung den Mendelschen Regeln entspricht.[21]
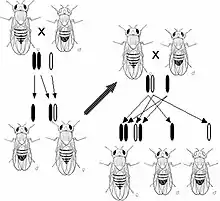
Eine sehr folgenreiche Entscheidung war die Wahl von Taufliegen als Versuchsobjekt durch die Arbeitsgruppe um Thomas Hunt Morgan im Jahre 1907, vor allem weil diese in großer Zahl auf kleinem Raum gehalten werden können und sich sehr viel schneller vermehren als die bis dahin verwendeten Pflanzen. So stellte sich bald heraus, dass es auch geringfügige Mutationen gibt, auf deren Grundlage allmähliche Veränderungen innerhalb von Populationen möglich sind (Morgan: For Darwin, 1909). Eine weitere wichtige Entdeckung machte Morgans Team etwa 1911, als man die schon 1900 von Correns publizierte Beobachtung, dass manche Merkmale meist zusammen vererbt werden (Genkopplung), mit Untersuchungen der Chromosomen verband und so zu dem Schluss kam, dass es sich bei den Koppelungsgruppen um Gruppen von Genen handelt, welche auf demselben Chromosom liegen. Wie sich weiter herausstellte, kann es zu einem Austausch von Genen zwischen homologen Chromosomen kommen (Crossing-over), und aufgrund der relativen Häufigkeiten dieser intrachromosomalen Rekombinationen konnte man eine lineare Anordnung der Gene auf einem Chromosom ableiten (Genkarte). Diese Erkenntnisse fasste Morgan 1921 in The Physical Basis of Heredity und 1926 programmatisch in The Theory of the Gene zusammen, worin er die Chromosomentheorie zur Gentheorie weiterentwickelte.
Diese Theorie war schon während ihrer allmählichen Herausbildung sehr umstritten. Ein zentraler Streitpunkt war die Frage, ob die Erbanlagen sich ausschließlich im Zellkern oder auch im Zytoplasma befinden. Vertreter der letzteren Ansicht waren u. a. Boveri, Correns, Hans Driesch, Jacques Loeb und Richard Goldschmidt. Sie postulierten, dass im Kern nur relativ geringfügige Erbfaktoren bis hin zu Artmerkmalen lokalisiert seien, während Merkmale höherer systematischer Kategorien (Gattung, Familie usw.) durch das Plasma vererbt würden. Der entschiedenste Vertreter der Gegenseite war Morgans ehemaliger Mitarbeiter Hermann Joseph Muller, der in The Gene as the Basis of Life (1929) die im Kern lokalisierten Gene als die Grundlage des Lebens überhaupt bezeichnete und die Bedeutung des Plasmas als sekundär einstufte.
Muller war es auch, der 1927 erstmals von der Erzeugung von Mutationen durch Röntgenstrahlung berichtete, wodurch die genetische Forschung nicht mehr darauf angewiesen war, auf spontan auftretende Mutationen zu warten. Der von de Vries, Morgan, Muller und Anderen vertretenen Ansicht der Zufälligkeit der Mutationen stand das u. a. von Paul Kammerer und Trofim Denissowitsch Lyssenko verfochtene Postulat gegenüber, dass Mutationen „gerichtet“ und qualitativ durch Umwelteinflüsse bestimmt seien.
Populationsgenetik
Nach dem allgemeinen Bekanntwerden von Mendels mathematisch exakter Beschreibung des dominant-rezessiven Erbgangs im Jahr 1900 wurde die Frage diskutiert, ob rezessive Merkmale in natürlichen Populationen allmählich verschwinden oder auf Dauer erhalten bleiben.[22] Hierzu fanden der deutsche Arzt Wilhelm Weinberg und der britische Mathematiker Godfrey Harold Hardy 1908 fast gleichzeitig eine Formel, die das Gleichgewicht dominanter und rezessiver Merkmale in Populationen beschreibt. Diese Entdeckung wurde jedoch unter Genetikern zunächst kaum beachtet. Erst 1917 führte Reginald Punnett das von ihm so genannte „Hardy-Gesetz“ in die Populationsforschung ein, was ein wichtiger Beitrag zur Begründung der Populationsgenetik als eigenständigem Forschungszweig in den 1920er Jahren war. Weinbergs Beitrag wurde sogar erst 1943 von Curt Stern wiederentdeckt, der die Formel daraufhin in „Hardy-Weinberg-Gesetz“ umbenannte.
Die Grundlagen der Populationsgenetik wurden parallel von Sewall Wright, Ronald A. Fisher und J. B. S. Haldane entwickelt.[23] Sie erkannten, dass Vererbungsvorgänge in der Natur sinnvollerweise auf der Ebene von Populationen zu betrachten sind, und formulierten dafür die theoretischen Grundlagen (Haldane: A Mathematical Theory of Natural and Artificial Selection. 1924–1932; Fisher: The Genetical Theory of Natural Selection. 1930; Wright: Evolution in Mendelian Populations. 1931).
Die Erbsubstanz
Seit 1889 (Richard Altmann) war bekannt, dass Chromosomen aus „Nucleinsäure“ und basischem Protein bestehen. Über deren Aufbau und Funktion konnte jedoch lange Zeit nur spekuliert werden. 1902 postulierten Emil Fischer und Franz Hofmeister, dass Proteine Polypeptide seien, also lange Ketten von Aminosäuren. Das war zu diesem Zeitpunkt allerdings noch sehr spekulativ. Als 1905 die ersten Analysen der Aminosäuren-Zusammensetzung von Proteinen publiziert wurden, erfassten diese lediglich ein Fünftel des untersuchten Proteins, und die Identifikation aller 20 proteinogenen Aminosäuren zog sich bis 1935 hin. Dagegen war bei der Nukleinsäure schon 1903 klar (Albrecht Kossel), dass sie neben Zucker und Phosphat lediglich fünf verschiedene Nukleinbasen enthält. Erste Analysen der Basenzusammensetzung durch Hermann Steudel ergaben 1906, dass die vier hauptsächlich vorhandenen Basen zu annähernd gleichen Anteilen enthalten sind. Daraus schloss Steudel (1907), dass die Nukleinsäure „ein relativ einfach gebauter Körper sei“,[24] dem man keine anspruchsvollen Funktionen beimessen könne. Dies etablierte sich als Lehrmeinung, die bis in die 1940er Jahre gültig blieb, und auf dieser Grundlage betrachtete man nicht die Nukleinsäure(n), sondern die Proteine als „Erbsubstanz“.
Zu der Einsicht, dass es sich gerade umgekehrt verhält und die Nukleinsäure DNA als Erbsubstanz angesehen werden muss, führten die Experimente der Arbeitsgruppe von Oswald Avery zur Transformation von Pneumokokken (1944)[25] und das Hershey-Chase-Experiment von 1952 mit Bakteriophagen. Außerdem zeigte Erwin Chargaff 1950, dass die vier Nukleotide, aus denen die DNA besteht, nicht zu gleichen, sondern zu paarweise gleichen Anteilen enthalten sind. Zusammen mit Röntgenstrukturanalyse-Daten von Rosalind Franklin war das die Grundlage für die Entwicklung des Doppelhelix-Strukturmodells der DNA durch James Watson und Francis Crick 1953.
Siehe auch
Literatur
- François Jacob: La logique du vivant: Une histoire de l'hérédité. Gallimard, Paris 1971. (deutsch: Die Logik des Lebenden. Fischer, Frankfurt am Main 1972, Neuausgabe 2002)
- Wilfried Janning, Elisabeth Knust: Genetik. 2. Auflage. Thieme, Stuttgart 2008, ISBN 978-3-13-149801-4.
- William S. Klug, Michael R. Cummings, Charlotte A. Spencer: Genetik. 8. Auflage. Pearson Studium, München 2007, ISBN 978-3-8273-7247-5.
- Hans-Peter Kröner: Genetik. In: Werner E. Gerabek, Bernhard D. Haage, Gundolf Keil, Wolfgang Wegner (Hrsg.): Enzyklopädie Medizingeschichte. de Gruyter, Berlin/ New York 2005, ISBN 3-11-015714-4, S. 468–475.
- Katharina Munk (Hrsg.): Taschenlehrbuch Biologie: Genetik. Thieme, Stuttgart 2010, ISBN 978-3-13-144871-2.
- Eberhard Passarge: Color atlas of genetics Taschenatlas der Genetik, Georg Thieme Verlag KG, Stuttgart 2018, 5. Auflage, ISBN 978-3-13-241440-2.
- Hans-Jörg Rheinberger, Staffan Müller-Wille: Vererbung – Geschichte und Kultur eines biologischen Konzepts. Fischer, Frankfurt am Main 2009, ISBN 978-3-596-17063-0.
Weblinks
- Die Geschichte der DNA. (Flash) (Nicht mehr online verfügbar.) In: schule-bw.de. Ehemals im Original (deutsche Fassung von „DNA from the Beginning“ des Dolan DNA Learning Center). (Seite nicht mehr abrufbar, Suche in Webarchiven)
- Genetic Science Learning Center – University of Utah (engl.), Gewinner des Science Prize for Online Resources in Education.[26]
- Michael Stang: Datenschutz – Gehackte Gene. In: Deutschlandfunk-Sendung Wissenschaft im Brennpunkt. 3. Oktober 2014.
Einzelnachweise
- genetikós. In: Henry George Liddell, Robert Scott: A Greek-English Lexicon. (perseus.tufts.edu).
- génesis. In: Henry George Liddell, Robert Scott: A Greek-English Lexicon. (perseus.tufts.edu).
- Ilse Jahn, Rolf Löther, Konrad Senglaub (Hrsg.): Geschichte der Biologie. 2. Auflage. Gustav Fischer Verlag, Jena 1985, S. 284 und 413.
- Peter Weingart, Jürgen Kroll, Kurt Bayertz: Rasse, Blut und Gene. Geschichte der Eugenik und Rassenhygiene in Deutschland. Suhrkamp, Frankfurt am Main 1992, S. 557 f.
- I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 56–59.
- Erna Lesky: Die Zeugungs- und Vererbungslehren der Antike und ihr Nachwirken. (= Akademie der Wissenschaften und der Literatur [zu Mainz]: Abhandlungen der geistes- und sozialwissenschaftlichen Klasse. [1950]. 19). Wiesbaden 1951, DNB 453020739.
- I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 68–71.
- Hans-Peter Kröner: Genetik. 2005, S. 468.
- Hans-Jörg Rheinberger, Staffan Müller-Wille: Vererbung. Geschichte und Kultur eines biologischen Konzepts. Frankfurt am Main 2009.
- François Jacob: Die Logik des Lebenden – Von der Urzeugung zum genetischen Code. Frankfurt am Main 1972, S. 27 f.
- F. Jacob: Die Logik des Lebenden – Von der Urzeugung zum genetischen Code. 1972, S. 32 f.
- F. Jacob: Die Logik des Lebenden – Von der Urzeugung zum genetischen Code. 1972, S. 72.
- I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 218–220 und 231.
- F. Jacob: Die Logik des Lebenden – Von der Urzeugung zum genetischen Code. 1972, S. 74–79; I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 232–249.
- F. Jacob: Die Logik des Lebenden – Von der Urzeugung zum genetischen Code. 1972, S. 123–139.
- I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 417 und 691.
- I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 418 f.
- F. Jacob: Die Logik des Lebenden – Von der Urzeugung zum genetischen Code. 1972, S. 232–235.
- I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 410–412.
- I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 463.
- I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 463 f.
- I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 468 f.
- I. Jahn u. a. (Hrsg.): Geschichte der Biologie. 1985, S. 482–484.
- H. Steudel: Hoppe-Seyler's Z. Physiol. Chem. 53, 1907, S. 18, zitiert nach Peter Karlson: 100 Jahre Biochemie im Spiegel von Hoppe-Seyler’s Zeitschrift für Physiologische Chemie. dito Band 358, 1977, S. 717–752, Zitat S. 747.
- Oswald T. Avery u. a.: Studies on the chemical nature of the substance inducing transformation of pneumococcal types. Inductions of transformation by a desoxyribonucleic acid fraction isolated from pneumococcus type III. In: J Exp Med. Band 79, Nr. 2, 1944, S. 137–158.
- Louisa A. Stark, Kevin Pompei: Making Genetics Easy to Understand. In: Science. Band 327, Nr. 5965, S. 538–539, doi:10.1126/science.1183029.