Paramylodon
Paramylodon ist eine ausgestorbene Gattung der Faultiere und gehört zur ebenfalls erloschenen Familie der Mylodontidae. Es war ein großes, bodenlebendes Faultier, das hauptsächlich im Pleistozän vor rund 1,8 Millionen Jahren bis vor etwa 11.000 Jahren in Nordamerika lebte. Die Gattung ist über zahlreiche Fossilfunde belegt, ihre Anzahl nahm in der Spätphase des Auftretens sichtlich zu. Sie stammen überwiegend aus dem Süden der heutigen USA, im westlichen Bereich streuen sie aber auch bis in den Süden von Kanada. Einen herausragenden Fundplatz stellen die Asphaltgruben von Rancho La Brea bei Los Angeles in Kalifornien dar, wo die Gattung recht häufig belegt ist. Anhand der Fossilreste lässt sich eine Länge der Tiere von knapp 280 cm und ein Gewicht von gut 1,4 t rekonstruieren.
| Paramylodon | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Paramylodon | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberes Pliozän bis Oberes Pleistozän | ||||||||||||
| 2,1? oder 1,8 Mio. Jahre bis 11.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Paramylodon | ||||||||||||
| Brown, 1903 | ||||||||||||
Aufgrund der teils sehr guten Fossilerhaltung ist bei den Tieren die für die Mylodonten typische Körperpanzerung in Form von Osteodermen überliefert. Zudem lässt sich ein geschlechtsspezifischer Unterschied im Schädelbau nachweisen. Den Funden zufolge lebten die Vertreter von Paramylodon in offenen Landschaften, teilweise auch in gebirgigen Lagen und ernährten sich höchstwahrscheinlich grasfresserisch oder von gemischter Pflanzenkost. Einmalige Spurenfossilien geben darüber hinaus Auskunft über die Fortbewegung der Tiere, die vierfüßig erfolgte. Zudem kann aufgrund der Gestaltung der Vordergliedmaßen auch eine gewisse grabende Lebensweise nicht ausgeschlossen werden.
Innerhalb der Gattung ist mit Paramylodon harlani nur eine Art anerkannt. Die ersten Fossilfunde stammen bereits aus den Beginn der 1830er Jahre. Sie gehen auf Richard Harlan zurück, zu dessen Ehre die Art benannt wurde. Die Gattung Paramylodon führte Barnum Brown zu Beginn des 20. Jahrhunderts ein. Insgesamt blickt der Faultiervertreter auf eine wechselvolle Forschungsgeschichte zurück. In deren ersten 150 Jahren wurden die heute Paramylodon zugeschriebenen Funde, später die Gattung selbst, immer wieder mit anderen Formen in Verbindung gebracht, zuerst mit Mylodon, seit den 1950er Jahren aber zunehmend mit Glossotherium. Vor allem mit letzterer Gattung verbindet Paramylodon zahlreiche Merkmale, die für eine enge Verwandtschaft sprechen. Erst seit den 1990er Jahren gelten beide Gattungen als eigenständig, wobei Glossotherium auf Südamerika beschränkt ist, während Paramylodon Nordamerika bewohnte.
Merkmale
Körpergröße
Paramylodon ist vor allem aufgrund der Funde aus Rancho La Brea in Kalifornien in reichhaltiger Zahl überliefert. Das dort geborgene Material von mehreren Dutzend Individuen diente als Basis zahlreicher Untersuchungen, auf die sich die folgenden beschreibenden Angaben weitgehend berufen.[1][2][3] Die Faultiergattung war ein mittelgroßer Vertreter der Mylodontidae. Ein vollständig rekonstruiertes Skelett aus Rancho La Brea besitzt eine Gesamtlänge von 279 cm, wovon der Schwanz etwa 118 cm einnimmt. An den Schultern erreicht es eine Höhe von 112 cm, am Becken misst es 122 cm. Das Gewicht für diese späten Angehörigen des Oberen Pleistozäns wird mit rund 1,39 t angegeben, frühere Formen waren aber durchaus kleiner.[4] Insgesamt stellte Paramylodon ein robust gebautes Tier dar. Es zeichnete sich durch einen langgestreckten Schädel, einen kurzen Nacken, einen kurzen und kompakten Körper mit breitem Becken sowie kräftigen Gliedmaßen und Schwanz aus.[5]
Schädel- und Gebissmerkmale
_5_(15444177835).jpg.webp)
Der Schädel von Paramylodon war langschmal. Er erreichte eine Gesamtlänge vom 42,9 bis 49,8 cm, ermittelt an etwa einem Dutzend Exemplaren.[6] Ein besonders großer Schädel maß 54,0 cm.[7] In der Aufsicht besaß er eine eher rechteckige Form mit einer durchschnittlichen Breite am Hinterhauptsbein von 18,8 cm, hinter den Augen von 12,2 cm und an der Schnauze von 14 cm. Typisch für zahlreiche Mylodonten war die sich nach vorn kontinuierlich verbreiternde Schnauze. Der Schädel zeigte sich aber insgesamt als deutlich schmaler als beim vergleichbar großen Glossotherium, letzteres wies in Seitenansicht an der Stirnlinie eine domartige Aufwölbung auf, die bei Paramylodon nicht auftrat. Allerdings war der Schädel von Paramylodon mit Ausnahme des mittleren Bereichs (der domartigen Aufwölbung bei Glossotherium) durchschnittlich höher, so maß er am Hinterhaupt rund 13,8 cm, an der Schnauze 13 cm. Das Nasenbein befand sich seitlich im Kontakt mit dem Oberkiefer. Dadurch entstand ein seitlich geschlossener und nur nach vorn geöffneter Naseninnenraum, der etwa so hoch wie breit wurde, was dem insgesamt schmaleren Schädel geschuldet war. Der Mittelkieferknochen war, typisch für die Faultiere, nur locker mit dem Oberkiefer verbunden. Am Stirnbein ragte das Nasenbein weit nach hinten, sodass die Naht zwischen den beiden Schädelknochen eher V-förmig verlief. Zudem stellte das Stirnbein den größten Knochen des gesamten Schädels dar. Zwischen den Scheitelbeinen bestand ein kräftiger Scheitelkamm, der jedoch wesentlich schmaler erschien als im Vergleich zu Glossotherium. Die Jochbögen waren abweichend von den meisten Faultieren sekundär wieder geschlossen. Der vordere, vom Jochbein ausgehende und nach hinten zeigende Bogenteil wies drei Fortsätze auf, von denen einer nach oben, einer nach unten und der mittlere horizontal orientiert war. Der hintere, am Schläfenbein ansetzende Bogenabschnitt besaß eine fingerförmige Gestalt und verband sich mit dem mittleren Fortsatz des vorderen Bogenabschnitts. Auf der Schädelunterseite ragte das Gaumenbein bei Paramylodon deutlich weiter nach hinten als bei Glossotherium, was durch die längere Ausdehnung des Knochens hinter dem letzten Backenzahn verursacht wurde. Wie bei vielen Mylodonten waren die beiden Flanken des Flügelbeins deutlich aufgebläht. Bei Paramylodon zeigte sich dies aber nicht ganz so deutlich wie bei Glossotherium, sodass die aufgewölbten Strukturen wesentlich weiter durch das Basisphenoid des Keilbeins auseinanderstanden.[1][3][6]
Der Unterkiefer erreichte nach Analyse von gut zwei Dutzend Objekten Längen von 31,5 bis 43,6 cm. Er war massiv gebaut und breit. Der horizontale Knochenkörper nahm kontinuierlich von vorn nach hinten an Höhe zu, unter dem hintersten Zahn betrug seine Höhe bis zu 10,5 cm.[8] Die robuste Symphyse wurde bis zu 11 cm lang und war – typisch für Mylodonten – breit. Sie zog nach vorne aus, was ein Kennzeichen fast aller Faultiere ist. Diese löffelartige Verlängerung der Symphyse kragte bei Paramylodon seitlich nicht so deutlich auseinander wie bei Glossotherium, sodass die Seitenkanten eher gerade verliefen und weniger deutlich geschwungen waren als bei Letzterem. Die Breite der Symphyse im vorderen Bereich betrug bis zu 15 cm. Der Gelenkfortsatz überragte die Kauebene nur unwesentlich, der Kronenfortsatz war deutlich höher. Seine Vorderkante verlief bei Paramylodon in einer geraden Linie, abweichend von der gebogenen Gestaltung bei Glossotherium. Das Gebiss bestand, wie allgemein üblich bei den Faultieren, aus 5 Zähnen je Oberkieferhälfte und 4 Zähnen je Unterkieferhälfte, insgesamt waren also 18 Zähne ausgebildet. Die jeweils vordersten Zähne hatten eine eckzahnartige (caniniforme) Gestalt, die übrigen waren molarenartig (molariform). Die Gebissstruktur gilt als stammesgeschichtlich urtümlich innerhalb der Faultiere. Allerdings waren bei den späteren Vertretern von Paramylodon die oberen caniniformen Zähne häufig reduziert, sodass das Gebiss dann aus nur 16 Zähnen bestand. Eine ähnliche Zahnreduktion ist bei Glossotherium nicht bekannt. Bei Mylodon hingegen waren die vordersten Zähne im Obergebiss ebenfalls nicht mehr ausgebildet, die unteren caniniformen Zähne ähnelten hingegen den hinteren Backenzähnen. Die caniniformen Zähne besaßen bei Paramylodon einen ovalen Querschnitt und waren nach hinten gekrümmt. Sie erreichten aber nicht die Größe wie bei Glossotherium oder gar bei Lestodon. Zur hinteren Zahnreihe bestand ein kurzes Diastema. Die molarenartigen Backenzähne hatten eine flache Gestalt mit einem etwas erhöhtem Rand. Sie besaßen im Umriss eine zweilappige Form mit starker mittlerer Einschnürung, außer dem ersten Oberkieferbackenzahn, der eher rechteckig geformt war und mit einer durchschnittlichen Länge von 3,7 cm den längsten Zahn im Oberkiefer bildete. Beim zweiten oberen Backenzahn zeichnete sich die lappenartige Struktur wesentlich deutlicher ab als bei Glossotherium. Allen Zähnen fehlte typischerweise der Zahnschmelz, vielmehr bestanden sie aus einer härteren Variante des Zahnbeins (Orthodentin), zusätzlich gab es noch eine äußere Schicht Zahnzement. Der Anteil des Orthodentins erreichte bei Paramylodon 28 %.[9] Die obere Zahnreihe wurde durchschnittlich 14,4 cm lang, wovon die hinteren Backenzähne 12,6 cm einnahmen. Aufgrund der sich nach vorn verbreiternden Schnauze verliefen die Zahnreihen divergierend zueinander.[1][3][6]
Körperskelett

Vor allem das umfangreiche Fundmaterial aus Rancho La Brea erlaubt eine umfassende Rekonstruktion des Körperskeletts. Die Wirbelsäule setzte sich aus 7 Hals-, 16 Brust- 8 bis 9 Lenden- und Kreuzbein- sowie 21 Schwanzwirbeln zusammen. Der Oberarmknochen war massiv, die Länge betrug 46 cm und der Kopf hob sich nicht sonderlich deutlich ab. Eine markante Knochenleiste (deltopectorale Leiste) saß am Humerusschaft an, war aber im oberen Teil weniger prominent ausgebildet als bei Glossotherium. Das untere Gelenkende kragte seitlich weit aus. Ein Foramen entepicondylaris, das gelegentlich bei einigen Faultieren vorkam, war hier nicht ausgebildet. Die Elle besaß einen stark ausgedehnten oberen Gelenkfortsatz, das Olecranon. Er wurde etwa 20 cm lang, der gesamte Knochen erreichte 40 cm Länge. Der Bau der Elle wirkte kürzer und robuster als bei Glossotherium, der Schaft war breit und oben vorn und hinten verschmälert. Ebenso war die Speiche kurz und massiv bei einer Länge von 29,6 cm. Den längsten Knochen repräsentierte der Oberschenkelknochen mit rund 54,6 cm. Sehr kurze Exemplare aus Rancho La Brea maßen nur 51 cm, sehr lange 58 cm. Markant war die bei den Bodenfaultieren typische, flache und breite Gestaltung, sodass der Knochen fast brettartig wirkte. Der Kopf erhob sich nur wenig von der Oberfläche und hatte eine mehr nach innen gerichtete Lage. Der Schaft war leicht nach innen gedreht, ein dritter Trochanter als Muskelansatzstelle, der bei Lestodon auftrat, war bei Paramylodon nicht sichtbar. Mit einer Länge von 24,6 cm war das Schienbein deutlich kürzer als der Oberschenkelknochen. Dies ist ein typisches Merkmal der Mylodonten, bei deren überwiegend späten Vertretern der untere Abschnitt des Hinterbeins häufig nur etwa die Hälfte der Länge des oberen erreichten.[10] Im Falle von Paramylodon wies die Tibia 45 % der Femurlänge auf. Ihr Schaft war wie der des Oberschenkelknochens abgeplattet und wies ebenso eine leichte Drehung auf. Das obere Gelenkende war seitlich ausladend, die Breite hier erreichte etwa drei Viertel der Länge des Gesamtknochens. Das Wadenbein war nicht mit dem Schienbein verwachsen, es wurde 26,3 cm lang.[3][11]
_7_(15444177065).jpg.webp)
Hände und Füße zeigten einen ähnlichen Aufbau wie bei den anderen großen Mylodonten Glossotherium und Lestodon, Abweichungen liegen im Detail vor. Die Hand wies insgesamt fünf Strahlen auf (I bis V), wobei nur die drei inneren Strahlen (I bis III) Krallen ausgebildet hatten. Der Mittelhandknochen des ersten Strahls war mit dem Großen Vieleckbein zu einer Einheit verschmolzen, was bei bodenbewohnenden Faultieren häufig belegt ist (sogenannter Metacarpal Carpal complex oder MCC). Die Mittelhandknochen des dritten bis fünften Strahls waren massiv und über 10 cm lang, der des Strahls IV besaß dabei den kräftigsten Bau. Am Fingerstrahl I waren zusätzlich noch die ersten beiden Phalangen miteinander verwachsen, an den Strahlen II und III bestanden jeweils drei Fingerglieder, wovon die ersten beiden aber deutlich reduzierte Längen aufwiesen. Die jeweiligen Endglieder der drei inneren Strahlen verfügten über ausgedehnte Krallenfortsätze, was auf dementsprechend große Krallen schließen lässt. Die Länge reichte von innen (I) nach außen (III) von 7,5 cm über 15,4 cm bis zu 17,4 cm, die Höhe variierte von 2,9 bis 5,7 cm. Die krallenlosen äußeren Finger besaßen in ihrer Größe stark reduzierte Fingerglieder. Der Fuß von Paramylodon verfügte über insgesamt vier Strahlen (II bis V), der innerste Strahl war vollständig reduziert. Krallen bestanden hier nur an den Zehen II und III, die auch am kräftigsten ausgebildet waren. Allerdings hatten die Mittelfußknochen hier eher kurze Längen von 3,6 beziehungsweise 6,5 cm, an den äußeren Strahlen wurden sie jeweils über 11,0 cm lang und waren sehr massiv. Wie bei den anderen beiden Mylodonten verfügte der zweiten Strahl nur über zwei Zehenglieder, da die erste und zweite Phalanx entsprechend der Hand zu einer Einheit verschmolzen waren. Abweichend von Glossotherium und Lestodon bestand bei Paramylodon auch der dritte Strahl häufig nur aus zwei Gliedern. Die jeweiligen Endphalangen mit Krallen wiesen analog der Hand einen äußerst kräftigen Bau auf, allein der Krallenfortsatz maß hier etwa 8,5 cm am zweiten und 11,1 cm am dritten Strahl und wurde jeweils 3,3 beziehungsweise 3,9 cm hoch. Die äußeren Strahlen besaßen dem gegenüber wiederum stark reduzierte Endglieder.[2][3]
Osteoderme
Die Mylodonten bilden die einzige bekannte Faultierlinie, bei deren Vertretern Knochenplättchen, sogenannte Osteoderme, in der Haut ausgebildet waren, analog den heutigen Gürteltieren. Im Gegensatz zu diesen bildeten sie aber bei den Mylodonten keinen festen Knochenpanzer, sondern waren eher locker verstreut, wie dies Funde von Hautresten von Mylodon zeigen.[12] Von Paramylodon liegen mehrere hundert solcher Osteoderme aus Rancho La Brea vor,[13][14][3] zusätzlich unter anderem auch als dichte Lage auf einer Platte aus dem Anza-Borrego State Park in Kalifornien[15] und aus Haile 15A, einer fossilreichen Kalksteinspalte in Florida. Die Knochenplättchen waren rund bis oval, manchmal auch irregulär geformt, und 5 bis 30 mm lang. Sie zeigten eine raue Oberfläche mit unregelmäßigen Eintiefungen, die Unterseite war dagegen glatt und konvex gestaltet. Im Querschnitt besaßen sie einen kompakten Bau bestehend aus zahlreichen Faserbündeln vermischt mit harten Knochenlamellen (Osteome). Prinzipiell waren die Knochenplättchen der Mylodonten einfacher strukturiert als die der Gepanzerten Nebengelenktiere.[16]
Verbreitung und wichtige Fossilfunde
Überblick und frühes Auftreten
Paramylodon war endemisch in Nordamerika und möglicherweise auch in Mittelamerika verbreitet. Die ältesten, eindeutig der Gattung zuweisbaren Funde sind aus dem Unteren Pleistozän bekannt. Ältere Formen von Mylodonten stammen aus dem Oberen Pliozän Mexikos[17] und des US-Bundesstaates Florida. Von letzterem ist das Teilskelett von der Fundstelle Haile 15A hervorzuheben, einer mit Sedimenten gefüllten Spalte im Kalkstein im Alachua County, dessen Alter auf 2,1 bis 1,8 Millionen Jahren geschätzt wird. Diese frühen Vertreter werden allgemein als „Glossotherium“ chapadmalensis bezeichnet,[18] die Stellung innerhalb der Gattung Glossotherium ist aber umstritten.[19][6] Nur wenig jünger sind die Funde der fossilreichen El Gulfo local fauna aus dem Mündungsgebiet des Colorado River im mexikanischen Bundesstaat Sonora. Sie werden bereits zu Paramylodon gestellt und datieren auf 1,8 bis 1,6 Millionen Jahre.[20] Insgesamt sind Fossilreste aus dem Unteren und Mittleren Pleistozän relativ selten und stammen in Nordamerika von rund 20 Fundstellen. Diese verteilen sich vor allem auf den südlichen und zentralen Bereich der heutigen USA und auf das nördliche Mexiko, streuen im Westen des Kontinents aber auch bis in den südlichen Teil von Alberta in Kanada. Sie befinden sich sowohl im Flachland als auch in Gebirgslagen, der höchste Fundpunkt erreicht in Colorado etwa 2900 m Höhe.[21][19][22] Einer der bedeutendsten Fundpunkte jener Zeit ist der Leisey Shell pit im Hillsborough County in Florida, von wo über mehrere Schädel und postcraniale Skelettelemente berichtet wurde, deren Alter bei etwa 1,2 Millionen Jahren liegt.[19] Bereits in den Übergang zum Mittelpleistozän gehört die Lokalität Fairmead Landfill im Madera County in Colorado, die ebenfalls mehrere Teilskelette hervorbrachte.[22]
Funde des Oberpleistozäns

Deutlich umfangreicher ist das Fundmaterial des Oberpleistozäns, dass mehr als 100 Fundstellen entstammt, allein in Kalifornien ist Paramylodon von über 60 Fundpunkten überliefert. Die Verbreitung der Gattung ist ähnlich wie im Unteren Pleistozän, zusätzlich kommt sie aber auch etwas weiter östlich im Mittleren Westen vor, so beispielsweise in Iowa.[23] Ihren nördlichsten Fundpunkt erreicht sie zu jener Zeit mit Sequim in Washington bei 48,1° nördlicher Breite, im Süden ist die Gattung zusätzlich aus Mexiko überliefert, wo bei Valsequillo bei 19° nördlicher Breite ihre südliche Ausbreitungsgrenze bestand, einige Funde weisen aber inzwischen darauf hin, dass Paramylodon eventuell auch aus Guatemala und El Salvador überliefert ist.[21][19] Unter anderem konnten Funde eines Jungtiers und eines ausgewachsenen Individuums von der Stevenson Bridge in Flussablagerungen des Putah Creek im Yolo County von Kalifornien geborgen werden, die an den Beginn der letzten Kaltzeit gehören.[24] Aus Shonto und Richville in Arizona sind zwei nahezu vollständige Skelette berichtet worden, die zu den wenigen bekannten Funden aus dem Bundesstaat gehören. Allgemein sind Fossilreste von Paramylodon auf dem Colorado-Plateau im Südwesten der heutigen USA und zusätzlich im Nordwesten Mexikos sehr selten, was eventuell mit dem damals trockeneren Klima in diesem Bereich zusammenhängt.[17][25]
Von weltweit herausragender Bedeutung sind aber die Funde aus den Asphaltgruben von Rancho La Brea im Süden von Kalifornien. Von hier stammt eine umfangreiche Fossilfauna, deren Alter von 45.000 bis 14.000 Jahren vor heute reicht. Die ersten Funde wurden bereits in der zweiten Hälfte des 19. Jahrhunderts entdeckt, das weitaus bedeutendere Material ist aber auf gezielte wissenschaftliche Untersuchungen zu Beginn des 20. Jahrhunderts zurückzuführen, die insgesamt über 100 dokumentierte Fundstellen einschließen.[1][3][26] Auffallend im Faunenspektrum ist die ungewöhnliche Dominanz von Raubtieren gegenüber Pflanzenfressern. Höchstwahrscheinlich wurden die Beutegreifer in größerer Zahl von im Bitumen feststeckenden Tieren angezogen und fielen den natürlichen Fallen dann selbst zum Opfer. Unter den Faultieren treten mit Paramylodon, Megalonyx und Nothrotheriops drei der vier in Nordamerika nachgewiesenen Gattungen auf (Eremotherium ist nur aus dem östlichen Teil der USA bekannt). Dabei stellt aber Paramylodon mit über 70 Individuen den weitaus häufigsten Vertreter dar, unter den Funden sind allein 30 Schädel hervorzuheben.[27][28] Ein weiterer sehr umfangreicher Fossilkomplex liegt mit der Diamond Valley Lake Local Fauna im Diamond Valley und im Domenigoni Valley im Riverside County ebenfalls im Süden von Kalifornien vor. Das Material wurde während der Konstruktion des Diamond Valley Lake seit Mitte der 1990er Jahre geborgen und umfasst gegenwärtig mehr als 100.000 Fundobjekte von mehr als 100 Taxa, die von über 2600 verschiedenen Fundlokalitäten stammen. Im Gegensatz zu Rancho La Brea dominieren hier die großen Pflanzenfresser, während der Anteil der großen Beutegreifer gering ausfällt. Dadurch kann auf einen ungestörten Charakter der Faunengemeinschaft geschlossen werden. Paramylodon ist mit rund 280 Einzelfunden belegt, was etwa 8 % der gesamten Säugetierfauna ausmacht. Das Bodenfaultier bildet damit nach den Bisons, den Pferden, dem urtümlichen Rüsseltier Mammut pacificus und dem Kamel Camelops den fünfthäufigsten Vertreter der Säugetiere in der Diamond Valley Lake Local Fauna. Die beiden anderen, auch in Rancho La Brea auftretenden Faultiere Megalonyx und Nothrotheriops spielen dagegen mit zusammen 0,5 % des Fundaufkommens nur eine untergeordnete Rolle. Das Alter der Diamond Valley Lake Local Fauna entspricht Radiocarbondatierungen zufolge dem von Rancho La Brea.[29][30]
Jüngste Nachweise
Wie die meisten anderen großen, bodenlebenden Faultiere verschwand auch Paramylodon gegen Ende des Pleistozäns im Zuge der Quartären Aussterbewelle. Im Gegensatz zu vielen anderen Gattungen liegen aber von Paramylodon kaum direkt am Fossilmaterial gemessene radiometrische Daten vor. Zu den jüngsten gehört ein Wert aus Rancho La Brea, der bei 20.450 BP liegt.[31] Allerdings sind noch eindeutig jüngere Funde bekannt, wenige davon kamen dabei aus archäologischen Fundstätten zum Vorschein, die im Zusammenhang mit der frühen Besiedlung des nordamerikanischen Kontinents durch den Menschen stehen. Einer der seltenen Nachweise stammt aus El Fin del Mundo („Das Ende der Welt“) in Sonora. Die im Jahr 2007 entdeckte Station konnte mit Hilfe der Radiocarbonmethode unter Verwendung von Holzkohle auf ein Alter von 13.390 Jahren BP datierte werden. Aufgrund des Vorkommens von sechs Clovis-Spitzen gehört sie in einen sehr frühen Abschnitt der Clovis-Kultur, welche eine der frühesten archäologischen Gruppen der ersten Besiedler Nordamerikas repräsentiert. Neben zwei Skeletten von Rüsseltieren, von denen eines eindeutig Cuvieronius vertritt und offensichtlich von den damaligen Jägern und Sammlern zerlegt worden war, kamen auch Reste von Paramylodon zum Vorschein.[32][33] Darüber hinaus sind mehr als 130 Osteoderme aus der Aubrey Clovis site im nordzentralen Texas belegt. Das die Funde umgebende Bodensubstrat wurde radiometrisch auf ein Alter von 12.860 Jahren BP datiert. Dort ebenfalls dokumentierte Steinartefakte, die etwa 9800 Stücke umfassen, können aufgrund einer Clovis-Spitze ebenfalls zur Clovis-Gruppe verwiesen werden. Die Reste von Paramylodon haben aber keinen unmittelbaren Bezug zu den frühen Siedlern, da sie mit Ausnahme eines einzigen Knochenplättchens in einer nahe gelegenen Wasserstelle gefunden wurden. Es ist aufgrund der bisher wenigen gemeinsamen Funde unklar, ob direkte Bejagung zum Aussterben der Tiere geführt hat.[34][35]
Dass die frühen Besiedler Nordamerikas mit den großen Bodenfaultieren interagierten, ihnen folgten sowie möglicherweise auch jagten, darauf verweisen Trittsiegel aus White Sands National Monument in New Mexico. Hier sind mehrere hundert Fußspuren größerer Faultiere mit denen von Menschen am Ufer eines ehemaligen Sees assoziiert. Teilweise überschneiden sie sich, in einem Fall liegen die menschlichen Spuren innerhalb der Fährte eines Faultiers. Eine Auffälligkeit bei den sich überkreuzenden Faultier- und Menschenspuren ist ein abrupter Richtungswechsel bei ersteren, was eine direkte Konfrontation durch die Verursacher annehmen lässt. Allerdings liegen von der Fundstelle keine Fossilreste vor und das Alter der Spuren ist bisher nur mittelbar datiert (zwischen 15.560 und 10.000 Jahren vor heute). Zudem sind die Faultierspuren nicht genauer bestimmt. Sie weisen starke Größenvariationen auf, was entweder auf Tiere verschiedenen Alters oder auf unterschiedliche Arten zurückgeführt werden kann. Im fraglichen Zeitraum kamen in der Region neben Paramylodon auch Nothrotheriops, ein kleineres Bodenfaultier aus der Gruppe der Nothrotheriidae, und Megalonyx, eine große Form der Megalonychidae, vor.[36]
Paläobiologie
Körpergrößenveränderung und Geschlechtsdimorphismus
Wie zahlreiche andere Tiergruppen auch durchlief Paramylodon im Laufe seiner Stammesgeschichte eine markante Körpergrößenzunahme. Das Gewicht bei den Angehörigen des Unteren Pleistozäns wird mit etwa 915 kg angegeben, die späten Vertreter aus dem Oberen Pleistozän erreichten dagegen wohl bis zu 1,39 t Körpergewicht. Grundlage für die jeweiligen Gewichtsschätzungen bilden die Oberschenkelknochen, deren entsprechenden Längen bei 48,4 beziehungsweise bei 54,6 cm liegen. Die frühesten Formen aus dem Pliozän, deren Stellung aber innerhalb der Gattung Paramylodon vielfach diskutiert wird, wiesen ein Gesamtgewicht von rund 310 kg bei einer Femurlänge vom 35,5 cm auf. Unter Berücksichtigung dieser frühen Vertreter erhöhte sich das Gewicht von Paramylodon im Laufe von gut 2,5 Millionen Jahren um den Faktor 4,5.[4] Besonders auffällig ist, dass gerade im ausgehenden Pleistozän zur Zeit der letzten Kaltzeit mit ihren extrem ausgeprägten Klimaschwankungen kaum Größenvariationen auftreten, wie Untersuchungen an den zahlreichen Funden von Rancho La Brea aus der Zeit vor 45.000 bis 10.000 Jahren vor heute zeigen. Erklärt wird dies mit einer hohen Flexibilität der Gattung in Bezug auf die Umwelt und damit einer hohen Anpassungsfähigkeit. Die Annahme lässt aber außer Acht, dass zunehmend kühlere Bedingungen laut der Bergmannschen Regel zu einer Körpergrößenzunahme führen müssten.[37]
Anhand der umfangreichen Fossilfunde aus dem späten Pleistozän können bei Paramylodon zwei Morphotypen unterschieden werden, eine grazile und eine robuste Variante. Die Morphotypen spiegeln sich dabei nicht in der allgemeinen Größe der Schädel wider, sondern betreffen hauptsächlich deren Ausprägung, etwa bei den Breitenverhältnissen. Auch lassen sich Unterschiede beispielsweise am Hinterhauptsbein feststellen, das bei der robusten Variante senkrecht steht, bei der grazileren aber schräg nach hinten. Dadurch sind bei letzterer die Gelenkflächen zum Ansetzen der Halswirbelsäule prominenter hervorgehoben als bei ersterer. Weitere Abweichungen finden sich in der Ausbildung der caniniformen Zähne die, sofern vorhanden, bei robusten Individuen spitz, bei grazilen dagegen stumpf enden. Möglicherweise handelt es sich bei den beiden Morphotypen nicht um Art- oder taxonomische Variationen im Sinne von Unterarten, wie es ursprünglich angenommen wurde, da sie häufig an ein und derselben Fundstelle auftreten. Vielmehr sind sie eher Ausdruck eines intraspezifischen Geschlechtsdimorphismus. Es ist allerdings momentan unmöglich, einen Morphotyp einem bestimmten Geschlecht zuzuweisen. Bei den 30 bekannten Schädelfunden von Rancho La Brea liegt das Verhältnis von robust zu grazil bei 3:1, im Americas Fall Reservoir in Idaho mit drei Schädeln bei 2:1 und in Ingleside in Texas mit ebenfalls drei Schädeln bei 3:0.[27] Bemerkenswert ist dabei, dass der Geschlechtsdimorphismus sich nicht im postcranialen Skelett niederschlägt und so, wie beim Schädel bereits bemerkt, kein Größendimorphismus in Form signifikanter Längenunterschiede bei den Extremitätenknochen auftritt. Im Gegensatz dazu ist bei Eremotherium, das zur gleichen Zeit ebenfalls in Nordamerika verbreitet war, aber zu den Megatheriidae gehört, ein ausgeprägter Größenunterschied zwischen den Geschlechtern bekannt.[38]
Fortbewegung
Allgemein wird für die bodenlebenden Faultiere eine vierfüßige Fortbewegung angenommen. Aufgrund des weit nach hinten verlagerten Körperschwerpunktes war es ihnen aber offensichtlich auch möglich, in eine zweifüßige Position zu wechseln, wobei sie sich dabei mit dem kräftigen – im Gegensatz zu den heutigen Baumfaultieren – sehr langen Schwanz abzustützen vermochten.[39] Der Hinterfuß von Paramylodon ist nach innen gedreht, sodass die Hauptlast beim Aufsetzen des Fußes auf dem äußeren Strahl (V) liegt. Dadurch entsteht der für zahlreiche Bodenfaultiere charakteristische pedolaterale Gang, der deutliche Umstrukturierungen in der Form und Lagerung der Fußwurzelknochen zueinander, vor allem beim Sprungbein und beim Fersenbein, erforderte. Bei Paramylodon war die äußere Fußkante wenig aufgewölbt, sodass sie eine mehr oder weniger gerade Kante bildete und das Fersenbein in nahezu voller Länge im Kontakt mit dem Boden stand. Dies ist übereinstimmend mit anderen Mylodonten, weicht aber stark von den nahe verwandten Scelidotheriidae ab, die einen hoch aufgewölbten Fuß besaßen, bei dem nur das hintere Ende des Fersenbeins den Boden berührte.[40][41] Eine weitere Besonderheit findet sich in den hinteren Gliedmaßen. Hier zeichnet sich der Bewegungsapparat durch einen extrem kurzen unteren Abschnitt aus. Bei Paramylodon erreicht der untere Abschnitt weniger als 50 % des oberen. Ein derartiger Bauplan, den nahezu alle Mylodonten aufweisen, lässt auf eine eher langsame und schwerfällige Bewegung schließen. Im Vergleich dazu hatten die Megatherien deutlich längere untere Gliedmaßenabschnitte.[42]
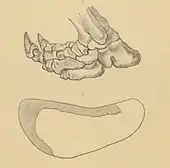
Spurenfossilien, die Hinweise auf die Fortbewegung der Bodenfaultiere geben, sind nur selten erhalten. Für Paramylodon konnten solche eindeutigen Trittsiegel im Nevada State Prison in der Nähe von Carson City nachgewiesen werden. Die Spuren wurden bereits in der zweiten Hälfte des 19. Jahrhunderts beim Sandsteinabbau entdeckt und anfänglich, 1882, als Hinweise auf riesenhafte Menschen gedeutet. Doch schon im Jahr darauf erkannte Othniel Charles Marsh einen Zusammenhang mit ausgestorbenen Bodenfaultieren und suchte den Verursacher der Trittsiegel unter den Mylodonten, von denen auch Knochenreste von der gleichen Fundstelle vorliegen.[43] Insgesamt sind bei Carson City neben Paramylodon zahlreiche Spuren weiterer Säugetiere – etwa Mammute, Pferde, Elche sowie Raubtiere – und zusätzlich von Vögeln entdeckt worden. Die Trittsiegel verteilen sich auf einer Fläche von rund 8000 m², sie sind zwar heute weitgehend durch den Gefängnisbau überdeckt, aber durch Abgüsse gut dokumentiert. Von Paramylodon konnten insgesamt zehn Fährten beobachtet werden, bestehend aus 15 bis 20 einzelnen, wechselseitig eingedrückten Spuren. Jeder einzelne Fußabdruck ist 47 bis 51 cm lang und rund 20 cm breit, im Umriss ähneln sie tatsächlich menschlichen Fußabdrücken, sie sind aber seitlich deutlich stärker eingedellt.[44] Weitere Studien zeigten, dass die Form der Abdrücke sehr gut mit der Form des Fußes von Paramylodon übereinstimmt[1][45] und der an Menschen erinnernde Umriss auf den auswärts gedrehten Fuß des Faultiers zurückzuführen ist. Der seitliche Abstand der Spuren zueinander beträgt etwa 60 cm, was in etwa mit dem Abstand der beiden Hüftgelenkspfannen am Becken von Paramylodon übereinstimmt, ebenso die Schrittlänge von rund 146 cm, was wiederum zur bekannten Hinterbeinlänge von 95 cm korrespondiert. Auffälligerweise sind dadurch fast ausschließlich Hinterfußabdrücke überliefert, was anfänglich auch mit einer zweifüßigen Fortbewegung der Tiere interpretiert wurde, analog zu entsprechenden Spurenfossilien von Megatherium in Südamerika. Es konnte aber ermittelt werden, dass die einzelnen Trittsiegel der Hinterfüße die der Vorderfüße überdecken. Da der vordere Fuß deutlich kleiner ist als der hintere und anders aufsetzt, erzeugt er einen wesentlich kleineren Abdruck. In Einzelfällen sind Trittsiegel überliefert, die von der Überdeckung des Vorderfußes durch den Hinterfuß zeugen. Aus anatomischen Gründen, etwa den angewinkelten Beinen beim Laufen, der Lage und Orientierung des Hinterfußes zum Bein und ähnlichen, ist eine dauerhafte Bipedie der großen Bodenfaultiere unwahrscheinlich. Die Schrittlänge von Paramylodon lässt eine durchschnittliche Geschwindigkeit von 1,8 bis 2,2 m/s annehmen, was in etwa der ermittelten Geschwindigkeit von Megatherium entspricht.[40]
Untersuchungen am Schulterblatt von sowohl jungen als auch ausgewachsenen Individuen zeigen eine signifikante Formveränderung, die von einer eher runden bei ersteren zu einer weitgehend ovoiden Form bei letzteren führt. Die ontogenetische Überprägungen sind vergleichbar mit denen der heutigen baumbewohnenden Faultiere. Die Ähnlichkeit der Schulterblätter der jungen Vertreter von Paramylodon und der Jungtiere der heutigen Faultiere lässt auf vergleichbare Verhaltensweisen schließen. Demnach verfügten die Jungen von Paramylodon noch über verschiedene kletternde Fähigkeiten und klammerten sich möglicherweise an ihre Muttertiere beim Transport.[46]
Ernährungsweise
Die Mylodonten gelten in der Regel als stärker an Grasnahrung angepasste Faultiere, die Annahme basiert auf den hochkronigen Zähnen und den abweichend von anderen Faultierlinien flachen Kauoberflächen, die dadurch denen der sich so ernährenden Huftiere ähneln.[8] Der fehlende Zahnschmelz bei den Zähnen der Faultiere macht aber Vergleiche schwierig. Die grasfresserische Ernährung wurde schon sehr früh aufgrund der speziellen Zahnausbildung angenommen,[3] Analysen des Kauapparates von Paramylodon ergaben, dass die Nahrung überwiegend in vor- und rückwärts gerichteten sowie seitlichen Kaubewegungen zerkleinert wurde, was auch durch dementsprechende Schleifspuren angezeigt wird. Dem widersprechen auch nicht die caniniformen vorderen Zähne, die – wenn ausgebildet – eher klein sind. Das Unterkiefergelenk ist bei Paramylodon breit ausgebildet und hat eine unspezialisierte Oberfläche, die zugehörige Glenoid-Grube am Schädel erscheint flach, was typisch für Pflanzenfresser mit ihren rotierenden Kaubewegungen ist. Allerdings tritt an der Innenseite eine zusätzliche, nahezu senkrecht stehende Gelenkfacette auf, die in einer Vertiefung an der Außenwand des Flügelbeins ankert. Dadurch waren allzu starke seitliche Kaubewegungen eher begrenzt. Die Anordnung des Musculus masseter bewirkte, dass Paramylodon sein Maul nur um 22° öffnen konnte, was deutlich weniger ist als bei den Zweifinger-Faultieren, die aber vergleichsweise längere caniniforme Zähne haben. Alles in allem lässt die Struktur des Kauapparates eher eine Bevorzugung gemischter Pflanzenkost (mixed feeder) annehmen.[47][48] Für eine derartige Ernährungsweise spricht auch der Bau der Schnauze, die nicht ganz so breit ist wie bei Lestodon, für das eine stärker auf Gräsern beruhende Ernährungsweise angenommen wird (grazer, analog dem heutigen Breitmaulnashorn), aber deutlich breiter als bei Megatherium, das eher Blätter bevorzugte (browser, vergleichbar zum Spitzmaulnashorn).[49][48] Die verlängerte Symphyse des Unterkiefers ragt weit über die Nasenregion hinaus. Da keine Verknöcherung der Nasenscheidewand vorliegt wie bei Mylodon, muss hier eine kräftige Knorpelentwicklung angenommen werden.[47] Möglicherweise hatte zudem die Zunge bei der Nahrungsaufnahme auch eine unterstützende Funktion. Durch die weit im Schädel zurückverlagerte Position des Zungenbeins und dessen robusten Bau mit kräftigen Muskelansatzstellen war beispielsweise der Musculus geniohyoideus besonders kräftig und lang ausgebildet, sodass eine sehr bewegliche Zunge anzunehmen ist.[50]
Da von Paramylodon im Gegensatz zu Mylodon keine Koprolithen bekannt sind, können die Nahrungsüberreste nicht direkt ermittelt werden. Zudem sind aufgrund des fehlenden Zahnschmelzes nur selten detaillierte Isotopenuntersuchungen möglich. Die Durchführung derartige Methoden bedarf daher einer exzellenten Fossilerhaltung, im Falle von Paramylodon gelang sie am Zahnbein einiger Zähne aus der oberpleistozänen Fundstelle von Ingleside in Texas. Die Ergebnisse, die dabei mit Hilfe der Kohlenstoffisotope gewonnen wurden, liegen zwischen den Bereichen von heutigen Herbivoren mit Spezialisierung auf eine harte (grazer) oder weiche (browser) Pflanzenkost und befürworten somit eine gemischte Nahrung, möglicherweise aber mit einer stärkeren Tendenz zu Gräsern.[51] Somit stimmen die bisherigen Ergebnisse gut mit der offenen Landschaft überein, in der Paramylodon lebte. Es wird aber auch angenommen, dass der Faultiervertreter möglicherweise nach Wurzeln grub. Hierfür sprechen etwa die kräftigen Vorderbeine, die über einen robusten, am unteren Gelenkende weit ausladenden Oberarmknochen, eine kurze Speiche mit lang ausgezogenem Olecranon für eine massige Unterarmmuskulatur und etwas abgeplatte Krallen verfügten und so sehr gut zum Graben geeignet waren. Darüber hinaus sind Unterschiede im Gebissaufbau zwischen frühen und späten Angehörigen der Gattung erkennbar. So besaßen die Formen aus dem Unterpleistozän noch weniger hohe Zahnkronen und dementsprechend einen niedrigeren Unterkiefer, während die des Oberpleistozäns deutlich höhere Zähne und einen massigeren Unterkiefer aufwiesen. Möglicherweise spiegelt dies eine zunehmend stärkere Anpassung von Paramylodon im Laufe der Zeit wider.[8][19]
Sonstiges
Der überwiegende Teil der Funde von Paramylodon umfasst einzelne Individuen, Massenansammlungen wie etwa in Rancho La Brea stellen Akkumulationen über mehrere Jahrtausende dar und haben so einen eher zufälligen Charakter. Es kann daher angenommen werden, dass die Faultiergattung wie die heutigen baumlebenden Vertreter einzelgängerisch auftrat und allenfalls Mutter-Jungtier-Gruppen bildete. Das Verdauungssystem war wahrscheinlich ähnlich strukturiert wie bei den rezenten Arten, womit auch bei Paramylodon ein eher langsamer Stoffwechsel vorherrschte mit langer Passagezeit der Nahrung, die dadurch aber in großem Umfang verwertet wurde. In Verbindung mit der eher langsamen Fortbewegung spricht auch dies gegen größere saisonale Wanderungen – im Gegensatz zu zahlreichen anderen Offenlandschaften bewohnenden Grasfressern. Demnach waren die Tiere relativ standorttreu. Auffällig an zahlreichen Fundstellen mit Paramylodon ist das häufige gemeinsame Auftreten mit dem Präriemammut und dem Bison. Beide Vertreter großer pflanzenfressender Säugetiere zeigten aber mit ihren Herdenbildungen und weiten Wanderungen sowie abweichendem Verdauungssystem eine grundlegend andere Lebensweise. Daher nutzte wohl Paramylodon eine unterschiedliche ökologische Nische zur Vermeidung direkter Konkurrenz mit den anderen Megaherbivoren der nordamerikanischen Steppenlandschaften.[21]
Systematik
Innere Systematik der Mylodontidae nach Boscaini et al. 2019 (basierend auf skelettanatomischen Analysen)[52]
|
Innere Systematik der Faultiere nach Presslee et al. 2019 (basierend auf Proteinanalysen)[53]
|
Paramylodon ist eine ausgestorbene Gattung aus der ebenfalls erloschenen Familie der Mylodontidae. Die Mylodontidae bilden wiederum einen Teil der Unterordnung der Faultiere (Folivora). In einer klassischen, auf skelettanatomischen Merkmalen beruhenden Systematik repräsentieren die Mylodontidae zusammen mit den Orophodontidae und den Scelidotheriidae die Überfamilie der Mylodontoidea (teilweise werden die Scelidotheriidae und die Orophodontidae aber auch nur als Unterfamilie innerhalb der Mylodontidae geführt[54]), welche wiederum neben den Megatherioidea die zweite große und bedeutende Faultierlinie darstellt. Gemäß molekulargenetischen und proteinbasierten Untersuchungen können neben den Mylodontoidea und den Megatherioidea auch die Megalocnoidea als dritte große Linie herausgestellt werden. Den Mylodontoidea sind dann mit den Zweifinger-Faultieren (Choloepus) auch eine der zwei heute noch bestehenden Faultiergattungen zuzuweisen.[55][53] Die Mylodontidae formen eine der vielfältigsten Gruppen innerhalb der Faultiere. Zu ihren Charakteristika gehören hochkronige Zähne mit abweichend von den Megatherioidea flacher (lobater) Kaufläche, was als eine Anpassung an stärker grashaltige Nahrung interpretiert wird. Die hinteren Zähne besitzen einen runden oder ovalen Querschnitt, die vordersten sind eckzahnartig gestaltet. Der Hinterfuß zeigt zudem eine deutlich seitlich ausgedrehte Gestalt.[56][23] Zu den frühesten Nachweise der Mylodonten wird Paroctodontotherium gezählt, welches in Salla-Luribay in Bolivien nachgewiesen ist und in das Oligozän datiert.[57]
Die innere Gliederung der Mylodontidae ist komplex und variiert je nach Bearbeiter. Zumeist anerkannt sind die späten Gruppen der Mylodontinae mit Mylodon als Typusform und der Lestodontinae, deren Charakterform Lestodon darstellt (auf tribaler Ebene als Mylodontini und Lestodontini bezeichnet). Teilweise wurden zahlreiche weitere Unterfamilien aufgestellt, etwa die Nematheriinae für Vertreter aus dem Unteren Miozän oder die Octomylodontinae für alle Basalformen, die aber nicht allgemein anerkannt sind.[58] Mit der Etablierung der Urumacotheriinae für spätmiozäne Gattungen des nördlichen Südamerikas konnte im Jahr 2004 eine weitere Linie innerhalb der Mylodonten herausgearbeitet werden.[59] Prinzipiell wird für die gesamte Familie eine Revision angemahnt, da zahlreiche der höheren taxonomischen Einheiten keine formale Diagnose besitzen.[10]
Die Untergliederung der terminalen Gruppe der Mylodonten in die Lestodontinae und Mylodontinae wurde generell auch in einer der bisher umfangreichsten Studien zur Stammesgeschichte der Faultiere aus dem Jahr 2004 bestätigt. Im Ergebnis dieser auf Schädelmerkmalen basierenden und von Timothy J. Gaudin durchgeführten Analyse steht Paramylodon in der Nähe von Mylodon, das relativ ähnliche Glossotherium bildet aber das Schwestertaxon zu den Lestodontinae.[60] In einigen späteren phylogenetischen Arbeiten ließen sich die Resultate reproduzieren.[61][62] Eine im Jahr 2019 vorgestellte Untersuchung von Luciano Varela und weiteren beteiligten Wissenschaftlern, die zahlreiche fossile Formen der gesamten Unterordnung der Faultiere einbezieht, sieht dies dagegen kritischer. Hier sind Paramylodon und Glossotherium eng verwandt, Mylodon hingegen formt die Bais der entwickelten Mylodonten und Lestodon gruppiert mit einigen Formen aus dem nördlichen Südamerika.[54] Im gleichen Jahr wurde eine höherauflösende phylogenetische Analyse der Mylodonten durch eine Arbeitsgruppe um Alberto Boscaini veröffentlicht. Diese untermauert die Zweigliederung der terminalen Vertreter und vereint Paramylodon, Glossotherium und Mylodon in einer Klade. Als fundamentaler Unterschied zwischen den Mylodontinae und Lestodontinae kann die Ausprägung der eckzahnartigen vorderen Zähne herangezogen werden. Bei letzteren sind diese groß und durch ein langes Diastema von den hinteren Zähnen getrennt, erstere habe nur kleine caniniforme Zähne, die mitunter auch teilweise reduziert sind, generell aber dichter an den molarenartigen Zähnen anstehen. Der Studie zufolge bilden Paramylodon, Glosstherium und Mylodon eine engere Beziehungsgemeinschaft innerhalb der Mylodontinae.[52] Diese Ansicht findet auch durch die bereits erwähnten biochemischen Daten Unterstützung, ebenfalls vorgelegt 2019.[53] Detaillierte, bereits im Jahr 2009 von Robert K. McAfee publizierte Schädelanalysen legen ebenfalls nahe, dass Paramylodon und Glossotherium sehr eng miteinander verwandt sind und sich höchstwahrscheinlich einen gemeinsamen Vorfahren teilen. Zu den Merkmalen, die die beiden Gattungen verbindet, gehören etwa der Gebissaufbau mit den vorderen caniniformen Zähnen und die Zahnstruktur, etwa des zweiten Backenzahns, oder die Lage der Knochennaht zwischen Gaumenbein und Oberkiefer nahe dem hintersten Zahn. Dagegen ist Mylodon mit seinem reduzierten Gebiss, den einfacher gestalteten Zähnen und der vorverlagerten Knochenverbindung zwischen Gaumenbein und Oberkiefer deutlicher abweichend.[6]
_BHL4368870.jpg.webp)
Innerhalb der Gattung Paramylodon ist mit P. harlani nur eine Art anerkannt. Eine weitere Art, P. nebrascensis, wurde 1903 von Barnum Brown anhand eines Teilskelettes aus Hay Spring in Nebraska beschrieben,[11] aber bereits in den 1920er Jahren mit der Nominatform vereint. Nur zehn Jahre später kreierte Glover Morrill Allen unter Zuhilfenahme eines weiteren Teilskelettes vom Niobrara River in Nebraska die Art Mylodon garmani,[63] die aber ebenfalls als synonym zu Paramylodon harlani angesehen wird. Gleiches gilt für mehrere von Edward Drinker Cope bereits in den 1870er und 1890er Jahren benannte Arten wie Mylodon sodalis und Mylodon sulcidens.[19] Die ursprünglich vorgenommene Untergliederung in zwei Unterarten, P. h. harlani für eine robuste und P. h. tenuiceps für eine grazile Form, wie es Chester Stock im Jahr 1917 vorschlug,[1][3] wird heute nicht mehr befürwortet.[27] Problematisch ist allerdings die Art „Glossotherium“ chapadmalense. Die Art wurde ursprünglich im Jahr 1925 von Lucas Kraglievich anhand eines 39 cm langen, nahezu unbeschädigten Schädels mit Unterkiefer aus Schichten des Mittleren Pliozän östlich von Miramar in der argentinischen Provinz Buenos Aires aufgestellt.[64] Sie weist Ähnlichkeiten zu Glossotherium robustum auf, besitzt aber auch einzelne Abweichungen, die möglicherweise einen eigenen Gattungsstatus rechtfertigen, vorgeschlagen wurde etwa die Bezeichnung Eumylodon (die Kraglievich bereits 1925 für Eumylodon chapadmalense verwendete). Die Form könnte damit eventuell der gemeinsame Vorfahr von Glossotherium und Paramylodon sein. Ob dies allerdings auch für die nordamerikanischen Funde aus dem Pliozän von Florida und Mexiko gilt, die von Jesse S. Robertson erstmals im Jahr 1976 unter dem gleichen Artnamen geführt wurden,[18] oder diese näher zu Paramylodon stehen, ist momentan aufgrund fehlender Vergleichsuntersuchungen unklar.[6] Teilweise werden die frühen Mylodontenreste auch unter der Bezeichnung P. garbanii geführt,[65] eine Artbezeichnung, die 1986 für einige pliozäne Unterkiefer- und Gliedmaßenreste aus Arroyo EI Tanque im mexikanischen Bundesstaat Guanajuato geprägt worden war (unter dem wissenschaftlichen Namen Glossotherium garbanii).[66] Die Art ist aber nicht vollständig anerkannt, andere Autoren sehen sie als synonym zu „Glossotherium“ chapadmalense.[17]
Forschungsgeschichte
Entdeckungen in Nord- und Südamerika

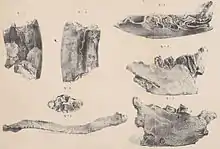
Die Forschungsgeschichte von Paramylodon ist komplex und von einer über mehr als 150 Jahre währenden Verwechslung und Gleichsetzung mit Mylodon und Glossotherium geprägt. Sie beginnt aber mit den ersten Entdeckungen von Richard Harlan (1796–1843) am Big Bone Lick im Boone County im US-Bundesstaat Kentucky im Jahr 1831, die einen rechten Unterkiefer und ein Schlüsselbein umfassen. Harlan erkannte, dass es sich um Reste eines ausgestorbenen Faultiers handelte und verwies sie zu Megalonyx, das damals aus Nordamerika bereits bekannt war, und innerhalb der Gattung zu der von ihm kurz zuvor aufgestellten Art Megalonyx laqueatus.[67][68] Die Funde wurden ursprünglich in New York aufbewahrt, sind aber heute verschollen.[19]
Zwischen den Jahren 1831 und 1836 unternahm Charles Darwin seine wegweisende Reise mit der HMS Beagle nach Südamerika und brachte von dort eine große Anzahl an Fossilien mit. Diese wurden dann von Richard Owen, einem der bedeutendsten Forscher des Viktorianischen Zeitalters, untersucht und die Ergebnisse veröffentlicht. In einer ersten Publikation zu den Säugetierresten allgemein im Jahr 1840 führte er die Gattung Mylodon mit der Art Mylodon darwinii ein. Gattung und Art basierten auf einem Unterkiefer, den Darwin in Punta Alta in der argentinischen Provinz Buenos Aires gefunden hatte. Als besonderes Kennzeichen hoben sich insgesamt vier molarenartige Zähne je Zahnreihe hervor. Zugleich bemerkte Owen auch Ähnlichkeiten im Zahnbau zwischen Harlans Unterkiefer und dem von Mylodon darwinii. Daraus folgernd verwarf er die von Harlan geprägte Bezeichnung Megalonyx laqueatus und schuf mit Mylodon harlani eine neue Art.[69] Der Gattungsname Mylodon setzt sich aus den griechischen Wörtern μύλη (myle „Molar“) und ὀδούς (odoús „Zahn“) zusammen, bedeutet übersetzt also so viel wie „Molarenzahn“. Harlan äußerte sich zwei Jahre später über die Verwendung des Namens, da dieser seiner Meinung nach kein herausragendes Charakteristikum des Tieres beschreibe und jedes ausgestorbene Säugetier meinen könnte, weil fast alle über die hinteren Backenzähne verfügten.[70]
Im selben Jahr, 1842, legte Owen eine umfassende Beschreibung eines Skeletts eines Mylodonten vor, das aus den Überschwemmungsebenen des Río de la Plata nördlich von Buenos Aires stammte; er etablierte für dieses die neue Art Mylodon robustus.[71] Zu diesem Zeitpunkt bestand also die Gattung Mylodon aus drei Arten, von denen zwei in Südamerika und eine in Nordamerika vorkam. Zudem sollte es sich als problematisch erweisen, dass Owen Mylodon darwinii als Typusart der Gattung auswies, obwohl diese, wie er zugab, nach Mylodon harlani die zweite bekannte und beschriebene Art sei. Demnach hätte eigentlich Mylodon harlani das Anrecht auf den Status als Nominatform. Im weiteren Verlauf wurden Mylodon unterschiedliche Typusarten zugeordnet, so sah Johannes Theodor Reinhardt 1879 diese in Mylodon robustus,[72] Richard Lydekker 1887 dagegen in Mylodon harlani.[73][6][74]
Paramylodon und die Mylodon-Glossotherium-Problematik

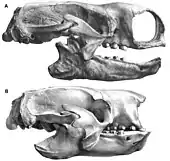
Im Jahr 1903 führte Barnum Brown (1873–1963) den Gattungsnamen Paramylodon ein. Er verwendete dazu ein Teilskelett aus Hay Spring in Nebraska, das 1897 bei einer Expedition des American Museum of Natural History entdeckt worden war. Der Gattung wies er mit Paramylodon nebrascensis eine Art zu. Als definierende Unterschiede zum nordamerikanischen Mylodon harlani, das Brown als Typusform von Mylodon ansah, gab er die fehlenden vorderen caniniformen Zähne im Oberkiefer an. Dadurch waren zu jener Zeit zwei unterschiedliche Vertreter der Mylodonten im Pleistozän Nordamerikas anerkannt.[11][19]
Später wies Chester Stock (1892–1950) aufgrund seiner Untersuchungen am Fundmaterial von Rancho La Brea darauf hin, dass das Merkmal der fehlenden oberen Vorderzähne bei Mylodon harlani sehr variabel ausgebildet ist. Daher synonymisierte er 1917 P. nebrascensis mit Mylodon harlani.[1][3] Im Jahr 1928 beschränkte Lucas Kraglievich allerdings die nordamerikanischen Funde auf Paramylodon und trennte die Gattung somit von den südamerikanischen Vertretern ab,[75] eine Meinung, der sich acht Jahre später auch Ángel Cabrera anschloss; sie fand aber in der Folgezeit bei den meisten Forschern kaum Resonanz. Kraglievich revidierte im gleichen Zug auch Glossotherium als eigenständige, von Mylodon zu unterscheidende Gattung.[75] Glossotherium war ursprünglich ebenfalls von Owen in seiner Schrift über Darwins Entdeckungen aus dem Jahr 1840 anhand eines Schädelbruchstückes vom Arroyo Sarandí im Südwesten des heutigen Uruguay aufgestellt worden, nur zwei Jahre später vereinte er es aber mit Mylodon.[69][71]
In der Folgezeit nach Kraglievich und Cabrera entwickelte sich Glossotherium aufgrund der Namenspriorität zu einem „Abfalleimer“-Taxon, in das zahlreiche nahe verwandte Formen eingestellt wurden. George Gaylord Simpson gab 1945 in seiner generellen Taxonomie der Säugetiere zum Ausdruck, dass, wenn Paramylodon nicht eindeutig von Glossotherium getrennt werden könne, aufgrund ebendieser Namenspriorität Glossotherium vorzuziehen wäre.[76] Durch die darauffolgende vollständige Eingliederung von Paramylodon in die Gattung, die Robert Hoffstetter im Jahr 1952 vollzog, gehörte Glossotherium zu den wenigen Faultierformen, die in Süd- und Nordamerika auftraten, sie besaß dadurch aber auch eine hohe Variabilität. Zahlreiche Forscher favorisierten im Verlauf des 20. Jahrhunderts die Ansicht der Kongenerität der beiden Faultierformen.[18] Im Jahr 1995 trennte H. Gregory McDonald jedoch das nordamerikanische Paramylodon wieder vom südamerikanischen Glossotherium. Er merkte dabei an, dass bisher keine Untersuchungen vorliegen, in denen nachgewiesen wurde, dass beide Gattungen tatsächlich identisch seien. Vielmehr spräche die Isolierung von Paramylodon in Nordamerika eher für eine Eigenständigkeit der Form.[19] In der darauffolgenden Zeit konnten mehrere Schädelstudien vorgelegt werden, die die beiden Gattungen und zuzüglich auch Mylodon eindeutig voneinander unterschieden.[6][77]
Literatur
- Robert K. McAfee: Reassessment of the cranial characters of Glossotherium and Paramylodon (Mammalia: Xenarthra: Mylodontidae). Zoological Journal of the Linnean Society, 155, 2009, S. 885–903
- H. Gregory McDonald und Steve Pelikan: Mammoths and mylodonts: Exotic species from two different continents in North American Pleistocene faunas. Quaternary International 142/143, 2006, S. 229–241
- Chester Stock: A mounted skeleton of Mylodon harlani. University of California Publications, Bulletin of the Department of Geology12 (6), 1920, S. 425–430 ()
- Chester Stock: Cenozoic gravigrade edentates of western North America with special reference to the Pleistocene Megalonychinae and Mylodontidae of Rancho La Brea. Carnegie. Institute of Washington 331, 1925, S. 1–206
Einzelnachweise
- Chester Stock: Further observation on the skull structure of the mylodont sloths from Rancho La Brea. University of California Publications, Bulletin of the Department of Geology 10, 1917, S. 165–178
- Chester Stock: Structure of the pes in Mylodon harlani. University of California Publications. Bulletin of the Department of Geology 10 (16), 1917, S. 267–286
- Chester Stock: Cenozoic gravigrade edentates of western North America with special reference to the Pleistocene Megalonychinae and Mylodontidae of Rancho La Brea. Carnegie. Institute of Washington 331, 1925, S. 1–206
- H. Gregory McDonald: Paleoecology of extinct Xenarthrans and the Great American Biotic Interchange. Bulletin of the Florida Museum of Natural History 45 (4), 2005, S. 313–333
- Chester Stock: A mounted skeleton of Mylodon harlani. University of California Publications, Bulletin of the Department of Geology12 (6), 1920, S. 425–430 ()
- Robert K. McAfee: Reassessment of the cranial characters of Glossotherium and Paramylodon (Mammalia: Xenarthra: Mylodontidae). Zoological Journal of the Linnean Society, 155, 2009, S. 885–903
- T. D. A. Cockerell: A fossil ground-sloth in Colorado. University of Colorado Studies 6, 1909, S. 309–312
- M. Susana Bargo, Gerardo de Iuliis und Sergio F. Vizcaíno: Hypsodonty in Pleistocene ground sloths. Acta Palaeontologica Polonica 51 (1), 2006, S. 53–61
- Daniela C. Kalthoff: Microstructure of Dental Hard Tissues in Fossil and Recent Xenarthrans (Mammalia: Folivora and Cingulata). Journal of Morphology 272, 2011, S. 641–661
- Ascanio D. Rincón, H. Gregory McDonald, Andrés Solórzano, Mónica Núñez Flores und Damián Ruiz-Ramoni: A new enigmatic Late Miocene mylodontoid sloth from northern South America. Royal Society Open Science 2, 2015, S. 140256, doi:10.1098/rsos.140256
- Barnum Brown: A new genus of ground sloth from the Pleistocene of Nebraska. Bulletin of the American Museum of Natural History 29, 1903, S. 569–583
- F. P. Moreno und A. Smith Woodward: On a Portion of Mammalian Skin, named Neomylodon listai, from a Cave near Consuelo Cove, Last Hope Inlet, Patagonia. By Dr. F. P. Moreno, C.M.Z.S. With a Description of the Specimen by A. Smith Woodward, F.Z.S. Proceedings of the Zoological Society1899, S. 144–156 ()
- John C. Merriam: Recent discoveries of Quaternary mammals in Southern California. Science24, 1906, S. 248–250
- William J. Sinclair: Dermal Bones of Paramylodon from the Asphaltum Deposits of Rancho la Brea, near Los Angeles, California. Proceedings of the American Philosophical Society 49 (195), 1910, S. 191–195
- H. Gregory McDonald: An Overview of the Presence of Osteoderms in Sloths: Implications for Osteoderms as a Plesiomorphic Character of the Xenarthra. Journal of Mammalogy 25, 2018, S. 485–493, doi:10.1007/s10914-017-9415-8
- Robert V. Hill: Comparative Anatomy and Histology of Xenarthran Osteoderms. Journal of Morphology 267, 2005, S. 1441–1460
- H. Gregory McDonald: Fossil Xenarthra of Mexico: a review. In: Marisol Montellano Ballesteros und Joaquín Arroyo Cabrales (Hrsg.): Avances en los Estudios Paleomastozoológicos en México. Serie Arqueología, Instituto Nacional de Antropología e Historia, Córdoba, 2002, S. 227–248
- Jesse S. Robertson: Latest Pliocene mammals from Haile XVA, Alachua County, Florida. Bulletin of the Florida Museum of Natural History. 20, 1976, S. 111–186 ()
- H. Gregory McDonald: Gravigrade xenarthrans from the early Pleistocene Leisey Shell Pit 1A, Hillsborough County, Florida. Bulletin of the Florida Museum of Natural History 37, 1995, S. 345–373 ()
- Fred W. Croxen III, Christopher A. Shaw und David R. Sussman: Pleistocene Geology and Paleontology of the Colorado River Delta at Golfo de Santa Clara, Sonora, Mexico. In: Robert E. Reynolds (Hrsg.): Wild, scenic & rapid-a trip down the Colorado River trough. The 2007 Desert Symposium field guide and abstracts from proceedings. California State University, 2007, S. 84–89
- H. Gregory McDonald und Steve Pelikan: Mammoths and mylodonts: Exotic species from two different continents in North American Pleistocene faunas. Quaternary International 142/143, 2006, S. 229–241
- H. Gregory McDonald, Robert G. Dundas und James C. Chatters: Taxonomy, paleoecology and taphonomy of ground sloths (Xenarthra) from the Fairmead Landfill locality (Pleistocene: Irvingtonian) of Madera County, California. Quaternary Research 79, 2013, S. 215–227
- H. Gregory McDonald: Harlan's Ground Sloth (Paramylodon harlani) (Xenarthra: Mylodontidae) from the Late Pleistocene (Rancholabrean) of Iowa. The Journal of the Iowa Academy of Science 119 (1-4), 2012, S. 16–21
- Robert G. Dundas und Laura M. Cunningham: Harlan's ground sloth (Glossotherium harlani) and a Columbian mammoth (Mammuthus columbi) from Stevenson Bridge, Yolo County, California. PaleoBios 15 (3), 1993, S. 47–62
- H. Gregory McDonald, Larry D. Agenbroad und Carol Manganaro Haden: Late Pleistocene mylodont sloth Paramylodon harlani (Mammalia: Xenarthra) from Arizona. The Southwestern Naturalist 49 (2), 2014, S. 229–238
- Anthony R. Friscia, Blaire van Valkenburgh, Lillian Spencer und John Harris: Chronology and spatial distribution of large mammal bones in Pit 91, Rancho La Brea. Palaios 23(1), 2008, S. 35–42
- H. Gregory McDonald: Sexual dimorphism in the skull of Harlan's ground sloth. Contribution in Science 510, 2006, S. 1–9
- Leslie F. Marcus: A census of the abundant large Pleistocene mammals from Rancho La Brea. Contributions in Science 38, 1960, S. 1–11
- Kathleen Springer, Eric Scott, J. Christopher Sagebiel und Lyndon K. Murray: The Diamond Valley Lake Local Fauna: Late Pleistocene vertebrates from inland Southern California. Museum of Northern Arizona Bulletin 65, 2009, S. 217–236
- Kathleen Springer, Eric Scott, J. Christopher Sagebiel und Lyndon K. Murray: Late Pleistocene large mammal faunal dynamics from inland southern California: The Diamond Valley Lake local fauna. Quaternary International 217, 2010, S. 256–265
- J. Tyler Faith und Todd A. Surovell: Synchronous extinction of North America’s Pleistocene mammals. PNAS 106 (49), 2009, S. 20641–20645
- Joaquín Arroyo-Cabrales, Alejandro Lopez-Jimenez, Guadalupe Sanchez und Edmund P. Gaines: Faunal remains from El Fin del Mundo archaeological site, Sonora, Mexico. Journal of Vertebrate Paleontology 31 (suppl.), 2011, S. 64
- Guadalupe Sanchez, Vance T. Holliday, Edmund P. Gaines, Joaquín Arroyo-Cabrales, Natalia Martínez-Tagüeña, Andrew Kowler, Todd Lange, Gregory W. L. Hodgins, Susan M. Mentzer und Ismael Sanchez-Morales: Human (Clovis)–gomphothere (Cuvieronius sp.) association∼13,390 calibrated yBP in Sonora, Mexico. PNAS 111, 2014, S. 10972–10977
- C. Reid Ferring: The Archaeology and Paleoecology of the Aubrey Clovis Site (41DN479) Denton County, Texas. Center for Environmental Archaeology, Department of Geography, University of North Texas, Denton, 2002, S. 1–276
- Donald K. Grayson und David J. Meltzer: Clovis Hunting and Large Mammal Extinction: A Critical Review of the Evidence. Journal of World Prehistory 16 (4), 2002, S. 313–359
- David Bustos, Jackson Jakeway, Tommy M. Urban, Vance T. Holliday, Brendan Fenerty, David A. Raichlen, Marcin Budka, Sally C. Reynolds, Bruce D. Allen, David W. Love, Vincent L. Santucci, Daniel Odess, Patrick Willey, H. Gregory McDonald, und Matthew R. Bennett: Footprints preserve terminal Pleistocene hunt? Human-sloth interactions in North America. Science Advances 4 (4), 2018, S. eaar7621, doi:10.1126/sciadv.aar7621
- Donald R. Prothero und Kristina R. Raymond: Stasis in the Late Pleistocene ground sloths (Paramylodon harlani) from Rancho La Brea Tar pits, California. New Mexico Museum of Natural History and Science Bulletin 53, 2011, S. 624–628
- Donald R. Prothero und Kristina R. Raymond: Variation and sexual size dimorphism in Pleistocene ground sloths (Xenarthra). New Mexico Museum of Natural History and Science Bulletin 44, 2008, S. 331–333
- M. Susana Bargo, Sergio F. Vizcaíno, Fernando M. Archuby und R. Ernesto Blanco: Limb bone proportions, strength and digging in some Lujanian (Late Pleistocene-Early Holocene) mylodontid ground sloths (Mammalia, Xenarthra). Journal of Vertebrate Paleontology 20 (3), 2000, S. 601–610
- H. Gregory McDonald: Biomechanical inferences of locomotion in ground sloths: Integrating morphological and track data. New Mexico Museum of Natural History and Science Bulletin 42, 2007, S. 201–208
- H. Gregory McDonald: Evolution of the Pedolateral Foot in Ground Sloths: Patterns of Change in the Astragalus. Journal of Mammalian Evolution 19, 2012, S. 209–215
- Gerardo De Iuliis, Guillermo H. Ré und Sergio F. Vizcaíno: The Toro Negro megatheriine (Mammalia, Xenarthra): A new species of Pyramiodontherium and a review of Plesiomegatherium. Journal of Vertebrate Paleontology 24 (1), 2004, S. 214–227
- Othniel Charles Marsh: On the supposed human foot-prints recently found in Nevada. American Journal of Science 152, 1883, S. 139–140
- Joseph LeConte: Carson footprints. Nature 1883, S. 101–102
- Chester Stock: Origin of the supposed human footprints of Carson City, Nevada. Science 51, 1920, S. 514
- Andy D. Grass: Inferring differential behavior between giant ground sloth adults and juveniles through scapula morphology. Journal of Vertebrate Paleontology 39 (1), 2019, S. e1569018, doi:10.1080/02724634.2019.1569018
- Virginia L. Naples: The feeding mechanism in the Pleistocene ground sloth, Glossotherium. Contributions in Science Los Angeles County Museum of Natural History 415, 1989, S. 1–23
- M. Susana Bargo und Sergio F. Vizcaíno: Paleobiology of Pleistocene ground sloths (Xenarthra, Tardigrada): biomechanics, morphogeometry and ecomorphology applied to the masticatory apparatus. Ameghiniana 45 (1), 2008, S. 175–196
- M. Susana Bargo, Néstor Toledo und Sergio F. Vizcaíno: Muzzle of South American Pleistocene Ground Sloths (Xenarthra, Tardigrada). Journal of Morphology 267, 2006, S. 248–263
- Leandro M. Pérez, Néstor Toledo, Gerardo De Iuliis, M. Susana Bargo und Sergio F. Vizcaíno: Morphology and Function of the Hyoid Apparatus of Fossil Xenarthrans (Mammalia). Journal of Morphology 271, 2010, S. 1119–1133
- Dennis R. Ruez, Jr.: Diet of Pleistocene Paramylodon harlani (Xenarthra: Mylodontidae): Review of methods and preliminary use of carbon isotopes. The Texas Journal of Science 57 (4), 2005, S. 329–344
- Alberto Boscaini, François Pujos und Timothy J. Gaudin: A reappraisal of the phylogeny of Mylodontidae (Mammalia, Xenarthra) and the divergence of mylodontine and lestodontine sloths. Zoologica Scripta 48 (6), 2019, S. 691–710, doi:10.1111/zsc.12376
- Samantha Presslee, Graham J. Slater, François Pujos, Analía M. Forasiepi, Roman Fischer, Kelly Molloy, Meaghan Mackie, Jesper V. Olsen, Alejandro Kramarz, Matías Taglioretti, Fernando Scaglia, Maximiliano Lezcano, José Luis Lanata, John Southon, Robert Feranec, Jonathan Bloch, Adam Hajduk, Fabiana M. Martin, Rodolfo Salas Gismondi, Marcelo Reguero, Christian de Muizon, Alex Greenwood, Brian T. Chait, Kirsty Penkman, Matthew Collins und Ross D. E. MacPhee: Palaeoproteomics resolves sloth relationships. Nature Ecology & Evolution 3, 2019, S. 1121–1130, doi:10.1038/s41559-019-0909-z
- Luciano Varela, P. Sebastián Tambusso, H. Gregory McDonald und Richard A. Fariña: Phylogeny, Macroevolutionary Trends and Historical Biogeography of Sloths: Insights From a Bayesian Morphological Clock Analysis. Systematic Biology 68 (2), 2019, S. 204–218
- Frédéric Delsuc, Melanie Kuch, Gillian C. Gibb, Emil Karpinski, Dirk Hackenberger, Paul Szpak, Jorge G. Martínez, Jim I. Mead, H. Gregory McDonald, Ross D.E. MacPhee, Guillaume Billet, Lionel Hautier und Hendrik N. Poinar: Ancient mitogenomes reveal the evolutionary history and biogeography of sloths. Current Biology 29 (12), 2019, S. 2031–2042, doi:10.1016/j.cub.2019.05.043
- H. Gregory McDonald und Gerardo de Iuliis: Fossil history of sloths. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 39–55.
- Bruce J. Shockey und Federico Anaya: Grazing in a New Late Oligocene Mylodontid Sloth and a Mylodontid Radiation as a Component of the Eocene-Oligocene Faunal Turnover and the Early Spread of Grasslands/Savannas in South America. Journal of Mammalian Evolution 18, 2011, S. 101–115
- Andrés Rinderknecht, Enrique Bostelmann T., Daniel Perea und Gustavo Lecuona: A New Genus and Species of Mylodontidae (Mammalia: Xenarthra) from the Late Miocene of Southern Uruguay, with Comments on the Systematics of the Mylodontinae. Journal of Vertebrate Paleontology 30 (3), 2010, S. 899–910
- Francisco Ricardo Negri und Jorge Ferigolo: Urumacotheriinae, nova subfamília de Mylodontidae (Mammalia, Tardigrada) do Mioceno Superior-Plioceno, América do Sul. Revista Brasileira de Paleontologia 7 (2), 2004, S. 281–288
- Timothy J. Gaudrin: Phylogenetic relationships among sloths (Mammalia, Xenarthra, Tardigrada): the craniodental evidence. Zoological Journal of the Linnean Society 140, 2004, S. 255–305
- Ascanio D. Rincón, Andrés Solórzano, H. Gregory McDonald und Mónica Núñez Flores: Baraguatherium takumara, Gen. et Sp. Nov., the Earliest Mylodontoid Sloth (Early Miocene) from Northern South America. Journal of Mammalian Evolution 24 (2), 2017, S. 179–191
- Luciano Brambilla und Damián Alberto Ibarra: Archaeomylodon sampedrinensis, gen. et sp. nov., a new mylodontine from the middle Pleistocene of Pampean Region, Argentina. Journal of Vertebrate Paleontology 2018, S. e1542308, doi:10.1080/02724634.2018.1542308
- Glover M. Allen: A new Mylodon. Memoirs of the Museum of Comparative Zoology 40 (7), 1913, S. 319–346
- Lucas Kraglievich: Cuatro nuevos gravigrados de la fauna Araucana „Chapadmalense“. Anales del Museo Nacional de Historia Natural de Buenos Aires 33, 1925, S. 215–235
- Gary S. Morgan: Vertebrate fauna and geochronology of the Great American Biotic Interchange in North America. New Mexico Museum of Natural History & Science Bulletin 44, 2008, S. 93–140
- Marisol Montellano-Ballesteros und Oscar Carranza-Castaneda: Descripcion de un milodontido del Blancano temprano de la mesa central de Mexico. Instituto de Geología, Universidad Nacional Autónoma de México, Revista 6, 1986, S. 193–203
- Richard Harlan: Description of the jaws, teeth, and clavicle of the Megalonyx laqueatus. The Monthly American journal of geology and natural science 1, 1831, S. 74–76 ()
- Richard Harlan: Description of the jaws, teeth, and clavicle of the Megalonyx laqueatus. Medical an physical researches, 1835, S. 334–336 ()
- Richard Owen: Fossil Mammalia. In Charles Darwin (Hrsg.): Zoology of the Voyage of H.M.S Beagle, under the command of Captain Fitzroy, during the years 1832 to 1836. Part I. Fossil Mammals. London, 1840, S. 12–111 (S. 63–73) ()
- Richard Harlan: Notice of two new mammals from Brunswick Cana, Georgia; with observations on the fossil quadrupeds of the United States. The American journal of science and arts 43, 1842, S. 141–144 ()
- Richard Owen: Description of the skeleton of an extinct gigantic Sloth, Mylodon robustus, Owen, with observations on the osteology, natural affinities, and probable habitats of the Megatherioid quadrupeds in general. London, 1842, S. 1–176 ()
- Johannes Theodor Reinhardt: Beskrivelse af Hovedskallen af et Kæmpedovendyr, Grypotherium darwini. Det Kongelige Danske Videnskabernes Selskabs Skrifter. 5 Række. Naturvidenskabelig og Mathematisk Afdeling 12 (4), 1879, S. 351–381 ()
- Richard Lydekker: Catalogue of the fossil Mammalia in the British Museum of Natural History. Part V. London, 1887, S. 1–345 (S. 106) ()
- Juan Carlos Fernicola, Sergio F. Vizcaíno und Gerardo De Iuliis: The fossil mammals collected by Charles Darwin in South America during his travels on board the HMS Beagle. Revista de la Asociación Geológica Argentina 64 (1), 2009, S. 147–159
- Lucas Kraglievich: „Mylodon darwini“ Owen, es la especie genotipo de „Mylodon“ Ow. Physis: Revista de la Sociedad Argentina de Ciencias Naturales 9, 1928, S. 169–185
- George Gaylord Simpson: The Principles of Classification and a Classification of Mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 71)
- Diego Brandoni, Brenda S. Ferrero und Ernesto Brunetto: Mylodon darwini Owen (Xenarthra, Mylodontinae) from the Late Pleistocene of Mesopotamia, Argentina, with Remarks on Individual Variability, Paleobiology, Paleobiogeography, and Paleoenvironment. Journal of Vertebrate Paleontology 30 (5), 2010, S. 1547–1558