Megatherium
Megatherium ist eine Gattung aus der ausgestorbenen Familie der Megatheriidae, einer Gruppe von teils riesigen Faultieren. Vor allem das elefantengroße M. americanum gehört zu den bekanntesten und am besten untersuchten Formen und stellt neben Eremotherium den größten bekannten Vertreter der großen Bodenfaultiere dar. Die Gattung trat vom frühen Pliozän vor rund 5 Millionen Jahren bis zum Übergang vom Oberen Pleistozän zum Unteren Holozän vor etwa 8.000, eventuell auch nur bis vor etwa 11.000 Jahren auf. Dabei sind innerhalb der Gattung zwei Entwicklungslinien zu unterscheiden. Die eine bildet die Tieflandform M. americanum, die die zentralen bis teils südlichen, unter temperiertem Klimaeinfluss stehenden Gebiete von Südamerika bewohnte, vornehmlich die Pamparegion im heutigen Argentinien, die andere trat überwiegend in den Hochgebirgslagen der Anden auf und schloss mehrere Arten ein. Beide Entwicklungslinien lassen sich anhand von Merkmalen am Skelett unterscheiden.
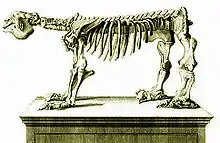
| Megatherium | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Megatherium americanum im Muséum national d’histoire naturelle in Paris | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Pliozän bis Oberes Pleistozän/Unteres Holozän | ||||||||||||
| 5,33 Mio. Jahre bis 11.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Megatherium | ||||||||||||
| Cuvier, 1796 | ||||||||||||
Als weitgehender Pflanzenfresser ernährte sich Megatherium von hauptsächlich weicher Pflanzenkost. Zur Nahrungsaufnahme vermochte es sich auf die Hinterbeine aufzurichten, wobei der kräftige Schwanz als Stütze am Boden diente. Die massigen Arme wurden zum Heranziehen von Ästen und Zweigen eingesetzt. Teilweise wird aber auch eine deutlich fleischhaltigere Ernährungsweise diskutiert. Ebenfalls im Zentrum einer wissenschaftlichen Debatte steht die Frage, ob Megatherium sich nicht nur zu Nahrungszwecken aufrichtete, sondern auch dauerhaft zu einer zweibeinigen Fortbewegung (Bipedie) imstande war, wobei es dann das größte bekannte Säugetier mit einer derartigen Bewegungsweise wäre. Sowohl die Ansicht einer räuberischen Lebensweise als auch eines dauerhaften zweibeinigen Ganges ist sehr umstritten.
Die Entdeckung von Fossilresten von Megatherium geht bis in das ausgehende 18. Jahrhundert zurück. Ein Skelett aus Luján im heutigen Argentinien führte 1796 zur wissenschaftlichen Erstbeschreibung der Gattung durch Georges Cuvier. Weitere bedeutende Funde wurden 1832 von Charles Darwin auf seiner historischen Reise mit der HMS Beagle um Südamerika in den Pampagebieten gesammelt. Megatherium spielte eine bedeutende Rolle in der Entwicklung und Etablierung der Paläontologie als Wissenschaftszweig im Verlauf des 19. Jahrhunderts. Die frühe Forschungsgeschichte der Gattung ist dabei nicht nur geprägt von zahlreichen neuen Erkenntnissen, sondern auch von einzelnen entscheidenden Irrtümern.
Merkmale
Körpergröße
Megatherium umfasst mittelgroße bis sehr große Vertreter der Bodenfaultiere. Die kleinste Art, M. altiplanicum, wog schätzungsweise zwischen 1,0 und 1,7 t,[1] M. americanum erreichte als größter Vertreter bei einer Gesamtlänge von 6 m und einer Schulterhöhe von 2 m vermutlich ein Körpergewicht von 3,8 bis 6,0 t (basierend auf verschiedenen Bestimmungsmethoden). Damit besaß Megatherium ähnliche Dimensionen wie das verwandte und etwa zeitgleich lebende Eremotherium.[2] Die Variabilität der Größe innerhalb der Gattung zeichnet sich unter anderem am charakteristischen Oberschenkelknochen ab. Dieser wies bei M. altiplanicum eine Länge von 39 cm auf, M. urbinai, M. sundti und M. tarijense repräsentieren mit 46 bis 53 cm langen Femora mittelgroße Arten, während am oberen Ende des Größenspektrums der Oberschenkelknochen von M. americanum zwischen 57 und 78 cm lang wurde.[3][4] Damit gehört Megatherium neben Eremotherium zu den größten bekannten Faultieren.[5][6] Allgemein war Megatherium nicht ganz so robust gebaut wie sein Verwandter Eremotherium und hatte vergleichsweise kürzere Gliedmaßen, aber einen längeren Schädel. Wie bei allen Bodenfaultieren waren Arme und Beine bezogen auf die Körperlänge aber deutlich kürzer als bei den heutigen Baumfaultieren. Weiterhin besaß Megatherium analog zu anderen bodenbewohnenden Faultieren einen kräftigen, langen Schwanz.[7]
Schädel- und Gebissmerkmale

Der Schädel der großen Arten wurde 72 bis 87 cm lang, an den Jochbögen erreichte er eine Breite von 36 bis 46 cm, am Hinterhauptsbein von 27 bis 31 cm.[8][9] Insgesamt war er röhrenförmig langgestreckt und schmal in der Aufsicht. Die Stirnlinie zeigte in der Seitenansicht ein markant sinusartig geschwungenes bis leicht domartiges Profil. Der paarige Mittelkieferknochen hatte eine Y- oder X- bis rechteckige Form, wobei die beiden Knochen fest miteinander verwachsen waren, ebenso wie eine feste Fusion mit dem Oberkiefer bestand. Allerdings war bei einigen kleineren Arten die Verbindung nicht sehr fest, so dass der Mittelkieferknochen teilweise getrennt vom Schädel aufgefunden wird oder mitunter auch unbekannt ist.[10][3] Der Oberkiefer war sehr hoch, um die extrem hochkronigen Zähne zu fassen. Ein Charakteristikum stellten die Jochbögen dar, die besonders massiv ausfielen und im Gegensatz zu den heutigen Baumfaultieren und zahlreichen fossilen Formen vollständig ausgebildet waren, so dass sich die beiden Bogenansätze des Joch- und des Schläfenbeins berührten. Weiterhin bestand ein kräftiger, abwärts gerichteter Knochenauswuchs am vorderen Jochbogenansatz, der als Muskelansatzstelle diente.[11] Das Scheitelbein wies einen kräftigen Scheitelkamm auf, der sich vorn teilte und jeweils über das Schläfenbein als scharfe Linie führte. Das Hinterhauptsbein besaß in der Seitenansicht eine rechtwinklige Form. Vor allem bei den kleineren Arten aus dem Andengebiet waren die Gelenkansätze für die Halswirbelsäule stark nach hinten verlängert und so sehr prominent.[12]
Der Unterkiefer zeigte einen massiven Bau und maß bei großen Arten zwischen 62 und 74 cm in der Länge.[9] Die Symphyse war ebenfalls kräftig, relativ flach und wie bei den meisten Faultieren vollständig verwachsen. Sie endete auf der Höhe des ersten oder zweiten Backenzahns. Nach vorn war sie deutlich verlängert und bildete so einen schmalen Knochenauswuchs, dessen Länge die der gesamten Zahnreihe in einigen Fällen übertraf. Sehr auffällig war der für Megatherien typische, nach unten gewölbte Rand des Unterkiefers. Hier erreichte der Knochenkörper teils bis zu 25 cm Höhe, was der höchste Wert für alle Megatherien ist, allerdings ist bei Megatherium die Ausbuchtung auch verglichen mit der Größe des Unterkiefers am deutlichsten.[13] In der Ausbuchtung lagen die Alveolen der Unterkieferzähne. Weiterhin ragten die Gelenkäste sehr weit nach oben. Das Gebiss bestand analog zu allen Megatherien aus fünf Zähnen je Oberkiefer- und vier je Unterkieferhälfte, insgesamt waren also 18 Zähne ausgebildet. Die Zähne waren homodont und molarenartig gestaltet, von quadratischer Form und besaßen auf der Kaufläche zwei quergestellte Leisten (bilophodont). Typischerweise besaßen die Zähne keinen Zahnschmelz und standen geschlossen in einer Reihe. Die Länge der unteren Zahnreihe variierte und lag beim großen M. americanum zwischen 18,6 und 25,9 cm, bei mittelgroßen Formen wie M. tarijense zwischen 15,2 und 20,4 cm oder bei M. medinae zwischen 13,0 und 17,2 cm und beim kleinen M. altiplanicum bei 14,4 cm.[13][3][10]
Körperskelett
Das Körperskelett ist aufgrund zahlreicher Funde weitgehend bekannt. Die Wirbelsäule umfasste 7 Hals-, 16 Brust-, 3 Lenden-, 3 Kreuzbein- und 17 bis 18 Schwanzwirbel. Im Gegensatz zu Eremotherium waren bei Megatherium die drei Gelenkflächen, die die Halswirbel Atlas und Axis miteinander verbinden, nicht untereinander fusioniert.[4][14] Die Gliedmaßen waren bei Megatherium generell durchschnittlich kürzer als bei Eremotherium. Der Oberarmknochen bestand als ein röhrenförmiger Knochen von bis zu 80 cm Länge, dessen unteres Gelenk wuchtig auslud und dabei bis zu 37 cm breit wurde. Generell zeigte sich der Humerus der Megatherien als länger, graziler und flacher als bei anderen Bodenfaultieren.[15] Die Elle besaß mit 69 cm Länge einen langen und schlanken Bau mit seitlich verschmälerten Gelenkenden. Das obere Gelenkende, das Olecranon, konnte bei den meisten kleineren Arten länglich und schmal gestaltet sein, bei den größten war es breit und sperrig. Der Oberschenkelknochen stellte wie bei allen Megatherien einen breiten und massiven Knochen dar, der aber vorn und hinten starke Verschmälerungen aufwies, so dass die Breite des Knochens die Tiefe um das Vierfache übertraf.[16] Der längste bekannte Knochen war 78 cm lang und gut 42 cm breit, der kleinste nur 39 cm lang und 20 cm breit. Teilweise war der Schaft deutlich gedreht. Der dritte Trochanter als Muskelansatzstelle am Schaft, der charakteristisch für Nebengelenktiere ist, fehlte wie bei allen Megatherien. Die Gelenkflächen standen deutlich hervor, wodurch der Knochen in der Gesamtansicht nicht so rechtwinklig wirkte wie bei Eremotherium.[1] Schien- und Wadenbein waren an beiden Enden miteinander verwachsen und nicht wie beim verwandten Eremotherium nur an einem. Die Länge des Schienbeins lag bei rund 54 cm.[8][12][11]


Die Hand war prinzipiell vierstrahlig aufgebaut (Strahlen II bis V) und wich damit von seinem nahen Verwandten Eremotherium mit einer dreistrahligen Hand (bei späten Vertretern) ab. Wie bei Eremotherium waren aber auch bei Megatherium die Mittelhandknochen der beiden äußeren Strahlen am stärksten entwickelt und erreichten Längen von 25 beziehungsweise 23 cm.[12] Sie übertrafen die beiden inneren Strahlen deutlich. Der Mittelhandknochen des ersten Strahles und einzelne Handwurzelknochen wie das Große Vieleckbein waren zum sogenannten Metacarpal-Carpal-Komplex (MCC) verwachsen (bei Eremotherium zusätzlich noch der zweite Metacarpus). Insgesamt besaßen nur der zweite, dritte und vierte Finger jeweils drei Phalangen, am dritten waren allerdings die beiden hintersten Phalangen zu einer Einheit verschmolzen. Die jeweiligen Endphalangen trugen Klauen und wiesen meist eine dreieckige Form im Querschnitt auf. Aufgrund der massigen Ausbildung des letzten Fingerglieds am dritten Strahl war hier wohl ursprünglich die größte Kralle ausgebildet. Am fünften Finger bestanden nur zwei Phalangen, das Endglied fehlte.[17][14][4] Der Fuß wies analog zu dem der anderen Megatherien drei Strahlen auf (Strahlen III bis V), wobei der Mittelfußknochen am dritten Strahl deutlich kürzer war. Dies war auch der einzige Zehenstrahl, der drei Phalangen umfasste – wie bei der Hand waren aber die ersten beiden Glieder miteinander verwachsen – und somit eine Kralle trug. Die beiden anderen, äußeren Strahlen hatten nur zwei, stark in der Länge reduzierte Zehenglieder und demzufolge keine Krallen ausgebildet.[4][18]
Verbreitung und bedeutende Fossilfunde

Megatherium war vom Pliozän bis zum frühen Holozän vor allem im mittleren und teils südlichen Bereich von Südamerika verbreitet, nur in den Hochgebirgslagen der Anden drang es weiter nach Norden vor. Funde sind überwiegend aus Argentinien, Uruguay, Chile, Bolivien, Peru und Ecuador bekannt. Die nördlicheren und eher tropisch geprägten Tieflandregionen wurden dagegen von der nahe verwandten Gattung Eremotherium bewohnt, die im Gegensatz zu Megatherium auch Nordamerika erreichte. Eines der bisher wenigen gemeinsamen Vorkommen der beiden riesigen Bodenfaultiere ließ sich in der mittel- bis oberpleistozänen Santa-Vitória-Formation an den Ufern des Arroio Chuí und weiterer Flüsse im brasilianischen Bundesstaat Rio Grande do Sul belegen. Megatherium ist hier mit einem Teilschädel und einzelnen Elementen des Körperskeletts überliefert.[19] Der Großteil der Funde stammt jedoch aus der Pamparegion in Argentinien und sie sind nach Auffassung vieler Forscher allesamt zu der sehr großen Art M. americanum zu stellen, die dieser Meinung nach auch den einzigen Vertreter im Tiefland östlich der Anden darstellte. Bedeutend sind unter anderem Punta Alta bei Bahía Blanca in der Provinz Buenos Aires, wo bereits Charles Darwin während seiner Reise mit der HMS Beagle im Jahr 1832 Funde sammelte,[20][21] oder Luján, eine der fossilreichsten Fundstellen Argentiniens in der gleichen Provinz, woher das Skelett stammte, das Georges Cuvier zur Erstbeschreibung von Megatherium diente.[22] Stellvertretend für die zahlreichen weiteren Funde in der Pampa sei hier der nahezu vollständige Unterkiefer von M. americanum von La Chumbiada am Río Salado im Norden der Provinz Buenos Aires genannt, der rund 12.000 Jahre alt ist und vergesellschaftet mit der Raubkatze Smilodon und dem Pferd Hippidion aufgefunden wurde.[23] Insgesamt stellt M. americanum nicht nur den größten Vertreter von Megatherium dar, sondern auch den am häufigsten dokumentierten und am besten untersuchten.[1] Allerdings sind vereinzelt auch Funde kleinerer Formen wie M. filholi aus den Pampagebieten bekannt, so dass die Gattung Megatherium vermutlich eine komplexere Verbreitungsgeschichte in der Region besaß. Hervorzuheben sind etwa die Fundstelle Rocas Negras bei Mar del Sur im Süden und Carmen de Areco sowie verschiedene weitere Plätze im Norden der Provinz Buenos Aires.[24]
Ansonsten sind zumeist kleinere Arten von Megatherium aus den Anden beschrieben worden. Von ihnen existiert aber in der Regel nur sehr geringes Fundmaterial und sie traten offensichtlich weniger häufig auf als ihr großer Verwandter in den Pampa-Tiefländern. Dadurch sind diese Arten aber auch meist weniger gut erforscht, wodurch beispielsweise kaum Aussagen zur innerartlichen Variabilität getroffen werden können.[4] Aus Chile ist M. medinae bekannt, eine mittelgroße und grazile Form, die unter Verwendung eines Gebissrestes und einzelner Hinterbeinknochen aus der Pampa del Tamarugal beschrieben wurde.[25][26] Weitere Funde, so ein Unterkiefer, Wirbel und Rippen, stammen aus der Nähe von Santiago de Chile.[27] Die kleinste und älteste Art stellt M. altiplanicum dar, die über einige Schädelreste und postcraniale Skelettelemente aus der unterpliozänen Umala-Formation nahe Ayo Ayo im Departamento La Paz im westlichen Bolivien nachgewiesen ist.[1] In geographischer Nähe ebenfalls im nördlichen Teil des Altiplano ist weiterhin aus der Ulloma-Formation des frühen Mittelpleistozäns nahe Ulloma am Río Desaguadero die mittelgroße Art M. sundti belegt. Ihr Fundmaterial umfasst neben zwei Schädeln, davon einer mit Unterkiefer, die 1893 zur Beschreibung der Art dienten,[25][26] unter anderem ein Teilskelett eines nicht ausgewachsenen Individuums.[28] Aus der Tarija-Formation nahe Tarija im südlichen Bolivien stammt hingegen das etwa ähnlich große M. tarijense in Form eines vollständigen Skeletts, welches bereits in den 1920er Jahren geborgen wurde.[3] Der gleichen Art kann auch ein Teilskelett aus Yuntac in Peru zugewiesen werden. Dieses gehört mit einer Lage in 4500 m über dem Meeresspiegel zu den höchstgelegenen Funden von Megatherium. Ein Oberschenkelknochen ist weiterhin aus dem Cuzco-Tal überliefert.[29] Ein unvollständiges Skelett aus der Santa-Rosa-Höhle bei Celendin in der peruanischen Provinz Cajamarca diente zur Beschreibung von M. celendinense, dem bisher größten Vertreter der andinischen Arten, der fast so groß wie M. americanum selbst war.[12] Deutlich kleiner und sehr grazil war dagegen M. urbinai. Von ihm sind ein Teilskelett aus Sacaco im Küstengebiet des südwestlichen Peru und ein weiteres aus der Tres-Vantanas-Höhle 70 km südöstlich von Lima in rund 4000 m Höhe geborgen worden.[4] Von einem dritten Skelett – einem Jungtier, in äolischen Sedimenten nahe Uyujalla ebenfalls im Küstengebiet gelegen – wurde ebenfalls berichtet, dieses ging aber noch vor der Bergung verloren. In La Brea bei Talara im nordwestlichen Küstenland Perus wurde ein Teilskelett von M. elenense gefunden, ebenso wie bei Cerro de Pasco in der Region Pasco in 4300 m Höhe. Dieser kleinere Vertreter von Megatherium war zuerst von Robert Hoffstetter im Jahr 1952 anhand einiger Funde von der Halbinsel Santa Elena in Ecuador eingeführt worden, allerdings unter dem Gattungsnamen Eremotherium.[29] Von der gleichen Halbinsel stammt auch das Typusmaterial von Eremotherium, das 1948 von Franz Spillmann beschrieben worden war.[30]
Paläobiologie
Fortbewegung

Im Gegensatz zu heutigen Faultieren und vergleichbar mit zahlreichen anderen ausgestorbenen Riesenfaultieren war Megatherium ein reiner Bodenbewohner, der sich hauptsächlich vierfüßig fortbewegte. Besonderes Kennzeichen sind die nach innen gedrehten Füße, so dass Megatherium sein Gewicht weitgehend auf den beiden äußersten Strahlen (IV und V) der Hinterfüße trug (pedolateral). Die Fußsohle zeigte so nach innen und hob sich in einem 35°-Winkel vom Boden ab. Der pedolaterale Gang stellt eine Besonderheit dar, die sich innerhalb der Faultiere mehrfach entwickelte und vor allem massive Umbauten am Sprungbein erforderte.[31][32] Auch die Hand wies eine vergleichbare Drehung auf. Beim Aufsetzen auf den Boden lag diese gleichfalls auf dem äußeren Strahl, wobei die Position Ähnlichkeiten zu den krallenbewehrten Chalicotheriidae besitzt, die aber zu den Unpaarhufern zu stellen sind. Die Drehung vor allem der Vorderfüße schränkte eine Manipulation von Objekten stark ein, sie konnten aber für das Heranziehen von Zweigen eingesetzt werden. Zudem wird aus der Stellung der Hände und Füße auf eine insgesamt langsame Fortbewegung geschlossen.[33]
Es ist aber möglich, dass Megatherium trotzdem markant agiler war als die heutigen baumbewohnenden Faultiere. Das wird aus Untersuchungen am Innenohr geschlossen, wobei die Größe der Bogengänge in direkter Beziehung zur Flinkheit eines Tieres steht. Den Ergebnissen zufolge sollte Megatherium über eine bis zu dreimal höhere Agilität als heutige Faultiere verfügt haben.[34] Allgemein ließen sich die deutlich gebogenen Krallen wohl auch gut zum Graben einsetzen und widerstanden durch ihre symmetrische Form den dabei auftretenden Zug- und Kompressionskräften, wie Belastungsanalysen aufzeigten, doch spricht der Bau des gesamten Unterarms weitgehend gegen eine bodenwühlende Lebensweise. So ist bei Megatherium unter anderem der obere Gelenkfortsatz der Elle (Olecranon) deutlich zu kurz und konnte dadurch die für das Aufbrechen des Bodensubstrats notwendigen Hebelbewegungen nicht erzeugen. Für andere sehr große Bodenfaultiere aus der Gruppe der Mylodontidae wie etwa Glossotherium lässt sich eine teils im Erdreich grabende Lebensweise aber gut belegen.[7][35][36]
Eine intensive Debatte wurde darüber geführt, ob Megatherium sich auch in aufrechter Position fortbewegte. Das Aufrichten auf die Hinterbeine, um in höheren Baumkronen nach Nahrung zu suchen, kann aufgrund der Konfiguration des Beckens und der Hinterbeine als wahrscheinlich angesehen werden, ebenso wie die Tatsache, dass der Schwerpunkt des Körpers sehr weit hinten lag, so dass rund 70 % des Gewichtes von den Hinterbeinen getragen wurden, und dadurch ein Aufrichten erleichterte.[7] Die Idee einer bipeden Fortbewegung kam schon Anfang des 20. Jahrhunderts auf und es wurde auch versucht, diese mit Hilfe von Ichnofossilien zu belegen. Solche Spurenfossilien von Megatherium liegen unter anderem vom Monte Hermosa und aus Pehuén-Có vor, beide Fundstellen liegen in der argentinischen Provinz Buenos Aires. Eine zentrale Rolle nehmen hier aber die Spuren von Pehuén-Có nahe Bahía Blanca ein. Diese 1986 entdeckte Fundstelle stellt eine der weltweit bedeutendsten von Ichnofossilien dar. Die Spuren verteilen sich auf einer Fläche von 1,5 km² und sind in einem ursprünglich weichen Substrat eingedrückt. Sie umfassen zahlreiche Säugetiere, etwa des Kamelartigen Megalamaichnum (Hemiauchenia), des südamerikanischen Huftiers Eumacrauchenichnus (Macrauchenia) oder des großen Gürteltierverwandten Glyptodontichnus (Glyptodon), und Vögel, etwa Aramayoichnus aus der Gruppe der Nandus. Ihr Alter wurde auf 12.000 Jahre datiert.[37][38]
Unter den Spurenfossilien befinden sich auch Abdrücke eines riesigen Faultiers, die der Ichnospezies Neomegatherichnum zugewiesen werden. Die Größe der einzelnen Trittsiegel liegt bei durchschnittlich 88 cm Länge und 48 cm Weite, sie entspricht in etwa den Maßen des aufsetzenden Hinterfußes vom M. americanum. Die Tiefe beträgt etwa 26 cm. Insgesamt sind von Megatherium rund 80 Fährten mit jeweils wenigstens fünf Einzelspuren überliefert, was die Faultiergattung zu einem der am häufigsten nachgewiesenen Tiere der Fundstelle macht. An einigen Trittsiegeln lassen sich noch die Krallen der Mittelzehe als separater Abdruck erkennen. Dieser steht zumeist in einem Winkel von 50 bis 90° zur Längsachse des Fußes und wird bis zu 15 cm lang.[20][38] Eine der längsten Fährten verläuft über circa 35 m Länge und setzt sich aus insgesamt 35 Einzelspuren zusammen. Der Abstand zueinander misst etwa 1,5 bis 1,8 m, was als Schrittlänge angenommen wird.[39] Da die Größe der Eindrücke relativ einheitlich ist, wurde geschlussfolgert, dass sich Megatherium überwiegend auf den Hinterbeinen fortbewegt habe. Nur etwa in der Mitte der Spurenfolge sind einzelne kleinere, nur 33 cm lange und 27 cm breite sowie 14 cm tiefe Abdrücke zu erkennen, wo sich das Tier offensichtlich kurz in eine vierfüßige Position begab. Die angenommene Geschwindigkeit liegt bei 1,2 bis 1,4 m/s, was etwas mehr als 4 km/h entspricht, die Höchstgeschwindigkeit wurde mit 1,7 m/s angesetzt. Biomechanische Untersuchungen zeigten weiterhin, dass die besondere Form des Oberschenkelknochens – hier vor allem der langschmale Querschnitt – seitlich auftretenden Beanspruchungen, die beim zweifüßigen Gang stärker wirken als beim vierfüßigen, besonders gut widerstehen konnte.[16][37]
Diese Interpretation wird aber zumeist abgelehnt, da aus funktionsmorphologischer Sicht eine vollständige Bipedie von Megatherium eher unwahrscheinlich ist. Eine alternative Erklärung der Spuren von Pehuén-Có wäre, dass die größeren Hinterfüße die Spuren der kleineren Vorderfüße jeweils überdecken, wie es auch an Spurenfossilien von Paramylodon aus Nevada beobachtet wurde.[40][41] Die Diskussion über die Bipedie von Megatherium beschränkt sich meist auf das große, in der Pamparegion verbreitete M. americanum. Für die Arten aus der Andenregion wird hingegen als Anpassung an das Gelände des Hochgebirges eine mehr oder weniger dauerhafte vierfüßige Fortbewegung angenommen. Dies lässt sich unter anderem an den nach oben verschobenen und prominenter herausragenden Gelenkflächen des Hinterhauptsbeins, an dem nur relativ flachen Oberschenkelknochen und an den teils stärker eingedrehten Hinterfüßen bei einigen Arten sowie an der Gestaltung der Hand belegen. So sind die beiden äußeren Mittelhandknochen, also diejenigen, die beim vierfüßigen Gang von Megatherium den Boden berühren, nahezu gleich lang und besitzen oberflächliche Aufrauungen, was auf einen häufigen Kontakt mit dem Untergrund hinweist.[42][4][12]
Ernährung
Megatherium gilt allgemein als Pflanzenfresser. Die hochkronigen Zähne würden eine Spezialisierung auf harte Grasnahrung indizieren, wie es bei zahlreichen heutigen Huftieren mit derartigen Zähnen der Fall ist. Aufgrund des Fehlens des Zahnschmelzes lassen sich detaillierte Studien zu Abnutzungsspuren nicht durchführen. Der Bau des Kauapparates widerspricht aber einer stärkeren Spezialisierung auf Gräser. So befürworten etwa die sehr hohen Gelenkfortsätze des Unterkiefers, die deutlich nach hinten versetzte Zahnreihe, aber auch der abwärts führende Knochenauswuchs am vorderen Jochbogen, an dem der Musculus masseter ansetzt, starke vertikale Kaubewegungen. Die bilophodonten Zähne mit ihren beiden querstehenden, scharfen Leisten lassen weiterhin vermuten, dass die Nahrung eher zerschnitten als aufwendig zerkaut wurde. Im Vergleich zu anderen Faultieren und in Bezug auf seine Körpergröße besaß Megatherium eine große Gesamtkaufläche aller Backenzähne, die etwa 10.500 bis 11.100 mm² umfasste. Das entspricht etwa dem zehnfachen Wert an Fläche, die das nahezu gleich große Lestodon aus der Gruppe der Mylodontidae aufwies, und liegt im Bereich heutiger Elefanten. Die effektive Vergrößerung der Kaufläche war dabei hauptsächlich das Resultat der Ausbildung der Querleisten auf den Backenzähnen. Die schneidende Wirkweise der Zähne führte aber gegenüber der mahlenden dazu, dass beim Kauen die Zellwände nicht immer aufgebrochen wurden und so weniger Nährstoffe direkt zur Verfügung standen. Jedoch konnte Megatherium aufgrund der großen Gesamtkaufläche mehr Nahrung gleichzeitig im Mundraum verarbeiten.[43]
Ob bei der Nahrungsaufnahme eine lange, bewegliche Zunge eingesetzt wurde, wie häufig angenommen, ist zweifelhaft. Der Bau des Zungenbeins ist bei Megatherium hochmodifiziert, der dort ansetzende Musculus geniohyoideus muss aufgrund der kurzen Distanz zur lang nach hinten gezogenen Symphyse des Unterkiefers sehr kurz gewesen sein, was beides gegen eine sehr mobile Zunge spricht.[44] Allerdings zeigen Rekonstruktionen der insgesamt schmalen Schnauze, dass höchstwahrscheinlich eine bewegliche Oberlippe ausgebildet war, ähnlich wie beim heutigen Spitzmaulnashorn. Diese bewegliche Lippe fungierte als Greiforgan bei der Nahrungsaufnahme und wird durch mehrere aufgeraute Flächen als Muskelansatzstellen nahe dem Augenfenster und dem Foramen infraorbitale sowie im Bereich des Oberkiefers und des Mittelkieferknochens angezeigt.[45]
Der Bau des Gebisses und der Kaumuskulatur von Megatherium sprechen für eine Ernährung von eher weicher Pflanzenkost.[10] Dies bestätigen auch Funde von Koprolithen von Megatherium, die neben Resten von Meerträubel auch solche von Fabiana, Acantholippia, Junellia und Chuquiraga enthielten.[46] Allerdings ergaben Isotopenuntersuchungen an Gebissresten aus der westargentinischen Provinz Mendoza, die in die Spätphase des Pleistozäns datieren, eine eher gemischte Pflanzenkost bestehend aus Blättern und Gräsern. Hier wird angenommen, dass die Tiere sich möglicherweise in ihrem Ernährungsverhalten den ökologischen Bedingungen der Region anpassten. Diese unterlag im Ausgang der letzten Kaltzeit größeren Veränderungen und war durch einen stärkeren montanen Einfluss geprägt.[47] Ähnliche Ergebnisse erbrachte eine Studie an Fossilresten aus der argentinischen Pamparegion, bei der auch Vergleichsmaterial von heutigen Säugetieren herangezogen wurden. Die entsprechenden Daten der Kohlenstoffisotope (δ13C) sprechen hier von einer rein pflanzlichen Ernährung, die recht hohen Werte bei den Stickstoffisotopen werden auf das trockene Klima zurückgeführt.[48] Untersuchungen von Abrasionsspuren an den Zähnen von Individuen aus dem nördlichen Argentinien ergaben bisher kein eindeutiges Bild zum Ernährungsverhalten von Megatherium in Bezug auf eine stärkere Bevorzugung von harter oder weicher Pflanzenkost. Die dabei bemerkte hohe Anzahl an Schliffspuren, die eigentlich typisch für Tiere mit Anpassung an eine eher härtere Pflanzenkost ist, wird ebenfalls auf das trockene Klima und der daraus resultierenden stärkeren Staubentwicklung in der Pamparegion zurückgeführt, was zu einer schnelleren Abnutzung der Zähne führte.[49]
Entgegen den oben genannten Analysen wurde vor allem im Übergang vom 20. zum 21. Jahrhundert auch die Ansicht vertreten, dass Megatherium sich von Fleisch ernährte und als Kleptoparasit lebte, der großen Raubtieren ihre Beute streitig machte oder als Aasfresser frisch verendete Kadaver nutzte. Erklärt wurde dies damit, dass das Vorderbein aufgrund der Beschaffenheit des Olecranon nicht zum Graben geeignet war, dafür allerdings für hohe und kraftvolle Geschwindigkeiten. Demnach konnte ein Tier mit einer Bewegung des Arms, dessen Gewicht mit etwa 115 kg veranschlagt wird, etwa 2700 J. freisetzen (vergleichbar mit der Energie, die beim Aufprall eines 10 kg schweren Objekts aus 20 m Höhe entsteht). Die so erreichte Kraft war ausreichend, um große Kadaver zu öffnen. Zudem besaß Megatherium lange scharfe Krallen an den drei mittleren Fingern (II bis IV), die außerdem seitlich verschmälert waren, und so das Abreißen von Fleisch befördert haben könnten. Demnach wäre es möglich, dass die Krallen neben einem Einsatz bei innerartlichen Rivalenkämpfen auch zum Erbeuten von Fleisch dienten.[39] Als Hinweis auf eine teils fleischliche Ernährung gilt auch eine Rippe eines großen Faultiers oder Rüsseltiers, auf der sich Nagespuren befinden, die typische Marken der bilophodonten Zähne von Megatherium zeigen. In der Regel gelten die Faultiere als weitgehend pflanzenfresserisch,[40] doch ernähren sich auch heutige Pflanzenfresser gelegentlich von Fleisch.[50]
Weichteilmorphologie und Pathologien

In zahlreichen Lebendrekonstruktionen wird Megatherium mit einem dichten Fell dargestellt. Begünstigt wird diese Vorstellung dadurch, dass einerseits die heutigen Baumfaultiere ebenfalls über Fell verfügen, andererseits von ausgestorbenen Faultieren wie Mylodon oder Nothrotheriops fossilisierte Fellreste vorliegen. Die enorme Größe der Tiere mit bis zu 6 t Körpergewicht und ihre Verbreitung in eher temperierten Regionen Südamerikas macht dies zumindest für die größten Vertreter wie M. americanum eher unwahrscheinlich. Hier spricht die notwendige Thermoregulation eines derartig großen Körpers zur besseren Ableitung von Wärme und Vermeidung von deutlicher Überhitzung eher für eine mehr oder weniger nackte Haut wie es auch bei anderen großen Säugetieren wie Elefanten, Nashörnern oder Flusspferden der Fall ist. Eine dichte Fellbedeckung würde eine enorme Menge an Trinkwasser voraussetzen, um den Wärmehaushalt auszugleichen, was aber in der gebietsweise semiariden Pamparegion Südamerikas nur schwer zur Verfügung gestanden haben dürfte.[51]
Ungeklärt ist die Frage, ob Megatherium ähnlich den großen Vertretern der Mylodontidae über Osteoderme in der Haut verfügte. Die Knochenplättchen sind bei den Mylodonten regellos verteilt, ähneln etwa denen der knöchernen Panzer der Gürteltiere und Glyptodontidae, haben aber eine einfachere Struktur. Gelegentlich werden solche Bildungen auch für Megatherium (und für das verwandte Eremotherium) beschrieben, für ersteres etwa von der Fundstelle Campo Laborde in der Pampasregion Argentiniens.[52] Da an den meisten Fundstellen mit Resten von Megatherium auch verschiedene Mylodonten vergesellschaftet sind und zudem auch Glyptodonten relativ häufig vorkommen, fehlt bisher ein eindeutiger anatomischer Beweis dafür.[53]
Krankheitsbedingte Knochenveränderungen sind bei Megatherium wie bei zahlreichen anderen Megatherien häufig zu finden. Sie betreffen meist die Wirbelsäule. Häufig treten sie an Schwanz und Becken auf. Dabei ist unter anderem das Typusskelett von M. celendinense von zahlreichen pathologischen Veränderungen überprägt, die nicht nur beide genannten Skelettabschnitte betreffen, sondern auch den Schädel und Rippen. Bei einem Fund von M. urbinai tritt weiterhin noch eine auf Osteoarthrose zurückzuführende Knochenschwellung auf, die möglicherweise auf einer Fraktur beruht.[12][4]
Systematik
|
Megatherium ist eine Gattung aus der ausgestorbenen Familie der Megatheriidae. Diese umfassen mittelgroße bis sehr große Vertreter aus der Unterordnung der Faultiere (Folivora). Innerhalb der Faultiere bilden die Megatheriidae wiederum zusammen mit den Megalonychidae und den Nothrotheriidae eine enger verwandte Gruppe, die Überfamilie Megatherioidea. In einer klassischen Auffassung, gestützt durch skelettanatomische Studien formen die Megatherioidea eine der beiden großen Faultierlinien, die andere findet sich in den Mylodontoidea. Molekulargenetische Analysen und Untersuchungen zur Proteinstruktur etablieren mit den Megalocnoidea eine dritte Linie. Den Ergebnissen der letzteren Studien zufolge gehören mit den Dreifinger-Faultieren (Bradypus) auch eine der beiden heute noch bestehenden Faultierlgattungen zu den Megatherioidea.[55][56] Zu den nächsten Verwandten von Megatherium innerhalb der Megatheriidae zählt das ähnlich große Eremotherium, welches das Schwestertaxon darstellt, im Gegensatz zu Megatherium aber eher die tropisch geprägten Tieflandgebiete Südamerikas bewohnte und auch nach Nordamerika vordrang. Gleichfalls in eine nähere Verwandtschaft gehören die stammesgeschichtlich älteren Vertreter Pyramiodontherium und Megatheriops. Alle diese Formen bilden die Unterfamilie der Megatheriinae, die die entwickelten großen Bodenfaultiere innerhalb der Megatheriidae stellen.[57][3]
Die Gattung gliedert sich in zwei Untergattungen mit heute insgesamt neun bekannten Arten:[4][1][12][28][3][24]
- Untergattung Megatherium Cuvier, 1796
- M. americanum Cuvier, 1796
- M. altiplanicum Saint-André & De Iuliis, 2001
- Untergattung Pseudomegatherium Kraglievich, 1931
- M. sundti Philippi, 1893
- M. urbinai Pujos & Salas, 2004
- M. celendinense Pujos, 2006
- M. elenense Hoffstetter, 1952
- M. medinae Philippi, 1893
- M. tarijense Gervais & Ameghino, 1880
- M. filholi Moreno, 1888
Bis auf M. americanum, die bekannteste und am besten untersuchte Art, sind alle anderen Vertreter bisher nur von wenig Fundmaterial bekannt. Die Untergattung Pseudomegatherium umfasst die weitgehend in den Anden oder im schmalen Küstenstreifen zum Pazifik nachgewiesene Arten. Sie unterscheidet sich von Megatherium, und da vor allem von dem bisher bekannten hauptsächlichen Vertreter M. americanum, durch eine Reduktion der Körpergröße um 20 bis 60 % – allerdings ist mit M. celendinense auch eine Art bekannt, die fast die Größe von M. americanum erreichte. Weitere Unterschiede sind die etwas abweichende Gestaltung der Gelenkflächen am Hinterhauptsbein für den Ansatz der Halswirbelsäule (höher gelegen und prominenter herausragend), kürzere Schnauzen, eine etwas andere Gestaltung der Hand mit zwei gleichlangen, äußeren Mittelhandknochen, flachere Oberschenkelknochen und bestimmte Merkmale am Sprungbein. Ein Teil dieser Eigenschaften, so die Form und Lage der Hinterhauptscondylen sowie die Gestaltung des Femurs oder der Hand, werden mit besonderen Anpassungen an die geographischen Verhältnisse der Anden in Verbindung gebracht, etwa einer stärkeren bis dauerhaft vierfüßigen Bewegung.[42][12]
Neben den heute anerkannten Arten wurden noch weitere benannt beziehungsweise beschrieben, etwa M. lundii, M. gaudryi, M. cuvieri oder M. parodii, sie gelten aber meist als synonym zu M. americanum. Uneinigkeit herrscht bei M. gallardoi, einem sehr großen Vertreter von Megatherium, der 1921 von Carlos Ameghino und Lucas Kraglievich anhand eines in das Mittelpleistozän datierenden, 83 cm langen, allerdings zahnlosen Schädels aus Buenos Aires beschrieben worden war.[9] Hier sehen einige Autoren die Art als valide an und heben unter anderem den kaum mit dem Oberkiefer verwachsenen Mittelkieferknochen und die geringere Höhe des Unterkieferkörpers resultierend aus einer geringeren Hypsodontie der Backenzähne als besondere Kennzeichen hervor.[8] Andere Wissenschaftler führen diese Merkmale wiederum auf die innerartliche Variabilität von M. americanum zurück, womit nach dieser Auffassung die Art der einzige Vertreter seiner Gattung in der Pamparegion wäre.[5] Problematisch ist die aus dem Unterpliozän stammende Art M. antiquum, die 1885 von Florentino Ameghino mit Hilfe isolierter Zähne eingerichtet wurde. Die Zähne ähneln denen von M. americanum, erreichen aber nur ein Drittel der Größe. Die Eigenständigkeit der Art wird angezweifelt, da das nur wenige Stücke umfassende Fundmaterial kaum diagnostische Merkmale erkennen lässt.[58][59]
Stammesgeschichte
Ursprünge
Über den Ursprung der Gattung Megatherium gibt es mehrere Theorien. Einerseits wird eine Entstehung allgemein im südlichen Teil Südamerikas angenommen. Von dort breitete sich die Gattung weiter nach Westen aus und erreichte später im Pleistozän im Andenraum ihre höchste Diversität. Diese Annahme ist kongruent zu den häufigen und mit rund einem halben Dutzend Gattungen recht vielfältigen Nachweisen pliozäner und obermiozäner Megatherien im zentralen und südlichen Südamerika.[60][54] So treten allein im obermiozänen Conglomerado osífero im unteren Abschnitt der Ituzaingó-Formation, die an den Uferbänken des Unterlaufs des Río Paraná nahe der argentinischen Stadt Paraná aufgeschlossen ist, mit Eomegatherium, Pliomegatherium, Pyramiodontherium und Promegatherium vier Gattungen auf.[58] Unterstützend zu dieser eher südlichen Herkunft datiert ein nur 28 cm langer Schädel eines Jungtiers von Megatherium aus der Chapadmalal-Formation bei San Eduardo del Mar in der argentinischen Pamparegion auf ein Alter von 3,58 Millionen Jahren und damit in die zweite Hälfte des Pliozäns.[61] Eine weit nördlichere Entstehung kann aber auch nicht ausgeschlossen werden, allerdings fehlen häufig Aufschlüsse aus dieser Zeit im nördlichen Südamerika. Die dort seit dem Beginn des 21. Jahrhunderts aufgefundenen Vertreter der späten Megatherien aus dem Übergang vom Miozän zum Pliozän, etwa Urumaquia und Proeremotherium,[62][60] lassen dabei nicht zwingend annehmen, dass die taxonomische Vielfalt hier geringer war als in den südlicheren Regionen.[46] Als dritte Möglichkeit besteht ein Ursprung von Megatherium im Andengebiet, was wiederum übereinstimmend wäre mit der höchsten hier nachgewiesenen Artenvielfalt.[63] Zudem ist mit M. altiplanicum auch einer der ältesten Nachweise der Gattung in dieser Region überliefert, der in den Beginn des Pliozäns datiert, wobei radiometrische Daten ein Alter zwischen 5,4 und 2,8 Millionen Jahren angeben.[1][63]
Im Pleistozän sind dann rund ein halbes Dutzend Arten aus der Andenregion überliefert, ihre zeitliche Einordnung ist aufgrund der wenigen Funde teilweise nur ungenau möglich.[8] Aus der Pamparegion wiederum stammt die bedeutendste und häufigste Art, M. americanum. Diese war dort weit verbreitet und trat wenigstens seit dem frühen Mittelpleistozän auf. Ihr Vorkommen wird in der lokalen Stratigraphie durch die Faunenstufe der Megatherium americanum biozone angezeigt, welche mit dem Beginn des Mittelpleistozäns vor rund 780.000 Jahren zusammenfällt und die die vorhergehende Tolypeutes pampaeus biozone ablöste, benannt nach einem heute ausgestorbenen Kugelgürteltier.[64]
Aussterben und Einfluss des Menschen
Der überwiegende Teil der Funde stammt aber aus dem Oberen Pleistozän. Ihr können auch die jüngsten Funde zugeschrieben werden, die in den Ausgang des Pleistozäns und in das frühe Holozän datieren. Sie gehören damit in die Zeit der Ankunft der ersten menschlichen Bewohner Südamerikas, die erstmals um etwa 14.800 vor heute nachweisbar sind, und zugleich in die Phase der quartären Aussterbewelle, der zahlreiche Großsäugetiere weltweit zum Opfer fielen. Inwiefern die frühen Besiedler Amerikas direkt am Aussterben der riesigen Faultiergattung beteiligt waren, ist in der Forschung umstritten. Hinweise auf Interaktionen zwischen Mensch und Megatherium liegen bisher nur wenige vor. Bedeutend in diesem Zusammenhang sind unter anderem die Funde von Arroyo Seco, einer mehrphasigen Freilandfundstelle in der Provinz Buenos Aires in Argentinien. In den unteren Fundhorizonten, die Radiocarbondatierungen zufolge zwischen 10.500 und 12.240 Jahren vor heute datieren, wurden neben Hinterlassenschaften von menschlichen Jäger-Sammler-Gruppen auch Reste von Megatherium gefunden, was die Jagd auf oder die Verwendung der Reste des riesigen Bodenfaultiers annehmen lässt.[65] Der archäologische Fundplatz Paso Otero 5 in der gleichen Provinz barg auf knapp 100 m² gut 80.000, zum großen Teil stark zerschlagene Knochenreste. Ihr dadurch fragmentierter Zustand lässt eine genaue Bestimmung nur eingeschränkt zu, jedoch wurden insgesamt 29 Exemplare Megatherium zugeordnet. Assoziiert waren die Faunenreste mit über 80 Steinartefakten aus Quarzit, darunter auch einzelne Fischschwanz-Spitzen (fish tail points). Das Alter des Fundplatzes wurde mit Hilfe der Radiokarbonmethode auf 10.440 bis 9.400 Jahre BP bestimmt. Als Besonderheit erwies sich ein Fersenbein von Megatherium, das von zahlreichen gebrannten Knochen umgeben war; sie werden als Brennmaterial interpretiert.[66][67][68][69]
In den Übergangszeitraum vom Oberen Pleistozän zum Unteren Holozän wird Campo Laborde eingeordnet, ebenfalls in der argentinischen Provinz Buenos Aires gelegen. Neben über 100 Abschlägen aus Quarzit als menschliche Relikte wurden bisher auch mehr als 99.000 Knochenfunde gemacht, darunter 108 zu zählende Reste von Megatherium, die einem Teilskelett ohne Schädel eines einzelnen Individuums angehören. Die Knochen insgesamt sind teilweise stark zerschlagen. Einzelne Rippen von Megatherium besitzen Schnittmarken und wurden teilweise auch zu Werkzeugen modifiziert. Insgesamt zeigt der Befund, dass das Skelett eindeutig von Menschen zerlegt wurde. Radiocarbonmessungen an einzelnen Knochen des Faultiers ergaben ursprünglich ein Alter von 7750 bis 8700 Jahre vor heute.[52] Sie ließen somit ein Überleben von Megatherium bis in das Untere Holozän vermuten, womit die Faultiergattung im Gegensatz zu zahlreichen anderen großen Bodenfaultieren sowohl die erste Ankunft des Menschen in Südamerika als auch die rapiden Klimaänderungen zum Ausgang der letzten Kaltzeit um mehrere Tausend Jahre überlebt hätte.[70][71] Neuere Datierungen aus dem Jahr 2019 erbrachten allerdings mit 10.250 bis 12.730 Jahren vor heute deutlich höhere Alterswerte.[72]
Zu den wenigen Spuren der Manipulation von Knochen von Megatherium durch den Menschen gesellt sich noch ein erster Halswirbel, der einzelne Schnittmarken von Steinartefakten aufweist. Er wurde in der Pampasregion entdeckt und gelangte Mitte des 19. Jahrhunderts in das Museo di Storia Naturale in Florenz. Es fehlen allerdings Hinweise zur genauen stratigraphischen Fundposition und somit zum Fundkontext, wodurch der Knochenrest als problematisch zu erachten ist.[73]
Forschungsgeschichte
Cuvier und das Megatherium

Der nachweislich früheste Fund von Megatherium ist ein nahezu vollständiges Skelett, welches nach unterschiedlichen Angaben zwischen 1787 und 1789 von Manuel Torres, einem Dominikaner, in Luján an den Ufern des Río Luján in der heutigen argentinischen Provinz Buenos Aires entdeckt worden war. Nicolás del Campo, damaliger Vizekönig von Río de la Plata, verschiffte das Skelett verpackt in sieben Kisten nach Spanien an das Real Gabinete de Historia Natural de Madrid, wo es am 29. September 1789 ankam. Dort nahm es Juan Bautista Bru de Ramón (1740–1799) in Empfang. Dieser arbeitete dort als Präparator und erstellte Skelettrekonstruktionen von verendeten Tieren für das naturhistorische Kabinett, so zum Beispiel von Elefanten. Er begann unmittelbar mit der Arbeit und beendete die Rekonstruktion vier Jahre später mit der Stellung des Skeletts in einer aus heutiger Sicht eher unnatürlichen Pose. Allerdings handelte es sich bei Brus Skelettrekonstruktion von Megatherium aus einer naturhistorischen Betrachtung heraus um die erste eines ausgestorbenen Wirbeltiers weltweit. Neben der Aufstellung des Skeletts fertigte Bru auch eine Monographie mit Beschreibung des Skelettes an, die auch 22 Abbildungen enthielt, eine von der Rekonstruktion und die restlichen über einzelne Knochen, die von Manuel Navarro gezeichnet worden waren. Allerdings wurde die Beschreibung nicht öffentlich publiziert, jedoch übergab man noch im Jahr 1793 einige Kopien der Zeichnungen einem französischen Repräsentanten.[22][6]

Die Kopien gelangten in die Hände des französischen Anatomen Georges Cuvier (1769–1832), der damals am Muséum national d’histoire naturelle in Paris arbeitete und über den Fund einen Bericht anfertigen sollte. Dieser erschien 1796 in der Zeitschrift Magasin encyclopédique, worin er dem riesigen Tier die wissenschaftliche Bezeichnung Megatherium americanum gab. Der Bericht gilt somit als Erstbeschreibung von Gattung und Art.[74] Der Gattungsname leitet sich von den griechischen Wörtern μέγας (mégas „groß“) und θηρίον (thērion „Tier“) her und bezieht sich auf die Größe des Faultiers. Cuvier nutzte bei seiner Beschreibung die für seine Zeit neue Methodik der vergleichenden Anatomie, wobei er die Zeichnungen als Grundlage nahm, das spanische Skelett hatte Cuvier selbst nie persönlich in Augenschein genommen. In seinem Bericht bezeichnete er die Herkunft des Skelettes fälschlicherweise mit „Paraguay“, doch ist Cuviers Artikel insofern bedeutend, als dass es sich um seine erste Arbeit über ausgestorbene Tiere handelt. Auch bemerkte Cuvier, dass ähnliche Tiere wie Megatherium heute nicht mehr existierten und damit ausgestorben waren, was damals eine revolutionäre Ansicht darstellte.[22][6][75]
Cuviers Bericht wurde von José Garriga nach Spanien gebracht, der ihn dort übersetzen wollte. Dabei fand er heraus, dass Bru bereits einen Aufsatz angefertigt hatte und publizierte ihn zusammen mit Cuvier im gleichen Jahr unter seiner Herausgeberschaft und auf eigene Kosten.[76] Dieses Buch bereitete den Weg für eine weitere Publikation von Georges Cuvier, die 1804 unter dem Titel Sur le Megatherium erschien und in dem er die Verwandtschaft mit den Faultieren weiter herausarbeitete. Für Cuvier sprachen primär die Merkmale des Schädels, so der Bau der Jochbögen, und der Schulterregion, wie das ausgebildete Schlüsselbein, für eine Beziehung zu den heutigen baumbewohnenden Faultieren, sekundär die Struktur des Gebisses und die Gestaltung der Gliedmaßen. Bei den Gliedmaßen bemerkte Cuvier, dass diese eher den heutigen Ameisenbären und Gürteltieren ähnelten und deutlich kürzer waren als bei den rezenten Faultieren. Später, im Jahr 1812, fügte er seinen zweiten Aufsatz und Brus Abbildungen in sein Werk Recherches sur les ossemens fossiles ein,[77] das zu den Grundlagenwerken zur Entwicklung der Paläontologie als wissenschaftliche Disziplin gehört.[22][6][75]
Darwin und Owen

Seit der Bergung des ersten Skeletts von Megatherium bei Luján sollte dies über mehr als vier Dekaden der einzige Fund dieses riesigen Bodenfaultiers bleiben. Zu Beginn der 1830er Jahre kam bei Villanueva am Río Salado, der die Pampaebenen südlich von Buenos Aires durchfließt, ein weiteres Teilskelett zu Tage, das bei Niedrigwasser nach einer ausgesprochen langen Trockenphase aus dem Fluss ragte. Die Fossilreste wurden geborgen und vom hochrangigen britischen Diplomaten Woodbine Parish nach England verschickt, wo sie nach eingehender Präparation William Clift am Royal College of Surgeons vorstellte und 1835 publizierte. Da Parish davon ausging, dass in der Pamparegion noch weitere Skelette aufzufinden seien, bat er den damaligen Gouverneur von Argentinien, Juan Manuel de Rosas, um Beihilfe bei der Suche, was in der Folgezeit zur Entdeckung zweier weiterer Skelette führte.[78] Von hoher Bedeutung für die Erforschung von Megatherium ist die Reise von Charles Darwin (1809–1882), die er mit der HMS Beagle zwischen 1831 und 1837 unternahm. Er entdeckte Ende August 1832 in felsigen Aufschlüssen an der Küste bei Punta Alta nahe Bahía Blanca im heutigen Argentinien zahlreiche neue Funde der Gattung. Da die Bordbibliothek der Beagle auch einen übersetzten Text mit Abbildung von Cuviers Beschreibung enthielt, war Darwin vertraut mit Megatherium, allerdings bezeichnete er in seinen Tagebüchern der Reise zahlreiche unterschiedliche Fossilien als zu Megatherium gehörig. Darwin besuchte die Region erneut im Oktober 1833 und erreichte auch Luján, von wo das erste Megatherium-Skelett stammte.[20][79]
Die Funde, die während der Reise gemacht worden waren, wurden nach England ins Royal College of Surgeons in London geschickt, wo sich ab 1836 der bedeutende Anatom Richard Owen (1804–1892) mit diesen zu beschäftigen begann. Im Jahr 1840 veröffentlichte Owen seine erste Abhandlung über das Riesenfaultier, das er als M. cuvieri bezeichnete[80] (ein Name, der 1822 von Anselme Gaëtan Desmarest als Alternativname für M. americanum eingeführt wurde, aber als nomen illegitimum gilt[21]). Zwischen 1851 und 1860 veröffentlichte Owen eine Serie von Monographien über Megatherium, betitelt jeweils mit On the Megatherium (Megatherium americanum, Cuvier and Blumenbach), die sich jeweils unterschiedlichen Skelettpartien widmeten.[81] Sein Abschlusswerk erschien 1861 und nannte sich Memoir on the Megatherium or giant Ground-sloth of America (Megatherium americanum, Cuvier).[82] So erkannte Owen, dass Megatherium aufgrund seiner Größe terrestrisch lebte und sein Gewicht auf die Außenstrahlen der Hände und Füße legte (pedolateral). Er rekonstruierte auch, dass das Tier befähigt war, sich auf den Hinterbeinen aufzurichten, wobei es sich dabei mit dem Schwanz abstützte, um mit den Vorderbeinen nach Nahrung zu suchen. Aufgrund der präzisen Darstellungen und Zeichnungen gelten diese Abhandlungen noch heute als Standardwerke über Bodenfaultiere. Das dadurch entstandene Bild über Megatherium prägte nachhaltig die Öffentlichkeit. Aufgrund des Enthusiasmus, den Owen für das Faultier an den Tag legte, wurde er mehrfach auch karikiert.[6][83]
Larrañaga und das gepanzerte Riesenfaultier
Eine weitere Episode, die das Bild über Megatherium zumindest im Verlauf des 19. Jahrhunderts nachhaltig prägte, war die Entdeckung eines Rücken- und Schwanzpanzers sowie eines Oberschenkelknochens, die nach heutiger Sicht zu einem Vertreter der Glyptodontidae, stark gepanzerten Verwandten der Gürteltiere, gehören; die Gruppe war damals aber noch nicht bekannt. Dàmaso Antonio Larrañaga (1771–1848), einer der damals führenden Naturforscher in den spanischen Kolonien, fertigte darüber im Jahr 1814 eine wissenschaftliche Beschreibung in seiner Schrift Diario de Historia Natural an und fügte dieser die Bezeichnung Dasypus (Megatherium Cuv) bei. Die Angabe von Megatherium als Untergattung für die heutigen Langnasengürteltiere führte dazu, dass in der Folgezeit die Vorstellung über gepanzerte Riesenfaultiere entstand. Cuvier persönlich gab Larrañagas taxonomische Bezeichnung 1824 Anlass, diese auch in der zweiten Auflage seines Werkes Recherches sur les ossemens fossiles einzufügen.[84][85] Die Idee über gepanzerte Riesenfaultiere war weit verbreitet, so hatte Clift 1835 bei der Beschreibung des zweiten Skelettfundes von Megatherium die mit den Resten assoziierten Panzerteile dem Faultier zugewiesen.[78] Selbst Darwin notierte auf seiner Beagle-Reise mehrfach Funde von Panzerfragmenten von Megatherium, teilweise ordnete er bestimmte Reste nur aufgrund von Beifunden von Knochenplättchen der Faultiergattung zu.[21][79]
Zwar wurden bereits in den 1830er Jahren Einwände gegen die Panzerung von Megatherium erhoben, so unter anderem 1833 von Joseph Eduard d’Alton anhand von Panzer- und Knochenfunden aus Uruguay und Brasilien, die er nach intensiven anatomischen Vergleichen mit riesigen Gürteltieren in Verbindung brachte.[86] Die Beschreibung von Hoplophorus durch Peter Wilhelm Lund im Jahr 1838[87] und von Glyptodon durch Owen im Jahr darauf,[88][89] beide zeigten als Vertreter der Glyptodontidae starke Ähnlichkeiten zu Larrañagas Dasypus (Megatherium Cuv), bewiesen aber letztendlich, dass die bisher bekannten Panzerfunde ganz anderen, aber ebenfalls riesigen Tieren angehörten. Owen revidierte in einer umfangreichen Beschreibung von Glyptodon im Jahr 1841 noch einmal die mit Megatherium verbundenen Panzerreste und korrigierte ihre tatsächliche Zugehörigkeit.[89] Im Jahr 1865 wies der englische Paläontologe Thomas Henry Huxley (1825–1895) in einer Schrift über die Skelettanatomie von Glyptodon noch einmal auf die eindeutigen Belege der unterschiedlichen Herkunft beider Fossilgruppen hin.[90] Anzumerken ist hierbei, dass es jedoch tatsächlich große Bodenfaultiere gab, in deren Haut Knochenplättchen ausgebildet waren, doch formten diese nie einen festen Panzer. Zudem gehören Faultiere allesamt einer anderen Linie an (Mylodontidae), die mit den Megatheriidae eher entfernt verwandt ist. Die Existenz von ähnlich locker gestreuten Osteodermen in der Haut bei Megatherium ist darüber hinaus umstritten.[91][53]
Die Wende zum 20. Jahrhundert
Zum Ende des 19. und zu Beginn des 20. Jahrhunderts waren vor allem die italienischstämmigen Gebrüder Florentino (1854–1911) und Carlos Ameghino (1865–1936) bedeutend für die paläontologische Forschung in Südamerika. Sie beschrieben meist unabhängig voneinander mehrere Arten von Megatherium, von denen aber nur M. tarijense aus dem Jahr 1880 bis heute gültig ist. Die Beschreibung der Art erfolgte weit nach der Entdeckung und Erstpublikation der Funde im Jahr 1855 durch Paul Gervais, sie zeigte aber auf, dass es auch Vertreter des Riesenfaultiers gab, die deutlich kleiner als M. americanum waren. Damit war zudem der erste Nachweis der Untergattung Pseudomegatherium erbracht.[3] Gut ein Dutzend Jahre später folgten dann mit M. sundti und M. medinae durch Rudolph Amandus Philippi zwei weitere Arten.[25][26] In der ersten Hälfte des 20. Jahrhunderts prägten dann Lucas Kraglievich (1886–1932) und Robert Hoffstetter (1908–1999) die Forschung nachhaltig.[83]
Literatur
- Paul S. Martin und Richard G. Klein (Hrsg.): Quaternary Extinctions. A Prehistoric Revolution. The University of Arizona Press, Tucson AZ 1984, ISBN 0-8165-1100-4.
- Richard A. Fariña, Sergio F. Vizcaíno und Gerardo De Iuliis: Megafauna. Giant beasts of Pleistocene South America. Indiana University Press, 2013, ISBN 978-0-253-00230-3, S. 254–256.
Einzelnachweise
- Pierre-Antoine Saint-André und Gerardo De Iuliis: The smallest and most ancient representative of the genus Megatherium Cuvier, 1796 (Xenarthra, Tardigrada, Megatheriidae), from the Pliocene of the Bolivian Altiplano. Geodiversitas 23 (4), 2001, S. 625–645.
- Carlos Castor Cartelle: Preguiças terrícolas, essas desconhecidas. Anales Ciência Hoje 27, 2000, S. 19–25.
- Gerardo De Iuliis, François Pujos und Giuseppe Tito: Systematic and Taxonomic Revision of the Pleistocene Ground Sloth Megatherium (Pseudomegatherium) Tarijense (Xenarthra: Megatheriidae). Journal of Vertebrate Paleontology 29 (4), 2009, S. 1244–1251.
- François Pujos und Rodolfo Salas: A new species of the genus Megatherium (Mammalia: Xenarthra: Megatheriidae) from the Pleistocene of Sacaco and Tres Ventanas. Palaeontology 47 (3), 2004, S. 579–604.
- Fariña, Vizcaíno und De Iuliis: Megafauna. 2013, S. 213–216.
- Sergio F. Vizcaíno, M. Susanna Bargo und Richard A. Fariña: Form, function, and paleobiology in xenarthrans. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 86–99.
- M. Susana Bargo, Segio F. Vizcaíno, Fernando M. Archuby und R. Ernesto Blanco: Limb bone proportions, strength and digging in some Lujanian (Late Pleistocene-Early Holocene) mylodontid ground sloths (Mammalia, Xenarthra). Journal of Vertebrate Paleontology 20 (3), 2000, S. 601–610.
- Diego Brandoni, Esteban Soibelzon und Alejo Scarano: On Megatherium gallardoi (Mammalia, Xenarthra, Megatheriidae) and the Megatheriinae from the Ensenadan (lower to middle Pleistocene) of the Pampean region, Argentina. Geodiversitas 30 (4), 2008, S. 793–804.
- Carlos Ameghino und Lucas Kraglievich: Descripción del „Megatherium gallardoi“ C. amegh descubierto en el Pampeano inferior de la ciudad de Buenos Aires. Anales del Museo Nacional de Historia Natural de Buenos Aires 31, 1921, S. 134–156.
- M. Susana Bargo: The ground sloth Megatherium americanum: Skull shape, bite forces, and diet. Acta Palaeontologica Polonica 46 (2), 2001, S. 173–192.
- H. Gregory McDonald: Xenarthran skeletal anatomy: primitive or derived? Senckenbergiana biologica 83, 2003, S. 5–17.
- François Pujos: Megatherium celendinense sp. nov. from the Pleistocene of the Peruvian Andes and the relationships of megatheriines. Palaeontology 49 (2), 2006, S. 285–306.
- M. Susana Bargo, Gerardo De Iuliis und Sergio F. Vizcaíno: Hypsodonty in Pleistocene ground sloths. Acta Palaeontologica Polonica 51 (1), 2006, S. 53–61.
- Gerardo De Iuliis und Cástor Cartelle: A new giant megatheriine ground sloth (Mammalia: Xenarthra: Megatheriidae) from the late Blancan to early Irvingtonian of Florida. Zoological Journal of the Linnean Society 127, 1999, S. 495–515.
- Gerardo De Iuliis: Toward the morphofunctional understanding of the humerus of Megatheriinae: The identity and homology of some diaphyseal humeral features (Mammalia, Xenarthra, Megatheriidae). Senckenbergiana biologica 83, 2003, S. 69–78.
- Adrià Casinos: Bipedalism and quadrupedalism in Megatherium: an attempt at biomechanical reconstruction. Lethaia 29, 1996, S. 87–96.
- Gerardo De Iuliis und Cástor Cartelle: The medial carpal and metacarpal elements of Eremotherium and Megatherium (Xenarthra: Mammalia). Journal of Vertebrate Paleontology 14, 1994, S. 525–533.
- Diego Brandoni, Alfredo A. Carlini, Francois Pujos und Gustavo J. Scillato-Yané: The pes of Pyramiodontherium bergi (Moreno & Mercerat, 1891) (Mammalia, Xenarthra, Phyllophaga): The most complete pes of a Tertiary Megatheriinae. Geodiversitas 26 (4), 2004, S. 643–659.
- Renato Pereira Lopes und Jamil Corrêa Pereira: On the presence of Megatherium Cuvier, 1796 (Xenarthra, Pilosa) in fossilferous deposits of the coastal plain of Southern Brazil. Revista Brasileira de Paleontologia 22 (1), 2019, S. 38–52.
- Richard D. Keynes: The Jayne Lecture. From Bryozoans to Tsunami: Charles Darwin’s Findings on the Beagle. Proceedings of the American Philosophical Society 147 (2), 2003, S. 103–127.
- Juan Carlos Fernicola, Sergio F. Vizcaíno und Gerardo De Iuliis: The fossil mammals collected by Charles Darwin in South America during his travels on board the HMS Beagle. Revista de la Asociación Geológica Argentina 64 (1), 2009, S. 147–159.
- José M. López Piñero: Juan Bautista Bru (1740–1799) and the Description of the Genus Megatherium. Journal of the History of Biology 21 (1), 1988, S. 147–163.
- Agustín Scanferla, Ricardo Bonini, Lucas Pomi, Enrique Fucks und Alejandro Molinari: New Late Pleistocene megafaunal assemblage with well-supported chronology from the Pampas of southern South America. Quaternary International 305, 2013, S. 97–103.
- Federico L. Agnolin, Nicolás R. Chimento, Diego Brandoni, Daniel Boh, Denise H. Campo, Mariano Magnussen und Francisco De Cianni: New Pleistocene remains of Megatherium filholi Moreno, 1888 (Mammalia, Xenarthra) from the Pampean Region: Implications for the diversity of Megatheriinae of the Quaternary of South America. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 289/3, 2018, S. 339–348.
- R. A. Philippi: Vorläufige Nachricht über fossile Säugethierknochen von Ulloma, Bolivia. Zeitschrift der deutschen geologischen Gesellschaft 45, 1893, S. 87–96.
- R. A. Philippi: Noticias preliminares sobre huesos fósiles de Ulloma. Anales de la Universidad de Chile 82, 1893, S. 499–506.
- Daniel Frassinetti und Valeria Azcárate: Presencia de Megatherium en los alrededores de Santiago (Chile). Boletín del Museo Nacional de Historia Natural 33, 1974, S. 35–42.
- Gerardo De Iuliis: On the taxonomic status of Megatherium sundti Philippi, 1893 (Mammalia: Xenarthra: Megatheriidae). Ameghiniana 46 (1), 2006, S. 161–169.
- François Pujos und Rodolfo Salas: A systematic reassessment and paleogeographic review of fossil Xenarthra from Peru. Bulletin de l’Institut Français d’Etudes Andines 33, 2004, S. 331–378.
- Franz Spillmann: Beiträge zur Kenntnis eines neuen gravigraden Riesensteppentieres (Eremotherium carolinense gen. et sp. nov.), seines Lebensraumes und seiner Lebensweise. Palaeobiologica 8, 1948, S. 231–279.
- H. Gregory McDonald: Evolution of the Pedolateral Foot in Ground Sloths: Patterns of Change in the Astragalus. Journal of Mammal Evolution 19, 2012, S. 209–215.
- Néstor Toledo, Gerardo De Iuliis, Sergio F. Vizcaíno und M. Susana Bargo: The Concept of a Pedolateral Pes Revisited: The Giant Sloths Megatherium and Eremotherium (Xenarthra, Folivora, Megatheriinae) as a Case Study. Journal of Mammalian Evolution 25 (4), 2018, S. 525–537, doi:10.1007/s10914-017-9410-0.
- Giuseppe Tito und Gerardo De Iuliis: Morphofunctional aspects and paleobiology of the manus in the giant ground sloth Eremotherium Spillmann 1948 (Mammalia, Xenarthra, Megatheriidae). Senckenbergiana biologica 83 (1), 2003, S. 79–94.
- G. Billet, D. Germain, I. Ruf, C. de Muizon und L. Hautier: The inner ear of Megatherium and the evolution of the vestibular system in sloths. Journal of Anatomy 223, 2013, S. 557–567.
- Santiago Pantiño und Richard A. Fariña: Ungual phalanges analysis in Pleistocene ground sloths (Xenarthra, Folivora). Historical Biology 29 (8), 2017, S. 1065–1075, doi:10.1080/08912963.2017.1286653.
- Santiago Pantiño, Jorge Peréz Zerpa und Richard A. Fariña: Finite element and morphological analysis in extant mammal's claws and Quaternary sloth' ungual phalanges. Historical Biology, 2019, doi:10.1080/08912963.2019.1664504.
- R. Ernesto Blanco und Ada Czerwonogora: The gait of Megatherium Cuvier 1796 (Mammalia, Xenarthra, Megatheriidae). Senckenbergiana biologica 83 (1), 2003, S. 61–68.
- Silvia A. Aramayo, Teresa Manera de Bianco, Nerea V. Bastianelli und Ricardo N. Melchor: Pehuen Co: Updated taxonomic review of a late Pleistocene ichnological site in Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 439, 2015, S. 144–165.
- Richard A. Fariña und R. Ernesto Blanco: Megatherium, the Stabber. Proceedings: Biological Sciences 263 (1377), 1996, S. 1725–1729.
- François Pujos, Timothy J. Gaudin, Gerardo De Iuliis und Cástor Cartelle: Recent Advances on Variability, Morpho-Functional Adaptations, Dental Terminology, and Evolution of Sloths. Journal of Mammal Evolution 19, 2012, S. 159–169.
- Fariña, Vizcaíno und De Iuliis: Megafauna. 2013, S. 254–256.
- François Pujos, Rodolfo Salas und Jean Mattos: Andean lineage of Pleistocene Megatherium: Geographical implications. Journal of Vertebrate Paleontology 22 (3 suppl), 2002, S. 97A.
- Sergio F. Vizcaíno, M. Susana Bargo und Guillermo H. Cassini: Dental occlusal surface area in relation to body mass, food habits and other biological features in fossil xenarthrans. Ameghiniana 43 (1), 2006, S. 11–26.
- Leandro M. Pérez, Néstor Toledo, Gerardo De Iuliis, M. Susana Bargo und Sergio F. Vizcaíno: Morphology and Function of the Hyoid Apparatus of Fossil Xenarthrans (Mammalia). Journal of Morphology 271, 2010, S. 1119–1133.
- M. Susana Bargo, Néstor Toledo und Sergio F. Vizcaíno: Muzzle of South American Pleistocene Ground Sloths (Xenarthra, Tardigrada). Journal of Morphology 267, 2006, S. 248–263.
- H. Gregory McDonald und Gerardo de Iuliis: Fossil history of sloths. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 39–55.
- Angel Praderio, Adolfo Gil und Analía M. Forasiepi: El registro de Megatherium (Xenarthra, Tardigrada) en Mendoza (Argentina): Aspectos taxonómicos, cronológicos y paleoecológicos. Mastozoología Neotropical 19 (2), 2012, S. 279–291.
- Hervé Bocherens, Martin Cotte, Ricardo A. Bonini, Pablo Straccia, Daniel Scian, Leopoldo Soibelzon und Francisco J. Prevosti: Isotopic insight on paleodiet of extinct Pleistocene megafaunal Xenarthrans from Argentina. Gondwana Research 48, 2017, S. 7–14.
- Jeremy L. Green und Daniela C. Kalthoff: Xenarthran Dental Microstructure and Dental Microwear Analyses, with New Data for Megatherium americanum (Megatheriidae). Journal of Mammalogy 96 (4), 2015, S. 645–657.
- Fariña, Vizcaíno und De Iuliis: Megafauna. 2013, S. 256–259 und 304–310.
- Fariña, Vizcaíno und De Iuliis: Megafauna. 2013, S. 259–260.
- Gustavo G. Politis und Pablo G. Messineo: The Campo Laborde site: New evidence for the Holocene survival of Pleistocene megafauna in the Argentine Pampas. Quaternary International 191, 2008, S. 98–114.
- H. Gregory McDonald: An Overview of the Presence of Osteoderms in Sloths: Implications for Osteoderms as a Plesiomorphic Character of the Xenarthra. Journal of Mammalian Evolution 25 (4), 2018, S. 485–493, doi:10.1007/s10914-017-9415-8.
- François Pujos, Rodolfo Salas-Gismondi, Guillaume Baby, Patrice Baby, Cyrille Goillot, Julia Tejada und Pierre-Oliver Antoine: Implications of the presence of Megathericulus (Xenarthra: Tardigrada: Megatheriidae) in the Laventan of Peruvian Amazonia. Journal of Systematik Palaeontology 11 (7–8), 2013, S. 973–991.
- Frédéric Delsuc, Melanie Kuch, Gillian C. Gibb, Emil Karpinski, Dirk Hackenberger, Paul Szpak, Jorge G. Martínez, Jim I. Mead, H. Gregory McDonald, Ross D.E. MacPhee, Guillaume Billet, Lionel Hautier und Hendrik N. Poinar: Ancient mitogenomes reveal the evolutionary history and biogeography of sloths. Current Biology 29 (12), 2019, S. 2031–2042, doi:10.1016/j.cub.2019.05.043.
- Samantha Presslee, Graham J. Slater, François Pujos, Analía M. Forasiepi, Roman Fischer, Kelly Molloy, Meaghan Mackie, Jesper V. Olsen, Alejandro Kramarz, Matías Taglioretti, Fernando Scaglia, Maximiliano Lezcano, José Luis Lanata, John Southon, Robert Feranec, Jonathan Bloch, Adam Hajduk, Fabiana M. Martin, Rodolfo Salas Gismondi, Marcelo Reguero, Christian de Muizon, Alex Greenwood, Brian T. Chait, Kirsty Penkman, Matthew Collins und Ross D. E. MacPhee: Palaeoproteomics resolves sloth relationships. Nature Ecology & Evolution 3, 2019, S. 1121–1130, doi:10.1038/s41559-019-0909-z.
- Timothy J. Gaudin: Phylogenetic relationships among sloths (Mammalia, Xenarthra, Tardigrada): the craniodental evidence. Zoological Journal of the Linnean Society 140, 2004, S. 255–305.
- Diego Brandoni: A review of Pliomegatherium Kraglievich, 1930 (Xenarthra: Phyllophaga: Megatheriidae). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 4, 2006, S. 212–224.
- Diego Brandoni: Los Tardigrada (Mammalia, Xenarthra) del Mioceno Tardío de Entre Ríos, Argentinia. In: Diego Brandoni und J. I. Noriega (Hrsg.): El Neógeno de la Mesopotamia argentina. Asociación Paleontológica Argentina, Publicación Especial 14, 2013, S. 135–144.
- Alfredo A. Carlini, Diego Brandoni und Rodolfo Sánchez: Additions to the knowledge of Urumaquia robusta (Xenarthra, Phyllophaga, Megatheriidae) from the Urumaco Formation (Late Miocene), Estado Falcón, Venezuela. Paläontologische Zeitschrift 82 (2), 2008, S. 153–162.
- Nicolás R. Chimento, Federico L. Agnolin, Diego Brandoni, Daniel Boh, Mariano Magnussen, Francisco De Cianni und Federico Isla: A new record of Megatherium (Folivora, Megatheriidae) in the late Pliocene of the Pampean region (Argentina). Journal of South American Earth Sciences, 2020, S. 102950, doi:10.1016/j.jsames.2020.102950.
- Alfredo A. Carlini, Diego Brandoni und Rodolfo Sánchez: First Megatheriines (Xenarthra, Phyllophaga, Megatheriidae) from the Urumaco (Late Miocene) and Codore (Pliocene) Formations, Estado Falcón, Venezuela. Journal of Systematic Palaeontology 4 (3), 2006, S. 269–278.
- François Pujos: Paleogeographic distribution and anatomical adaptions in Peruvian megatheriine ground sloths (Xenarthra, Megatherioidea). In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 56–63.
- Paulina E. Nabel, Alberto Cione und Eduardo P. Tonni: Environmental changes in the Pampean area of Argentina at the Matuyama–Brunhes (C1r–C1n) Chrons boundary Palaeogeography, Palaeoclimatology, Palaeoecology 162, 2000, S. 403–412.
- Gustavo G. Politis, Clara Scabuzzo und Robert H. Tykot: An Approach to Pre-Hispanic Diets in the Pampas during the Early/Middle Holocene. International Journal of Osteoarchaeology 19, 2009, S. 266–280.
- Gustavo Martínez: „Fish-Tail“ projectile points and megamammals: new evidence from Paso Otero 5 (Argentina). Antiquity 75, 2001, S. 523–528.
- Gustavo Martínez, María Gutiérrez und José Luis Prado: New Archaeological Evidences from the Late-Pleistocene/Early-Holocene Paso Otero 5 Site (Pampean Region, Argentina). Current Research in the Pleistocene 21, 2004, S. 16–18.
- Gustavo Martínez: Arqueología del curso medio del Río Quequén Grande: Estado actual y aportes a la arqueología de la región Pampeana. Relaciones de la Sociedad Argentina de Antropología 31, 2006, S. 249–275.
- Gustavo Martínez, María A. Gutiérrez und Eduardo P. Tonni: Paleoenvironments and faunal extinctions: Analysis of the archaeological assemblages at the Paso Otero locality (Argentina) during the Late Pleistocene–Early Holocene. Quaternary International 299, 2013, S. 53–63.
- Marína A. Gutiérrez und Gustavo A. Martínez: Trends in the faunal human exploitation during the Late Pleistocene and Early Holocene in the Pampean region (Argentina). Quaternary International 191, 2008, S. 53–68.
- Anthony D. Barnosky und Emily L. Lindsey: Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. Quaternary International 217, 2010, S. 10–29.
- Gustavo G. Politis, Pablo G. Messineo, Thomas W. Stafford Jr. und Emily L. Lindsey: Campo Laborde: A Late Pleistocene giant ground sloth kill and butchering site in the Pampas. Science Advances 5, 2019; S. eaau4546, doi:10.1126/sciadv.aau4546.
- Karina V. Chichkoyan, Bienvenido Martínez-Navarro, Anne-Marie Moigne, Elisabetta Cioppi, Margarita Belinchón und José L. Lanata: Description and interpretation of a Megatherium americanum atlas with evidence of human intervention. Rivista Italiana di Paleontologia e Stratigrafia 123 (1), 2017, S. 51–64.
- Georges Cuvier: Notice sur le squelette d’une très grande espèce quadrupède inconnue jusqu’à présent, trouvé au Paraguay, et déposé au Cabinet d’Histoire naturelle de Madrid. Magasin encyclopédique 1, 1796, S. 303–310 ().
- Christine Argot: Changing Views in Paleontology: The Story of a Giant (Megatherium, Xenarthra). In: Eric J. Sargis und Marian Dagosto (Hrsg.): Mammalian Evolutionary Morphology. Vertebrate Paleobiology and Paleoanthropology Series 2008, S. 37–50.
- Don Joseph Garriga: Descripción del esqueleto de un quadrúpedo muy corpulento y raro, que se conserva en el Real Gabinete de Historia Natural de Madrid. Madrid, 1796 ().
- Georges Cuvier: Recherches sur les ossemens fossiles de quadrupèdes. Paris, 1812, Kapitel 8 und zugehörige Tafel 2 und 3 ().
- William Clift: Some account of the remains of the Megatherium sent to England from Buenos Ayres by Woodbine Parish. Transactions of the Geological Society 2 (3), 1835, S. 437–450 ().
- Paul D. Brinkman: Charles Darwin’s Beagle Voyage, Fossil Vertebrate Succession, and ‘‘The Gradual Birth & Death of Species’’. Journal of the History of Biology 43, 2010, S. 363–399.
- Richard Owen: Fossil Mammalia. In: Charles Darwin (Hrsg.): Zoology of the Voyage of H.M.S Beagle, under the command of Captain Fitzroy, during the years 1832 to 1836. Teil 1 von 4, London, 1840, S. 13–111 (S. 100–106) ().
- Richard Owen: On the Megatherium (Megatherium americanum Blumebach). Philosophical Transactions of the Royal Society of London: Part I. Preliminary Observations on the Exogenous Processes of Vertebrae. 141, 1851, S. 719–764 (), Part II. Vertebrae of the Trunk. 145, 1855, S. 359–388 (), Part III. The skull. 146, 1856, S. 571–589 (), Part IV. The bones of the anterior extremities. 148, 1858, S. 261–278 (), Part V. The bones of the posterior extremities. 149, 1859, S. 809–829 ().
- Richard Owen: Memoir on the Megatherium, or giant ground-sloth of America (Megatherium americanum, Cuvier). London, 1861, S. 1–84 ().
- Fariña, Vizcaíno und De Iuliis: Megafauna. 2013, S. 31–71.
- Georges Cuvier: Recherches sur les ossemens fossiles, où l'on rétablit les caractères de plusieurs animaux dont les révolutions du globe ont détruit les espèces. Tome Cinquième. Paris, 1823, S. 1–405 (S. 174–192 ).
- Juan-Carlos Fernicola, Serdio F. Vizcaíno und Richard a. Fariña: The evolution of armored xenarthrans and the phylogeny of the glyptodonts. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 79–85.
- Joseph Eduard d’Alton: Über die von dem verstorbenen Herrn Sellow aus der Banda oriental mitgebrachten fossilen Panzerfragmente und die dazugehörigen Knochen-Überreste. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin, 1833, S. 369–424 ().
- Peter Wilhelm Lund: Blik paa Brasiliens Dyreverden för Sidste Jordomvaeltning. Anden Afhandling: Patteedyrene. Det Kongelige Danske Videnskabernes Selskbas Naturvidenskabelige og Matematiske Afhandlinger 8, 1839, S. 61–144 (S. 83) ().
- Richard Owen: Note on the Glyptodon. In: Woodbine Parish (Hrsg.): Buenos Ayres, and the provinces of the Rio de La Plata: their present state, trade and dept; with some account from original documents of the progress of geographical discovery in those parts of South America during the last sixty years. London, 1839, S. 178b–178e ().
- Richard Owen: Description of a tooth and part of the skeleton of the Glyptodon clavipes, a large quadrupede of the Edentate order, to which belongs the tesselated bony armour described and figured by Mr. Clift in the former Volume of the Transactions of the Geological Society; with a consideration of the question whether the Megatherium possessed an analogous dermal armor. Transactions of the Geological Society 2 (6), 1841, S. 81–106 ().
- Thomas Henry Huxley: On the Osteology of the Genus Glyptodon. Philosophical Transactions of the Royal Society of London 155, 1865, S. 31–70 ().
- Robert V. Hill: Comparative Anatomy and Histology of Xenarthran Osteoderms. Journal of Morphology 267, 2005, S. 1441–1460.