Kürbisgewächse
Die Kürbisgewächse (Cucurbitaceae) sind eine Familie der Bedecktsamigen Pflanzen (Magnoliopsida). Die Vertreter sind meist krautige Pflanzen mit Ranken, eingeschlechtigen Blüten und unterständigem Fruchtknoten. Die Blütenkrone ist meist verwachsen, die Staubblätter sind untereinander verbunden bis verwachsen.
| Kürbisgewächse | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
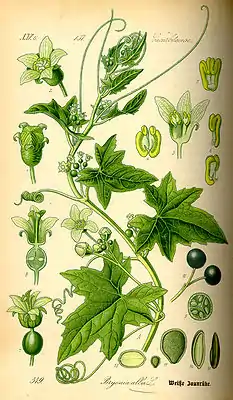
Weiße Zaunrübe (Bryonia alba), Illustration | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Cucurbitaceae | ||||||||||||
| Juss. |
Die Kürbisgewächse sind eine mittelgroße Familie und umfassen rund 800 Arten in rund 130 Gattungen, die in die beiden Unterfamilien Nhandiroboideae und Cucurbitoideae aufgeteilt werden. Sie sind weltweit in den tropischen und subtropischen Klimazonen vertreten, nur wenige Arten kommen auch in gemäßigten Gebieten vor; in Mitteleuropa etwa ist nur die Gattung der Zaunrüben (Bryonia) heimisch. Zur Familie zählen etliche Nutzpflanzen, die kommerziell bedeutendsten sind Gartenkürbis (Cucurbita pepo), Zuckermelone (Cucumis melo), Gurke (Cucumis sativus) und Wassermelone (Citrullus lanatus).
Beschreibung
Wurzeln und Speicherorgane
Die meisten Vertreter haben eine ausgeprägte Pfahlwurzel, die etwa bei Gurke (Cucumis sativus) einen und bei Kürbissen (Cucurbita) zwei Meter in die Tiefe reicht. Nahe der Oberfläche bilden sie ein dichtes Netz aus Seitenwurzeln, die mindestens eine so große, wenn nicht größere Bodenfläche bedecken als die oberirdischen Organe. An liegenden Sprossachsen bilden sich häufig Adventivwurzeln, auch ohne direkten Kontakt der Sprossachse mit dem Boden.
Einige xerophytische Vertreter bilden Speicherwurzeln, mit denen sie Trockenzeiten überstehen. Bei Acanthosicyos reichen sie bis 15 Meter tief.
Sprossachse und Ranken
Die Sprossachsen sind krautig, selten auch leicht verholzt. Meist sind sie niederliegend, kriechend oder kletternd. Im Querschnitt sind sie häufig eckig und besitzen ein hohles Zentrum. Die Seitenzweige erster und zweiter Ordnung können bis 15 Meter lang werden. Bei Kürbissen gibt es strauchförmige Formen mit kurzen Internodien.

Die Leitbündel sind bikollateral, das heißt, das Phloem befindet sich innen und außen am Xylem. Meist gibt es zehn Leitbündel, die in zwei Ringen um die zentrale Höhle angeordnet sind. Die Siebröhren sind relativ groß und sind bei manchen Sippen auch in der Rinde verstreut und verbinden so die einzelnen Leitbündel.
Viele Vertreter sind sowohl an der Sprossachse als auch an den Blättern weich oder rau behaart, nur wenige sind unbehaart. Die Form der Trichome ist sehr variabel: sie können drüsig sein, ein- oder vielzellig, einfach oder verzweigt.
In etlichen nicht näher verwandten Sippen innerhalb der Familie tritt eine Ausbildung dicker Stämme als Speicherorgane (Pachycaulie) auf. Pachycaule Vertreter in der Unterfamilie Nhandiroboideae sind Gerrardanthus, Neoalsomitra, Xerosicyos und Zygosicyos. Innerhalb der Unterfamilie Cucurbitoideae in der Tribus Benincaseae Cephalopentandra, Coccinia, Trochomeria, in der Tribus Coniandreae Corallocarpus, Doyerea, Ibervillea, Kedrostis, Seyrigia, sowie verstreut Marah, Sinobaijiana, Momordica, Odosicyos und Tricyclandra. Der extremste Vertreter ist Dendrosicyos.[1]




Die Ranken sind modifizierte Sprossachsen und das deutlichste abgeleitete Merkmal der Familie. Sie kommen bei allen außer einigen stark abgewandelten, holzigen Arten vor. Bei einigen dieser Arten (Acanthosicyos horridus, Acanthosicyos naudiana) sind sie zu Dornen umgewandelt, bei anderen (Citrullus ecirrhosus, Dendrosicyos socotranus, Ecballium elaterium, Melancium campestre, Myrmecosicyos messorius und Trochomeria polymorpha) fehlen sie ganz. Meist wird zwischen drei Rankentypen unterschieden:
- einfache, unverzweigte Ranken hat beispielsweise die Gurke (Cucumis sativus).
- verzweigte Ranken, deren unterer Teil berührungsunempfindlich ist und sich nicht aufrollt. Die Ranken können bis zu acht Seitenranken tragen. Dieser Typ ist hauptsächlich bei den Cucurbitoideae vertreten. Einige Arten haben scheibenförmige Haftorgane ähnlich denen der Jungfernreben (Parthenocissus) entwickelt, etwa Alsomitra macrocarpa, Neoalsomitra sarcophylla oder Trichosanthes cucumerina.
- verzweigte Ranken, deren unterer Teil sich einrollt, auch zaninoide Ranken genannt. Dieser Typ tritt vor allem in der Tribus Zanonieae (= Unterfamilie Nhandiroboideae) und einigen basalen Vertretern der Unterfamilie Cucurbitoideae auf.[2]
Bei einigen Arten sind sogenannte Probrakteen (Probracts) ausgebildet. Dies sind oft abfallende oder beständige tragblatt- oder blattartige und oft drüsige Strukturen beim Blütenstands- oder Blütenstiel oder seltener an sterilen Knoten oder Blattknoten.
Blätter
Die Laubblätter sind wechselständig an der Sprossachse angeordnet in einer Schraube mit 2/5-Phyllotaxis. Laubblätter sind in Blattstiel und Blattspreite gegliedert. Im Blattstiel sind die Leitbündel halbmond- oder ringförmig angeordnet. Die Leitbündel sind ungleich, die größeren sind bikollateral. Die Blattspreiten sind einfach und leicht bis tief drei- bis siebenfach handförmig gelappt. Auch die Aderung ist handförmig. Sukkulente Blätter sind selten, auch bei den xerophytischen Arten. Xerosicyos besitzt große wasserspeichernde Zellen im Mesophyll und ist eine CAM-Pflanze. Bei den meisten Vertretern in trocken-heißen Gebieten wird eine Hitzeschädigung der Blätter durch hohe Transpirationsraten verhindert. Bei einigen Arten finden sich auf den Blättern extraflorale Nektarien. Nebenblätter fehlen in der Regel. Bei Acanthosicyos sind Nebenblätter vorhanden und in photosynthetisch aktive Dornen umgewandelt.
Die Spaltöffnungen sind vorwiegend anomocytisch, ihnen fehlen die Begleitzellen.
Blüten

Die Blüten der meisten Vertreter sind groß und auffällig und werden von Insekten bestäubt. Einige Gattungen wie Echinocystis und Sechium bilden kleine, unauffällige Blüten. Die Blüten aller Vertreter stehen in den Blattachseln, entweder einzeln oder in Blütenständen. Häufige Farben sind weiß und gelb, seltener sind andere Farben wie rot bei Gurania.
Die Blüten sind meist eingeschlechtig, besitzen also entweder männliche oder weibliche Organe. Die Blütenanlagen sind jeweils zwittrig angelegt, es entwickeln sich jedoch nur die männlichen beziehungsweise die weiblichen Organanlagen weiter. In der männlichen (staminaten) Blüte ist oft ein Stempel-Rudiment (Pistillodium) erhalten, in der weiblichen können Staubblatt-Rudimente (Staminodien) vorkommen. Die Pflanzen sind dabei je nach Art oder Gattung einhäusig (beide Geschlechter an einer Pflanze) oder zweihäusig (nur je ein Geschlecht an einer Pflanze). Von den etwa 800 Arten sind rund 460 einhäusig, 340 zweihäusig. Auf weitere, seltene Geschlechtsverteilungen wird weiter unten eingegangen.
Die Blüten besitzen einen ausgeprägten „falschen“ Blütenbecher (Hypanthium), der becher- bis glockenförmig ist und aus der Blütenachse und/oder aus den verwachsenen Basalteilen von Kelch und Krone (Kelch-Kronbecher) gebildet wird. Die Blüten sind meist fünfzählig. Die beiden Kreise der doppelten Blütenhülle bestehen meist aus je fünf, selten zwischen drei und sechs, Blütenblättern. Die Kelchblätter sind mehr oder weniger verwachsen. Die Kronblätter sind frei oder verwachsen, meist radiärsymmetrisch und bilden eine rad- oder glockenförmige Krone.
Die Staubblätter sind fast immer monothezisch (besitzen nur eine Theka), außer bei Fevillea, setzen am Hypanthium an und stehen alternierend zu den Kronblättern. Die Grundzahl der Staubblätter ist fünf. Manche Gattungen wie Fevillea haben fünf freie Staubblätter, bei manchen sind alle Staubblätter verwachsen wie bei Cyclanthera. Andere Gattungen wie Gurken (Cucumis) haben zwei verwachsene Paare und ein freies Staubblatt, sodass der Eindruck von drei Staubblättern entsteht. Diese drei sind meist über ihre Antheren miteinander verbunden.
Der Fruchtknoten ist meist unterständig oder (epiperigyn), das Perianth hat eine kurze bis lange „epigyne Zone“, die den Kelch trägt. Er besteht aus meist drei verwachsenen Fruchtblättern, seltener ein bis fünf. Die Zahl der Griffel ist stark konserviert: bei den Nhandiroboideae sind sie stets frei und meist drei an der Zahl, selten zwei. Die Cucurbitoideae besitzen einen Griffel mit zwei, drei oder fünf Narben. Diese sind meist vergrößert und ähneln oft den Staubblättern, wohl um Bestäuber anzulocken.[1] Der Fruchtknoten ist nicht in Fächer unterteilt, die Scheidewände sind aufgelöst und die Plazenten, an denen die Samenanlagen sitzen, befinden sich an der Fruchtwand (parietal).[3] Bei den Nhandiroboideae ist die Zahl der Samenanlagen meist gering, die Samenanlagen sind stets hängend. Die Cucurbitoideae besitzen meist große Plazenten mit manchmal sehr vielen Samenanlagen, Sechium edule jedoch nur eine. Diese stehen meist aufrecht oder waagrecht, selten hängend. Die Samenanlagen in der ganzen Familie sind crassinucellat, anatrop und bitegmisch.[4] Der Nucellus ist flaschenförmig, was ein charakteristisches Familienmerkmal darstellt. Der Embryosack entwickelt sich nach dem Polygonum-Typ.[4]
Die Blüten besitzen häufig, aber nicht immer Nektarien, durch die die bestäubenden Insekten angelockt werden. Sie befinden sich an der Basis des Hypanthiums. In weiblichen Blüten bildet das Nektarium einen durchgehenden Ring um die Basis des Stempels, während es bei den männlichen Blüten oft zusammen mit dem manchmal ausgebildeten Pistillodium einen knopfartigen Hügel im Zentrum der Blüte bildet.
Pollen
Die Pollenkörner sind innerhalb der Familie recht vielgestaltig. Die Apertur kann tri- bis mehrfach zonocolpat, colporat, tri- bis mehrfach porat oder pantoporat sein. Die Oberfläche der Exine ist netzförmig, regelmäßig granuliert oder mit Stacheln versehen.[5]
Der Pollen wird in Form von Einzelkörnern, seltener in Pollentetraden ausgebreitet. Die Nhandiroboideae besitzen stets tricolpate Pollenkörner mit einem Durchmesser von unter 40 Mikrometern. Ihre äußere Schicht (Exine) ist meist gestreift (striat), selten netzförmig (reticulat). Die Cucurbitoideae besitzen mit bis zu 200 Mikrometern ungewöhnlich große Pollenkörnern, die Exine ist netzförmig oder stachelig (echinat), die Aperturen sind porat oder colpat. Zwei Triben können anhand der Pollenkörner identifiziert werden: die Cucurbiteae besitzen echinate Pollenkörner, die drei bis viele Keimporen besitzen (triporat bis stephanoporat sind); die Sicyeae besitzen fein stachelige Pollenkörner mit Keimfalten (colpat) oder gemischten Poren/Falten (colporat). Bei den übrigen Triben überwiegen tricolporate Pollenkörner.[1]
Früchte und Samen

Die Früchte sind vielfach nicht öffnende Beeren, häufig mit einer harten Schale, sogenannte Panzerbeeren, seltener werden öffnende Beeren oder Kapselfrüchte gebildet, sehr selten sind Steinfrüchte oder Flügelnüsse. Gestalt und Oberfläche sind dabei sehr variabel. Sie sind kugelig bis länglich. Die Flaschenkürbisse (Lagenaria) sind rund, birnen-, flaschenförmig oder länglich. Bei Kürbissen kann die Oberfläche gestreift, gefleckt oder einfarbig sein, die Oberfläche glatt, gerunzelt oder warzig. Die Früchte des Riesen-Kürbis (Cucurbita maxima) sind die größten des Pflanzenreichs: am 29. September 2007 wurde in Massachusetts ein 766 kg schwerer Kürbis gewogen.[6] Die meisten Arten mit Panzerbeeren sind in saisonal trockenen Gebieten heimisch: hier ermöglichen die hartschaligen, wasserspeichernden Früchte den Samen eine längere Reifezeit, auch wenn der Rest der Mutterpflanze bereits verwelkt ist. Die meisten bleiben bei der Reife geschlossen. Bei den Gattungen Momordica und Cyclanthera öffnen sich die Früchte. Die Früchte der Zaunrüben (Bryonia) sind rund fünf Millimeter groß, kugelig und grün, rot oder schwarz.
Bei der Spritzgurke (Ecballium elaterium) und bei der Explodiergurke (Cyclanthera brachystachya) lösen sich die Früchte vom Stiel und stoßen das Fruchtinnere, das unter Druck steht, explosionsartig aus. Cucumis humifructus verbirgt die Blüten nach der Befruchtung in der Erde (Geokarpie), wo die Früchte reifen und vom Erdferkel (Orycteropus afer) ausgegraben und verzehrt werden, das so die Samen ausbreitet.[7] Auch die amerikanische Gattung Apatzingania hat geokarpe Früchte.
Bei den Nhandirobideae kommen vorwiegend Kapselfrüchte vor, die geflügelte Samen entlassen.

Die Samen sind meist zu vielen, oft zu hunderten in einer Frucht enthalten, sehr selten ist nur ein einzelner pro Frucht. Die Samen sind meist flach, in seltenen Fällen geflügelt. Unter der Samenschale befindet sich ein kollabiertes Perisperm. Ein Endosperm fehlt oder ist spärlich vorhanden. Der Embryo ist sehr ölreich, seine beiden Keimblätter füllen den Großteil des Samens aus.
Größe, Form und Farbe sind sehr vielfältig. Den größten ungeflügelten „Steinkern“ bildet Hodgsonia mit bis 8,5 Zentimeter Länge, den kleinsten die Zaunrüben (Bryonia) mit manchmal nur 3 Millimeter Durchmesser.
Cytologie und Genetik
Die vorherrschende Chromosomengrundzahl beträgt x = 12, sie reicht jedoch von 7 bis 24. Die Cucurbiteae haben eine fixierte Polyploidie mit einem Chromosomensatz von n = 20. Auch in anderen Triben kommen recht häufig Polyploidie und Aneuploidie vor. Die gesamte Familie ist durch eine 35 Basenpaare lange Inversion im Bereich zwischen den Genen für Leucin-tRNA und Phenylalanin-tRNA gekennzeichnet, die bei den übrigen Familien der Ordnung Cucurbitales nicht vorkommt, also eine Synapomorphie darstellt. Bei Cucurbita digitata und bei Neoalsomitra ist diese Inversion wieder – unabhängig voneinander – umgekehrt worden.
Das Genom der Cucurbitaceae ist im Vergleich zu anderen Familien ungewöhnlich labil. So kommen hier zwei der bekannten sechs Fälle vor, bei denen das ansonsten plastidäre Gen rbcL (für die große Untereinheit der RuBisCO) in das Mitochondrien-Genom transferiert ist: bei der Gurke (Cucumis sativus) und bei den Kürbissen (Cucurbita pepo, Cucurbita maxima), dies jedoch unabhängig voneinander.
Die Gattung Cucumis verfügt auch über die größten bekannten Mitochondrien-Genome, die 1500 kb bei der Gurke und 2400 kb bei der Zuckermelone (Cucumis melo) groß sind. Bei beiden Arten erfolgt die mitochondriale Vererbung über den Vater (paternal), nicht wie sonst üblich über die Mutter.[2]
Inhaltsstoffe
Die für die Familie charakteristischsten Inhaltsstoffe sind die Bitterstoffe aus der Gruppe der Cucurbitacine. Dies sind tetrazyklische Triterpene und kommen glykosyliert oder als Aglykon vor. Sie sind vor allem in den Früchten vorhanden, mit Ausnahme von Zuchtformen, die auf geringe Bitterkeit selektiert wurden. Samen sind stets frei von Cucurbitacin.[8]
Neben den Cucurbitacinen kommen ebenfalls bittere Triterpen-Saponine vor, die auf den Grundgerüsten Dammaran, Cucurbitan und Lanostan beruhen.[8] Besonders reich an Saponinen sind die Gattungen Luffa und Trichosanthes.
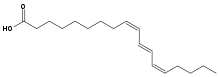
Die Samen sind generell reich an Ölen. Bei den kommerziell genutzten Arten variiert der Ölgehalt zwischen 38 und 45 Prozent. Die häufigsten Fettsäuren sind Palmitinsäure, Stearinsäure, Ölsäure, Linolsäure und Linolensäure, die zusammen mehr als 80 Prozent der Fettsäuren ausmachen. Daneben sind Phosphatidylcholin, Lysophosphatidyl-Ethanolamin, Phosphatidyl-Inositol und Phosphatidylethanolamin, Sterol-Glykoside, Cerebrosid und Diphosphatidyl-Glycerol häufig. In Trichosanthes-Arten ist Punicasäure, eine konjugierte Triensäure, häufig, in Momordica α-Elaeostearinsäure. In Summe wurden in Samen der Kürbisgewächse über 150 Fettsäuren identifiziert.[4]
In Samen und Früchten kommen verschiedentlich nicht-proteinogene Aminosäuren vor, besonders Citrullin.[8] Die Speicherproteine in den Samen sind vorwiegend Globuline.[4]
Im Phloem transportieren die Kürbisgewächse als Kohlenhydrate außer Saccharose auch die Vertreter der Raffinose-Familie, sie werden deshalb zu den symplastischen Phloem-Beladern gerechnet.

Wachstum und Entwicklung
Samen können bei einigen Arten, wie bei Chayote (Sechium edule), bereits in der Frucht auskeimen (Viviparie). Bei vielen Arten gibt es eine Samen-Dormanz, die jedoch bereits durch einige Wochen bei kühlen Temperaturen gebrochen wird. Für die Keimung sind Dunkelheit und hohe Temperaturen (meist 25 bis 30 °C) nötig. Die Keimung erfolgt epigäisch, das heißt, dass die Keimblätter photosynthetisch aktiv sind. Sie sind meist querelliptisch.
Kürbisgewächse wachsen bei hohen Temperaturen sehr rasch. Ein wildes Exemplar von Cucurbita foetidissima bildete in einer Saison 360 Sprosse in einem Kreis von 12 Meter Radius und einer Gesamtlänge von 2000 Meter. Der Flaschenkürbis wächst bis zu 60 Zentimeter in 24 Stunden. Bei den großfrüchtigen, rankenden Pflanzen wächst die Blattfläche exponentiell bis zum Zeitpunkt des Fruchtansatzes.
Geschlechtsbestimmung
Die Geschlechtigkeit der Kürbisgewächse ist äußerst vielfältig und variiert teilweise auch innerhalb einer Art. Die ursprüngliche Geschlechtsverteilung innerhalb der Familie ist die Zweihäusigkeit (Diözie). Monözie und andere Formen sind davon abgeleitet. Androdiözie (männliche Pflanzen und zwittrige Pflanzen) kommt bei Pflanzen äußerst selten vor, etwa bei Schizopepon bryoniifolius. Bei den Kürbisgewächsen äußerst selten sind zwittrige Blüten, die nur von wenigen Varietäten von Nutzpflanzen bekannt sind, etwa 'Satputia' des Schwammkürbis Luffa acutangula.
Bei den wenigen untersuchten Arten wird die Geschlechtsverteilung durch ein oder zwei Gene gesteuert, von denen es jeweils zwei oder drei Allele gibt. Je nach Allel-Kombination gibt es, etwa bei Luffa acutangula monözische, andromonözische (männliche und zwittrige Blüten), andrözische (nur männliche), gynomonözische (weibliche und zwittrige), gynözische (nur weibliche Blüten) und zwittrige Individuen. Bei manchen Arten ändern Individuen ihr Geschlecht im Laufe ihrer Entwicklung, etwa in Gurania.
In den Gattungen Coccinia und Trichosanthes gibt es – zum Teil heteromorphe – Geschlechtschromosomen, die das Geschlecht der Einzelpflanzen bestimmen. Bei Coccinia grandis sind, ähnlich wie bei Menschen, Individuen mit zwei X-Chromosomen weiblich. Das größere Y-Chromosom bestimmt das männliche Geschlecht und dominiert, sodass auch tetraploide Individuen mit drei X- und einem Y-Chromosom männlich sind.
Die Ausbildung des Geschlechtes unterliegt nicht nur genetischer Regulation, sondern wird auch durch den Hormonhaushalt, durch Tageslänge und Temperatur beeinflusst. Bei hohen Temperaturen werden männliche Blüten gefördert, bei tiefen weibliche. Kurztagsbedingungen (acht Stunden Licht) fördern vielfach weibliche Blüten. Generell werden weibliche Blüten unter Bedingungen gefördert, bei denen Kohlenhydrate akkumuliert werden.
Diese hohe Variabilität wird in der Züchtung ausgenützt. So sind die meisten in Europa angebauten Gurken-Sorten gynözisch, bilden also nur weibliche Blüten, die auch ohne Bestäubung Früchte ansetzen (Parthenokarpie). Zur Samenzucht wird mit Hilfe von Silber-Ionen (einem Ethylen-Inhibitor) die Bildung von männlichen Blüten angeregt.
Bestäubung
Die Blüten öffnen sich meist am frühen Morgen, die Bestäubung ist meist bis Mittag erfolgt. Manche weißblütige Arten, wie der Flaschenkürbis, öffnen sich am Abend und bleiben bis zum Morgen geöffnet. Diese Arten werden meist von Schwärmern bestäubt. Die tagsüber blühenden Arten werden überwiegend von Bienen und anderen Insekten bestäubt, die Pollen und Nektar sammeln. Darunter gibt es auch einige Spezialisten, so besuchen die Bienen-Gattungen Peponapis und Xenoglossa vorwiegend Blüten der Gattung Cucurbita.
Samen- und Fruchtbildung

Das Endosperm entwickelt sich nukleär (durch freie Kernteilung ohne Zellbildung). Das junge Endosperm bildet ein Haustorium in Richtung Chalaza. Dieses bleibt coenocytisch (keine Zellbildung, also freie Zellkerne) oder wird später zellulär. Die Zellen des Endosperms sind im späteren, zelligen Stadium zum Teil hochpolyploid, bei Echinocystis lobata wurde 3072n gezählt. Im reifen Samen ist das Endosperm stark reduziert und weitgehend degeneriert. Häufig umfasst es nur zwei Zellschichten.[4]
Bei einigen Arten kommt gelegentlich Polyembryonie vor, es werden in einem Samen mehrere Embryonen gebildet. Bei Hodgsonia macrocarpa (im „Steinkern“) kommt sie stets vor.[4]
Die Samenschale wird bei den Kürbisgewächsen nur vom äußeren Integument der Samenanlage gebildet, das innere degeneriert. Sie besteht von außen nach innen aus einer häufig dreischichtigen Epidermis, einer Hypodermis, einem Sklerenchym, einem Aerenchym und einem Chlorenchym. Dieser fünfschichtige Aufbau ist für die Familie charakteristisch, teilweise gibt es Abwandlungen davon.[4]
Für die Entwicklung der Früchte ist in der Regel eine erfolgreiche Befruchtung nötig. Am Ende der Wachstumsperiode können sich bei etlichen Arten jedoch die letzten Blüten ohne Befruchtung entwickeln, wenn bis dahin noch keine Blüte der Pflanze befruchtet wurde. Auf der anderen Seite wird durch sich entwickelnde Früchte vielfach die Bildung weiterer weiblicher Blüten unterdrückt. Die Form der Früchte, die bei den Kürbisgewächsen sehr variabel ist, ist häufig bereits in der Form des Fruchtknotens vorgebildet.
Systematik








_at_Pogallapalli_Khammam.jpg.webp)


.JPG.webp)

_flor_M02_antesis_vista_superior_poros_de_acceso_al_n%C3%A9ctar.JPG.webp)
Die Familie der Cucurbitaceae wurde 1789 durch Antoine Laurent de Jussieu in Genera Plantarum, Seiten 393–394 aufgestellt. Typusgattung ist Cucurbita L. Synonyme für Cucurbitaceae Juss. nom. cons. sind: Bryoniaceae G.Mey., Nhandirobaceae T.Lestib. nom. illeg., Zanoniaceae Dumort., Cyclantheraceae Lilja.
Die nächstverwandten Familien der Kürbisgewächse (Cucurbitaceae) innerhalb der Ordnung Cucurbitales sind die Begoniengewächse (Begoniaceae), Scheinhanfgewächse (Datiscaceae) und Tetramelaceae.[9] Die traditionelle Systematik der Kürbisgewächse beruht vorwiegend auf Merkmalen des Androeceums, des Gynoeceums, der Ranken, seit Mitte des 20. Jahrhunderts auch auf Pollenstruktur und dem Bau der Samenschale. Eine der neuesten Klassifikationen der Familie ist die von C. Jeffrey (2005).[10] Folgende Aufstellung ist aus Kocyan u. a. (2007) übernommen, die das System von Jeffrey durch neue Gattungen erweitert hatten, so dass die Familie 126 Gattungen umfasst. Änderungen gegenüber Kocyan u. a. sind gekennzeichnet:
- Unterfamilie Nhandiroboideae (Syn.: Zanonioideae)
- Tribus Zanonieae
- Subtribus Zanoniinae
- Alsomitra (Blume) M.Roem.: Sie enthält nur eine Art:
- Alsomitra macrocarpa (Blume) M.Roem.: Sie ist im tropischen Asien in Thailand, Malaysia, Indonesien, Neuguinea sowie auf den Philippinen verbreitet.
- Bayabusua W.J.de Wilde (3 Arten auf der malaiischen Halbinsel)
- Gerrardanthus Harv. ex Benth. & Hook.f. (4 Arten im tropischen und südlichen Afrika)
- Neoalsomitra Hutch. (12 Arten in Indien, der Volksrepublik China, Polynesien und Australien)
- Siolmatra Baill.: Die nur zwei Arten kommen im Amazonasbecken vor.
- Xerosicyos Humbert (2 bis 3 Arten auf Madagaskar)
- Zanonia L.: Sie enthält nur eine Art:
- Zanonia indica L.: Sie ist in Indien, Indochina, China, Malaysia und auf den Philippinen verbreitet.
- Zygosicyos Humbert: Die nur zwei Arten kommen auf Madagaskar vor.
- Alsomitra (Blume) M.Roem.: Sie enthält nur eine Art:
- Subtribus Fevilleinae
- Fevillea L. (7 Arten in Zentral- und im tropischen Südamerika)
- Fevillea cordifolia L.: Karibik, Zentralamerika bis südliches Mexiko und nördliches bis nordwestliches Südamerika
- Fevillea L. (7 Arten in Zentral- und im tropischen Südamerika)
- Subtribus Gomphogyninae
- Gomphogyne Griff. (1–2 Arten in China, Indochina, Malesien)
- Gynostemma Blume (5–13 Arten von Indien bis Japan, Malaysia)
- Gynostemma pentaphyllum (Thunb.) Makino
- Hemsleya Cogn. ex F.B.Forbes & Hemsl.: 3 bis 24 Arten in China (Himalaya)
- Subtribus Actinostemmatinae
- Actinostemma Griff. (1–3 Arten in Asien)
- Bolbostemma Franquet (2 Arten in China)
- Subtribus Sicydiinae
- Chalema Dieterle: Sie enthält nur eine Art:
- Chalema synanthera Dieterle: Sie kommt in Mexiko vor.
- Cyclantheropsis Harms (2–3 Arten in Ostafrika und Madagaskar)
- Pseudosicydium Harms: Sie enthält nur eine Art:
- Pteropepon (Cogn.) Cogn. (4 Arten in Südamerika)
- Sicydium Schltdl. (6 Arten in Mexiko, Zentral- und im tropischen Südamerika)
- Chalema Dieterle: Sie enthält nur eine Art:
- Subtribus Zanoniinae
- Tribus Zanonieae
- Unterfamilie Cucurbitoideae
- Tribus Joliffieae
- Subtribus Telfairiinae
- Odosicyos Keraudren: Sie enthält nur eine Art:
- Odosicyos bosseri Keraudren: Sie kommt auf Madagaskar vor.
- Telfairia Hook. (3 Arten im tropischen Afrika)
- Odosicyos Keraudren: Sie enthält nur eine Art:
- Subtribus Thladianthinae
- Baijiania A.M.Lu & J.Q.Li: Sie enthält nur eine Art:
- Baijiania borneensis (Merr.) A.M.Lu & J.Q.Li: Sie kommt auf Borneo vor.
- Indofevillea Chatterjee: Sie enthält nur eine Art:
- Microlagenaria (C. Jeffrey) A.M.Lu & J.Q.Li: Sie enthält nur eine Art:
- Bittermelonen (Momordica L.) (47 Arten in Afrika, tropisches Asien, Australien und als Neophyt in der Neuen Welt), darunter:
- Momordica balsamina L. (Balsamapfel, Jerusalemsapfel)[11][12]
- Bittermelone (Momordica charantia L.)
- Momordica cochinchinensis (Lour.) Spreng.
- Sinobaijiania C. Jeffrey & W.J.de Wilde (4 Arten in China, Indonesien, Taiwan, Thailand)
- Siraitia Merr. (4 Arten in Indien, China, Thailand, Vietnam, Indonesien). Darunter:
- Siraitia grosvenorii (Swingle) A. M. Lu & Zhi Y. Zhang
- Thladiantha Bunge (25 Arten in Asien)
- Thladiantha dubia Bunge: Zentrales China bis in die Mandschurei und südliche Mongolei bis nach Korea, Sachalin sowie in ins angrenzende Ostrussland.
- Baijiania A.M.Lu & J.Q.Li: Sie enthält nur eine Art:
- Subtribus Telfairiinae
- Tribus Bryonieae
- Austrobryonia H.Schaef. (4 Arten in Australien; 2008 erstbeschrieben[13])
- Zaunrüben (Bryonia L.) (10 Arten im Mittelmeerraum, Nordafrika und Zentralasien)
- Ecballium A.Rich.: Sie enthält nur eine Art:
- Spritzgurke (Ecballium elaterium (L.) A.Rich.): Sie ist im Mittelmeerraum, Nordafrika und Zentralasien verbreitet.
- Tribus Trichosantheae
- Subtribus Ampelosicyinae
- Ampelosicyos Thouars (3 Arten auf Madagaskar)
- Tricyclandra Keraudren: Sie enthält nur eine Art:
- Tricyclandra leandrii Keraudren: Sie kommt auf Madagaskar vor.
- Subtribus Hodgsoniinae
- Hodgsonia Hook.f. & Thomson (2 Arten von Nordost-Indien bis Borneo)
- Hodgsonia macrocarpa (Blume) Cogn.: Östlicher Himalaya bis nach Südostasien.
- Hodgsonia Hook.f. & Thomson (2 Arten von Nordost-Indien bis Borneo)
- Subtribus Trichosanthinae
- Gymnopetalum Arn. (4 Arten in China, Indochina, Indien)
- Trichosanthes L. (100 Arten in China, Indien, Malesien, Australien), darunter:
- Schlangenhaargurke (Trichosanthes cucumerina L.)
- Subtribus Ampelosicyinae
- Tribus Herpetospermeae
- Biswarea Cogn.: Sie enthält nur eine Art:
- Biswarea tonglensis (C.B.Clarke) Cogn.: Sie kommt in Indien, Myanmar sowie China vor.
- Edgaria C.B. Clarke: Sie enthält nur eine Art:
- Edgaria darjeelingensis C.B.Clarke: Sie ist in Inden, Nepal sowie China verbreitet.
- Herpetospermum Wall. ex Benth. & Hook.f.: Sie enthält nur eine Art:
- Herpetospermum pedunculosum (Ser.) C.B.Clarke: Sie kommt in Nepal, Tibet sowie China vor.
- Biswarea Cogn.: Sie enthält nur eine Art:
- Tribus Schizopeponeae
- Schizopepon Maxim. (8 Arten in Russland, Indien, Myanmar, China, Japan)
- Tribus Luffeae
- Luffa Mill. (7 Arten in Afrika, Arabien, Asien, Australien, Zentral- und Südamerika), darunter:
- Schwammkürbis (Luffa cylindrica (L.) M.Roem.)
- Luffa operculata (L.) Cogn.
- Luffa Mill. (7 Arten in Afrika, Arabien, Asien, Australien, Zentral- und Südamerika), darunter:
- Tribus Sicyeae
- Subtribus Cyclantherinae
- Brandegea Cogn.: Sie enthält nur eine Art:
- Brandegea bigelovii (S.Watson) Cogn.: Dieser Endemit kommt nur in Niederkalifornien vor.
- Cyclanthera Schrad. (inkl. Cremastopus Paul G.Wilson): 25 Arten von Nordamerika bis Argentinien
- Echinocystis Torr. & A.Gray: Sie enthält nur eine Art:
- Stachelgurke (Echinocystis lobata (Michx.) Torr. & A.Gray): Sie ist in Nordamerika verbreitet und ist in Europa ein Neophyt.
- Echinopepon Naudin (inkl. Apatzingania Dieterle) (18 Arten von Arizona bis Argentinien)
- Elateriopsis Ernst (5 Arten in Zentral- bis tropisches Südamerika)
- Hanburia Seem. (2 Arten in Mexico und Guatemala)
- Marah Kellogg (7 Arten in Nordamerika)
- Pseudocyclanthera Mart. Crov.: Sie enthält nur eine Art:
- Rytidostylis Hook. & Arn. (5 Arten in Zentral- bis tropisches Südamerika)
- Vaseyanthus Cogn.: Sie enthält nur eine Art:
- Vaseyanthus brandegeei (Cogn.) Rose: Sie kommt in Mexiko vor.
- Brandegea Cogn.: Sie enthält nur eine Art:
- Subtribus Sicyinae
- Microsechium Naudin (2 Arten in Mexiko und Guatemala)
- Parasicyos Dieterle (2 Arten in Mexiko und Guatemala)
- Sechiopsis Naudin (inkl. Pterosicyos Brandegee) (5 Arten in Mexiko und Guatemala)
- Sechium P.Browne (11 Arten in Zentralamerika bis tropisches Südamerika)
- Sicyos L. (50 Arten von Nordamerika bis Argentinien, Hawaii)
- Haargurke (Sicyos angulatus L.): Aus dem mittleren bis östlichen Nordamerika
- Sicyosperma A.Gray: Sie enthält nur eine Art:
- Sicyosperma gracile A.Gray: Sie kommt nur in Arizona vor.
- Subtribus Cyclantherinae
- Tribus Coniandreae
- Apodanthera Arn. (15 Arten in Nord- und Südamerika)
- Ceratosanthes Burm. ex Adans. (4 Arten in Zentral- bis tropisches Südamerika)
- CorallocarpusWelw. ex Benth. & Hook.f. (17 Arten in Afrika, Madagaskar, Indien)
- Cucurbitella Walp.: Sie enthält nur eine Art:
- Cucurbitella asperata (Gillies ex Hook. & Arn.) Walp.: Sie ist in Südamerika verbreitet.
- Dendrosicyos Balf.f.: Sie enthält nur eine Art:
- Dendrosicyos socotrana Balf.f.: Dieser Endemit kommt nur auf dem Socotra-Archipel vor.
- Dieterlea E.J. Lott (2 Arten in Nordamerika)
- Doyerea Grosourdy: Sie enthält nur eine Art:
- Doyerea emetocathartica Grosourdy ex Bello: Sie ist von Zentral- bis ins nördliche Südamerika verbreitet.
- Gurania (Schltdl.) Cogn. (35 Arten in Zentral- bis tropisches Südamerika)
- Guraniopsis Cogn.: Sie enthält nur eine Art:
- Guraniopsis longipedicellata Cogn.: Sie kommt in Peru vor.
- Halosicyos Mart. Crov.: Sie enthält nur eine Art:
- Halosicyos ragonesei Mart. Crov.: Sie kommt in Argentinien vor.
- Helmontia Cogn. (1 oder 2 Arten in Guyana und Brasilien)
- Ibervillea Greene (5 Arten in Texas, Mexiko, Guatemala)
- Kedrostis Medik. (20 bis 25 Arten im tropischen und subtropischen Afrika, Madagaskar, Indien, Sri Lanka, Westmalesien)
- Melothrianthus Mart. Crov.: Sie enthält nur eine Art:
- Melothrianthus smilacifolius (Cogn.) Mart. Crov.: Sie kommt in Brasilien vor.
- Psiguria Neck. ex Arn. (12 Arten in Zentral- bis tropisches Südamerika)
- Seyrigia Keraudren (4 Arten auf Madagaskar)
- Trochomeriopsis Cogn.: Sie enthält nur eine Art:
- Trochomeriopsis diversifolia Cogn.: Sie kommt auf Madagaskar vor.
- Tumamoca Rose (2 Arten in Nordamerika und Mexiko):
- Wilbrandia Silva Manso (5 Arten in Südamerika)
- Tribus Benincaseae
- Subtribus Benincasinae
- Acanthosicyos Welw. ex Benth. & Hook.f.: Die etwa zwei Arten kommen im südlichen Afrika vor.
- Bambekea Cogn. (2 Arten im tropischen Afrika)
- Wachskürbisse (Benincasa Savi): Sie enthält seit 2011 zwei Arten, beispielsweise:[14]
- Wachskürbis (Benincasa hispida (Thunb.) Cogn.): Er ist in Asien und Australien verbreitet.
- Borneosicyos W.J.de Wilde: Sie enthält nur eine Art:
- Borneosicyos simplex W.J.de Wilde: Sie kommt in Indonesien und Sabah vor.
- Cephalopentandra Chiov.: Sie enthält nur eine Art:
- Cephalopentandra ecirrhosa (Cogn.) C.Jeffrey: Sie ist im tropischen und subtropischen Afrika verbreitet.
- Citrullus Schrad. (3 Arten im tropischen und subtropischen Afrika)
- Coccinia Wight & Arn. (30 Arten im tropischen und subtropischen Afrika, Asien)
- Cogniauxia Baill. (2 Arten im tropischen Afrika)
- Ctenolepis Hook.f. (2 Arten im tropischen und subtropischen Afrika, Indien)
- Dactyliandra (Hook.f.) Hook.f. (2 Arten im tropischen und subtropischen Afrika, Indien)
- Diplocyclos (Endl.) Post & Kuntze (4 Arten im tropischen und subtropischen Afrika, Asien)
- Eureiandra Hook.f. (8 Arten im tropischen und subtropischen Afrika)
- Indomelothria W.J.de Wilde & Duyfjes (2 Arten in Südostasien)
- Khmeriosicyos W.J.de Wilde & Duyfjes: Sie enthält nur eine Art:
- Khmeriosicyos harmandii W.J.de Wilde & Duyfjes: Sie kommt nur in Kambodscha vor.
- Flaschenkürbisse (Lagenaria Ser.) (6 Arten im tropischen und subtropischen Afrika, Asien), darunter:
- Flaschenkürbis (Lagenaria siceraria (Molina) Standley)
- Lemurosicyos Keraudren: Sie enthält nur eine Art:
- Lemurosicyos variegata (Cogn.) Keraudren: Sie kommt auf Madagaskar vor.
- Neoachmandra W.J.de Wilde & Duyfjes (30 Arten in Afrika, Australien, pazifische Inseln)
- Nothoalsomitra I. Telford (1 Art im tropischen Australien):
- Nothoalsomitra suberosa (F.M.Bailey) I.Telford
- Papuasicyos Duyfjes: Sie enthält nur eine Art:
- Papuasicyos papuanus (Cogn.) Duyfjes: Sie kommt in Papua-Neuguinea vor.
- Peponium Engl. (20 Arten im tropischen Afrika, Madagaskar)
- Praecitrullus Pangalo: Sie enthält nur eine Art:
- Praecitrullus fistulosus (Stocks) Pangalo: Sie kommt in Indien sowie Pakistan vor.
- Raphidiocystis Hook.f. (5 Arten im tropischen Afrika, Madagaskar)
- Ruthalicia C.Jeffrey (2 Arten im tropischen Afrika)
- Scopellaria W.J.de Wilde & Duyfjes (2 Arten in Südostasien)
- Solena Lour. (3 Arten in China, Indochina, Indien)
- Trochomeria Hook.f. (8 Arten im tropischen und subtropischen Afrika)
- Urceodiscus W.J.de Wilde & Duyfjes (7 Arten in Neuguinea)
- Zombitsia Keraudren: Sie enthält nur eine Art:
- Zombitsia lucorum Keraudren: Sie kommt auf Madagaskar vor.
- Subtribus Cucumerinae
- Cucumeropsis Naudin: Sie enthält nur eine Art:
- Cucumeropsis mannii Naudin: Sie ist im tropischen Afrika verbreitet.
- Gurken (Cucumis L.) (52 Arten in Afrika, Madagaskar, Asien bis Nordaustralien; inklusive Cucumella Chiov. (11 Arten), Dicoelospermum C.B.Clarke (1 Art), Mukia Arn. (6 Arten), Myrmecosicyos C.Jeffrey (1 Art) und Oreosyce Hook.f. (1 Art), 2007 zu Cucumis gestellt[15])
- Melancium Naudin: Sie enthält nur eine Art:
- Melancium campestre Naudin: Sie kommt in Brasilien vor.
- Melothria L. (10 Arten im tropischen Zentral- und Südamerika)
- Muellerargia Cogn. (2 Arten auf Madagaskar, in Malaysia, Queensland)
- Posadaea Cogn.: Sie enthält nur eine Art:
- Posadaea sphaerocarpa Cogn.: Sie ist in Zentral- und Südamerika verbreitet.
- Zehneria Endl. (25 Arten im tropischen und subtropischen Afrika, Madagaskar, Asien)
- Cucumeropsis Naudin: Sie enthält nur eine Art:
- Subtribus Benincasinae
- Tribus Cucurbiteae
- Abobra Naudin: Sie enthält nur eine Art:
- Abobra tenuifolia (Gillies ex Hook. & Arn.) Cogn.: Sie ist in Südamerika verbreitet.
- Anacaona Alain: Sie enthält nur eine Art:
- Anacaona sphaerica Alain: Dieser Endemit kommt nur in der Dominikanischen Republik vor.
- Calycophysum H.Karst. & Triana (5 Arten von Kolumbien bis Bolivien)
- Cayaponia Silva Manso (60 Arten Nordamerika bis Argentinien, Westafrika, Madagaskar)
- Cionosicys Griseb. (3 Arten in Zentralamerika, Jamaika)
- Kürbisse und Zucchini (Cucurbita L.) (20 Arten von Nordamerika bis Argentinien)
- Penelopeia Urb.: Sie enthält nur eine Art:
- Penelopeia suburceolata Urb.: Dieser Endemit kommt nur in der Dominikanischen Republik vor.
- Peponopsis Naudin (1 Art in Mexiko):
- Peponopsis adhaerens Naudin
- Polyclathra Bertol. (1 Art von Mexiko bis Panama):
- Polyclathra cucumerina Bertol.
- Schizocarpum Schrad. (6 Arten in Mexiko)
- Selysia Cogn. (4 Arten von Nicaragua bis Peru)
- Sicana Naudin (1 Art von Mexiko bis Panama)
- Sicana odorifera Naudin
- Tecunumania Standl. & Steyerm. (1 Art in Zentralamerika):
- Tecunumania quetzalteca Standl. & Steyerm.
- Abobra Naudin: Sie enthält nur eine Art:
- Tribus Joliffieae
Diese auf morphologischen Merkmalen basierende Gliederung konnte von Kocyan u. a. 2007 in einer phylogenetischen Untersuchung, die 123 der 130 Gattungen umfasste, bis auf Tribus-Ebene im Wesentlichen bestätigen, die Subtribus sind allerdings künstliche Unterteilungen. Aus der Arbeit von Kocyan u. a. kann folgendes, auf die Triben vereinfachtes Kladogramm abgeleitet werden, wobei Wellenlinien Triben anzeigen, die paraphyletisch im Hinblick auf die anderen Äste derselben Klade sind:
| Nhandiroboideae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cucurbitoideae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Unter den von Jeffrey 2005 anerkannten Triben ist Jollifeae hochgradig polyphyletisch. Die Gattungen der Jollifeae bilden eine Gruppe an der Basis der Cucurbitoideae. Innerhalb dieser Gruppe sind einige Gattungen anderer Triben eingeordnet (Cogniauxia von den Benincaseae, Ampelosicyos und Tricyclandra von den Trichosantheae). Die übrigen Trichosantheae bilden eine Gruppe an der Basis der Sicyeae. Einige Gattungen sind nach den Ergebnissen von Kocyan u. a. para- oder polyphyletisch.
Nutzung
| Erntemengen und Anbauländer 2019[16] | ||
|---|---|---|
| Art | Ernte (t) | wichtigste Anbauländer |
| Wassermelonen | 100 Mio. | China, Türkei, Indien, Brasilien, Algerien |
| Gurken und Essiggurken | 88 Mio. | China, Türkei, Russland, Ukraine, Iran |
| Andere Melonen (z. B. Zuckermelonen) | 28 Mio. | Nigeria, China, Ghana, Elfenbeinküste, Benin |
| Kürbisse | 23 Mio. | China, Ukraine, Russland, Spanien, Mexico |
Die Nutzung der Kürbisgewächse durch den Menschen ist sehr vielfältig. Die Früchte werden reif oder unreif geerntet und gebacken (Kürbisse), frittiert (Bittermelone), gekocht (Wachskürbis, Schlangenhaargurke), gefüllt (Cyclanthera pedata), eingelegt (Gurke), kandiert (Wassermelone, Wachskürbis, Cucurbita ficifolia) oder frisch als Salat (Gurke) oder Dessert (Zuckermelone, Wassermelone) gegessen. Zur Haltbarmachung können sie zu Konserven verarbeitet, eingefroren oder eingelegt werden. Der Fruchtsaft einiger Arten wird zu alkoholischen Getränken vergoren, in Japan wird etwa aus der Zuckermelone Likör hergestellt.
Die Samen sind aufgrund ihres hohen Ölgehalts sehr nahrhaft. In Afrika können die Samen der Wassermelone, der Naras und anderer Arten einen wesentlichen Bestandteil der Nahrung ausmachen. In Mexiko und Zentralamerika, aber auch Teilen Mittel- und Südosteuropas werden Kürbissamen als Snack verspeist. Die Auslese von „nackten“, also dünnschaligen Samen hat die Beliebtheit auch in anderen Ländern gefördert. Aus den Samen folgender Arten wird Öl als Nahrung, zur Beleuchtung, Seifenherstellung und anderem hergestellt: Gartenkürbis (darunter auch der Steirische Ölkürbis), Wassermelone, Luffa, Fevillea cordifolia, Hodgsonia spp. und Cucumeropsis mannii. Neben Früchten und Samen werden auch Blüten, Blätter, Sprossspitzen und Wurzeln (Chayote) zubereitet.
Die Früchte des Flaschenkürbis werden ausgehöhlt zu Vorratsgefäßen, Flaschen, aber auch Tabak-Pfeifen, Musikinstrumenten, Penis-Futteralen, Masken, Bojen für Fischnetze, Baby-Rasseln usw. verarbeitet. Auch die Früchte des Wachskürbis und verschiedener Cucurbita-Arten werden als Vorratsgefäße verwendet. Der Schwammkürbis und Momordica angustisepala werden als Schwamm verwendet.
Die seit der Antike belegbare medizinische Verwendung der Kürbisgewächse ist äußerst vielseitig, sowohl bei den verwendeten Arten als auch bei den Anwendungsgebieten. Guha und Sen geben in ihrem auf Indien konzentrierten tabellarischen Überblick 79 Anwendungsgebiete an.[4] Weit verbreitet sind Anwendungen als Abführmittel, Emetikum und Wurmmittel.[17] Die Wirkung ist meist auf den Gehalt an Cucurbitacinen zurückzuführen. Die Wurzeln von Trichosanthes dioica werden als Gift verwendet.
Belege
Der Artikel beruht überwiegend auf folgenden Quellen:
- Die Familie der Cucurbitaceae bei der APWebsite. (engl.)
Literatur
- Alexander Kocyan, Li-Bing Zhang, Hanno Schaefer, Susanne S. Renner: A multi-locus chloroplast phylogeny for the Cucurbitaceae and its implications for character evolution and classification. In: Molecular Phylogenetics and Evolution. Band 44, August 2007, S. 553–577. doi:10.1016/j.ympev.2006.12.022, Volltext (Systematik; PDF; 381 kB)
- R. W. Robinson, D. S. Decker-Walters: Cucurbits. CAB International, Wallingford 1997, ISBN 0-85199-133-5. (Merkmale, Wachstum und Entwicklung, Nutzung)
- N. M. Nayar, T. A. More: Cucurbits. Science Publishers, Enfield 1998, ISBN 1-57808-003-7.
Einzelnachweise
- Alexander Kocyan, Li-Bing Zhang, Hanno Schaefer, Susanne S. Renner: A multi-locus chloroplast phylogeny for the Cucurbitaceae and its implications for character evolution and classification. In: Molecular Phylogenetics and Evolution. Band 44, August 2007, S. 553–577.
- Dieser Abschnitt beruht vorwiegend auf: Kocyan, Zhang, Schaefer, Renner: A multi-locus chloroplast phylogeny for the Cucurbitaceae and its implications for character evolution and classification. In: Molecular Phylogenetics and Evolution. Band 44, August 2007, S. 553–577.
- L.-B. Zhang, M. P. Simmons, A. Kocyan, S. S. Renner: Phylogeny of the Cucurbitales based on DNA sequences of nine loci from three genomes: Implications for morphological and sexual system evolution. In: Molecular Phylogenetics and Evolution. Band 39, 2006, S. 305–322. doi:10.1016/j.ympev.2005.10.002
- J. Guha, S. P. Sen: Physiology, biochemistry and medicinal importance. In: N. M. Nayar, T. A. More: Cucurbits. Science Publishers, Enfield 1998, ISBN 1-57808-003-7, S. 97–127.
- P. K. K. Nair, R. Satheesh: Palynology. In: N. M. Nayar, T. A. More: Cucurbits. Science Publishers, Enfield 1998, ISBN 1-57808-003-7, S. 85–95.
- 2007 – Joe Jutras and his 1689 pound World Record! www.bigpumpkins.com
- Susanne S. Renner, Hanno Schaefer, Alexander Kocyan: Phylogenetics of Cucumis (Cucurbitaceae): Cucumber (C. sativus) belongs in an Asian/Australian clade far from melon (C. melo). In: BMC Evolutionary Biology. Nr. 7, 2007, S. 58. doi:10.1186/1471-2148-7-58, (Volltext; pdf; 391 kB)
- Dietrich Frohne, Uwe Jensen: Systematik des Pflanzenreichs unter besonderer Berücksichtigung chemischer Merkmale und pflanzlicher Drogen. 4. neubearbeitete Auflage. Gustav Fischer, Stuttgart/Jena/New York 1992, ISBN 3-437-20486-6.
- Li-Bing Zhang, Mark P. Simmons, Alexander Kocyan, Susanne S. Renner: Phylogeny of the Cucurbitales based on DNA sequences of nine loci from three genomes: Implications for morphological and sexual system evolution. In: Molecular Phylogenetics and Evolution. Band 39, 2006, S. 305–322. doi:10.1016/j.ympev.2005.10.002
- Charles Jeffrey: A new System of Cucurbitaceae. In: Botanicheskii Zhurnal. Band 90, 2005, S. 332–335.
- Josef Domes: Momordica-Traktat. In: Verfasserlexikon, VI, Sp. 645 f.
- Josef Domes, Gundolf Keil: Der ‚Momordica-‘ oder ‚Balsamöl-Traktat‘. Ein Beispiel für die Indikationsübertragung vom Erdöl auf den Jerusalemsapfel. In: Gundolf Keil (Hrsg.): „Ein teutsch puech machen“. Untersuchungen zur landessprachlichen Vermittlung medizinischen Wissens. (= Ortolf-Studien. 1 ; = Wissensliteratur im Mittelalter. 11). Reichert, Wiesbaden 1993, ISBN 3-88226-539-6, S. 480–498.
- Hanno Schaefer, Ian R. H. Telford, Susanne S. Renner: Austrobryonia (Cucurbitaceae), a New Australian Endemic Genus, is the Closest Living Relative to the Eurasian and Mediterranean Bryonia and Ecballium. In: Systematic Botany. Band 33, 2008, S. 125–132. (Volltext; pdf; 456 kB)
- Hanno Schaefer, Susanne Renner: Phylogenetic relationships in the order Cucurbitales and a new classification of the gourd family (Cucurbitaceae). In: Taxon, Volume 60, Issue 1, 2011, S. 122–138. doi:10.1002/tax.601011 JSTOR 41059827 PDF.
- Hanno Schaefer: Cucumis (Cucurbitaceae) must include Cucumella, Dicoelospermum, Mukia, Myrmecosicyos, and Oreosyce: a recircumscription based on nuclear and plastid DNA data. In: Blumea. Band 52, 2007, S. 165–177. (online)
- nach: FAOSTAT-Produktionsstatistik, Daten für „watermelons“, „cucumbers and gherkins“, „other melons (inc. cantaloupes)“, „pumpkins, squash and gourds“, abgerufen 4. Februar 2021.
- R. W. Robinson, D. S. Decker-Walters: Cucurbits. CAB International, Wallingford 1997, ISBN 0-85199-133-5, S. 26.
Weblinks
- Informationen zur Familie der Cucurbitaceae von T.C.Andres. (Memento vom 28. Oktober 2011 im Internet Archive)
Weiterführende Literatur
- Amanuel G. Ghebretinsae, Mats Thulin, Janet C. Barber: Relationships of cucumbers and melons unraveled: molecular phylogenetics of Cucumis and related genera (Benincaseae, Cucurbitaceae). In: American Journal of Botany. Volume 94, Issue 7, 2007, S. 1256–1266. doi:10.3732/ajb.94.7.1256