Embryosack
Der Embryosack oder Megagametophyt ist der weibliche Gametophyt in der Samenanlage der Samenpflanzen. Er enthält die Eizelle, den weiblichen Gameten. Im Unterschied zu den sonstigen Teilen der Pflanze ist er nicht diploid, sondern haploid, denn die Embryosackzelle, aus der er sich entwickelt, geht aus einer Meiose (Reduktionsteilung) hervor. Der ebenfalls haploide männliche Gametophyt befindet sich im Pollenkorn.
Der Aufbau des Embryosacks wurde erstmals 1849 von Wilhelm Hofmeister beschrieben. 1851 zeigte er als Erster, dass bei den Samenpflanzen wie bei den Farnen und Moosen ein Wechsel von geschlechtlichen und ungeschlechtlichen Generationen (Generationswechsel) vorliegt und dass der Embryosack dem weiblichen Gametophyten der Farne und Moose entspricht (Homologie).[1] Den genauen Ablauf der Entwicklung beschrieb Eduard Strasburger 1879 zuerst bei einem Vogelknöterich (Polygonum).[2]
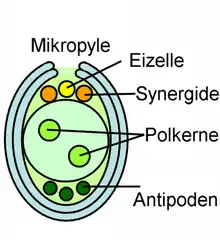
In der Embryosackzelle finden zunächst freie Kernteilungen (Mitosen) ohne Zellteilung statt, und erst später werden Zellmembranen und teils auch dünne Zellwände ausgebildet. Der vollständig entwickelte Embryosack besteht dann im häufigsten Fall (bei den meisten Bedecktsamern) aus sieben Zellen: der Eizelle und zwei Synergiden, die zusammen den Eiapparat bilden und an einem Ende des länglichen Embryosacks liegen, des Weiteren aus der großen Zentralzelle und drei Antipoden am anderen Ende. Die Zentralzelle enthält anfangs zwei Zellkerne, die dann miteinander zu einem diploiden Kern verschmelzen. Die Eizelle ist fast vollständig von den Synergiden umgeben, und der ganze Eiapparat ist, mit Ausnahme seiner Verbindung zur Wand des Embryosacks, von der Zentralzelle umschlossen.
Bei der Befruchtung bildet das Pollenkorn einen Pollenschlauch, der gewöhnlich durch die Mikropyle zum Eiapparat vordringt. Dort entlässt er zwei Spermazellen, die männlichen Gameten, in eine der Synergiden. Eine Spermazelle befruchtet dann die Eizelle, woraus die Zygote und aus dieser dann der diploide Embryo hervorgeht. Die andere Spermazelle befruchtet die diploide Zentralzelle, und daraus geht das triploide Endosperm als Nährgewebe hervor. Diese doppelte Befruchtung ist eine Besonderheit der Bedecktsamer, zu der keine Parallele bei anderen Lebewesen bekannt ist.
Bei den nacktsamigen Pflanzen (Gymnospermae), zu denen unsere Nadelbäume, die tropischen Palmfarne und der Ginkgo gehören, ist der Embryosack sehr viel größer und besteht aus zahlreichen Zellen. Dies ist der evolutionär ursprüngliche Zustand. Die winzigen, wenigzelligen Embryosäcke der Bedecktsamer sind das Ergebnis einer extremen Reduktion. Am weitesten geht diese bei den Nachtkerzengewächsen und den Seerosenartigen, bei denen die Antipoden fehlen und die Zentralzelle nur einen haploiden Kern enthält.[3]
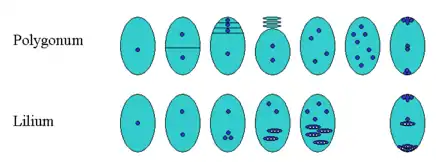
Neben dem oben beschriebenen Normalfall gibt es bei Bedecktsamern viele Abweichungen bei der Entwicklung des Embryosacks. Dies beginnt schon bei der vorangehenden Meiose. Aus dieser gehen gewöhnlich vier in einer Reihe liegende Tochterzellen hervor, von denen drei zugrunde gehen und die übrigbleibende als Embryosackzelle sich zum Embryosack entwickelt. Bei manchen Bedecktsamern unterbleibt jedoch die Zellteilung nach der zweiten meiotischen Kernteilung (Meiose II), und es entstehen zwei Tochterzellen mit jeweils zwei Kernen, von denen eine vergeht und die andere zum Embryosack wird. Im Extremfall findet auch nach der Meiose I keine Zellteilung statt, und es resultiert ein vierkerniger Coenocyt, der sich in vielfältiger Weise weiter entwickeln kann. Zum Beispiel vereinigen sich bei Lilien bei der ersten Kernteilung (Mitose) nach den beiden meiotischen Teilungen drei Kernteilungsspindeln, und es entstehen zwei triploide Tochterkerne (s. Abb.). Einen anderen Sonderfall zeigen die schon erwähnten Nachtkerzen: Da entwickelt sich – in Umkehrung der Verhältnisse beim Normaltyp (Polygonum) – nach der Meiose nur die oberste der vier Zellen weiter, und nach nur zwei Mitosen wird außer dem Eiapparat nur ein haploider Kern in der Zentralzelle angelegt, während Antipoden fehlen.[4]
Die größte Variabilität zeigen unter den Bestandteilen des Embryosacks die Antipoden. Vielfach sterben sie bald ab, in anderen Fällen entwickeln sie aber einen regen Stoffwechsel und scheinen an der Versorgung der Zentralzelle mit Nährstoffen beteiligt zu sein. Bei Gräsern vermehren sie sich bis zu annähernd hundert Zellen.[5] Die Synergiden haben eine essenzielle Funktion bei der Befruchtung (Siphonogamie), und man nimmt an, dass sie Substanzen ausscheiden, die den Pollenschlauch anlocken (Chemotropismus). Bei manchen Pflanzen (Bleiwurzgewächse) fehlen sie allerdings, und ihre Funktionen werden von der Eizelle übernommen.[6]
Literatur
- Lexikon der Biologie: Embryosack. Spektrum, Heidelberg 1999.
- M.T.M. Willemse, J.L. van Went: The female gametophyte. In: B.M. Johri (Hg.): Embryology of Angiosperms. Springer, Berlin/Heidelberg/New York/Tokyo 1984, Nachdruck 2011. S. 159–196.
Einzelnachweise
- Ilse Jahn (Hrsg.): Geschichte der Biologie. 3. Aufl., Sonderausgabe Nikol, Hamburg 2004, ISBN 978-3-937872-01-8, S. 315.
- B.M. Johri (Hg.): Embryology of Angiosperms. Springer, Berlin/Heidelberg/New York/Tokyo 1984, S. 1f.
- C. Baroux, C. Spillane, U. Grossniklaus: Evolutionary origins of the endosperm in flowering plants. In: Genome biology. Band 3, Nummer 9, August 2002, S. reviews1026, PMID 12225592, PMC 139410 (freier Volltext) (Review).
- B.M. Johri (Hg.): Embryology of Angiosperms. Springer, Berlin/Heidelberg/New York/Tokyo 1984, S. 159f. (Näheres zu Lilium auf S. 12.)
- B.M. Johri (Hg.): Embryology of Angiosperms. Springer, Berlin/Heidelberg/New York/Tokyo 1984, S. 188–190.
- B.M. Johri (Hg.): Embryology of Angiosperms. Springer, Berlin/Heidelberg/New York/Tokyo 1984, S. 178–181.