Griffel (Botanik)
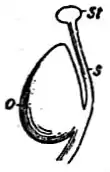
Der Griffel oder Stylus in einer Blüte ist der Teil eines Fruchtblatts oder Stempels, der die Narbe trägt. Der Begriff Stilus bedeutet ursprünglich Schreibstift oder Griffel, er wurde bereits von Clusius verwendet und setzte sich seit Malpighi und Jungius allgemein durch.[1]


.jpg.webp)
Ein Griffel kann als Teil des Fruchtblattes nur bei Bedecktsamern vorkommen. Auch bei nicht verwachsenen (apokarpen) Fruchtblättern kann der obere Bereich des Fruchtblattes, der keine Samenanlagen mehr beinhaltet, Griffel genannt werden. Meist wird der Begriff Griffel nur bei verwachsenen (coenokarpen) Fruchtblättern verwendet. Je nachdem, wie weit die Fruchtblätter miteinander verwachsen sind, können mehrere Griffel vorhanden sein. Sie können teilweise verwachsen sein und am oberen Ende freie Griffeläste bilden („Stylodien“), oder sie können zu einem einzigen Griffel verwachsen sein, an dessen Ende dann noch so viele Narben wie Fruchtblätter vorhanden sind, oder es kann auch nur ein einheitliches Narbengewebe vorhanden sein.
Stylodien, Stylodia werden teilweise auch als verlängerte Narben aufgefasst, die wie Griffeläste erscheinen; falsche Griffel (Stylode), auch bei reduzierten Pistilloden mit nur noch den (reduzierten) Griffelästen. Oder auch wenn jedes Fruchtblatt einen eigenen Griffel hat.
Der Griffel ist im Normalfall endständig, wenn er auf der Spitze des Fruchtknotens steht, oder seitenständig, wenn er neben oder unter der Spitze liegt, wie bei den Mangos (Mangifera), sowie grundständig, wenn er am Grunde des Fruchtknotens steht, sind hierbei die Fruchtblätter verwachsen und der Griffel steht am Blütenboden, dann nennt man diese Stellung gynobasisch, wie bei den Raublattgewächsen.[2] Bei wenigen Arten sind Zwischenformen möglich; subend-, -grund- oder -endständig; z. B. bei einigen Ficus-Arten, und subgynobasisch wie beim Lauch (Allium).[3][4][5]
Im Inneren des Griffels befindet sich das Leit- oder Transmissionsgewebe, das sich von der Narbe bis hinunter in den Fruchtknoten erstreckt. In diesem Transmissionsgewebe wachsen die Pollenschläuche zu den einzelnen Samenanlagen. Man unterscheidet hier meistens zwei Arten von Griffeln, in denen das Transmissionsgewebe unterschiedlich ist; üblicherweise solide mit einem (Compitum), aber auch hohle, hier ist ein spezieller Kanal (Lumen) vom Fruchtknoten zur Narbe vorhanden.[6][7] Von einigen Autoren wird noch ein halboffenes Gewebe unterschieden.[8][9]
Bei manchen Pflanzen, darunter die Doldengewächse und die Gattung Shorea, bildet sich ein Stylopodium (Griffelpolster, -fuß, -scheibe), eine kissenartige, kegel- oder scheibenförmige Verdickung, ein Podest an der Basis, am Fuß des Griffels, die oft diskusartig Nektar ausscheidet.
Bei einigen Pflanzenarten, z. B. bei den Asternartigen, können die Griffel, der meist zwittrigen und protandrischen Blüten, mit speziellen Haaren ausgestattet sein (Bärtig, Griffelbürste), die Haare (Sammelhaare, Fegehaare) dienen zum Auffangen des zuvor ausgetreuten Pollens der Antheren und übernehmen so die Funktion einer sekundären Pollenpräsentation.[2][10][11][12] Neuere Untersuchungen zeigen aber auch, dass sie zu einem effizienten Mechanismus gehören, der die Öffnung der Narbe kontrolliert und die Fremdbestäubung fördert. Eine berührende Stimulation der Haare bewirkt eine Verkürzung der männlichen Phase und beschleunigt die Narbenreifung. Je häufiger die Haare stimuliert werden, desto kürzer ist die männliche Phase.[13] Es kann auch ein Griffel-, Pollenbecher (oder Indusium)[14] vorhanden sein, ein Auswuchs, Saum am Griffelende, der die Narbe becherförmig umschließt, wie bei den Goodeniengewächsen. Auch kann ein löffel- oder hakenförmiger Auswuchs am Griffelende vorhanden sein, an den Pollen angelagert wird, wie bei den Kreuzblumengewächsen. Diese verschiedenen Pollenauffangstrukturen werden als „Pollen-Presenter“ bezeichnet.[12][15]
Bei wenigen Arten (Pfeilwurzgewächse Marantaceae) kann der Griffel sich bei Berührung eines Bestäubers irreversibel und explosionsartig einrollen und so Pollen vom Bestäuber abstreifen und in eine spezielle Höhlung befördern und gleichzeitig eigenen Pollen von der Pollenplatte abgeben, der hier vorher in der Blütenentwicklung von der Narbe abgegeben wurde.[16][17]
Auch möglich ist die Flexistylie, die bei einigen Arten vorkommt, abhängig vom Reifungsgrad der Staubblätter bewegt sich der Griffel hier hoch oder runter, wie beim Echten Galgant. Man vermutet, dass dadurch die Bestäubung mit eigenem Pollen, also eine Selbstbestäubung verhindert wird. Man kann noch unterscheiden in Cata- (bei protogynen Blüten) und Anaflexistylie (bei protandrischen Blüten).[18]
Bei einigen Arten (Apocynaceae) wird eine sogenannte „Clavuncula“ ausgebildet, hier verbreitert sich der Griffel direkt unterhalb der Narbe. Dies wird auch als Griffel- oder Narbenkopf bezeichnet.
Der Griffel fällt meistens nach der Befruchtung ab, er kann aber auch bleibend sein und dann welken oder fortwachsen.[19] Ist er bleibend sind die Früchte oft geschnäbelt.
Siehe auch
Literatur
- Peter Sitte, Elmar Weiler, Joachim W. Kadereit, Andreas Bresinsky, Christian Körner: Lehrbuch der Botanik für Hochschulen. Begründet von Eduard Strasburger. 35. Auflage. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X, S. 764 f.
Einzelnachweise
- Gerhard Wagenitz: Wörterbuch der Botanik. Die Termini in ihrem historischen Zusammenhang. 2. erweiterte Auflage. Spektrum Akademischer Verlag, Heidelberg / Berlin 2003, ISBN 3-8274-1398-2, S. 315.
- Hermann Hager: Botanischer Unterricht in 160 Lectionen. 3. Auflage, Springer, 1885, ISBN 978-3-642-89788-7 (Reprint), S. 198.
- Michael G. Simpson: Plant Systematics. Academic Press, 2006, ISBN 0-12-644460-9, S. 378 f.
- Chittaranjan Kole: Wild Crop Relatives: Genomic and Breeding Resources. Springer, 2011, ISBN 978-3-642-20449-4, S. 1.
- Rong Shu: Ficus. In: Flora of China. 5, 2003, S. 37–71, online (PDF; 493 kB), bei eFloras.org, abgerufen am 18. Mai 2018.
- K. R. Shivanna, Rajesh Tandon: Reproductive Ecology of Flowering Plants: A Manual. Springer, 2014, ISBN 978-81-322-2002-2, S. 51 ff.
- Focko Weberling: Morphology of Flowers and Inflorescences. Cambridge Univ. Press, 1989, ISBN 0-521-25134-6, S. 192.
- Valayamghat Raghavan: Molecular Embryology of Flowering Plants. Cambridge Univ. Press, 1997, ISBN 0-521-55246-X, S. 192.
- Peter K. Endress: Diversity and Evolutionary Biology of Tropical Flowers. Cambridge Univ. Press, 1994, 1998, ISBN 0-521-42088-1, S. 53 f, 76 f, 204.
- Veit M. Dörken: Campanulaceae – Glockenblumengewächse (Asterales). online (PDF; 1,0 MB), auf cms.uni-konstanz.de, abgerufen am 7. Mai 2018.
- Strasburger − Lehrbuch der Pflanzenwissenschaften. 37. Auflage, 2014, ISBN 978-3-642-54434-7, S. 714, eingeschränkte Vorschau in der Google-Buchsuche.
- P. G. Ladd: Pollen presenters in the flowering plants-form. In: Botanical Journal of the Linnean Society. 115(3), 1994, S. 165–195, doi:10.1006/bojl.1994.1040, online (PDF; 2,2 MB), bei Oxford Academic - Oxford University Press, abgerufen am 18. Mai 2018.
- Yvonne Nyman: The Pollen-Collecting Hairs of Campanula (Campanulaceae). II. Function and Adaptive Significance in Relation to Pollination. In: American Journal of Botany. Vol. 80, No. 12, 1993, S. 1437–1443, doi:10.2307/2445673.
- Eug. Warming: Handbuch der Systematischen Botanik. Bornträger, 1911, Salzwasser, 2012, ISBN 978-3-86444-713-6, S. 473.
- P. F. Yeo: Secondary Pollen Presentation: Form, Function and Evolution. Springer, 1993, 2012, ISBN 978-3-7091-7375-6 (Reprint), S. 140–150.
- Maria Thurner: Biologie der Marantaceae (Pfeilwurzgewächse) speziell Calathea sp. Bakkalaureatsarbeit, Univ. Wien, 2003, online (PDF), bei Universität Wien, abgerufen am 18. Mai 2018.
- Alexandra C. Ley, Regine Claßen-Bockhoff: Floral synorganization and its influence on mechanical isolation and autogamy in Marantaceae. In: Botanical Journal of the Linnean Society. Volume 168, Issue 3, 2012, S. 300–322, doi:10.1111/j.1095-8339.2011.01202.x.
- Michael Allaby: A Dictionary of Plant Sciences. Third Edition, Oxford Univ. Press, 2012, ISBN 978-0-19-960057-1, S. 198.
- Stephan Endlicher, Franz Unger: Grundzüge der Botanik. Gerold, 1843, S. 274–280.