Stoma (Botanik)
Ein Stoma oder eine Spaltöffnung (Plural Stomata; altgriechisch στόμα stóma, deutsch ‚Mund‘, ‚Mündung‘, ‚Öffnung‘) ist eine Pore in der Epidermis von Pflanzen. Der Stoma wird normalerweise von zwei bohnenförmigen Zellen, den Schließzellen, gebildet. Zählt man die Zellen, die um die Schließzellen herumliegen (Nebenzellen), noch hinzu, spricht man vom Spaltöffnungsapparat oder stomatären Komplex. Während die Epidermiszellen keine Chloroplasten enthalten, befinden sich in den ausdifferenzierten Schließzellen Chloroplasten.[1]
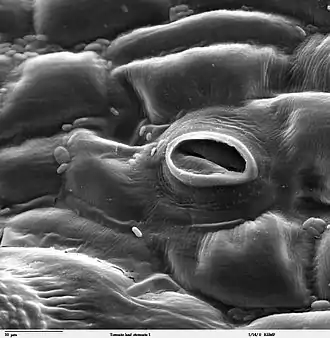
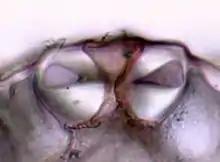
Die Stomata regulieren den Gasaustausch der Pflanze mit der Umgebungsluft. Im Allgemeinen handelt es sich dabei um die Abgabe von Sauerstoff und Wasser (Transpiration), sowie die Aufnahme von Kohlenstoffdioxid. Unabhängig von Pflanzentyp und der Anpassung an besondere Standortbedingungen erfolgt das Öffnen und Schließen der Stomata nach dem gleichen Mechanismus.
Die Stomata befinden sich bei den meisten Pflanzen in der unteren Epidermis der Blätter, bei Gräsern auf beiden Blattseiten und bei Schwimmblattpflanzen nur auf der Blattoberseite. Stomata sind auch in der Epidermis von Sprossachsen und Blütenblättern zu finden, jedoch nie an Wurzeln.
Bedeutung
Der Gasaustausch mit der umgebenden Luft ist insbesondere für die Versorgung mit CO2 wichtig. Kohlenstoffdioxid wird von den Pflanzen durch physikalische Prozesse (Gasaustausch) aus der Luft aufgenommen. Würde die CO2-Aufnahme ausschließlich über die Zellwände erfolgen, müssten diese extrem dünn sein, um eine ausreichende Versorgung der Pflanze zu gewährleisten. Dies hätte allerdings eine verringerte Stabilität sowie einen erhöhten Wasserverlust zur Folge. Über die Trennung des Interzellularraums im Blatt von der trockenen Außenluft durch die Stomata bekommt die Pflanze Kontrolle über ihren Wasserhaushalt.
Weitere Bedeutung: Durch die Poren findet Verdunstung (stomatäre Transpiration) statt, die einen Transpirationssog erzeugt, durch den Wasser von den Wurzeln bis in die Blätter transportiert wird. Mit dem Wasser werden Mineralstoffe aus dem Boden aufgenommen und in der Pflanze verteilt. Zusätzlich kühlt die Verdunstung die Blätter, diese überhitzen bei starker Sonneneinstrahlung nicht und das spezifische Temperaturoptimum der Enzyme in den Blattgeweben wird nicht überschritten.
Die Transpiration allein über die Fläche der Stomata, die nur ein bis zwei Prozent der gesamten Blattoberfläche ausmachen, beträgt bis zu zwei Drittel der Evaporation, also der widerstandslosen Verdunstung, einer gleich großen Wasserfläche. Untersuchungen haben gezeigt, dass an vielen kleinen Öffnungen bei gleicher Oberfläche mehr Wasser verdunstet. Der Grund ist der so genannte „Randeffekt“: Moleküle am Rand eines Stomas können auch zur Seite diffundieren, während die in der Mitte sich gegenseitig behindern.[2]
Der Anteil der kutikulären Transpiration ist sehr gering, bei Hygrophyten (Pflanzen in feuchten Gebieten) mit zarten Blättern weniger als zehn Prozent der Evaporation einer freien Wasserfläche, bei Bäumen weniger als 0,5 Prozent und bei Kakteen sogar nur 0,05 Prozent.
Struktur des Spaltöffnungsapparates
Die äußerste Zellschicht eines Blattes, die Epidermis, ist eine meist einlagige Schicht aus in der Regel chlorophyllfreien Zellen. Die Epidermis ist nach außen durch die Cuticula, eine nahezu wasserundurchlässige Schicht aus Cutin mit einer aufgelagerten Wachsschicht, abgegrenzt. Das zwischen den Epidermisschichten liegende Blattgewebe, das Mesophyll, besteht aus dem Palisadengewebe, in dem hauptsächlich die Photosynthese stattfindet, dem ebenfalls photosynthetisch aktiven Schwammgewebe und den Blattadern. Das Schwammgewebe ist mit Wasserdampf gesättigt und erleichtert die Diffusion aufgrund seiner erhöhten Oberfläche. Zwischen seinen Zellen befinden sich Interzellularräume, auch Atemhöhlen genannt, mit deren Luft der Gasaustausch erfolgt. Sie münden in die Stomata.
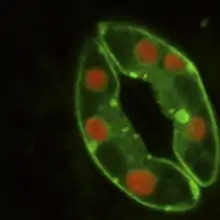
Der Spaltöffnungsapparat besteht aus zwei Schließzellen, in der Regel bohnenförmige Zellen, die an beiden Enden aneinander haften. Zwischen ihnen ist ein Interzellularspalt, der Porus, der die Verbindung zwischen Außenluft und Atemhöhle darstellt.
Bei manchen Pflanzen sind die beiden Schließzellen von spezialisierten epidermalen Zellen, den Nebenzellen (in den Abbildungen hellblau), umgeben, die an Öffnung und Verschluss der Spaltöffnung indirekt beteiligt sind. In den Nebenzellen lassen sich oft Leukoplasten erkennen.
Die Schließzellen enthalten Chloroplasten, können also Photosynthese durchführen. Die Öffnungsweite des Porus ist variabel, bei Sonnenlicht und genügender Wasserzufuhr sind sie in der Regel weit geöffnet, bei Nacht oder bei Wassermangel geschlossen.
Typen
Von der Form der Schließzellen kann man drei Haupttypen unterscheiden: Zu dem hantelförmigen Gramineen-Typ bei Gräsern und dem nierenförmigen Helleborus-Typ bei Ein- und Zweikeimblättrigen gesellt sich noch der Mniumtyp der Moose. Zuweilen trennt man davon noch den Xerophytentyp, der auf Nadelblättern zu finden ist und einen einzelligen Spaltöffnungsapparat, der bei wenigen Moosen und Farnen auftritt.
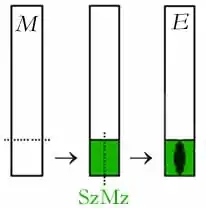 Entwicklung des Spaltöffnungsapparates bei Iris (Amaryllidaceen-Typ) |
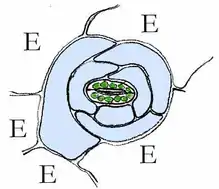 Spaltöffnungsapparat bei Kalanchoe (Crassulaceen-Typ) |
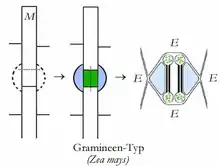 Spaltöffnungsapparat bei Zea mays (Poaceen- oder Gramineentyp) |
| Ranunculaceen-Typ | 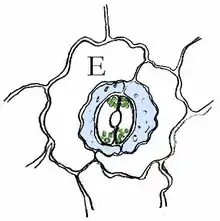 Spaltöffnungsapparat bei Coffea arabica (Rubiaceen-Typ) |
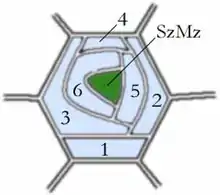 Entwicklungsschema des Spaltöffnungsapparates vom Brassicaceen-Typ (=Cruciferen-Typ) (Die Zahlen geben die Reihenfolge an, in der Nebenzellen entstanden sind.) |
| Caryophyllaceen-Typ | Rhoeo spathacea Tradescantia-Typ |
Abkürzungen:
E = Epidermiszelle
M = epidermale Meristemoide
SzMz = Schließzellen-Mutterzelle
Zuordnungskriterien sind Zahl und Anordnung der Nebenzellen:
| Typ | Beschreibung |
|---|---|
| Brassicaceen-Typ | anisocytisch (gr. an ‚nicht‘, isos ‚gleich‘) |
| Ranunculaceen-Typ | anomocytisch: ohne Nebenzellen |
| cyclocytisch: (gr. kyklos ‚Kreis‘) | |
| Caryophyllaceen | diacytisch (gr. diá ‚zwischen‘) 2 Nebenzellen um 90° versetzt zu den Schließzellen |
| Rubiaceen-Typ | paracytisch (gr. pará ‚neben‘) 2 Nebenzellen parallel zu den Schließzellen |
| tetracytisch (gr. tetra ‚vier‘) |
Blatt-Typen nach Lage der Stomata
Je nach Lage der Stomata kann man drei Blatt-Typen unterscheiden.
- Hypostomatisch
- Stomata liegen nur auf der Blattunterseite (häufig)
- Epistomatisch
- Stomata liegen nur auf der Blattoberseite (selten) z. B. bei Seerosen
- Amphistomatisch
- Stomata liegen auf der Blattoberseite und auf der -unterseite. z. B. Gräser, Nadelblätter
Entstehung
Nebenzellen entstehen aus epidermalen Meristemoiden (embryonalen Zellen der Epidermis) durch inäquale, also ungleiche Zellteilungen, deren Anzahl je nach Typ variiert.
Die Zellen des Spaltöffnungsapparates können auf zwei verschiedene Arten entstehen:
- Fortlaufende Zellteilungen einer Meristemoide
- Die letzte Zellteilung ergibt die Schließzellen-Mutterzelle. Beispiel: Brassicaceen-Typ
- Abgliederung der Nebenzellen von benachbarten Meristemoiden
- Beispiel: Gramineen-Typ
Als Letztes entstehen aus der Schließzellen-Mutterzelle die beiden Schließzellen durch Einziehen von zwei Zellwänden in der Mitte (äquale Teilung, es entstehen also zwei gleich große Tochterzellen), die durch Auflösen der Mittellamelle voneinander getrennt werden und damit die Spaltöffnung bilden.
Allgemeiner Mechanismus
Wasserverlust führt zu einer Verringerung des Innendrucks (Turgordruck) der Schließzellen. Bei niedrigem Turgor stehen ihre Zellwände in direktem Kontakt miteinander, wodurch der Zentralspalt geschlossen ist. Eine Erhöhung des Turgors durch Wasseraufnahme in die Zelle führt aufgrund der speziellen Zellform und den ungleichmäßig stark verdickten Zellwänden zu einer Verformung. Die Zellen wölben sich im zentralen Kontaktbereich voneinander weg. Der Spalt öffnet sich.
Verbildlichen lässt sich dieses Prinzip anhand eines Fahrradschlauchs. Hält man den luftleeren Schlauch in der Hand hängt er schlaff nach unten – seine Innenflächen berühren sich. Wird er aufgepumpt, lässt sich das Auseinanderbewegen beider Seiten erkennen. Somit vergrößert sich der zentrale Raum, der dem Zentralspalt des Stomas entspricht.
Molekulare Mechanismen
Die Öffnungsweite der Spaltöffnungen wird durch Lichtstärke, Lichtqualität, Wasserversorgung und die CO2-Konzentration gesteuert. Dabei spielen auch die Phytohormone Auxin und Abszisinsäure eine Rolle.[3]
Soll die Spaltöffnung erweitert werden, fällt das Membranpotential ab und Kalium-Ionen strömen ins Innere der Schließzellen. Als Ladungsausgleich strömen auch Anionen, überwiegend Chlorid nach innen und Malationen werden synthetisiert. Aufgrund der erhöhten Ionenkonzentration strömt nun Wasser über Aquaporine zunächst ins Cytosol und dann in die Vakuole (Osmose) ein.
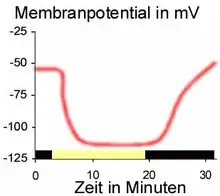
Im Dunkeln weist die Schließzelle ein negatives Membranpotential von −55 mV auf. Bei einem Lichtreiz verstärkt sich diese negative Spannung auf −110 mV (Hyperpolarisation). Diese Hyperpolarisation erfolgt durch eine lichtgesteuerte ATPase, die Protonen unter ATP-Verbrauch von innen nach außen pumpt. Das dazu notwendige ATP könnte aus der Photosynthese der Schließzellen stammen, die in der Regel als einzige Epidermiszellen Chloroplasten aufweisen.
Die Hyperpolarisation ist notwendig, da im Cytosol die K+-Konzentration höher (ca. 100 mM) als im Apoplasten (ca. 1 mM) – der Quelle der Kalium-Ionen – ist. Die Kaliumkanäle öffnen sich ab −100 mV, und die Kaliumionen können gegen ihr Konzentrationsgefälle, aber mit dem Potentialgefälle von außen nach innen diffundieren.
Auslöser für das Öffnen dieser Kaliumkanäle ist eine erhöhte Protonenkonzentration (also eine pH-Erniedrigung) im Apoplasten. Die Protonen lösen aber nicht nur die Öffnung aus, sondern erleichtern sie auch, indem sie das Schwellenpotential zur Öffnung zu positiveren Werten verschieben.
Im Dunkeln stellt die ATPase ihre Tätigkeit ein, das Membranpotential steigt wieder auf −55 mV und die Kaliumionen strömen wieder entsprechend ihrem Konzentrationsgefälle nach außen. Gleichzeitig wandern auch die Chloridionen nach außen, Wasser strömt jetzt wieder nach außen, der Turgordruck sinkt und die Spaltöffnung schließt sich.
Bei KST1 (Kalium-Kanal von Solanum tuberosum) liegt ein Einwärtsgleichrichter mit pH-Sensor vor. Bei dem entsprechenden Kanal von Arabidopsis werden höhere Protonenkonzentration im Apoplasten zum Öffnen des Kanals benötigt.
Beispiel C4-Pflanze: Bei Sonneneinstrahlung wird in den Schließzellen Kohlenstoffdioxid an Phosphoenolpyruvat (PEP) gebunden und wird zu Äpfelsäure. Diese dissoziiert zu Malat und es werden H+-Ionen freigesetzt. Die H+-Ionen werden durch Ionenpumpen in der Membran mit Energieaufwand (Aufspaltung von ATP in ADP und P) in die Nachbarzellen befördert. Dadurch entsteht eine negative Ladung in den Schließzellen, durch die positive K+-Ionen angezogen werden. Sie diffundieren in das Zellinnere und erhöhen so den osmotischen Wert. Dadurch diffundiert Wasser aus den Nachbarzellen in die Schließzellen, diese erweitern sich bis zu ihrem doppelten Volumen und geben so die Spaltöffnungen frei. Solange Sonne auf die Schließzelle scheint, wird durch diese Reaktionen das Stoma offengehalten, je stärker die Sonne, desto praller sind die Schließzellen und umso weiter ist die Spaltöffnung geöffnet. Lässt die Lichtintensität nach, so finden alle Reaktionen nicht mehr in vollem Umfang statt, der osmotische Wert (Turgordruck) der Schließzellen nimmt ab und sie werden schlaff – die Stomata schließen sich. Nimmt stattdessen die Wasserzufuhr der Pflanze ab, verringert sich insgesamt der osmotische Druck (Turgor) der Pflanzenzellen, weniger Wasser diffundiert in die Schließzellen und die Stomata schließen sich ebenfalls. Durch die Verengung der Stomata transpiriert die Pflanze weniger, sie trocknet langsamer aus.
Literatur
- Strasburger: Lehrbuch der Botanik, 29. Auflage neubearbeitet von v. Denffer et al., Gustav Fischer, Stuttgart 1967, S. 87–90 (C 1. a) Spaltöffnungsapparate), S. 217–218, S. 244, S. 361.
- G. Czihak, H. Langer, H. Ziegler (Hrsg.): Biologie. Ein Lehrbuch. 6. unveränderte Auflage, Springer, Berlin/Heidelberg/New York 1996, S. 420–424.
Weblinks
Einzelnachweise
- Ulrich Kutschera: Kurzes Lehrbuch der Pflanzenphysiologie. Quelle&Meyer, Wiesbaden 1995, S. 383.
- Gerhard Richter: Stoffwechselphysiologie der Pflanzen. Georg Thieme, Stuttgart 1976, S. 295.
- Rainer Franz Hedrich: Über den Stoffwechsel von Schließzellen im Licht und im Dunkeln. Univ., Diss., Göttingen 1985.