Rapoport-Luebering-Zyklus
Der Rapoport-Luebering-Zyklus, auch als Rapoport-Luebering-Shunt, Rapoport-Luebering-Shuttle, Phosphoglyceratzyklus oder 2,3-BPG-Zyklus bezeichnet, ist in der Biochemie ein vor allem in roten Blutkörperchen (Erythrozyten) von Säugetieren ablaufender Stoffwechselweg, also eine Abfolge von enzymatisch gesteuerten chemischen Reaktionen. Es handelt sich um einen aus drei Teilreaktionen bestehenden Nebenweg der Glycolyse, die für die Energiegewinnung und den Kohlenhydratstoffwechsel fast aller Lebewesen von zentraler Bedeutung ist. Der Rapoport-Luebering-Zyklus gehört somit zu den biochemischen Prozessen zum Abbau von Traubenzucker im tierischen Organismus.
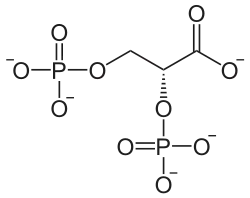
Seine Hauptreaktion ist die durch das Enzym Bisphosphoglyceratmutase gesteuerte Bildung des Zwischenprodukts 2,3-Bisphosphoglycerat (2,3-BPG) aus dem in der Glycolyse entstehenden 1,3-Bisphosphoglycerat. Das im Rapoport-Luebering-Zyklus entstehende 2,3-BPG wirkt als wichtiger biochemischer Effektor in der Regulation der Bindungsfähigkeit (Affinität) des Blutfarbstoffs Hämoglobin für das Atemgas Sauerstoff, insbesondere für deren längerfristige Anpassung an Sauerstoffmangelzustände, und ist damit für die Freisetzung des Sauerstoffs aus den Erythrozyten ins Gewebe von Bedeutung. Es ist darüber hinaus an der enzymatischen Steuerung der Glycolyse beteiligt und fungiert in den Erythrozyten als Energie- und Phosphatspeicher.
Die Entdeckung des Rapoport-Luebering-Zyklus und der Bedeutung des 2,3-BPG für den Energiehaushalt der roten Blutkörperchen in den 1940er Jahren durch den Biochemiker Samuel Mitja Rapoport und seine Assistentin Janet Luebering war medizinisch von großer Bedeutung, da durch das Verständnis dieser Prozesse die Haltbarkeit von Blutkonserven erheblich gesteigert werden konnte.
Biochemische Aspekte
Ablauf
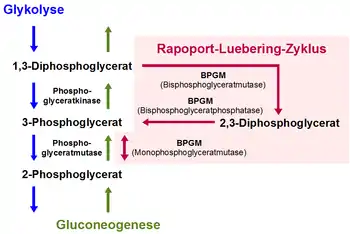
Beim Rapoport-Luebering-Zyklus handelt es sich um einen Nebenweg der Glycolyse in den roten Blutkörperchen der Säugetiere einschließlich des Menschen. Er führt ausgehend vom 1,3-Bisphosphoglycerat (1,3-BPG) aus der Glycolyse zur Bildung von 2,3-Bisphosphoglycerat (2,3-BPG). Aus diesem entstehen die zum Reaktionsablauf der Glycolyse gehörenden Phosphoglycerinsäure-Verbindungen 3-Phosphoglycerat (3-PG) und, durch dessen Isomerisierung, 2-Phosphoglycerat (2-PG).[1]
Das für diese Reaktionen verantwortliche Enzym Bisphosphoglyceratmutase (BPGM) ist in seinem Vorkommen im Wesentlichen auf Erythrozyten und erythropoetisches Gewebe beschränkt und weist als trifunktionales Enzym drei verschiedene Aktivitäten auf.[2][3] Es fungiert in Abhängigkeit vom pH-Wert entweder als Synthase (2,3-BPG-Synthase, synonym Bisphosphoglyceratmutase; EC-Nummer 5.4.2.4) für die Umwandlung des 1,3-BPG zu 2,3-BPG oder als Phosphatase (2,3-Bisphosphoglyceratphosphatase; EC-Nummer 3.1.3.13) für die Umsetzung des 2,3-BPG zu 3-PG. Darüber hinaus katalysiert sie als Mutase (Monophosphoglyceratmutase; EC-Nummer 5.4.2.1) die Gleichgewichtsreaktion zwischen 3-PG und 2-PG.[3]
Hauptaktivität der BPGM ist dabei die Synthasereaktion vom 1,3-BPG zu 2,3-BPG, die irreversibel verläuft. Der letzte Schritt im Rapoport-Luebering-Zyklus, die Umsetzung von 3-PG zu 2-PG, ist über das Enzym Phosphoglyceratmutase eine auch in anderen Zellen ablaufende Teilreaktion der Glycolyse. Für die Phosphoglyceratmutase, die der BPGM hinsichtlich ihrer Molekülmasse, ihrer Untereinheitenstruktur und ihrer Aminosäuresequenz ähnelt, wurden darüber hinaus auch geringe Aktivitäten als 2,3-BPG-Synthase und -phosphatase gefunden.[4] Sie fungiert somit wahrscheinlich ähnlich der BPGM als trifunktionales Enzym, jedoch mit einem anderen Verhältnis der drei Enzymaktivitäten zueinander. Dies gilt, neben der Expression der BPGM in einigen nicht-erythropoetischen Geweben wie in der Plazenta und in der Leber, als mögliche Erklärung für das Auftreten geringer Mengen von 2,3-BPG in nicht-erythroiden Zellen.[5] Die Rückreaktionen vom 2-PG über 3-PG zum 1,3-BPG und damit der Teilprozesse der Glycolyse, die parallel zum Rapoport-Luebering-Zyklus ablaufen, finden im Rahmen der Gluconeogenese statt.
Bilanz
Der erste Schritt des Rapoport-Luebering-Zyklus, die Umlagerung vom 1,3-BPG zum 2,3-BPG, ist eine Isomerisierung mit neutraler stofflicher Bilanz. Die Bisphosphoglyceratmutase als Enzym dieser Reaktion benötigt allerdings das Vorhandensein von Magnesium-Ionen.[6] Die hydrolytische Spaltung vom 2,3-BPG zum 3-PG im zweiten Schritt verläuft unter Verbrauch eines Wassermoleküls und Freisetzung eines anorganischen Phosphats. Im Rapoport-Luebering-Zyklus entsteht – im Gegensatz zur Umsetzung von 1,3-BPG zu 3-PG durch die Phosphoglyceratkinase in der Glycolyse – kein Adenosintriphosphat (ATP). Die Energieausbeute ist damit beim Nebenweg über das 2,3-BPG geringer als beim direkten Weg in der Glycolyse.
Regulation
Die im Rapoport-Luebering-Zyklus entstehenden Verbindungen 2,3-BPG und 3-PG inhibieren diesen Nebenweg, der damit autoregulatorisch verläuft.[7] 2,3-BPG hemmt außerdem einige in der Reaktionsfolge der Glycolyse vor dem Rapoport-Luebering-Zyklus liegende Enzyme wie beispielsweise die Hexokinase und die Phosphofructokinase.[1] Darüber hinaus fungiert es als Kofaktor der Phosphoglyceratmutase in der Glycolyse.[8] Ein Anstieg der 1,3-BPG-Menge stimuliert die Entstehung von 2,3-BPG. Alle Prozesse der Glycolyse, die über Aktivierung oder Hemmung von Enzymen zu einer Erhöhung der 1,3-BPG-Konzentration führen, beschleunigen damit die 2,3-BPG-Bildung.[7]
Auch durch einen Anstieg des pH-Wertes entsteht vermehrt 2,3-BPG, da der optimale pH-Wert für die Synthase-Aktivität der BPGM bei etwa 7,2 liegt, während die Phosphatase-Aktivität ihr Optimum im sauren Bereich hat und dann gegenüber der 2,3-BPG-Bildung überwiegt. Die Hormone Thyroxin, Somatotropin, Testosteron und Erythropoietin stimulieren ebenfalls die Bildung des 2,3-BPG.[9] Durch Chlorid, Phosphat und vor allem durch den physiologischen Phosphataseaktivator 2-Phosphoglycolat kommt es hingegen zu einer verstärkten Spaltung von 2,3-BPG zu 3-PG durch die Phosphatase-Funktion der BPGM.[3]
Bedeutung
Physiologische Funktion
Da die roten Blutkörperchen der Säugetiere im Gegensatz zu den meisten anderen Körperzellen nicht über einen Zellkern oder über Mitochondrien verfügen, besitzen sie einen spezialisierten Kohlenhydrat- und Energiestoffwechsel ohne Citratzyklus und Atmungskette. Die Glycolyse ist in den Erythrozyten neben dem Pentosephosphatweg die einzige Möglichkeit zur Energiegewinnung.[10] Etwa 20 Prozent des in den Erythrozyten in der Glycolyse entstehenden 1,3-BPG wird dabei über den Rapoport-Luebering-Zyklus umgesetzt, der Anteil des gebildeten 2,3-BPG beträgt etwa 50 Prozent aller Intermediate der Glycolyse in den Erythrozyten[1] und etwa zwei Drittel der gesamten erythrozytären Phosphate.[11] Unter physiologischen Bedingungen liegt 2,3-BPG in den roten Blutkörperchen etwa in der gleichen molaren Konzentration wie der Blutfarbstoff Hämoglobin und rund dem Vierfachen der ATP-Konzentration vor.[7] Die Menge an 2,3-BPG wird bestimmt durch das Verhältnis zwischen der Synthase- und der Phosphatase-Aktivität der BPGM.
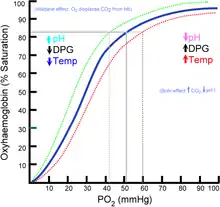
grün: Linksverschiebung
rot: Rechtsverschiebung
Das im Rapoport-Luebering-Zyklus entstehende 2,3-BPG fungiert vor allem als allosterischer Inhibitor des Hämoglobins, indem es dessen nicht mit Sauerstoff beladene Deoxy-Form stabilisiert, und reguliert damit umgekehrt proportional zu seiner Konzentration in den roten Blutkörperchen die Bindungsfähigkeit (Affinität) des Hämoglobins für Sauerstoff.[7] 2,3-BPG bindet dabei zwischen den beiden Beta-Untereinheiten des Hämoglobins in der Tasche, die sich in dem auch als T-Form bezeichneten unbeladenen Zustand bildet.[12] Die biophysikalische Grundlage der Bindung sind Wechselwirkungen zwischen den negativ geladenen Gruppen des 2,3-BPG und positiv geladenen Aminosäureresten in der Bindungstasche. Ein Anstieg der 2,3-BPG-Konzentration verschiebt die Sauerstoffbindungskurve des Hämoglobins nach rechts, wodurch der gebundene Sauerstoff leichter abgegeben wird. Umgekehrt führt ein Absinken der 2,3-BPG-Konzentration zu einer Linksverschiebung der Sauerstoffbindungskurve und damit einer stärkeren Bindung des Sauerstoffs an das Hämoglobin.
Weitere Faktoren, die zu einem Anstieg der Sauerstoffaffinität des Hämoglobins führen und dabei zum Teil auch den 2,3-BPG-Spiegel beeinflussen, sind eine Abnahme der Temperatur, ein Anstieg des pH-Wertes und ein Absinken der Kohlendioxidkonzentration. Die kombinierte Wirkung des pH-Werts und des Kohlendioxidpartialdrucks auf die Bindungsfähigkeit des Hämoglobins für Sauerstoff wird auch als Bohr-Effekt bezeichnet und ist die physikochemische Grundlage der Regulation des Gasaustausches in der Lunge sowie der Versorgung stoffwechselaktiver Gewebe mit Sauerstoff. Kohlenmonoxid verringert hingegen die Bindungskapazität des Hämoglobins für Sauerstoff, da es mit dem Sauerstoff um die gleiche Bindungsstelle im Hämoglobinmolekül konkurriert. Ein Anstieg der 2,3-BPG-Menge verbessert die Sauerstoffabgabe in der Körperperipherie und damit die Sauerstoffversorgung in den Geweben, insbesondere unter ungünstigen Bedingungen wie Zuständen, die mit einem Sauerstoffmangel einhergehen. So führt beispielsweise ein Aufenthalt in höheren Lagen zu einer Steigerung der 2,3-BPG-Konzentration, die etwa zwei Tage nach Rückkehr auf die Ausgangshöhe wieder Normalwerte erreicht.[7] Auch körperliche Kurz- oder Langzeitbelastung sowie Ausdauertraining haben in unterschiedlichem Ausmaß Auswirkungen auf die Konzentration des 2,3-BPG.[13]
Neben dieser Funktion als Kompensationsmechanismus spielt der Rapoport-Luebering-Zyklus wahrscheinlich auch eine Rolle in der Regulation der Stoff- und Energiebilanz der Glycolyse.[9][13] So ermöglicht er eine verstärkte Bildung des Koenzyms Nicotinamidadenindinukleotid (NADH) in der Glycolyse ohne eine nachfolgende Zunahme der ATP-Konzentration sowie den Ablauf der Glycolyse auch bei geringem ATP-Bedarf. Darüber hinaus stellt 2,3-BPG einen erythrozytären Energie- sowie Phosphatspeicher dar.
Medizinische Relevanz
Enzymdefekte in denjenigen Reaktionen der Glycolyse, die nach der Bildung des 2,3-BPG ablaufen, bewirken durch einen Anstieg von dessen Konzentration eine Abnahme der Sauerstoffaffinität von Hämoglobin und damit eine gesteigerte Freisetzung von Sauerstoff im Gewebe.[1] Umgekehrt führen Defekte in den Reaktionen der Glycolyse vor dem Rapoport-Luebering-Zyklus zu einer Abnahme der 2,3-BPG-Konzentration und damit zu einer Verminderung der Sauerstoffabgabe im Gewebe.
Eine gezielte Regulierung der Bisphosphoglyceratmutase zur Beeinflussung der 2,3-BPG-Konzentration in den Erythrozyten wäre beispielsweise therapeutisch von Interesse zur Behandlung von Ischämien und der Sichelzellenanämie.[3][14] Für Diabetes-Patienten wurde eine Verringerung der BPGM-Aktivität durch Glykation beschrieben.[2] Eine angeborene Defizienz der BPGM ist bisher erst in wenigen Fällen dokumentiert worden.[15] Die betroffenen Personen waren abgesehen von einer sekundären Erythrozytose (vermehrte Bildung roter Blutkörperchen) weitestgehend beschwerdefrei. Eine labormedizinische Bestimmung des 2,3-BPG in Erythrozyten und im Serum ist möglich, aufgrund der geringen diagnostischen Aussagekraft jedoch nicht gebräuchlich und nur für spezielle Fragestellungen von Interesse.
Das 2,3-BPG in den Erythrozyten hat ebenso wie das ATP Einfluss auf die Lagerungsfähigkeit von Blutkonserven. Aufgrund des Anstiegs der Laktatkonzentration mit fortschreitender Lagerungsdauer verschiebt sich der pH-Wert des entnommenen Bluts in den sauren Bereich, wodurch das 2,3-BPG verstärkt abgebaut und seine Neubildung gehemmt wird. Durch die Zugabe von Zusätzen wie Dextrose und Adenin, wie sie beispielsweise in den gegenwärtig verwendeten CPDA- beziehungsweise CPD/SAGM-Blutbeuteln enthalten sind, lässt sich die Abnahme des 2,3-BPG-Gehaltes verzögern und damit die Haltbarkeit und Funktion von gelagertem Blut verbessern.[16]
Veterinärphysiologische Aspekte
Die Konzentration von 2,3-BPG in den Erythrozyten und das Ausmaß seiner Wirkung auf das Hämoglobin unterscheidet sich zwischen verschiedenen Säugetieren.[9][13][17] Die Hämoglobine von Menschen, Pferden, Hunden, Schweinen, Kaninchen, Meerschweinchen, Mäusen und Ratten, deren Erythrozyten eine hohe 2,3-BPG-Konzentration aufweisen, reagieren dementsprechend stark. Demgegenüber ist die Wirkung des 2,3-BPG auf das Hämoglobin ebenso wie der 2,3-BPG-Gehalt in den roten Blutkörperchen von Schafen, Ziegen und Rindern, von Hirschen, Antilopen und Giraffen sowie von Hyänen und Katzen geringer.
In Vögeln fungiert 2,3-BPG nur während der Embryonalentwicklung als Regulator der Sauerstoffaffinität des Hämoglobins. Wenige Tage nach dem Schlüpfen aus dem Ei wird es dann vollständig abgebaut, die Funktion des 2,3-BPG übernehmen im weiteren Leben Inositolphosphate wie Inositolhexaphosphat (IHP).[18] In Fischen wurde 2,3-BPG nur in wenigen Arten gefunden, die dominierenden Organophosphate in Fischerythrozyten sind ATP und Guanosintriphosphat (GTP).[19] In den Erythrozyten von Reptilien kommen als Organophosphate vorwiegend ATP, IHP und Myo-Inositol-5-Phosphat (IP5) vor.
Als Grund für die Unterschiede zwischen Säugetieren und anderen Wirbeltieren gilt der besondere Energiestoffwechsel der Erythrozyten der Säugetiere. In den kernhaltigen Erythrozyten anderer Wirbeltiere ist die Atmungskette der primäre energieliefernde Stoffwechselweg und nicht, wie in den roten Blutkörperchen der Säugetiere, die Glycolyse.[19]
Entdeckungsgeschichte
_507-516.jpg.jpg.webp)
2,3-BPG, das Reaktionsprodukt des Rapoport-Luebering-Zyklus, wurde 1925 erstmals beschrieben und isoliert,[20] die Ausgangssubstanz 1,3-BPG durch Erwin Negelein im Jahr 1939.[21] Der in Österreich geborene Biochemiker Samuel Mitja Rapoport und seine damalige technische Assistentin Janet Luebering entdeckten dann in den 1940er Jahren in den USA die für die 2,3-BPG-Bildung wesentlichen Reaktionen und beschrieben diese Anfang der 1950er Jahre in mehreren gemeinsamen Veröffentlichungen.[22][23] Die Erforschung dieses Stoffwechselweges führte zur Entwicklung des Citrat- und Dextrose-haltigen ACD-Mediums, mit dem die Haltbarkeit von Blutkonserven von einer auf rund drei Wochen gesteigert werden konnte. Aufgrund der Bedeutung dieser Entdeckung für die Militärmedizin während des Zweiten Weltkrieges wurde Samuel Mitja Rapoport vom US-Präsidenten Harry S. Truman mit dem „President’s Certificate of Merit“ geehrt.[24]
Aufgrund seiner politischen Überzeugungen ging Samuel Mitja Rapoport, der 1937 ein einjähriges Stipendium am Kinderkrankenhaus der University of Cincinnati erhalten hatte und nach der deutschen Annexion Österreichs wegen seiner jüdischen Abstammung nicht nach Europa zurückgekehrt war, im Jahr 1952 in die Deutsche Demokratische Republik (DDR). Er wurde hier zu einem der führenden Biochemiker des Landes und führte seine Forschung zum Stoffwechsel der roten Blutkörperchen weiter. Zusammen mit seiner als Kinderärztin tätigen Frau Ingeborg Rapoport und seinem Sohn Tom Rapoport, der 1995 an die Harvard University wechselte, veröffentlichte er unter anderem in den 1970er Jahren Beiträge zur pH-Abhängigkeit der 2,3-BPG-Bildung und zur Regulation der Glycolyse in den Erythrozyten.
Die Eigenschaften der Bisphosphoglyceratmutase als dem zentralen Enzym des Rapoport-Luebering-Zyklus und ihre trifunktionale Aktivität wurden in den 1960er und 1970er Jahren näher charakterisiert.[4][25] 1967 erfolgte die Aufklärung der Wirkung von 2,3-BPG auf Hämoglobin,[26] 1978 wurde das angeborene Auftreten einer vollständigen BPGM-Defizienz in einem Patienten beschrieben.[27] Zehn Jahre später erfolgte die Isolierung und Charakterisierung des auf dem menschlichen Chromosom 7 liegenden Gens für das Enzym.[5] Die molekularen Grundlagen der Funktion der BPGM wurden in den 1990er Jahren näher untersucht,[14][28] 2004 wurde die Kristallstruktur des Enzymmoleküls aufgeklärt.[3] Vier Jahre später wurde beschrieben, dass das in verschiedenen Geweben vorkommende Enzym Multiple Inositol-Polyphosphat-Phosphatase (MIPP) ebenfalls eine Aktivität als 2,3-BPG-Phosphatase aufweist.[29] Diese Entdeckung ist für die Regulation der Abgabe von Sauerstoff aus dem Hämoglobin und damit für die physiologische Rolle des Rapoport-Luebering-Zyklus von Bedeutung.
Einzelnachweise
- R. van Wijk, W.W. van Solinge: The energy-less red blood cell is lost: erythrocyte enzyme abnormalities of glycolysis. In: Blood. 106(13)/2005. American Society of Hematology, S. 4034–4042.
- T. Fujita et al.: Human Erythrocyte Bisphosphoglycerate Mutase: Inactivation by Glycation In Vivo and In Vitro. In: Journal of Biochemistry. 124(6)/1998. Japanese Biochemical Society, S. 1237–1244.
- Y. Wang et al.: Crystal Structure of Human Bisphosphoglycerate Mutase. In: Journal of Biological Chemistry. 279/2004. The American Society for Biochemistry and Molecular Biology, S. 39132–39138.
- R. Sasaki, K. Ikura, E. Sugimoto, H. Chiba: Purification of bisphosphoglycerate mutase, bisphosphoglycerate phosphatase and phosphoglycerate mutase from human erythrocytes: three enzyme activities in one protein. In: European Journal of Biochemistry. 50(3)/1975. Federation of European Biochemical Societies, S. 581–593.
- V. Joulin et al.: Isolation and characterization of the human 2,3-bisphosphoglycerate mutase gene. In: Journal of Biological Chemistry. 263/1988. The American Society for Biochemistry and Molecular Biology, S. 15785–15790.
- Gerhard Michal: Biochemical Pathways: Biochemie-Atlas. Spektrum Akademischer Verlag, Heidelberg 1999, ISBN 3-86-025239-9, S. 27/28.
- Georg Löffler, Petro E. Petrides: Biochemie und Pathobiochemie. 7. Auflage. Springer-Verlag, Berlin und Heidelberg 1998, ISBN 3-540-42295-1, S. 986 und 994/995.
- H. Chiba, R. Sasaki: Functions of 2,3-bisphosphoglycerate and its metabolism. In: Current Topics in Cellular Regulation. 14/1978. Academic Press, S. 75–116.
- Larry Rex Engelking: Review of Veterinary Physiology. Teton NewMedia, Jackson WY 2002, ISBN 1-89-344169-5, S. 130.
- Gerhard Thews, Ernst Mutschler, Peter Vaupel: Anatomie, Physiologie und Pathophysiologie des Menschen. Wissenschaftliche Verlagsgesellschaft, Stuttgart 1999, ISBN 3-8047-1616-4, S. 117.
- John P. Greer, Maxwell Myer Wintrobe: Wintrobe's Clinical Hematology. Lippincott Williams & Wilkins, Philadelphia 2009, ISBN 0-78-176507-2, S. 143.
- Albert L. Lehninger, David L. Nelson, Michael M. Cox: Prinzipien der Biochemie. Zweite Auflage. Spektrum Akademischer Verlag, Heidelberg 1994, ISBN 3-86-025106-6, S. 267/268.
- Nemi C. Jain: Essentials of Veterinary Hematology. Lea & Febiger, Philadelphia 1993, ISBN 0-81-211437-X, S. 145.
- P. Ravel, C.T. Craescu, N. Arous, J. Rosa, M.C. Gare: Critical Role of Human Bisphosphoglycerate Mutase Cys22 in the Phosphatase Activator-binding Site. In: Journal of Biological Chemistry. 272/1997. The American Society for Biochemistry and Molecular Biology, S. 14045–14050.
- Bisphosphoglycerate Mutase Deficiency of Erythrocyte. In: Online Mendelian Inheritance in Man. (englisch), OMIM-Eintrag zur BPGM-Defizienz (englisch).
- J.R. Hess, T.G. Greenwalt: Storage of Red Blood Cells: New Approaches. In: Transfusion Medicine Reviews. 16(49)/2002. Elsevier, S. 283–295.
- Diphosphoglycerate Pathway. In: Jiro J. Kaneko, John W. Harvey, Michael Bruss: Clinical Biochemistry of domestic Animals. Fünfte Auflage. Academic Press, San Diego 1997, ISBN 0-12-396305-2, S. 178–180.
- R.E. Isaacks, L.L. Lai, P.H. Goldman, C.Y. Kim: Studies on avian erythrocyte metabolism. XVI. Accumulation of 2,3-bisphosphoglycerate with shifts in oxygen affinity of chicken erythrocytes. In: Archives of Biochemistry and Biophysics. 257(1)/1987. Academic Press, S. 177–185.
- Organic Phosphate Effects on Oxygen Affinity. In: Stephen C. Wood, Claude Lenfant: Evolution of Respiratory Processes. A Comparative Approach. Informa Health Care, 1979, ISBN 0-82-476793-4, S. 212–214.
- R. Juel: 2,3-Diphosphoglycerate: its role in health and disease. In: CRC Critical Reviews in Clinical Laboratory Sciences. 10(2)/1979. CRC Press, S. 113–146.
- Erwin Negelein, Heinz Brömel: R-Diphosphoglycerinsäure, ihre Isolierung und Eigenschaften. In: Biochemische Zeitschrift. 303/1939. Springer, S. 132–144.
- S. Rapoport, J. Luebering: The formation of 2,3-diphosphoglycerate in rabbit erythrocytes: The existence of a diphosphoglycerate mutase. In: Journal of Biological Chemistry. 183/1950. S. 507–516.
- S. Rapoport, J. Luebering: Glycerate-2,3-diphosphatase. In: Journal of Biological Chemistry. 189/1951. S. 683–694.
- A. Tuffs: Samuel Mitja Rapoport. Nachruf in: British Medical Journal. 329/2004. BMJ Group, S. 353.
- Z.B. Rose: The Purification and Properties of Diphosphoglycerate Mutase from Human Erythrocytes. In: Journal of Biological Chemistry. 243(18)/1968. The American Society for Biochemistry and Molecular Biology, S. 4810–4820.
- Reinhold Benesch, Ruth Benesch: The effect of organic phosphates from the human erythrocyte on the allosteric properties of hemoglobin. In: Biochemical and Biophysical Research Communications. 26(2)/1967. Academic Press, S. 162–167.
- R. Rosa, M.-O. Prthu, Y. Beuzard, J. Rosa: The first case of a complete deficiency of diphosphoglycerate mutase in human erythrocytes. In: Journal of Clinical Investigation. 62/1978. American Society for Clinical Investigation, S. 907–915.
- M.C. Garel, V. Lemarchandel, M.C. Calvin, N. Arous, C.T. Craescu, M.O. Prehu, J. Rosa, R. Rosa: Amino acid residues involved in the catalytic site of human erythrocyte bisphosphoglycerate mutase. Functional consequences of substitutions of His10, His187 and Arg89. In: European Journal of Biochemistry. 213(1)/1993. Federation of European Biochemical Societies, S. 493–500.
- J. Cho, J.S. King, X. Qian, A.J. Harwood, S.B. Shears: Dephosphorylation of 2,3-bisphosphoglycerate by MIPP expands the regulatory capacity of the Rapoport-Luebering glycolytic shunt. In: Proceedings of the National Academy of Sciences. 105(16)/2008. United States National Academy of Sciences, S. 5998–6003.
Weblinks
- UniProt: Bisphosphoglycerate mutase - Homo sapiens (Human) UniProt-Informationen zur Bisphosphoglyceratmutase (englisch)