Glyptodontidae
Die Glyptodontidae oder Glyptodonten sind eine ausgestorbene Familie der Nebengelenktiere (Xenarthra) und waren in Südamerika und in einigen Teilen Nordamerikas verbreitet. Sie stehen in der näheren Verwandtschaft mit den heute noch lebenden Gürteltieren und besaßen wie diese einen ausgebildeten Körperpanzer, der aber im Gegensatz zu dem der Gürteltiere starr war und über keine beweglichen Bänder verfügte. Darüber hinaus war auch der Schwanz vollständig gepanzert, einige Formen wiesen ein keulenartiges, verlängertes Ende auf. Die Vertreter der Familie erreichten vor allem im Pleistozän teilweise riesige Ausmaße und wogen bis zu 2 t, womit sie die größten Mitglieder der Gepanzerten Nebengelenktiere stellten. Neben einigen Merkmalen des Schädels verfügten die Glyptodonten auch über Besonderheiten im Skelettbau, wozu unter anderem elefantenähnliche Hinterbeine und eine extrem stark verknöcherte Wirbelsäule zu zählen sind. Als möglicherweise angepasste Grasfresser hatten sie zudem Zähne mit hohen Kronen. Die Glyptodonten bewohnten meist offene, von kühlem bis hin zu tropischem Klima beeinflusste Landschaften und kamen auch in höheren Gebirgslagen vor.
| Glyptodontidae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion mit Panzer von Glyptodon | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mittleres Eozän bis Unteres Holozän | ||||||||||||
| 48 Mio. Jahre bis 8.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Glyptodontidae | ||||||||||||
| Gray, 1869 | ||||||||||||
Die ersten Vertreter der Familie sind aus dem Mittleren Eozän vor rund 48 Millionen Jahren bekannt und wurden in Patagonien entdeckt, wo wahrscheinlich auch der Ursprung der Gruppe liegt. Die jüngsten Funde stammen aus dem Beginn des Holozän und sind rund 7500 Jahre alt. Während der Stammesgeschichte kam es zu einer starken Aufgliederung der Glyptodonten in mehrere Entwicklungslinien. Im Pliozän vor rund 3,5 Millionen Jahren erreichten einige Vertreter durch die Entstehung einer Landbrücke auch Nordamerika, wo sie sich aber nicht sehr lange hielten. Da häufig nur Reste des Panzers fossil belegt sind, ist die systematische Gliederung nur ungenügend untersucht und basiert weitgehend auf der Gestaltung und den Formenveränderungen der einzelnen Elemente der Rücken- und der Schwanzpanzerung. Insgesamt repräsentieren die Glyptodonten neben den Faultieren aber eine der formenreichsten Gruppen der Nebengelenktiere.
Die Forschungsgeschichte der Glyptodonten reicht bis in die zweite Hälfte des 18. Jahrhunderts zurück und begann mit der Entdeckung eines Rückenpanzers in der Pamparegion. Weitere Funde folgten Anfang des 19. Jahrhunderts, der für die Familie namengebende Gattungsvertreter Glyptodon erhielt 1839 durch Richard Owen seine Beschreibung. Diese baute auf einem Teilskelett aus der Pamparegion südlich von Buenos Aires auf, das nach England verbracht worden war. Einen wichtigen Beitrag leistete Charles Darwin auf seiner Reise nach Südamerika, der zahlreiche Reste von Glyptodonten in der Pampa barg. Die ersten Funde in Nordamerika gelangen erst im Übergang vom 19. zum 20. Jahrhundert. Im gleichen Zeitraum erarbeitete Florentino Ameghino eine erste systematische Untergliederung der Glyptodonten, die als Grundlage für zahlreiche weitere Forschungen diente.
Merkmale
Allgemein und Körpergröße
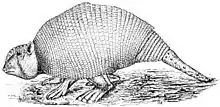
Die Glyptodontidae ähnelten den heutigen Gürteltieren (Dasypoda), mit denen sie enger verwandt sind, waren aber bedeutend größer und besaßen einen kurzschnauzigen Schädel und einen starren, kuppelartig gewölbten Rückenpanzer sowie einen vollständig gepanzerten, vergleichsweise kurzen Schwanz. Frühe Vertreter dieser Familie waren noch verhältnismäßig klein. So erreichte Cochlops aus dem Unteren Miozän eine Gesamtlänge von rund einem Meter bei einem Gewicht von knapp 90 kg, das etwa gleichalte Eucinepeltus brachte bis zu 115 kg auf die Waage.[1] Vor allem die Formen des späten Pleistozäns besaßen dem gegenüber aber riesige Ausmaße. Panochthus wurde über 2,6 m lang, während das Gewicht zwischen 1,1 und 1,3 t betrug. Größere Ausmaße besaß Doedicurus mit einer Länge von mehr 3,3 bis 4 m und einer Höhe von 1,5 m. Es wog zwischen 1,4 und 1,76 t, wobei für einen sehr späten Angehörigen der gleichen Gattung anhand eines fragmentierten Oberarmknochens ein Gewicht von 2,3 t ermittelt werden konnte.[2][3] Die größten Vertreter von Glyptodon brachten es auf eine Länge von über 3,5 m bei einem Gewicht von rund 2 t.[4] Damit waren die Glyptodonten die größten Vertreter der Gepanzerten Nebengelenktiere (Cingulata) und deutlich größer als die verwandten Gürteltiere und Pampatheriidae.[5]
Schädel- und Gebissmerkmale
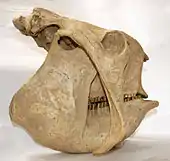
Der Schädel der Glyptodonten war massiv und dabei kurz und hoch. Er wies dadurch einen sehr charakteristischen Bau auf, der zusammen mit dem Unterkiefer fast einem Kubus ähnelte. Nur stammesgeschichtlich ältere Formen besaßen noch ein etwas verlängertes Rostrum und ähnelten dadurch den heutigen Gürteltieren.[6] Kleinere Formen wie Eosclerocalyptus wiesen Schädellängen von etwa 22 cm auf, bei großen wie Panochthus erreichten sie mit bis zu 42 cm gut die doppelte Länge. Charakteristisch war vor allem der deutlich verkürzte rostrale Bereich, wodurch der Gesamtschädel wie teleskopartig zusammengeschoben wirkte.[7][8] Ein weiteres markantes Merkmal stellten die überaus massigen Jochbögen dar, die weit auskragten und bei Panochthus einen Abstand von 31 cm erreichten.[9][10] Als auffällig erwies sich hier ein zweiter Knochenauswuchs, der vom vorderen Ansatz des Jochbogens weit nach unten bis unterhalb der Zahnreihe des Unterkiefers reichte und vorne und hinten verschmälert war. Eine ähnliche Bildung kann bei den Faultieren beobachtet werden, doch ist diese seitlich abgeflacht. In beiden Fällen dient der Auswuchs als Ansatzstelle des Musculus masseter der Kaumuskulatur. Da der Muskel aber in beiden Gruppen unterschiedlich angeordnet ist, geht man bei dem knöchernen Fortsatz von jeweils unabhängigen Bildungen aus.[11] Daneben treten bei den Glyptodonten weitere Schädelmerkmale auf, die sie von anderen Säugetieren unterscheiden. Vor allem die Knochenabschnitte im Bereich des Rostrums, so der Oberkiefer und das Gaumenbein, zeigten vertikal ausgerichtete Verlängerungen, um die extrem hochkronigen (hypdodonten) Zähne aufzunehmen. Ebenso waren die Stirnhöhlen stark vergrößert, was möglicherweise eine Rolle beim Zerkauen der Nahrung spielte. Der Unterkiefer war massiv gestaltet, am horizontalen Knochenkörper aufgrund der Zähne äußerst hoch und mit einer robusten Symphyse versehen. Das Gelenkende ragte steil auf, weit über die Höhe der Zahnreihe hinaus. Aufgrund des ungewöhnlichen Baus des Schädels besaßen die Glyptodonten im Bezug auf die Schädellänge den im Verhältnis am höchsten gelegenen Ansatzpunkt für die Gelenke des Unterkiefers (craniomandibulare Verbindung) innerhalb der Säugetiere.[7][12]
Wie bei allen Nebengelenktieren wich das Gebiss in seinem Aufbau von dem der anderen Höheren Säugetiere ab. Schneide- und Eckzähne waren nicht ausgebildet. Die hintere Bezahnung war homodont gestaltet, so dass zwischen Prämolaren und Molaren nicht unterschieden werden kann. Prinzipiell standen in jeder Kieferhälfte acht molarenartige Zähne, insgesamt verfügte das Gebiss also über 32 Zähne. Eine Besonderheit war das Fehlen von Zahnschmelz, was ebenfalls ein generelles Merkmal der Nebengelenktiere ist. Die Zähne wurden weitgehend aus Orthodentin und Osteodentin, die beiden härtesten Varianten des Zahnbeins, gebildet. Mit Ausnahme der vordersten beiden Zähne wiesen die Kauflächen aller Backenzähne drei querstehende lappenartige (trilobate) Bildungen auf. Jede einzelne Lobe bestand aus einem Mantel aus deutlich mineralisiertem Orthodentin (ursprünglich auch als Zahnzement beschrieben[7]) und einem Inneren aus weniger stark ausgehärtetem Zahnbein. Im Zentrum erhob sich jeweils eine kleine, querstehende Rippe aus sehr hartem Osteodentin, wobei eine entlang der Zahnmittelachse verlaufende Leiste die drei Rippen miteinander verband. Damit unterschieden sich die Zähne der Glyptodonten von denen der heutigen Gürteltiere mit ihrer generell stift- oder nagelartigen Form. Sie ähnelten dagegen den Backenzähnen der Pampatheriidae, ebenfalls panzertragende Verwandte der Gürteltiere, die zwei querstehende Loben besaßen. Generell waren die Zähne sehr hochkronig und wurzellos und wuchsen dadurch lebenslang, wodurch es wie bei anderen Nebengelenktieren auch nicht zu einem Zahnwechsel kam.[7][13][14] Im Gegensatz zu anderen Gruppen der Höheren Säugetiere sind bei Glyptodonten nur wenige Fälle bei der Ausbildung überzähliger Zähne bekannt. Eines der wenigen Beispiele liegt von einem Individuum der Gattung Boreostemma aus dem Mittleren Miozän vor, das im Oberkiefer einen zusätzlichen vorderen Zahn besaß.[15]
Körperskelett

Besondere Skelettmerkmale befinden sich an der Wirbelsäule. Charakteristisch für alle Gepanzerten Nebengelenktiere ist die Fusion der Wirbel der Halswirbelsäule, wobei in der Regel der Atlas (vorderster Halswirbel) frei beweglich ist, die hinteren aber eine zusammengewachsene Knochenstruktur darstellen. Zusätzlich waren bei den Glyptodonten noch der zweite, dritte und vierte Brustwirbel fest verwachsen (trivertebrales Element). Darauf folgte eine knöcherne Röhre, bestehend aus neun Brustwirbeln. Die Anzahl der Lendenwirbel variierte innerhalb der Glyptodonten auf Artebene, Vertreter der nordamerikanischen Gattung Glyptotherium wiesen zwischen 5 und 9 auf. Allerdings waren die Wirbel der Lendenwirbelsäule ebenfalls fest verwachsen, womit die Glyptodonten den höchsten Grad an Wirbelfusionen unter allen Säugetieren zeigten. Dadurch traten bei diesen aber die für die Nebengelenktiere typischen und namengebenden xenarthrischen Gelenke (Nebengelenke oder Xenarthrale) an den seitlichen Gelenkfortsätzen der Lenden- und der hinteren Brustwirbel nicht auf, was als einzigartiges Merkmal anzusehen ist.[11]
Der Bewegungsapparat zeigte vor allem an den Hinterbeinen Besonderheiten. Diese waren ähnlich den heutigen Elefanten gestaltet und wiesen Anpassungen an einen extrem schweren Körperbau auf. In den Proportionen übertrafen die Glyptodonten dabei die heutigen Elefanten und gehörten damit zu den am stärksten auf einen schweren Gang angepassten Landwirbeltieren. Weiterhin war das Becken vollständig mit dem Panzer verwachsen und dadurch unbeweglich.[12] Am Oberschenkelknochen bildete sich typisch für Nebengelenktiere ein dritter Trochanter als Muskelansatzstelle aus, der aber sehr weit unten lag und kontinuierlich in das untere Gelenkende überging. Schien- und Wadenbein waren fest an den Enden miteinander verwachsen. Die Hände und Füße besaßen einen relativ ursprünglichen Aufbau ohne stärkere Spezialisierungen. Die jeweils ersten Phalangen (Finger- und Zehenglieder) waren deutlich gekürzt.[11] Finger und Zehen endeten in eher huf- als krallenartigen Bildungen.[12][16] Insgesamt erinnerten die Vorder- und Hinterfüße an jene der Elefanten. Frühe Glyptodonten hatten jeweils fünfstrahlige Hände und Füße (pentadactyl). Die späteren Formen reduzierten an den Autopodien den innersten Strahl (Strahl I) und verfügten demzufolge über vierstrahlige Hände und Füße (tetradactyl), mit Ausnahme von Glyptodon und seiner näheren Verwandtschaft, die den pentadactylen Hinterfuß beibehielten.[6][17]
Panzer



Die Gepanzerten Nebengelenktiere sind die einzigen Säugetiere, bei denen ein äußerer, knöcherner Panzer ausgebildet ist. Im Gegensatz zu den heutigen Gürteltieren war der Rückenpanzer bei den Glyptodonten starr und unbeweglich. Er bedeckte den gesamten Rumpf der Tiere und bestand aus einem Mosaik von mehreren Hundert bis zu 1800 einzelnen Knochenplättchen (Osteoderme),[12] die sich in der Haut bildeten und meist wie bei den heutigen Gürteltieren mit Horn überzogen waren. Die Form des Panzers variierte und war bei Doedicurus etwa stark kuppelartig aufgewölbt, bei Glyptodon weniger deutlich und bei Neosclerocalyptus markant flach. Bei größeren Formen wie Panochthus und Glyptodon maß der Panzer zwischen 127 und 194 cm in der Länge, wobei sich unter Berücksichtigung der Krümmung die Werte auf 146 bis 220 cm belaufen.[18][19] Auch die Knochenplättchen zeigten je nach Gattung eine unterschiedliche Gestaltung, häufig besaßen sie aber einen vieleckigen Umriss. Teilweise wiesen sie oberflächlich eine auffällig rosettenartige Ornamentierung mit einer mehr oder weniger zentral gelegenen Musterung auf, um die konzentrisch in einer oder mehreren Reihen weitere angeordnet waren. Die einzelnen Musterungen hoben sich durch Furchen (Sulci) voneinander getrennt ab. Im Querschnitt hatten die Osteoderme analog zu denen der Gürteltiere einen mehrlagigen Aufbau: Innen und außen befand sich eine feste Knochenschicht, dazwischen ein Raum mit luftgefüllten Kammern, in denen möglicherweise auch Schweiß- und Talgdrüsen sowie Haarfollikel eingebettet waren. Dieser Innenraum nimmt bis zur Hälfte des Volumens des gesamten Knochenplättchens ein. Bei den Glyptodonten war er wesentlich regelmäßiger aufgebaut als im Vergleich zu den Gürteltieren.[20]

Auch der Schädel war durch einen Kopfschild geschützt, der den Kopf helmartig bedeckte. Zudem sind bei einigen Vertretern auch auf der Bauchseite und an den Beinen Osteoderme nachgewiesen, die aber unregelmäßig gestaltet und nicht miteinander verwachsen waren.[21] Der eher kurze Schwanz war ebenfalls vollständig gepanzert. Die Panzerung hier bestand aus mehreren knöchernen Ringen, die lose miteinander verbunden waren und dem Schwanz so eine hohe Flexibilität gaben. Jeder Ring war aus zwei oder drei Reihen von Knochenplättchen aufgebaut. Bei einigen Gattungen wie Glyptodon wies der Schwanz ein relativ kurzes Ende auf. Andere Formen wie Doedicurus, Panochthus oder Castellanosia hatten nur im vorderen Bereich Ringe ausgebildet. Die Osteoderme des hinteren Schwanzteils waren bei diesen dagegen komplett verwachsen und umhüllten den Schwanz vollständig. Sie bildeten eine röhrenartige, knöcherne Keule, die bis zu über einen Meter lang werden konnte und einen Durchmesser von bis zu 30 cm besaß. Teilweise war das Ende auch verdickt, häufig befanden sich aber auf der Oberfläche ovale Eintiefungen mit aufgerauter Oberfläche. Bei Hoplophorus und Panochthus ragten aus diesen Vertiefungen kegelförmig gestaltete Knochenbildungen auf, bei anderen Vertretern nehmen einige Wissenschaftler an, dass dort ursprünglich dornartige Stachel aus Keratin ausgebildet waren. Die Schwanzgestaltung ist einmalig unter den Säugetieren und stellt eine konvergente Entwicklung zu den Ankylosauriern dar.[22][21] Diese Konvergenzen betreffen etwa die Versteifung und Verdickung des hinteren Schwanzabschnittes, aber auch die Teilverwachsung der Rückenwirbel und die Ausbildung eines knöchernen Hautpanzers sowie eine enorme Körpergewichtszunahme.[23][24]
Verbreitung und Lebensraum
Die Familie der Glyptodontidae lebte ausschließlich auf dem amerikanischen Doppelkontinent. Sie entstand im Eozän in Südamerika, möglicherweise in der Region des heutigen Patagonien, und breitete sich von dort sukzessive nach Norden und Süden aus. Bedeutend sind hier die Funde der Santa-Cruz-Formation im Übergang vom Unteren zum Mittleren Miozän im Süden des Kontinents, deren zahlreichen Funde angeben, dass die Glyptodonten in ihrer frühen Entwicklungsphase damals in einem Mosaik aus offenen und teils geschlossenen Landschaften lebten.[1] Im Pliozän erreichten sie erstmals Nordamerika, was durch die Schließung des Isthmus von Panama und der Entstehung einer Landbrücke ermöglicht wurde, wonach der Große Amerikanische Faunenaustausch einsetzte. Dadurch besaßen die Glyptodonten im Pleistozän ihr größtes Verbreitungsgebiet und kamen dann von Südamerika vom 49. südlichen Breitengrad bis nach Nordamerika etwa um den 36. nördlichen vor. Sie bewohnten damals weitgehend offene Landschaften in sowohl kühlen bis kalten, trockenen, wüstenartigen Regionen, kamen aber auch in tropischen Gebieten vor. Zudem waren sie in dieser Zeit auch in der Andenregion auf bis mehr als 3300 m,[25][26] teilweise auch bis über 4000 m, über Meeresspiegelhöhe anzutreffen, was zu den höchsten Nachweisen von Nebengelenktieren gehört.[18][19] Eine mitunter für einige Formen angenommene Lebensweise in sumpfigen Gebieten oder Auenlandschaften[12] wird meist mit Hinweis auf den Bau der Gliedmaßen und den Anpassungen an offene Landschaften abgelehnt.[27]
Paläobiologie
Fortbewegung

Heutige Gürteltiere sind mit wenigen Ausnahmen, etwa den Kugelgürteltieren, gute Gräber, die mit spezialisierten Vorderfüßen und robusten Vordergliedmaßen an eine grabende Lebensweise angepasst sind. Dazu gehören unter anderem der kräftige und teilweise verlängerte Mittelfinger und der sehr ausgedehnte obere Gelenkfortsatz der Elle, das Olecranon. Es wird angenommen, dass die Grabfähigkeit ursprünglich bei allen Gepanzerten Nebengelenktieren ausgebildet war und erst später eine Anpassung an rein bodenbewohnende Lebensweisen erfolgte. Ein Großteil der Glyptodontidae, so unter anderem die frühen Vertreter wie Propalaehoplophorus, aber auch zahlreiche spätere Formen besitzen ein moderat gebautes Olecranon, das nicht auf spezialisierte Gräber hinweist. Ebenso ist der Aufbau der Hand eher ursprünglich und somit abweichend von den Gürteltieren. Die extrem großen späten Formen wie Glyptodon und Doedicurus weisen aber massige obere Ulnagelenkfortsätze auf, die bei jeweils 24 cm Knochenlänge bis zu 11 cm lang sind. Hier wird argumentiert, dass dieser kräftig gebaute Vorderarm eine Anpassung an das sehr hohe Körpergewicht ist. Zudem diente er wohl auch dazu, den Körper abzufedern, wenn ein Tier aus einem zweifüßigen Stand in eine vierfüßige Position zurückkehrte.[28][1] Dass sich die Glyptodonten auf die Hinterbeine aufrichten konnten, zeigen vor allem die extrem starken Knochen der Hinterbeine, die auch nach histologischen Untersuchungen an Gliedmaßenelementen von Lomaphorus deutlich kompakter gebaut sind als die Vorderbeine.[29] Zudem befindet sich das Zentrum der Körpermasse sehr weit hinten am Rumpf und unterstützte so das Aufrichten. Auch die heutigen Gürteltiere und Ameisenbären vermögen in eine zweibeinige Position zu wechseln, die überwiegend bei der Nahrungssuche und zur Verteidigung eingenommen wird.[4][30]
Häufig wird der Bau des Innenohrs für die Bewegungsfähigkeit zu Rate gezogen. Hier sind vor allem die Bogengänge von Bedeutung. Deren Orientierung und Lage zueinander ermöglichen Aussagen über den Gleichgewichtssinn und somit auch die Rotationsfähigkeit des Kopfes. Darüber hinaus hat die Weite der Bogengänge Einfluss auf die Agilität eines Tieres. Die optimale Lage der drei Bogengänge zueinander liegt bei jeweils rund 90°. Bei den untersuchten Glyptodonten, hier die Gattungen Glyptodon, Doedicurus, Panochthus und Pseudoplohophorus, stehen die Bogengänge in einem spitzeren Winkel zueinander, was mit zahlreichen grabenden Tieren einschließlich der Gürteltiere übereinstimmt und eine eher geringe Beweglichkeit beziehungsweise Rotationsfähigkeit des Kopfes impliziert. Die Bogengänge selbst sind eng und vergleichsweise dick gegenüber denen der Gürteltiere und der Pampatheriidae. Generell wird daher die Agilität der Tiere als niedrig eingeschätzt. Jedoch bestehen bei den verschiedenen untersuchten Formen der Glyptodonten einzelne Unterschiede. So weist Glyptodon in Relation zu den anderen drei Gattungen einen kleineren seitlichen Bogengang auf. Dies könnte koordinative Auswirkungen beinhalten, da Glyptodon als einziger Angehöriger kein keulenartiges Schwanzende besaß, das von den drei anderen Vertretern eventuell als Verteidigungswaffe eingesetzt wurde.[31]
Direkte Belege für die Fortbewegung der Glyptodonten in Form von Spurenfossilien sind bisher nur wenig bekannt. Die umfangreichsten kamen in Pehuén-Có bei Bahía Blanca in der argentinischen Provinz Buenos Aires zu Tage. Diese 1986 entdeckte Fundstelle mit einem Alter von rund 12.000 Jahren birgt auf einer Fläche von rund 1,5 km² eine große Anzahl an Trittsiegeln unterschiedlichster Säugetiere und Vögel, die in ein ursprünglich weiches Substrat eingedrückt sind. Darunter befinden sich rundliche Abdrücke mit fünf beziehungsweise drei kurzen Zehen. In Form und Größe stimmen sie mit der anatomischen Rekonstruktion der Füße von Glyptodon überein, wobei die fünfstrahligen Abdrücke als Hinterfüße, die dreistrahligen als Vorderfüße interpretiert werden (die Vorderfüße von Glyptodon sind eigentlich vierstrahlig, der äußere Strahl V steht seitlich ab und ist klein, so dass er möglicherweise keinen Abdruck hinterließ). Die Maße der Hinterfußabrücke betragen 18,5 × 18,5 cm, die der Vorderfüße 17 × 10 cm. Die Funde werden der Spurengattung Glyptodontichnus zugewiesen. Die Seltenheit von Glyptodonten-Spurenfossilien im Gegensatz zu dem reichhaltigen anatomischen Fossilreport erklären einige Forscher damit, dass die großen Tiere Landschaften mit weichem Untergrund aufgrund größerer Verletzungsgefahr mieden.[32]
Ernährung
Aufgrund der zahnschmelzlosen Zähne sind keine Abriebspuren erkennbar, auch gibt es keine Überreste von Nahrung in Form von Koprolithen, um die Ernährungsweise genauer zu untersuchen. Die extrem hochkronigen Zähne lassen möglicherweise eine Ernährung hauptsächlich von Gras annehmen. Für stammesgeschichtlich frühe Vertreter wird aber auch gemischte Pflanzenkost als Hauptnahrung vermutet.[1] Dies lässt sich unter anderem aus der relativen Breite der Schnauze schlussfolgern. Heutige grasfressende Huftiere haben meist ein vergleichsweise weites Maul gegenüber den schmalschnauzigeren, stärker selektiven Blattfressern. Für zahlreiche frühe Glyptodonten aus dem Miozän wie Propalaehoplophorus oder Cochlops kann ein eher schmales Rostrum rekonstruiert werden, während dieses bei späten Formen aus dem Pleistozän wie Doedicurus oder Panochthus deutlich breiter war. Erstere beiden ernährten sich wohl demzufolge stärker wählerisch als letztere beiden. Ähnlich wie Propalaehoplophorus und Cochlops ist auch Glyptodon einzuschätzen, dessen Schnauze allgemein schmaler war als bei seinen pleistozänen Zeitgenossen.[33] In der Struktur des Gebisses ähneln Glyptodonten den Rindern und afrikanischen Nashörnern, ersteren fehlen Schneidezähne im Oberkiefer, letzteren zudem auch im Unterkiefer. Beide Gruppen benutzen ihre beweglichen Lippen zur Nahrungsaufnahme, was auch bei den Glyptodonten angenommen werden kann. Aufgrund der besonderen Gestaltung des Schädels mit der kurzen Schnauze hat sich bei den Glyptodonten ein abweichender Kauapparat entwickelt. Die Schädelkürzungen hatten zur Folge, dass der Hirnschädel über die hintere Hälfte der Zahnreihe ragt, eine Kondition, die bei den verwandten Gürteltieren nicht auftritt und zu einer abweichenden Anordnung der Kaumuskulatur führt.[7][8]
Im Vergleich zur Körpergröße der Tiere ist darüber hinaus die gesamte verfügbare Kauoberfläche des Gebisses eher klein. So besaß Plohophorus bei einem Körpergewicht von schätzungsweise 260 kg eine Kaufläche von knapp 900 mm², einem etwa gleich großen Steppenzebra stehen dagegen 2600 bis 2830 mm² zur Verfügung, beim Flachlandtapir bewegt sie sich zwischen 1930 und 2240 mm². Für Doedicurus lässt sich wiederum eine Kauoberfläche von etwa 1800 mm² bei einem Körpergewicht von rund 1,5 t ermitteln. Hier liegen Vergleichswerte beim etwa gleich großen Java-Nashorn bei rund 6250 mm². Aufgrund dieser Merkmale und in Verbindung mit der besonderen Gestaltung des Unterkiefers – so liegt der hinterste Zahn durch den schräg nach vorn ragenden Gelenkast direkt unterhalb des Unterkiefergelenkes und nicht wie sonst üblich deutlich davor – ergibt sich, dass Glyptodonten ihre Nahrung offensichtlich weniger effizient kauen konnten. Da dadurch die Nahrung in einem geringeren Maß zerkleinert wurde, durch die teils enorme Größe der Tiere aber entsprechend hohe Nahrungsmengen aufgenommen werden mussten, gehen Wissenschaftler von einer insgesamt niedrigen Stoffwechselrate für diese Tiere aus.[7][1][34][33] Allerdings zeigt das Zungenbein eine robuste Gestaltung, die eine sehr gut ausgeprägte und bewegliche Zunge annehmen lassen, welche möglicherweise die Nahrungsaufnahme und den Verdauungsprozess im Maul unterstützte.[35][36]
Sozialverhalten und Individualentwicklung
Über das Sozial- und Fortpflanzungsverhalten der Glyptodonten ist kaum etwas bekannt, heutige Gürteltiere sind einzelgängerisch und kommen nur zur Paarungszeit zusammen. Die Wurfgröße ist artspezifisch und variiert zwischen 1 und 3 Jungtieren bei den meisten Vertretern und bis zu 12 bei einigen Angehörigen der Langnasengürteltiere.[37] Jungtiere von Glyptodonten sind teils gut belegt. Mit diesen aufgefundene Osteoderme besitzen eine hohe Dicke und eine wenig ausgeprägte Oberflächenmusterung. Dies gibt an, dass der Panzer analog zu den heutigen Gürteltieren erst nach der Geburt vollständig ausformte und aushärtete.[38][39][40] Als äußerst selten sind Funde von ungeborenen Individuen im Rückenpanzer von Alttieren anzusehen. Ein Beispiel wurde in Monte Cercado im südlichen Bolivien entdeckt und gehört zu Glyptodon, ein weiteres liegt aus der Sopas-Formation in Uruguay vor und repräsentiert Neuryurus. Am Unterkiefer des erstgenannten Fundes waren bereits einzelne Zähne durchgebrochen, ob Glyptodonten aber voll bezahnt zur Welt kamen, ist unklar.[41][42] Schädeluntersuchungen zeigten wiederum auf, dass die Zähne von hinten nach vorn durchbrechen, die ausgesprochene Hochkronigkeit jedoch schon bei jungen Individuen bestand.[43]
Funktion des Panzers und Verteidigung
Es gilt als wahrscheinlich, dass sich der Panzer ursprünglich nicht aufgrund der Bedrohung durch Fressfeinde entwickelte, sondern möglicherweise als Schutz vor stacheliger Vegetation fungierte. Erst in zweiter Linie schützte er vor größeren Beutegreifern, die damals in Südamerika von den Phorusrhacidae („Terrorvögel“) und Raubbeutlern wie etwa die Vertreter der Borhyaenidae und der Thylacosmilidae gestellt wurden.[12] Als sekundäre Funktion diente er wohl auch als Abschluss eines Fettspeichers oberhalb des Brustbereiches, ähnlich den Höckern der Kamele. Hier ist der Panzer nicht mit der Wirbelsäule oder den Rippen verbunden und erhebt sich frei darüber, wodurch unter anderem bei Doedicurus ein Freiraum mit einer Höhe von rund 25 cm bestand. Da dieser Bereich kaum von Muskeln beansprucht worden sein dürfte, gehen Fachleute hier von einem Fettpolster aus.[30]

Der lange und massive Schwanz diente wahrscheinlich auch als Gegengewicht bei der Fortbewegung, was durch die starre Wirbelsäule und das fest mit ihr verwachsene Becken notwendig wurde. Allerdings konnten ihn einige Vertreter, bei denen das Ende keulenartig verwachsen war, auch als Waffe einsetzen. Am Schwanzansatz war eine kräftige Muskulatur ausgebildet, was die massiven Fortsätze der Wirbel und der große Durchmesser der Panzerringe anzeigen. Diese allein wog Berechnungen auf Basis der Schwanzgröße bei Panochthus rund 74 kg, bei Doedicurus möglicherweise bis zu 108 kg. Das keulenartige Ende ist bei beiden 89 respektive 105 cm lang, bei einem Gewicht von 30 und 65 kg. Die vorhandene Muskelmasse ermöglichte es einzelnen Berechnungen zufolge, über das keulenartige Ende bis zu 3000 J an Energie punktuell bei einem Schlag freizusetzen (das entspricht in etwa der Menge, die ein Kugelstoßer benötigt, um die 7,3 kg schwere Kugel 16 m weit zu stoßen), wobei der Schlagpunkt im hinteren Teil der Schwanzkeule lag. Auf derartige Einsätze des Schwanzes im intraspezifischen Kampf deuten auch einige Panzer hin, die Frakturen aufweisen, so unter anderem ein Rückenpanzer von Doedicurus mit verheilten Narben von etwa 35 cm Durchmesser.[30][22] Offensichtlich bestanden hierbei aber auch funktionelle Unterschiede. So weisen unter anderem frühe Vertreter von Panochthus ein eher konisches Schwanzende auf, während dieses bei späteren Formen an ein Wikingerschwert erinnert. Erstere konnten ihren Schwanz in alle Richtungen kreisen, ohne dass bei den Schlägen eine hohe Präzision erreicht wurde. Für letztere ließen sich überwiegend horizontale Bewegungen rekonstruieren, die wiederum zielgerichteter erfolgten.[44] Bei einigen Formen, die wie Glyptodon nicht über ein keulenartiges Schwanzende verfügten, befanden sich am Rand des Körperpanzers bis zu drei Reihen von hochmodifizierten Knochenplättchen, die ihn umschlossen, aber nicht mit diesem fest verwachsen waren und eine spitz-konische Form aufwiesen. Möglicherweise besaßen diese Osteoderme eine Schutzfunktion für besonders wichtige Körperteile wie den Nacken.[21] Der älteste Nachweis vollständig ausgebildeter Schwanzkeulen wurde bisher bei Kelenkura aus dem Oberen Miozän vor rund 9 Millionen Jahren erbracht.[45]
Ob die Glyptodonten ihren Schwanz auch gegen Fressfeinde einsetzten, ist unklar, heutige Stirnwaffenträger als Vergleich benutzen ihre Hörner und Geweihe nur äußerst selten zur Abwehr von Beutegreifern. Zumindest für die größten Vertreter wird eine derartige Bedrohung im ausgewachsenen Alter eher ausgeschlossen.[30] Aus Nordamerika ist lediglich der Schädel eines jungen Individuums von Glyptotherium aus dem Pliozän bekannt, dessen Kopfpanzer noch nicht vollständig ausgebildet war und der Bissmarken aufweist, die zum Tod des Tieres führten.[12] Aus der argentinischen Provinz Buenos Aires wiederum ist ein Panzer mit Resten des Körperskelettes von Eosclerocalyptus aufgefunden worden, der ebenfalls ins Pliozän datiert und dessen Wirbel wiederum Bisspuren tragen. Diese stammen wahrscheinlich von einem Kleinbären, möglicherweise von Chapalmalania, der zu jener Zeit in der Region fossil belegt ist. Allerdings entstanden sie wohl erst nach dem Tod des Glyptodonten und gehen somit auf Aasfresserei zurück.[46]
Gehirn
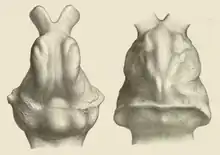
Mehrere vollständige Schädel ermöglichen die Untersuchung der Gehirnstruktur. Ausgüsse der Gehirnkapsel ergeben für große Vertreter wie Glyptodon, Doedicurus und Panochthus mit Körpergewichten von 1,2 bis 2 t ein Volumen von 213 bis 234 cm³, bei kleineren wie Pseudohoplophorus, das nur etwas mehr als 200 kg wog, von 101 cm³. Der Enzephalisationsquotient liegt bei 0,12 bis 0,4, wobei das kleinere Pseudohoplophorus den größten Wert aufweist. Die Angaben liegen im unteren Bereich der heutigen Gürteltiere (0,44 bis 1,06) und entsprechen zusätzlich denen der Pampatherien. Das Gehirn der Glyptodonten verfügte über einen ausgedehnten Riechkolben, der zwischen 4,8 und 9,7 % des Gesamtgehirns in Anspruch nahm. Rund zwei Drittel wurden darüber hinaus vom Großhirn und der Rest vom Kleinhirn eingenommen. Im generellen Aufbau stimmt dies mit den Gürteltieren überein, allerdings ist bei letzteren das Großhirn im Verhältnis voluminöser und das Kleinhirn weniger umfangreich. Abweichend von den Gürteltieren mit ihrem breiten Riechkolben war dieser bei den Glyptodonten lang-schmal und spitz, ähnlich wie es sich auch bei den Pampatherien wiederfindet. Korrespondierend zu den Gürteltieren zeigte sich das Relief der Großhirnrinde als relativ einfach gestaltet. An Furchen ließ sich lediglich der Sulcus suprasylvianus des Scheitellappens ausmachen, was auch für die Pampatherien zutrifft. Der bei den Gürteltieren zusätzlich ausgebildete Sulcus praesylvianus fehlte dagegen.[47][48]
Generell haben die heutigen Gürteltiere in Relation kleinere Gehirne als die Ameisenbären und Faultiere. Die Gründe hierfür sind nicht eindeutig. Sie könnten einerseits in einer kürzeren Aufzuchtphase des Nachwuchses liegen, andererseits auch mit der Ausbildung des Panzers und den damit verbundenen zahlreichen biologischen und funktionalen Einschränkungen zusammenhängen. Auch der extrem niedrige Stoffwechsel der Gürteltiere wäre eine potentielle Möglichkeit, da dadurch weniger Energie in die aufwendige Entwicklung des Gehirns fließt. Für die Glyptodonten wird ebenfalls ein geringer Metabolismus angenommen. Ihre gegenüber den Gürteltieren weitaus größere Statur lässt noch weitere Überlegungen zu dem kleinen Gehirn zu. Die teils enorme Körpergröße und der massive Panzer erforderten kaum Verteidigungs- und Fluchtstrategien gegenüber großen Beutegreifern, was wiederum gegen ein größeres Gehirn spricht. Ähnliches ist von den gepanzerten Ankylosauriern mit einem ebenfalls kleinen Enzephalisationsquotienten im Verhältnis zu ungepanzerten Sauriern überliefert. Als eine einschränkende funktionale Komponente kommt der Panzer aber selbst in Betracht. Dieser ermöglichte bedingt durch seine Kompaktheit nur eine schwach ausgebildete Halsmuskulatur, die den Kopf trägt und stabilisiert. Eine verminderte Gehirngröße unterstützte damit die Gewichtsverringerung des Schädels, was sich dann vor allem bei den riesigen Formen des Pleistozäns mit ihren großen Schädeln stärker auswirkte.[47][48]
Parasiten und Pathologien

An einigen Rücken- und Schwanzpanzern, unter anderem von Glyptodon und Panochthus, treten mitunter rundliche Durchbohrungen von nur wenigen Millimetern Durchmesser und mit konischem Querschnitt auf. Sie ähneln parasitischen Fraßspuren, die von Flöhen, speziell der Gattung Tunga, verursacht werden. Vergleichbare Befunde wurden auch bei fossilen Gürteltieren berichtet[49] und sind ebenso von den heutigen Vertretern der Gruppe bekannt. Da sich derartige Eintiefungen sowohl bei den Gürteltieren als auch den Glyptodonten belegen lassen und bereits an Fossilien aus dem Miozän dokumentiert sind, kann von einer langen Koevolution zwischen den gepanzerten Tieren und den Flöhen ausgegangen werden. Des Weiteren kommen oberflächliche Veränderungen an den Ornamenten der Knochenplättchen vor. Diese gehen womöglich auf Entzündungen zurück, deren Verursacher Bakterien oder Pilze waren.[50]
Eher selten lassen sich bisher pathologische Veränderungen nachweisen. Dazu gehören unter anderem Knochendeformationen an den Füßen, besonders im Bereich der Gelenke. Sie sind in der Regel auf Enthesiopathien und Pseudogicht zurückzuführen. Einzelne Individuen zeigen dabei unter Umständen mehrere Krankheitsbilder gleichzeitig auf, was womöglich auf die Anfälligkeit vor allem der großen Glyptodonten für solche Gelenkerkrankungen verweist.[51][52][53]
Systematik
Äußere Systematik
Verwandtschaftsverhältnis der Glyptodonten und Gürteltiere laut morphologischen Daten nach Billet et al. 2011[54]
|
Die Glyptodontidae bilden eine Familie innerhalb der Ordnung der Gepanzerten Nebengelenktiere (Cingulata). Diese Ordnung war einst formenreich in Süd- und Nordamerika vertreten. Ihr gehören weiterhin auch die ebenfalls ausgestorbenen Pampatheriidae, die Peltephilidae, die Palaeopeltidae, die Pachyarmatheriidae sowie die heute noch bestehenden Gürteltiere (Dasypoda) an. Die Gepanzerten Nebengelenktiere stellen wiederum einen Teil der Überordnung der Nebengelenktiere (Xenarthra) dar, welche zusätzlich noch die Zahnarmen (Pilosa) mit den heutigen Faultieren (Folivora) und den Ameisenbären (Vermilingua) beinhalten. Die Nebengelenktiere repräsentieren eine der vier Hauptlinien der Höheren Säugetiere, die den anderen drei (zusammengefasst als Epitheria) als Schwestergruppe gegenübersteht.[55] Eine generelle Gemeinsamkeit der Nebengelenktiere findet sich in den namengebenden xenarthrischen Gelenke (Nebengelenke, auch Xenarthrale) an den Gelenkfortsätzen der hinteren Brust- und der Lendenwirbel. Hier bilden allerdings die Glyptodontidae eine Ausnahme, da deren Lendenwirbel als einzigartiges Merkmal miteinander verwachsen sind.[56] Der Ursprung ist bisher unbekannt, die ältesten Fossilfunde stammen aus Südamerika und datieren in das Paläozän vor mehr als 56 Millionen Jahren, die zu den Gürteltieren gerechnet werden.[57] Molekulargenetische Untersuchungen erbrachten eine Abspaltung der Nebengelenktiere von den anderen Höheren Säugetieren bereits in der ausgehenden Unterkreide vor etwa 103 Millionen Jahren. Die Gürteltiere als heute lebende, nächste Verwandte der Glyptodonten trennten sich von der gemeinsamen Linie mit den Zahnarmen zu Beginn des Paläozän vor etwa 65 Millionen Jahren ab.[58][59]

Die Innere Gliederung der Gepanzerten Nebengelenktiere ist nicht vollständig gesichert und gegenwärtig im Fluss. Von größere Bedeutung sind hier die Pampatheriidae, die in einer klassischen Auffassung als die nächsten Verwandten der Glyptodonten gelten. Diese charakterisieren sich durch einen Rückenpanzer, der ähnlich den Gürteltieren mit beweglichen Bändern zwischen einem starren Schulter- und Beckenschild versehenen ist. Abweichend von den Gürteltieren mit ihrer variablen Anzahl an beweglichen Bändern waren es bei den Pampatherien zumeist drei. Mit einem Gewicht von über 200 kg für einige Formen des späten Pleistozäns, etwa Pampatherium und Holmesina, erreichten sie auch deutlich größere Ausmaße als die Gürteltiere, allerdings zeichnet sie die Gestaltung der Vorderbeine als nicht so geschickte Gräber aus.[60] Aufgrund des markanten Baus des Panzers hielt man die Pampatherien ursprünglich für näher verwandt mit den Gürteltieren und führte sie innerhalb dieser als Unterfamilie. Untersuchungen an Schädeln und Zähnen erwiesen dagegen, dass sich Pampatherien und Glyptodonten näher stehen. Dies ergab sich unter anderem aus der Struktur des Gehörganges und dem Bau des Kauapparates, wie der hohe Unterkiefer, aber auch aufgrund der komplexer gestalteten Zähne. Letztere sind bei den Gürteltieren einfach nagelartig gebaut, bei den Pampatherien aber durch zwei querstehende Loben, bei den Glyptodonten durch drei charakterisiert. Beide Familien bilden zusammen die übergeordnete Gruppe der Glyptodonta.[61][62] Die weiteren Gruppen – Palaeopeltidae, Peltephilidae und Pachyarmatheriidae – stehen in einem mehr oder weniger engen Verhältnis zu den übrigen Gepanzerten Nebengelenktieren, wobei letztere möglicherweise die Schwestergruppe der Glyptodonta repräsentieren.[63]
Verwandtschaftsverhältnis der Glyptodonten und Gürteltiere laut molekulargenetischen Daten nach Delsuc et al. 2016[64]
|
Die Beziehungen der Glyptodonten zu den heutigen Gürteltieren unterliegen einer bis heute anhaltenden fachwissenschaftlichen Debatte. Die anfänglich favorisierte Zweiteilung der Gepanzerten Nebengelenktiere in die Großgruppen der Gürteltier- und Glyptodonten-Verwandtschaft wurde durch modernere phylogenetische Untersuchungen basierend auf anatomischen Merkmalen im Jahr 2006 weitgehend aufgehoben. Die Analyse ergab eine deutlich nähere Bindung der Glyptodonta (Glyptodonten und Pampatherien) an die Gürteltiere und ordnete sie in eine evolutionäre Entwicklungslinie. Im Ergebnis bildeten die Glyptodonta eine gemeinsame Klade mit den Euphractinae, denen unter anderem das heutige Sechsbinden-Gürteltier angehört. Verfeinerte Studien in Verbindung mit weiterem Fossilmaterial im Jahr 2011 rückten die Glyptodonta noch tiefer in die Gürteltiere hinein, da sie eine Nahbeziehung zu den Eutatini vermuten ließen, letztere, benannt nach der Charakterform Eutatus, stellen einen ausgestorbenen Seitenzweig der euphractinen Gürteltiere dar. Dadurch wurden aber die Gürteltiere an sich zu einer paraphyletischen Gruppe.[61][54] Den skelettanatomischen Analysen stehen molekulargenetische Untersuchungen aus dem Jahr 2016 gegenüber, die neben den Gürteltieren auch die ausgestorbene Glyptodonten-Gattung Doedicurus einschlossen. Sie bestätigen generell die sehr nahe Verwandtschaft der beiden Gruppen. Im Gegensatz zu der skelettanatomisch festgestellten Beziehung zu den Euphractinae unterstützen die genetischen Daten jedoch ein Nahverhältnis der Glyptodonten zu einer Klade bestehend aus den Chlamyphorinae, welche die Gürtelmulle stellen, und den Tolypeutinae, innerhalb derer die Kugelgürteltiere, die Nacktschwanzgürteltiere sowie das Riesengürteltier vereint sind. Demnach wären die Glyptodonten die Schwestergruppe dieser beiden Unterfamilien, die Trennung der beiden Linien erfolgte im Übergang vom Eozän zum Oligozän vor etwa 35 Millionen Jahren. Sowohl nach den anatomischen wie auch den molekulargenetischen Untersuchungsergebnissen wären die Glyptodonten somit lediglich als Seitenzweig der Gürteltiere anzusehen und würden keine eigenständige Entwicklungslinie innerhalb der Gepanzerten Nebengelenktiere darstellen. Zum genauen taxonomischen Status der Glyptodonten finden sich unterschiedliche Auffassungen. Einige Forscher sehen die Glyptodonten lediglich auf der Ebene einer Unterfamilie (Glyptodontinae) innerhalb der Gürteltiere, andere behalten den Familienstatus vorerst bei. Die Position der Pampatherien ist unklar, da bisher noch keine DNA-Untersuchungen dazu vorliegen.[64][3] Allerdings unterstützen anatomische Untersuchungen am Innenohr sowohl die nahe Position der Glyptodonten zu den Chlamyphorinae als auch ihre enge Verwandtschaft mit den Pampatherien.[31]
Innere Systematik
Innere Systematik der Glyptodonten nach Croft et al. 2007[65]
|
In einer häufig zitierten Ansicht wird die Familie der Glyptodonten in fünf Unterfamilien unterteilt. Sehr urtümlich erscheinen die Glyptatelinae als basale Gruppe. Sie waren auf das Eozän und das Oligozän beschränkt und wiesen noch sehr ursprüngliche Merkmale der Osteoderme und Zähne auf. So sind erstere nicht so symmetrisch-rosettenartig aufgebaut wie bei stammesgeschichtlich jüngeren Glyptodonten, sondern weisen ein dezentral liegendes Hauptmuster auf. Insgesamt gelten sie aber als nur wenig erforscht.[66] Die Propalaehoplophorinae traten erstmals im Oberen Oligozän auf, verschwanden aber im Oberen Miozän wieder. Sie waren deutlich weiter entwickelt als die Glyptatelinae, aber weniger modern als die späteren Glyptodonten. Gegenüber diesen zeichnen sie sich durch etwas längere Schädel mit einer weiter ausgedehnten Schnauze aus, ebenso wie durch das Vorhandensein von bis zu sechs beweglichen Bändern an den Seitenrändern des Rückenpanzers, was als ursprünglicher Zustand innerhalb der Glyptodonten gilt. Allerdings weisen die Osteoderme bereits eine symmetrisch Form und meist eine zentrale Musterung auf.[65][6] In ihrer Körpergröße blieben sie mit etwa 70 bis 115 kg noch deutlich hinter den späteren Formen zurück.[1]
Die anderen drei Unterfamilien umfassen die „Hoplophorinae“ (= Sclerocalyptinae), die Doedicurinae und die Glyptodontinae. Die „Hoplophorinae“ stellen die formenreichste Gruppe mit circa 60 % aller bekannten Taxa dar. Die Unterfamilie ist insgesamt über einige, nur unzureichend definierte Merkmale charakterisiert, daher sehen einige Untersuchungen sie nicht als monophyletische Gruppe an. Sie wird daher teilweise als Sammelgruppe für Vertreter aufgefasst, die moderner erscheinen als die Propalaehoplophorinae, aber außerhalb der Glyptodontinae und Doedicurinae stehen.[62][17] Die große Heterogenität der Unterfamilie schlägt sich auch in der Aufstellung von wenigstens sechs weiteren Untergruppierungen nieder (etwa die Hoplophorini, Panochthini, Plohophorini), die in sich genommen teilweise wieder stark angezweifelt werden. Mitunter als Resultat daraus lösten einzelne Fachwissenschaftler verschiedentlich weitere Unterfamilien aus den „Hoplophorinae“ heraus.[67][68][69] Zu den bedeutendsten Entwicklungen innerhalb der „Hoplophorinae“ gehört die Ausbildung eines keulenartigen Schwanzendes.[67] Die Doedicurinae und Glyptodontinae umfassen stammesgeschichtlich jüngere Glyptodonten, die moderner waren als die Propalaehoplophorinae. Dabei charakterisieren die Doedicurinae eine ausgeprägte Schwanzkeule und Knochenplättchen, die kaum ornamentiert sind. Die Gruppe ist aber bis auf den namengebenden Vertreter weitgehend wenig erforscht und fast nur anhand der Panzerung bekannt. Die Glyptodontinae wiederum behielten die Gestaltung der Osteoderme der Propalaehoplophorinae weitgehend bei, entwickelten aber kurze Schwänze ohne keulenartig verwachsenes Ende.[70]
|
Ein von dieser traditionellen Gliederung abweichendes Ordnungsschema der Glyptodonten ergaben kladistische Analysen des Jahres 2008. Die gesamte Gruppe der Glyptodonten in der klassischen Auffassung wurde auf die Ebene der Unterordnung der Glyptodontia angehoben. Die eigentlichen Glyptodontidae beschränken sich auf die stammesgeschichtlich moderneren Formen. Innerhalb der Glyptodontidae wurden die Doedicurinae mit den Glyptodontinae vereint, eine Aufteilung erfolgt hier nur noch auf der Ebene der Tribus. Zur Seite der Glyptodontinae stehen die Plohophorinae und einzelne weitere Triben. Nicht zu den Glyptodontidae gehörig, sondern als eigenständig erwiesen sich die Panochthidae, die in die beiden Triben der Panochthini und der Neosclerocalyptini differenziert werden. Die Glyptodontidae und die Panochthidae wiederum formen die Überfamilie der Glyptodontoidea. Die häufig als paraphyletisch angenommenen Propalaehoplophorinae stellten sich als monophyletisch heraus, formen aber nun als eigenständige Familie eine Linie außerhalb der Glyptodontoidea. Hier erhielten zudem die Glyptatelinae einen Familienstatus. Die problematischen „Hoplophorinae“ wurden dagegen aufgelöst, die meisten der enthaltenen Gattungen sind aber in diesem Gliederungsschema nicht einer genaueren Gruppe zuordenbar, sie gelten somit als incertae sedis. In dieser Sichtweise bestehen die Glyptodontia derzeit aus vier Familien: den Glyptatelidae, den Propalaehoplophoridae, den Panochthidae und den Glyptodontidae.[71][17]
In weiteren phylogenetischen Studien wird teilweise zwischen einer „nördlichen“ und „südlichen Klade“ unterschieden.[19][72][45] Erstere umfasst weitgehend die klassischen Glyptodontinae, letztere die Propalaeohoplophorinae, die „Hoplophorinae“ und die Doedicurinae. Beide Linien bilden in sich geschlossene Gruppen, wobei in der „südlichen Klade“ die angenommenen höheren taxonomischen Einheiten sich stärker mischen und so als eher paraphyletisch aufzufassen sind. Definierende Unterschiede zwischen den beiden Gruppen finden sich unter anderem in der Panzergestaltung. So kam es in der „nördlichen Klade“ nicht zu einer Ausbildung einer komplexen Schwanzpanzerung, während sich in der „südlichen“ eine teils massive Schwanzkeule herausformte.[45]
Überblick über die Gattungen
Insgesamt sind etwa 65 Gattungen der Glyptodontidae bekannt, womit die Familie sehr formenreich war. Die meisten der Taxa sind aber nur unzureichend überliefert, ihre Beschreibung basiert in der Regel auf Resten der Panzerung. Nur bei den wenigsten Formen sind auch Schädel oder Körperskelettteile bekannt. Traditionell erfolgt die Unterscheidung der einzelnen Arten und Gattungen der Glyptodonten anhand der Form der Knochenplättchen des Panzers und der Gestaltung der Schwanzpanzerung und nicht, wie sonst bei Säugetieren üblich, mit Hilfe von Schädel- und Gebissmerkmalen.[62][70] Aus diesem Grund ist es teilweise auch schwierig, isoliertes Skelettmaterial bestimmten Taxa zuzuweisen, die sonst nur durch Panzerreste bekannt sind.[73][74] Das hier verwendete Ordnungssystem folgt weitgehend McKenna und Bell 1997,[75] wobei neuere Entwicklungen berücksichtigt wurden.[76][77][65][6][78][79][39][67][80][81][45]
- Familie: Glyptodontidae Gray, 1869
- Paraeucinepeltus González-Ruiz, Zurita, Scillato-Yané, Zamorano & Tejedor, 2011
- Parapropalaehoplophorus Croft, Flynn & Wyss, 2007
- Kelenkura Barasoain, Zurita, Croft, Montalvo, Contreras, Miño‑Boilini & Tomassini, 2022
- Unterfamilie: Glyptatelinae Castellanos, 1932
- Glyptatelus Ameghino, 1897
- Clypeotherium Scillato-Yané, 1977
- Unterfamilie: Propalaeohoplophorinae Castellanos, 1932
- Propalaehoplophorus (= Propalaeohoplophorus) Ameghino, 1887
- Cochlops (= Metopotoxus) Ameghino, 1889
- Eucinepeltus Ameghino, 1891
- Asterostemma Ameghino, 1889
- Unterfamilie: „Hoplophorinae“ Huxley, 1864
- Asymmetrura Fariña, 1981
- Caudaphorus Fariña, 1981
- Uruguayurus Mones, 1987
- Tribus: Hoplophorini Huxley, 1864
- Hoplophorus (= Sclerocalyptus) Lund, 1838
- Eosclerocalyptus Ameghino, 1919
- Hoplophractus Cabrera, 1939
- Trachycalyptus Ameghino, 1908
- Berthawyleria Castellanos, 1939
- Parahoplophorus Castellanos, 1932
- Isolinia Castellanos, 1951
- Stromaphoropsis Kraglievich, 1932
- Eosclerophorus Castellanos, 1948
- Trabalia Kraglievich, 1932
- Neosclerocalyptus Paula Couto, 1957
- Eonaucum Scillato-Yané & Carlini, 1998
- Chacus Zurita, 2002
- Tribus: Palaehoplophorini Hoffstetter, 1958
- Palaehoplophorus (= Palaeohoplophorus) Ameghino, 1883
- Aspidocalyptus Cabrera, 1939
- Chlamyphractus Castellanos, 1939
- Pseudoeuryurus Ameghino, 1889
- Protoglyptodon Ameghino, 1885
- Palaehoplophoroides Scillato-Yané & Carlini, 1998
- Tribus: Lomaphorini Hoffstetter, 1958
- Peiranoa Castellanos, 1946
- Lomaphorops Castellanos, 1932
- Lomaphorus Ameghino, 1889
- Trachycalyptoides Saint-André, 1996
- Tribus: Plohophorini Castellanos, 1932
- Coscinocercus Cabrera, 1939
- Phlyctaenopyga Cabrera, 1944
- Stromaphorus Castellanos, 1926
- Plohophorops Rusconi, 1933
- Plohophorus (= Urotherium) Ameghino, 1887
- Nopachtus Ameghino, 1888
- Propanochthus Castellanos, 1925
- Pseudoplohophorus Castellanos, 1926
- Tesseiria Kraglievich, 1932
- Plohophoroides Castellanos, 1928
- Zaphilus Ameghino, 1889
- Tribus Panochthini Castellanos, 1927
- Panochthus Burmeister, 1866
- Parapanochthus Moreira, 1971
- Tribus: Neuryurini Hoffstetter, 1958
- Neuryurus (= Euryurus) Ameghino, 1889
- Unterfamilie: Doedicurinae Ameghino, 1889
- Eleutherocercus Koken, 1888
- Prodaedicurus (= Palaeodoedicurus, Thoracophorus) Castellanos, 1927
- Comaphorus Ameghino, 1886
- Castellanosia Kraglievich, 1932
- Xiphuroides Castellanos, 1927
- Doedicurus Burmeister, 1874
- Doedicuroides Castellanos, 1941
- Plaxhaplous Ameghino, 1884
- Unterfamilie: Glyptodontinae Gray, 1869
- Boreostemma Carlini, Zurita, Scillato-Yané, Sánchez & Aguilera, 2008
- Tribus: Glyptodontini Gray, 1869
- Glyptodontidium Cabrera, 1944
- Glyptodon (= Chlamydotherium, Glyptocoileus, Glyptopedium, Lepitherium, Neothoracophorus?, Pachypus, Paraglyptodon, Pseudothoracophorus, Schistopleurum) Owen, 1839
- Stromatherium Castellanos, 1953
- Glyptostracon Castellanos, 1938
- Heteroglyptodon Roselli, 1976
- Tribus: Glyptotheriini Castellanos, 1953
- Glyptotherium (= Brachyostracon, Boreostracon, Neothoracophorus?, Xenoglyptodon) Osborn, 1903
Stammesgeschichte
Adaptive Radiation und Ursprünge
Zahlreiche Linien der Glyptodonten erfuhren eine starke Körpergrößenzunahme, vor allem im letzten Abschnitt des Pleistozäns. Erklärt wird diese teilweise mit der Bergmannschen Regel, wodurch die Tiere unter den kühleren Bedingungen der Eiszeit größer wurden. Einen stärkeren Einfluss hierbei haben allerdings auch die verfügbaren Nahrungsressourcen, so dass Formen aus höheren Bergregionen mit ihren begrenzteren Pflanzenangebot dieser Regel nicht zwingend unterlagen.[82] Andere Erklärungsversuche führen den Konkurrenzdruck an, der durch einwandernde Pflanzenfresser aus Nordamerika im Zuge des Großen Amerikanischen Faunenaustausches seit dem Pliozän entstand.[2] Weitere evolutive Veränderungen betreffen die Reduktion der Zehenanzahl in mehreren Linien und die Überprägung der Form der Knochenplättchen und des Schwanzpanzers. Dabei zeigt sich aber, dass ähnlich gestaltete Strukturen nicht unbedingt auf nähere Verwandtschaft schließen lassen, sondern teilweise Ausdruck einer konvergenten Entwicklung sind.[70]
Die frühesten bekannten Funde, einige Knochenplättchen des Rückenpanzers, stammen aus Patagonien und gehören mit einem Alter von 48 bis 42 Millionen Jahren ins Mittlere Eozän. Sie werden der Gattung Glyptatelus aus der Gruppe der Glyptatelinae zugewiesen. Das Material wurde bereits Ende des 19. Jahrhunderts vorgestellt und soll aus entsprechend alten Gesteinseinheiten geborgen worden sein,[83][84] genauere Daten zu den Lokalitäten liegen aber nicht vor.[85][86] Damit ist der älteste Nachweis der Glyptodonten etwas jünger als der der verwandten Gürteltiere, welcher in Ablagerungen des Paläozän mit Alterswerten von vor über 56 Millionen Jahren im südlichen Brasilien gefunden wurde.[87] Möglicherweise ist im südlicheren Teil des Kontinentes auch die Ursprungsregion beider Gruppen zu suchen. Im Oberen Eozän ist dann Clypeotherium aus der gleichen Verwandtschaftsgruppe nachgewiesen.[88]
Oligozän
Im Verlauf des Oligozän treten weiterhin Clypeotherium und Glyptatelus auf, so unter anderem mit einzelnen Knochenplättchen in Quebrada Fiera im westlichen Argentinien belegt.[89] Im ausgehenden Oligozän, einer Zeit mit stärkerer Diversifizierung der Glyptodonten, sind erstmals Vertreter der Propalaehoplophorinae überliefert, eine der am besten untersuchten basalen Gruppen dieser Gürteltierverwandten. Sie gelten zudem als Ausgangsgruppe der Entstehung der moderneren Glyptodonten.[88][6] Ähnlich den Glyptatelinae liegen die frühesten Nachweise der Propalaehoplophorinae aus Patagonien vor, bedeutend ist hier etwa die Fundstelle El Pajarito in der argentinischen Provinz Chubut. Doch bereits im Oberen Oligozän hatten sie auch weiter nördlich gelegene Regionen erreicht, wie einige Osteoderme aus der Fray-Bentos-Formation in der argentinischen Provinz Entre Ríos zeigen.[90] Neben diesen beiden bedeutenden Linien kommt mit Pseudoglyptodon eine Gattung vor, die Mischmerkmale der Faultiere und der Glyptodonten aufweist. So erinnert der Gebissaufbau mit fünf Zähnen je Oberkiefer- und vier je Unterkieferhälfte an die Faultiere, die Gestaltung der Zahnkauflächen mit drei querstehenden Loben dagegen an die Glyptodonten. Abweichend von den Glyptodonten fehlten den Zähnen die erhöhten Rippeln aus hartem Zahnbein innerhalb der Loben. Bedeutende Funde stammen unter anderem aus Salla-Luribay in Bolivien und aus der Tinguiririca-Fauna aus dem zentralen Chile. Möglicherweise aber steht Pseudoglyptodon den Faultieren näher.[86]
Miozän

Recht formenreich traten die Glyptodonten im Miozän auf. Im Unteren Miozän und im Übergang zum Mittleren Miozän aus der Zeit vor 18 bis 16 Millionen Jahren ist die Santa-Cruz-Formation in Patagonien bedeutend, wo die Glyptodonten mit den Propalaehoplophorinae recht zahlreich in Erscheinung treten. Nachgewiesen sind mit Propalaehoplophorus, Cochlops, Asterostemma und Eucinepeltus wenigstens vier Gattungen, wobei letztere durch ebenfalls wenigstens vier Arten repräsentiert wird. Alle Vertreter waren deutlich größer als die heutigen Gürteltiere und wogen zwischen 67 kg bei Propalaehoplophorus und 115 kg bei Eucinepeltus. Erstmals für Glyptodonten sind aus der Santa-Cruz-Formation auch vollständige Skelette erhalten. Dadurch gelten die Propalaehoplophorinae als gut untersucht. Die Angehörigen der Propalaehoplophorinae ernährten sich hauptsächlich von gemischter Pflanzenkost und bewegten sich ausschließlich am Boden fort.[91][1] Neben den Funden der Santa-Cruz-Formation sind auch noch weitere bedeutende aus anderen Bereichen Südamerikas bekannt. Hierzu gehören Knochenplättchen und umfangreiche Reste des Körperskeletts von Propalaehoplophorus aus der zeitlich etwas jüngeren Río-Mayo-Formation in der Chubut-Provinz von Argentinien.[92] Ein Teilskelett von Parapropalaehoplophorus wurde aus der Chucal Formation im nördlichen Chile berichtet. Der Fund ist etwa gleichalt zu jenen der Santa-Cruz-Formation, aufgrund abweichender Merkmale der Zähne und der Osteoderme, letztere zeigen kaum Ornamentierungen, ist dessen genaue systematische Zuweisung unklar.[65]
Im Mittleren Miozän treten noch einzelne frühe Vertreter auf, etwa das mit beweglichen Bändern an den seitlichen Panzerrändern ausgestattete Paraeucinepeltus aus dem südlichen Argentinien. Darüber hinaus sind aber auch einige modernere Formen nachgewiesen, etwa Eonaucum.[79] Die frühesten Vertreter der Glyptodontinae erscheinen erstmals im Mittleren Miozän vor rund 12 Millionen Jahren und gehören der Gattung Boreostemma an. Aus dieser Zeit liegt ein vollständiges, rund 1,4 m langes Exemplar von Boreostemma aus den Monkey Beds der Villavieja-Formation in La Venta in Kolumbien vor.[93] Da auch weitere frühe Funde aus dem nördlichen Südamerika stammen, etwa aus der formenreichen Fitzcarrald-Lokalfauna des westlichen Amazonasgebietes in Peru,[94] kann ein Ursprung der Gruppe in dieser Region angenommen werden. Vom Norden aus breiteten sich die modernen Glyptodonten nach Süden aus, so dass sie im ausgehenden Miozän auch in der Pamparegion mit Glyptodontidium und möglicherweise schon Glyptodon nachweisbar sind.[95][96][80] Weiter südlich im heutigen zentralen Argentinien ist mit Kelenkura aus der Arroyo-Chasicó-Formation und der Loma-de-Las-Tapias-Formation des Oberen Miozäns eine Form dokumentiert, die erstmals über eine vollständige gepanzerte Schwanzkeule verfügte.[45] In regionaler und zeitlicher Nähe liegt eine sehr umfangreiche Glyptodontenfauna aus dem Conglomerado osífero der Ituzaingó-Formation vor, welche im nordöstlichen Argentinien am Unterlauf des Río Paraná nahe der Stadt Paraná aufgeschlossen ist und rund ein Dutzend Gattungen einschließt.[97] Wiederum in den Übergang om Miozän zum Pliozän lässt sich im südlichen Südamerika mit Eleutherocercus eines der frühesten Mitglieder der Doedicurinae feststellen, wie einzelne Skelettfunde einschließlich Schädel, Gliedmaßen und Panzerreste aus der Andalhuala-Formation im nordwestlichen Argentinien nahelegen.[98]
Plio- und Pleistozän

Vor allem im Pliozän und im Pleistozän kam es zu einer starken Aufsplitterung der Glyptodonten, was möglicherweise mit der Ausbreitung offener Landschaften in Folge des sich abkühlenden Klimas einherging.[99] Waren im Pliozän mit Boreostemma aus der bedeutenden Codore-Formation in Venezuela[6] noch frühe Formen der modernen Glyptodonten anzutreffen, ist im Altpleistozän vor etwa 1,8 Millionen Jahren erstmals eindeutig Glyptodon im südlichen Südamerika überliefert.[100] Zu den frühesten Funden gehört unter anderem ein 28 cm langer Schädelfund aus der Nähe von Tarija im südlichen Bolivien.[101] Ähnlich alte Funde stammen auch aus dem angrenzenden Pampagebiet, so dass der wahrscheinliche Ursprung der Gattung in dieser gesamten Region lag.[95] In der darauf folgenden Zeit stellt Glyptodon den dominierenden Vertreter der Glyptodonten dar. Die Gattung kam hauptsächlich vom südlichen Brasilien bis in die südlichen Bereiche des Kontinentes vor, war aber ebenfalls weiter nördlich in den Anden zu finden. Ihr Verbreitungsgebiet erstreckte sich zwischen dem 20. und 38. südlichen Breitengrad. Der ursprünglich angenommene Artenreichtum der Gattung ist allerdings möglicherweise ein Relikt der Forschung, da moderne Analysen lediglich zwei Formen in den Tieflandsgebieten unterscheiden können, die sich weitgehend zeitlich abgrenzen, zuzüglich einer dritten aus den Anden.[102][103][19] Auch Vertreter anderer Linien bewohnten weite Bereiche des Kontinentes. So hatte Panochthus eine größere geographische Reichweite und ökologische Toleranz und besiedelte sowohl tropische Areale als auch temperierte Gebiete vom heutigen nordöstlichen Brasilien bis weit in das südliche Argentinien hinein. Außerdem gehört Panochthus neben Glyptodon zu den wenigen Angehörigen der Gepanzerten Nebengelenktiere, die zu jener Zeit bis in Hochgebirgslagen von teils über 4000 m vordrangen.[18][19] Andere Formen waren dagegen regional stärker begrenzt. Hierzu zählen unter anderem Hoplophorus, das überwiegend in tropischen Landschaften vorkam, und Doedicurus sowie Plaxhaplous in der Pampa und im angrenzenden Mesopotamia.[39] Ein ähnliches Verbreitungsgebiet wies auch Neosclerocalyptus auf, das mit geschätzten 295 bis 470 kg Körpergewicht der kleinste alle pleistozänen Vertreter der Glyptodonten war. Einzigartig war bei ihm eine stark verknöcherte Nasenregion.[104][105][72]

Mit der Entstehung des Isthmus von Panama vor rund 3,5 Millionen Jahren während des Pliozän und der damit verbundenen Ausbildung einer Landbrücke zwischen Süd- und Nordamerika kam es zum Großen Amerikanischen Faunenaustausch. Die frühesten Glyptodontenreste nördlich von Südamerika, die aber nur einige wenige Osteoderme umfassen, sind im San-Miguel-Allende-Becken des mexikanischen Bundesstaates Guanajuato im zentralen Teil des Landes entdeckt worden und datieren auf etwa 3,6 bis 3,9 Millionen Jahre.[106] Im ausgehenden Pliozän ist dann erstmals der nordamerikanische Vertreter Glyptotherium nachgewiesen, dessen Knochenplättchen stärker ornamentiert sind als bei seinem Verwandten Glyptodon. Dabei traten zuerst eher kleine Angehörige dieser Gattung auf, die nur rund 230 kg wogen, erst im Verlauf des Pleistozän bildeten sich bis zu 790 kg schwere Formen aus.[107] In Nordamerika war Glyptotherium, der bisher einzige bisher anerkannte Vertreter der Glyptodonten dort, hauptsächlich entlang der Golfküste verbreitet, zahlreiche Funde sind aus Texas, Arizona und Florida bekannt. Einer der bedeutendsten und reichhaltigsten Fundpunkte ist mit der 111 Ranch in Arizona belegt. In Oklahoma erreichte Glyptotherium bei 36° 37′ sein nördlichstes Vorkommen, wie ein Rückenpanzer aus Carmen im Alfalfa County anzeigt.[108] Im Verlauf des Mittelpleistozäns verschwanden die Glyptodonten allmählich wieder aus ihren nördlichsten Refugien,[95][12] blieben aber noch sehr weit über das heutige Mexiko verbreitet.[27] Offensichtlich kam es zu dieser Zeit teilweise zu einer Rückwanderung nach Südamerika, da im ausgehenden Pleistozän Glyptotherium auch aus Venezuela[100] und Brasilien,[109][110] bekannt ist und dort Küstenflachländer bewohnte.[25]
Aussterben
Zum Ende des Pleistozän im Übergang zum Holozän starben die Glyptodonten im Zuge der Quartären Aussterbewelle aus. Für Glyptodon liegen die jüngsten direkt datierten Funde bei einem Alter von rund 25.500 Jahren von Inciarte in Venezuela. Jüngere Daten von 9600 bis 10.500 Jahren von Pay Paso in Uruguay wurden an beigefundener Holzkohle gewonnen, wobei eine gemeinsame Ablagerung der Reste von Glyptodon und Holzkohle nicht sicher ist. Deutlich länger überlebte Doedicurus. Es war noch während der Besiedlung Südamerikas durch den frühen Menschen anzutreffen, die vor etwa 14.500 Jahren begann.[111] Ob der Mensch aber ursächlich für das Verschwinden der großen Säugetierfauna verantwortlich ist, gehört zu einer vielfach diskutierten Debatte. In der Pamparegion hielt sich Doedicurus noch bis in das frühe Holozän. So sind Funde der Gattung, die einen Unterkiefer, mehrere Halswirbel, Knochen des Fußskelettes und andere Elemente des Körperskelettes sowie des Panzers einschließen, zusammen mit Steinartefakten früher Jäger-Sammler-Gruppen von La Moderna im nordöstlichen Argentinien auf ein Alter von 7500 Jahre BP datiert worden. Unklar ist hier, ob die Menschen das Tier selbst jagten oder einen Kadaver zerlegten.[112][113] Mit rund 8480 Jahren BP wenig älter ist ein Fragment eines Oberarmknochens, das südlich von Buenos Aires aufgefunden wurde.[2]
Forschungsgeschichte
Erste Entdeckungen im 18. und 19. Jahrhundert

Der früheste bekannt gewordene Fund eines Glyptodonten geht auf das Jahr 1774 zurück, als der englische Jesuit Thomas Falkner von einem rund 2,7 m langen Körperpanzer, bestehend aus sechseckigen Knochenplättchen berichtete, der am Ufer des Rio Carcarañá nahe Santa Fe in Argentinien entdeckt worden war und den er mit dem der heutigen Gürteltiere verglich.[114] Im Jahr 1814 erstellte Dàmaso Antonio Larrañaga (1771–1848) die erste wissenschaftliche Beschreibung eines Glyptodonten in seiner Schrift Diario de Historia Natural anhand eines Rückenpanzers, eines Oberschenkelknochens und eines Schwanzpanzers. Er fügte dieser die Bezeichnung Dasypus (Megatherium Cuv) zu, was 1823/24 von Georges Cuvier (1769–1832) in die zweite Auflage seines Werkes Recherches sur les ossemens fossiles, eines der Grundlagenwerke zur Entwicklung der Paläontologie, eingebracht wurde.[115] Die Angabe der Untergattung Megatherium, eigentlich ein riesiges Bodenfaultier, welches Cuvier 1796 selbst wissenschaftlich eingeführt hatte, führte dazu, dass in der darauf folgenden Zeit die Ansicht von gepanzerten Riesenfaultieren vertreten wurde. So etwa beschrieb 1827 Christian Samuel Weiss Panzerreste eines Glyptodonten aus dem heutigen Uruguay und Brasilien als zu Megatherium gehörig.[116] Auch William Clift verband in einer Publikation von 1835 ein Skelett von Megatherium aus der Umgebung von Villanueva am Río Salado in der Pamparegion südlich von Buenos Aires mit ebenfalls dort aufgefundenen Panzerresten von Glyptodonten und bildete diese auch ab.[117]
Sechs Jahre später untersuchte Joseph Eduard d’Alton das von Weiss verwendete Material erneut und schloss auch zusätzlich gefundene Skelettreste ein. Er kam nach ausführlichen anatomischen Vergleichen zu dem Schluss, dass es sich um riesige Gürteltiere handelte.[118] Heinrich Georg Bronn (1800–1862) verwendete die gleiche Fundkollektion für die Aufstellung der Gattung Chlamydotherium.[119] Allerdings ist der Name Chlamydotherium problematisch, einerseits weil er nahezu gleichzeitig von Peter Wilhelm Lund für einen Vertreter der Pampatheriidae, nahen Verwandten der Glyptodonten, verwendet worden war, andererseits, weil Bronn ihn als Genus coelebs (ungebundene Gattung) eingeführt hatte, ohne der Gattung eine spezifische Art zuzuweisen. Chlamydotherium gilt heute als synonym zu Glyptodon.[39] Im gleichen Zeitraum entdeckte eine Expedition unter der Leitung von Teodoro Vilardebó mehrere Knochen und einen Rückenpanzer in Uruguay, den sie nach wissenschaftlicher Analyse 1838 in einer lokalen Tageszeitung veröffentlichten. Die darin verwendete Bezeichnung Dasypus antiquus ist allerdings ungültig. Erst 1844, nachdem die Reste an das Muséum national d’histoire naturelle in Paris verschickt worden waren, konnte der Schweizer Zoologe François Jules Pictet diese als zu Glyptodon gehörig klassifizieren.[62][120]
Wissenschaftliche Namensgebungen

Den Gattungsnamen Glyptodon führte der englische Paläontologe Richard Owen (1804–1892) im Jahr 1839 ein,[121] eine weitere ausführliche Vorstellung der Gattung hier auch verbunden mit dem Artzusatz Glyptodon clavipes folgte zwei Jahre später.[122] Die Erstbeschreibung basierte auf einem Teilskelett, das in den 1830er Jahren in der Umgebung von Cañuelas am Río Matanza-Riachuelo südlich von Buenos Aires gefunden worden war. Woodbine Parish, ein hochrangiger britischer Diplomat in Buenos Aires, sandte die Fossilreste nach England, wo sie Owen eingehend untersuchte und dabei erkannte, dass es sich um einen Verwandten der Gürteltiere handelte. Er revidierte dabei auch alle angeblichen Panzerreste, die mit Megatherium in Verbindung gebracht worden waren. Der Name Glyptodon setzt sich aus den griechischen Wörtern γλύφειν (glyphein „einschneiden“; Partizip Perfekt γλύπτω, glypto) und ὀδούς (odoús „Zahn“) zusammen und bezieht sich auf die besondere Gestaltung der Zähne, die Owen im Vergleich zu den Gürteltieren sah.[121]
Etwa zur gleichen Zeit unternahm Charles Darwin (1809–1882) mit der HMS Beagle seine für die Evolutionsforschung bedeutende Forschungsreise und landete zwischen 1832 und 1834 mehrfach an der Küste Argentiniens an. Dort sammelte er über 5000 Fossilfunde, zumeist aus dem Pleistozän, die er an das Royal College of Surgeons in London weiterleitete (von denen haben aber nur insgesamt 175 Objekte die Bombardements Londons 1941 überstanden). Dort begann Owen ab 1836 mit dem Studium der Fossilien. Darwin selbst war aufgrund von Cuviers Angabe zu der Körperpanzerung von Megatherium in der neuen Ausgabe dessen Werkes Recherches sur les ossemens fossiles von gepanzerten Riesenfaultieren überzeugt und wies diesen in seinen Reiseaufzeichnungen zahlreiche Funde zu, was Owen im Nachfolgenden korrigierte.[123] Parallel zu Darwins Unternehmungen und Owen Analysen arbeitete der dänische Forscher Peter Wilhelm Lund (1801–1880) in Brasilien, wo er in der Region des Rio das Velhas eine über 12.000 Fossilien umfassende Kollektion von rund 800 Fundstellen, überwiegend Höhlen, zusammentrug und nach Kopenhagen verschiffte. Diese Tätigkeiten führten 1838 zur Beschreibung von Hoplophorus durch Lund.[124][125][120]
_(18169666261).jpg.webp)

Im Jahr 1869 etablierte John Edward Gray (1800–1875) den heute für die Familie genutzten Namen Glyptodontidae. In diese schloss er neben Glyptodon auch Hoplophorus, Panochthus und Schistopleurum ein, letzteres ist heute ein Synonym zu Hoplophorus. Als Hauptmerkmale nannte er:
Dorsal shield entire, not revolute, immovable affixed to the upper part of the very large pelvis.
„Rückenschild vollständig, nicht rollbar, unbeweglich am oberen Teil des sehr großen Beckens befestigt.“
Wenige Jahre zuvor hatte der deutsche Forscher Hermann Burmeister (1807–1892), der jahrelang in Südamerika tätig war, den Begriff Biloricata („Zweischaler“) für die Glyptodonten eingeführt, unter der Annahme, das auf dem Bauch ebenfalls ein, wenn auch dünnerer Panzer ausgebildet sei.[126] Diese Überlegung arbeitete Gray in seine Beschreibung ein, wobei er zusätzlich vermerkte, dass wohl der Kopf analog zu den Schildkröten in den Panzer zurückgezogen werden könne.[127]
Im Jahr 1875 wurden erstmals außerhalb Südamerikas Reste von Glyptodonten in Form eines vollständigen Rückenpanzers im Tal von Mexiko entdeckt, denen einige weitere bis Anfang des 20. Jahrhunderts folgten, so 1912 ebenfalls ein vollständiger Panzer nebst einigen Skelettteilen.[128] Bereits 1888 beschrieb Edward Drinker Cope (1840–1897) einzelne Osteoderme aus Texas und lieferte damit den frühesten Nachweis der Glyptodonten nördlich von Mexiko. Erst 1903 publizierte Henry Fairfield Osborn (1857–1935) ein nahezu vollständiges Skelett von Glyptotherium und beschrieb damit die einzige heute anerkannte Gattung Nordamerikas.[129][12]
Die Wende vom 19. zum 20. Jahrhundert
Im Übergang vom 19. zum 20. Jahrhundert heben sich die Arbeiten der Gebrüder Carlos und Florentino Ameghino heraus. Im Jahr 1889 erarbeitete Florentino Ameghino erstmals eine systematische Gliederung der Glyptodonten, die 13 der 19 damals bekannten Gattungen umfasste. Er ordnete diese in drei Familien (Glyptodontidae, Doedicuridae und Hoplophoridae). Grundlagen seiner systematischen Aufteilung waren die unterschiedliche Gestaltung der Osteoderme und der Schwanzpanzerung. Dabei erkannte Ameghino als erster die zwei grundsätzlichen Schwanztypen der Glyptodonten: zum einen der Schwanz, der vollständig umringt war und in einer kurzen Spitze endete, typisch für Glyptodon und zum anderen der Schwanz mit einem keulenartigen Ende, wie er bei Doedicurus vorkam.[130] Diese Methode zur Untergliederung der Glyptodonten wurde auch später von anderen Wissenschaftlern verwendet.[70] Zu den weiteren herausragenden Forschern zu den Glyptodonten gehören unter anderem Lucas Kraglievich in den 1930er Jahren sowie Robert Hoffstetter in den 1950er Jahren.[131]
Literatur
- Richard M. Fariña, Sergio F. Vizcaíno und Gerardo de Iuliis: Megafauna. Giant beasts of Pleistocene South America. Indiana University Press, 2013, ISBN 978-0-253-00230-3
- Paul S. Martin und Richard G. Klein (Hrsg.): Quaternary Extinctions. A Prehistoric Revolution. The University of Arizona Press, Tucson AZ 1984, ISBN 0-8165-1100-4
Einzelnachweise
- Sergio F. Vizcaíno, Juan C. Fernicola und M. Susana Bargo: Paleobiology of Santacrucian glyptodonts and armadillos (Xenarthra, Cingulata). In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene paleobiology in Patagonia: High-latitude paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 194–215
- Leopold Héctor Soibelzon, Martín Zamorano und Gustavo Juan Scillato-Yané: Un Glyptodontidae de gran tamaño en el Holoceno Temprano de la Region Pampeana, Argentinia. Revista Brasileira de Paleontologia 15 (1), 2012, S. 105–112
- Kieren J. Mitchell, Agustin Scanferla, Esteban Soibelzon, Ricardo Bonini, Javier Ochoa und Alan Cooper: Ancient DNA from the extinct South American giant glyptodont Doedicurus sp. (Xenarthra: Glyptodontidae) reveals that glyptodonts evolved from Eocene armadillos. Molecular Ecology 25 (14), 2016, S. 3499–3508, doi:10.1111/mec.13695
- Sergio F. Vizcaíno, R. Ernesto Blanco, J. Benjamí Bender und Nick Milne: Proportions and function of the limbs of glyptodonts. Lethaia 44, 2011, S. 93–101
- Fariña, Vizcaíno und de Iuliis 2013. S. 223–234
- Alfredo A. Carlini, Alfredo E. Zurita, Gustavo J. Scillato-Yané, Rodolfo Sánchez und Orangel A. Aguilera: New Glyptodont from the Codore Formation (Pliocene), Falcón State, Venezuela, its relationship with the Asterostemma problem, and the paleobiogeography of the Glyptodontinae. Paläontologische Zeitschrift 82 (2), 2008, S. 139–152
- Richard A. Fariña und Sergio F. Vizcaíno: Carved teeth and strange jaws. How glyptodonts masticated. Acta Palaeontologica Polonica 46 (2), 2001, S. 219–234
- Sergio F. Vizcaíno, Richard A. Fariña, M. Susana Bargo und Gerardo de Iuliis: Functional and phylogenetic assessment of the masticatory adaptations in Cingulata (Mammalia, Xenarthra). Ameghiniana 41 (4), 2004, S. 651–664
- Alfredo Eduardo Zurita und Silvia A. Aramayo: New remains of Eosclerocalypto tapinocephalus (Cabrera) (Mammalia, Xenarthra, Glyptodontidae): Description and implication for its taxonomic status. Revista Italiana di Paleontologia e Stratigrafia 113 (1), 2007, S. 57–66
- Alfredo Eduardo Zurita, Martín Zamorano, Gustavo J. Scillato-Yané, Laureano R. González-Ruiz, Santiago Rodríguez-Bualó, Boris Rivas Durán und Ricardo Céspedes Paz: An exceptional Pleistocene specimen of Panochthus Burmeister (Xenarthra, Glyptodontoidea) from Bolivia: Its contribution to the understanding of the Early-Middle Pleistocene Panochthini. Comptes Rendus Palevol 10, 2011, S. 655–664
- H. Gregory McDonald: Xenarthran skeletal anatomy: primitive or derived? Senckenbergiana biologica 83, 2003, S. 5–17
- David D. Gillette und Clayton E. Ray: Glyptodonts of North America. Smithonian Contributions to Paleobiology 40, 1981, S. 1–251
- Sergio F. Vizcaíno: The teeth of the “toothless”: novelties and key innovations in the evolution of xenarthrans (Mammalia, Xenarthra). Paleobiology 35 (3), 2009; S. 343–366
- Daniela C. Kalthoff: Microstructure of Dental Hard Tissues in Fossil and Recent Xenarthrans (Mammalia: Folivora and Cingulata). Journal of Morphology 272, 2011, S. 641–661
- Laureano R. González-Ruiz, Martin R. Ciancio, Gabriel M. Martin und Alfredo E. Zurita: First Record of Supernumerary Teeth in Glyptodontidae (Mammalia, Xenarthra, Cingulata). Journal of Vertebrate Paleontology 35 (1), 2015, S. e885033
- Kevin F. Downing und Richard S. White: The cingulates (Xenarthra) of the Leisey Shell Pit local fauna (Irvingtonian), Hillborough County, Florida. Bulletin of the Florida Museum of Natural History 37 (Part II), 1995, S. 375–396
- Kleberson de O. Porpino, Juan C. Fernicola und Lílian P. Bergqvist: Revisiting the Intertropical Brazilian Species Hoplophorus euphractus (Cingulata, Glyptodontoidea) and the Phylogenetic Affinities of Hoplophorus. Journal of Vertebrate Paleontology 30 (3), 2010, S. 911–927
- Alfredo Eduardo Zurita, Martín Zamorano, Gustavo Juan Scillato-Yané, Sergio Fidel, Martín Iriondo und David D. Gillette: A new species of Panochthus Burmeister (Xenarthra, Cingulata, Glyptodontidae) from the Pleistocene of the Eastern Cordillera, Bolivia. Historical Biology 29 (8), 2017, S. 1076–1088, doi:10.1080/08912963.2016.1278443
- Francisco Cuadrelli, Alfredo E. Zurita, Pablo Toriño, Angel R. Mio-Boilini, Daniel Perea, Carlos A. Luna, David D. Gillette und Omar Medina: A new species of glyptodontine (Mammalia, Xenarthra, Glyptodontidae) from the Quaternary of the Eastern Cordillera, Bolivia: phylogeny and palaeobiogeography. Journal of Systematic Palaeontology, 18, 2020, S. 1543–1566, doi:10.1080/14772019.2020.1784300
- Robert V. Hill: Comparative Anatomy and Histology of Xenarthran Osteoderms. Journal of Morphology 267, 2005, S. 1441–1460
- Alfredo Eduardo Zurita, Leopoldo Hector Soibelzon, Esteban Soibelzon, Germán Mariano Gasparini, Marcos Martín Cenizo und Héctor Arzani: Accessory protection structures in Glyptodon Owen (Xenarthra, Cingulata, Glyptodontidae). Annales de Paléontologie 96, 2010, S. 1–11
- R. Ernesto Blanco, Washington W. Jones und Andrés Rinderknecht: The Sweet Spot of a Biological Hammer: The Centre of Percussion of Glyptodont (Mammalia:Xenarthra) Tail Clubs. Proceedings of the Royal Society B 276 (1675), 2009, S. 3971–3978
- Victoria M. Arbour und Lindsay E. Zanno: The evolution of tail weaponization in amniotes. Proceedings of the Royal Society B 285, 2018, S. 20172299, doi:10.1098/rspb.2017.2299
- Victoria M. Arbour und Lindsay E. Zanno: Tail Weaponry in Ankylosaurs and Glyptodonts: An Example of a Rare but Strongly Convergent Phenotype. The Anatomical Record 303 (4), 2020, 988–998, doi:10.1002/ar.24093
- Alfredo E. Zurita, Ángel R. Miño-Boilini, Analía Francia und José E. Arenas-Mosquera: The Pleistocene Glyptodontidae Gray 1869 (Xenarthra: Cingulata) of Colombia and some considerations about the South American Glyptodontidae. Revista Brasileira de Paleontologia 15 (3), 2012, S. 273–280
- François Pujos und Rodolfo Salas: A systematic reassessment and paleogeographic review of fossil Xenarthra from Peru. Bulletin de l'Institut Français d'Etudes Andines 33 (2), 2004, S. 331–377
- Jim I. Mead, Sandra L. Swift, Richard S. White, H. Greg McDonald und Arturo Baez: Late Pleistocene (Rancholabrean) Glyptodont and Pampathere (Xenarthra, Cingulata) from Sonora, Mexico. Revista Mexicana de Ciencias Geológica 24 (3), 2007, S. 439–449
- Nick Milne, Sergio F. Vizcaíno und Juan Carlos Fernicola: A 3D geometric morphometric analysis of digging ability in the extant und fossil cingulated humerus. Journal of Zoology 278, 2009, S. 48–56
- Fiona R. Straehl, Torsten M. Scheyer, Analía M. Forasiepi, Ross D. MacPhee und Marcelo R. Sánchez Villagra: Evolutionary Patterns of Bone Histology and Bone Compactness in Xenarthran Mammal Long Bones. PLoS ONE 8 (7), 2013, S. e69275, doi:10.1371/journal.pone.0069275
- R. McNeill Alexander, Richard A. Fariña und Sergio F. Vizcaíno: Tail blow energy and carapace fractures in a large glyptodont (Mammalia, Xenarthra). Zoological Joumal of the Linnean Society 126, 1999, S. 41–49
- P. Sebastián Tambusso, Luciano Varela, Flávio Góis, Jorge Felipe Moura, Chiara Villa und Richard A. Fariña: The inner ear anatomy of glyptodonts and pampatheres (Xenarthra, Cingulata): Functional and phylogenetic implications. Journal of South American Earth Sciences, 2021, S. 103189, doi:10.1016/j.jsames.2021.103189
- Silvia A. Aramayo, Teresa Manera de Bianco, Nerea V. Bastianelli und Ricardo N. Melchor: Pehuen Co: Updated taxonomic review of a late Pleistocene ichnological site in Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 439, 2015, S. 144–165
- Sergio F Vizcaíno, Guillermo H. Cassini, Juan C. Fernicola und M. Susana Bargo: Evaluating habitats and feeding habits through ecomorphological features in glyptodonts (Mammalia, Xenarthra). Amegjiniana 48 (3), 2011, S. 305–319
- Sergio F. Vizcaíno, M. Susana Bargo und Guillermo H. Cassini: Dental occlusal surface area in relation to body mass, food habits and other biological features in fossil xenarthrans. Ameghiniana 43 (1), 2006, S. 11–26
- Leandro M. Pérez,Néstor Toledo, Gerardo De Iuliis, M. Susana Bargo und Sergio F. Vizcaíno: Morphology and Function of the Hyoid Apparatus of Fossil Xenarthrans (Mammalia). Journal of Morphology 271, 2010, S. 1119–1133
- Martín Zamorano, Gustavo Juan Scillato-Yané, Esteban Soibelzon, Leopoldo Héctor Soibelzon, Ricardo Bonini und Sergio Gabriel Rodriguez: Hyoid apparatus of Panochthus sp. (Xenarthra; Glyptodontidae) from the Late Pleistocene of the Pampean Region (Argentina). Comparative description and muscle reconstruction. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 288/2, 2018, S. 205–219
- Mariella Superina: Biologie und Haltung von Gürteltieren (Dasypodidae). Universität Zürich, 2000, S. 1–248 (S. 74)
- Carlos A. Luna und Jerónimo M. Krapovickas: Primer registro de un ejemplar juvenil de Glyptodon sp. (Cingulata, Glyptodontidae) del Cuaternario de la provincia de Córdoba, Argentina. Mastozoología Neotropical 18 (1), 2011, S. 135–141
- Alfredo Eduardo Zurita, Edison Vicente Oliveira, Pablo Toriño, Santiago Martín Rodriguez-Bualó und Gustavo Juan Scillato-Yané: On the taxonomic status of some Glyptodontidae (Mammalia, Xenarthra, Cingulata) from the Pleistocene of South America. Annales de Paléontologie 97, 2011, S. 63–83
- Carlos A. Luna, Ignacio A. Cerda, Alfredo E. Zurita, Romina Gonzalez, M. Cecilia Prieto, Dimila Mothé, und Leonardo S. Avilla: Distinguishing Quaternary glyptodontine cingulates in South America: How informative are juvenile specimens? Acta Palaeontologica Polonica 63 (1), 2018, S. 159–170, doi:10.4202/app.00409.2017
- Alfredo Eduardo Zurita, Angel R. Miño-Boilini, Esteban Soibelzon, Gustavo J. Scillato-Yané, Germán M. Gasparini und Freddy Paredes-Ríos: First record and description of an exceptional unborn specimen of Cingulata Glyptodontidae: Glyptodon Owen (Xenarthra). Comptes Rendus Palevol 8, 2009, S. 573–578
- Daniel Perea, Pablo Toriño und Martín Ghizzoni: First endoskeletal remains of Neuryurus (Xenarthra, Glyptodontidae), an emended diagnosis of the genus, and bodymass estimations. Journal of Vertebrate Paleontology 39 (4), 2019, S. e1668400, doi:10.1080/02724634.2019.1668400
- Laureano R. González Ruiz, Diego Brandoni, Alfredo R. Zurita, Jeremy L. Green, Nelson M. Novo, Adan A. Tauber und Marcelo F. Tejedor: Juvenile glyptodont (Mammalia, Cingulata) from the Miocene of Patagonia, Argentina: Insights into mandibular and dental characters. Journal of Vertebrate Paleontology 40 (1), 2020, S. 1768398, doi:10.1080/02724634.2020.1768398
- Martín Zamorano und Richard A. Fariña: Changes in form and function of the caudal tubes in Panochthus (Xenarthra; Glyptodontidae) along the Pleistocene. Historical Biology: An International Journal of Paleobiology, 2021, doi:10.1080/08912963.2021.2012767
- Daniel Barasoain, Alfredo E. Zurita, Darin A. Croft, Claudia I. Montalvo, Víctor H. Contreras, Ángel R. Miño-Boilini und Rodrigo L. Tomassini: A New Glyptodont (Xenarthra: Cingulata) from the Late Miocene of Argentina: New Clues About the Oldest Extra-Patagonian Radiation in Southern South America. Journal of Mammalian Evolution, 2022, doi:10.1007/s10914-021-09599-w
- Martín de los Reyes, Daniel Poiré, Leopoldo Soibelzon, Alfredo E. Zurita und M. J. Arrouy: First evidence of scavenging of a Glyptodont (Mammalia, Glyptodontidae) from the Pliocene of the Pampean region (Argentina): taphonomic and paleoecological remarks. Palaeontologia Electronica 16 (2), 2013, 15A (online)
- P. Sebastián Tambusso und Richard A. Fariña: Digital cranial endocast of Pseudoplohophorus absolutus (Xenarthra, Cingulata) and its systematic and evolutionary implications. Journal of Vertebrate Paleontology 35 (5), 2015, S. e967853, doi:10.1080/02724634.2015.967853
- P. Sebastián Tambusso und Richard A. Fariña: Digital endocranial cast of Pampatherium humboldtii (Xenarthra, Cingulata) from the Late Pleistocene of Uruguay. Swiss Journal of Palaeontology 134, 2015, S. 109–116, doi:10.1007/s13358-015-0070-5
- Rodrigo L. Tomassini, Claudia I. Montalvo und María C. Ezquiaga: The oldest record of flea/armadillos interaction as example of bioerosion on osteoderms from the late Miocene of the Argentine Pampas. International Journal of Paleopathology 15, 2016, S. 65–68, doi:10.1016/j.ijpp.2016.08.004
- Fábio Cunha Guimarães de Lima und Kleberson de Oliveira Porpino: Ectoparasitism and infections in the exoskeletons of large fossil cingulates. PLoS ONE 13 (10), 2018, S. e0205656, doi:10.1371/journal.pone.0205656
- Fernando Henrique de Souza Barbosa und Carlos A. Luna: Lesiones en la coraza y endoesqueleto de un ejemplar de Panochthus Burmeister, 1866 (Mammalia, Glyptodontia) de la provincia de Córdoba, Argentina. Revista del Museo Provincial de Ciencias Naturales “Dr. Arturo Umberto Illia” 1, 2014, S. 9–26
- Fernando Henrique de Souza Barbosa, Kleberson de Oliveira Porpino, Ana Bernadete Lima Fragoso und Edison Vicente Oliveira: Arthritis in a Glyptodont (Mammalia, Xenarthra, Cingulata). PLoS ONE 9 (2), 2014, S. e88646, doi:10.1371/journal.pone.0088646
- Fernando Henrique de Souza Barbosa, Kleberson de Oliveira Porpino, Bruce M. Rothschild, Uiara G. Cabral und Lilian P. Bergqvist: Arthritic lesions and congenital fusion in foot bones of Panochthus sp. (Xenarthra, Cingulata). Anais da Academia Brasileira de Ciências 91(Suppl. 1), 2019. S. e20160812, doi:10.1590/0001-3765201720160812
- Guillaume Billet, Lionel Hautier, Christian de Muizon and Xavier Valentin: Oldest cingulate skulls provide congruence between morphological and molecular scenarios of armadillo evolution. Proceedings of the Royal Society B, 278, 2011, S. 2791–2797
- Maureen A. O'Leary, Jonathan I. Bloch, John J. Flynn, Timothy J. Gaudin, Andres Giallombardo, Norberto P. Giannini, Suzann L. Goldberg, Brian P. Kraatz, Zhe-Xi Luo, Jin Meng, Xijun Ni, Michael J. Novacek, Fernando A. Perini, Zachary S. Randall, Guillermo W. Rougier, Eric J. Sargis, Mary T. Silcox, Nancy B. Simmons, Michelle Spaulding, Paúl M. Velazco, Marcelo Weksler, John R. Wible und Andrea L. Cirranello: The Placental Mammal Ancestor and the Post–K-Pg Radiation of Placentals. Science 339, 2013, S. 662–667, doi:10.1126/science.1229237
- Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, S. 1–431 (S. 200–204)
- Sergio F. Vizcaíno und W. J. Loughry: Xenarthran biology: Past, present and future. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 1–7
- Frédéric Delsuc, Michael J. Stanhope und Emmanuel J.P. Douzery: Molecular systematics of armadillos (Xenarthra, Dasypodidae): contribution of maximum likelihood and Bayesian analyses of mitochondrial and nuclear genes. Molecular Phylogenetics and Evolution 28, 2003, S. 261–275
- Frédéric Delsuc, Sergio F Vizcaíno und Emmanuel J. P. Douzery: Influence of Tertiary paleoenvironmental changes on the diversification of South American mammals: a relaxed molecular clock study within xenarthrans. BMC Evolutionary Biology 4 (11), 2004, S. 1–13
- Sergio F. Vizcaíno, Marcelo Zaráte, M. Susana Bargo und Alejandro Dondas: Pleistocene burrows in the Mar del Plata area (Argentina) and their probable builders. Acta Palaeontologica Polonica 46 (2), 2001, S. 289–301
- Timothy J. Gaudin und John R. Wible: The Phylogeny of Living and Extinct Armadillos (Mammalia, Xenarthra, Cingulata): A Craniodental Analysis. In: M. T. Carrano, T. J. Gaudin, R. W. Blob und J. R. Wible (Hrsg.): Amniote Paleobiology. Chicago/London: University of Chicago Press, 2006, S. 153–198
- Juan-Carlos Fernicola, Sergio F. Vizcaíno und Richard a. Fariña: The evolution of armored xenarthrans and the phylogeny of the glyptodonts. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 79–85
- Juan C. Fernicola, Andrés Rinderknecht, Washington Jones, Sergio F. Vizcaíno und Kleberson Porpino: A new species of Neoglyptatelus (Mammalia, Xenarthra, Cingulata) from the late Miocene of Uruguay provides new insights on the evolution of the dorsal armor in cingulates. Ameghiniana 55, 2018, S. 233–252
- Frédéric Delsuc, Gillian C. Gibb, Melanie Kuch, Guillaume Billet, Lionel Hautier, John Southon, Jean-Marie Rouillard, Juan Carlos Fernicola, Sergio F. Vizcaíno, Ross D. E. MacPhee und Hendrik N. Poinar: The phylogenetic affinities of the extinct glyptodonts. Current Biology 26, 2016, S. R155–R156, doi:10.1016/j.cub.2016.01.039
- Darin A. Croft, John J. Flynn und André R. Wyss: A new basal glyptodontid and other Xenarthra of the Early Miocene Chucal fauna, Northern Chile. Journal of Vertebrate Paleontology 27 (4), 2007, S. 781–797
- Sergio F. Vizcaíno, Andrés Rinderknecht und Ada Czerwonogora: An enigmatic Cingulata (Mammalia: Xenarthre) from the Late Miocene of Uruguay. Journal of Vertebrate Paleontology 23 (4), 2003, S. 981–983
- Martín Zamorano und Diego Brandini: Phylogenetic analysis of the Panochthini (Xenarthra, Glyptodontidae), with remarks on their temporal distribution. Alcheringa 37, 2013, S. 442–451
- Kleberson De O. Porpino, Juan C. Fernicola, Laura Edith Cruz und Lílian P. Bergqvist: The Intertropical Brazilian Species of Panochthus (Xenarthra, Cingulata, Glyptodontoidea): A Reappraisal of Their Taxonomy and Phylogenetic Affinities. Journal of Vertebrate Paleontology 34 (5), 2014, S. 1165–1179
- Pablo Toriño und Daniel Perea: New contributions to the systematics of the "Plohophorini" (Mammalia, Cingulata, Glyptodontidae) from Uruguay. Journal of South American Earth Sciences 86, 2018, S. 410–430, doi:10.1016/j.jsames.2018.07.006
- Juan Carlos Fernicola und K. O. Porpino: Exoskeleton and Systematics: A Historical Problem in the Classification of Glyptodonts. Journal of Mammal Evolution 19, 2012, S. 171–183
- Juan Carlos Fernicola: Nuevos aportes para la sistemática de los Glyptodontia Ameghino 1889 (Mammalia, Xenarthra, Cingulata). Ameghiniana 45, 2008, S. 553–574
- Sofía I. Quiñones, Martin De los Reyes, Alfredo E. Zurita, Francisco Cuadrelli, Ángel R. Miño-Boilini und Daniel G. Poiré: Neosclerocalyptus Paula Couto (Xenarthra, Glyptodontidae) in the late Pliocene-earliest Pleistocene of the Pampean region (Argentina): Its contribution to the understanding of evolutionary history of Pleistocene glyptodonts. Journal of South American Earth Sciences 103, 2020, S. 102701, doi:10.1016/j.jsames.2020.102701
- Alfredo E. Zurita, M. Taglioretti, Martín de los Reyes, Martín Zamorano, F. Scaglai und Gustavo Juan Scillato-Yané: Neogene skulls of Glyptodontidae Doedicurinae (Mammalia, Xenarthra): the first evidence. In: Diego Brandoni u. a. (Hrsg.): II Simposio del Mioceno-Pleistoceno del Centro y Norte de Argentina. Diamante, 25-27 de Septiembre de 2013. Diamante, 2013, S. 32
- Alfredo E. Zurita, Matías Taglioretti, Martín de los Reyes, Cristian Oliva und Fernando Scaglia: First Neogene skulls of Doedicurinae (Xenarthra, Glyptodontidae): morphology and phylogenetic implications. Historical Biology: An International Journal of Paleobiology 28 (3), 2016, S. 423–432, doi:10.1080/08912963.2014.969254
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 87–91)
- Alfredo A. Carlini, Sergio F. Vizcaíno und Gustavo J. Scillato-Yané: Armored Xenarthtans: A unique taxonomic and ecologic assemblage. In: Richard F. Kay, Richard H. Madden, Richard L. Cifelli und John J. Flynn (Hrsg.): Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, 1997, S. 213–226
- Gustavo J. Scillato-Yané und Alfredo A. Carlini: Nuevos Xenarthra del Friasense (Mioceno medio) de Argentina. Studia Geologica Salmanticensia, 34, 1998, S. 43–67
- Alfredo E. Zurita, Alfredo A. Carlini und Gustavo J. Scillato-Yané: Paleobiogeography, biostratigraphy and systematics of the Hoplophorini (Xenarthra, Glyptodontoidea, Hoplophorinae) from the Ensenadan Stage (early Pleistocene to early-middle Pleistocene). Quaternary International 210, 2009, S. 82–92
- Laureano Raúl González-Ruiz, Alfredo Eduardo Zurita, Gustavo Juan Scillato-Yané, Martin Zamorano und Marcelo Fabián Tejedor: Un nuevo Glyptodontidae (Mammalia, Xenarthra, Cingulata) del Mioceno de Patagonia (Argentina) y comentarios acerca de la sistemática de los gliptodontes “friasenses”. Revista Mexicana de Ciencias Geológicas 28 (3), 2011, S. 566–579
- Laura E. Cruz, Juan C. Fernicola, Matias Taglioretti und Nestor Toledo: A reassessment of the taxonomic status of Paraglyptodon Castellanos, 1932 (Mammalia, Cingulata, Glyptodontia). Journal of South American Earth Sciences 66, 2016, S. 32–40
- Alfredo E. Zurita, Francisco Cuadrelli, Rodrigo L. Tomassini, Martín de los Reyes, Carlos Luna und Pablo Toriño: On the status of “Urotherium antiquum” (Ameghino) (Xenarthra, Glyptodontidae). Comptes Rendus Palevol 16 (3), 2017, S. 303–311, doi:10.1016/j.crpv.2016.08.008
- Martín Zamorano, Esteban Soibelzon und Eduardo P. Tonni: Giants of the Pampean plains (Argentina) during Early Pleistocene (Ensenadan). The case of Panochthus (Xenarthra, Glyptodontidae): comparative descriptions. Neues Jahrbuch für Geologie und Paläontologie 302 (1), 2021, S. 35–51
- Florentino Ameghino: Mammifères crétacés de l'Argentine. (Deuxième contribution à la connaissance de la faune mammalogique des couches à Pyrotherium). Boletin del Instituto Geográfico Argentino 18, 1897, S. 406–521 (S. 507) ()
- Florentino Ameghino: Notice préliminaires sur des mammifères nouveaux des terrains crétacés de Patagonie. Boletin de la Academia Nacional de Ciencias en Córdoba, República Argentina 17, 1902, S. 5–70 (S. 51) ()
- George Gaylord Simpson: The beginning of the age of mammals in South America. Part 1. Introduction. Systematics: Marsupialia, Edentata, Condylarthra, Litopterna and Notioprogonia. Bulletin of the American Museum of Natural History 91, 1948, S. 1–232 (S. 93)
- Malcolm McKenna, André R. Wyss und John J. Flynn: Paleogene Pseudoglyptodont Xenarthrans from Central Chile and Argentine Patagonia. American Museum Novitates 3536, 2006, S. 1–18
- Lílian Paglarelli Bergqvist, Érika Aparecida Leite Abrantes und Leonardo dos Santos Avilla: The Xenarthra (Mammalia) of São José de Itaboraí Basin (upper Paleocene, Itaboraian), Rio de Janeiro, Brazil. Geodiversitas 26 (2), 2004, S. 323–337
- Rosendo Pascual, Edgardo Ortiz Jaureguizar und José Luis Prado: Land Mammals: Paradigm for Cenozoic South American geobiotic evolution. Münchner Geowissenschaftliche Abhandlungen A 30, 1996, S. 265–319 ()
- François Pujos, Martin R. Ciancio, Analía M. Forasiepi, Michel Pujos, Adriana M. Candela, Bárbara Vera, Marcelo A. Reguero, Ana María Combina und Esperanza Cerdeño: The late Oligocene xenarthran fauna of Quebrada Fiera (Mendoza, Argentina) and its implications for sloth origins and the diversity of Palaeogene cingulates. Papers in Paleontology, 2021, doi:10.1002/spp2.1356
- Alfredo E. Zurita, Laureano R. González-Ruiz, Angel R. Miño-Boilini, Rafael Herbst, Gustavo J. Scillato-Yané und Pedro Cuaranta: Paleogene Glyptodontidae Propalaehoplophorinae (Mammalia, Xenarthra) in extra-Patagonian areas. Andean Geology 43 (1), 2016, S. 127–136, doi:10.5027/andgeoV43n1-a07
- Sergio F. Vizcaíno, M. Susana Bargo, Richard F. Kay, Richard A. Fariña, Mariana Di Giacomo, Jonathan M. G. Perry, Francisco J. Prevosti, Néstor Toledo, Guillermo H. Cassini und Juan C. Fernicola: A baseline paleoecological study for the Santa Cruz Formation (late–early Miocene) at the Atlantic coast of Patagonia, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 292, 2010, S. 507–519
- Laureano R. González-Ruiz, Augustina Reato, Martín Cano und Oscar Martínez: Old and new specimens of a poorly known glyptodont from the Miocene of Patagonia and their biochronological implications. Acta Palaeontologica Polonica 62 (1), 2017, S. 181–194, doi:10.4202/app.00280.2016
- Alfredo E. Zurita, Laureano R. González Ruiz, Arley J. Gómez-Cruz und Jose E. Arenas-Mosquera: The Most Complete Known Neogene Glyptodontidae (Mammalia, Xenarthra, Cingulata) from Northern South America: Taxonomic, Paleobiogeographic, and Phylogenetic Implications. Journal of Vertebrate Paleontology 33 (3), 2013, S. 696–708
- Julia V. Tejada-Lara, Rodolfo Salas-Gismondi, François Pujos, Patrice Baby, Mouloud Benammi, Stéphane Brusset, Dario De Franceschi, Nicolas Espurt, Mario Urbina und Pierre-Olivier Antoine: Life in proto-Amazonia: Middle Miocene mammals from the Fitzcarrald Arch (Peruvian Amazonia). Palaeontology 58 (2), 2015, S. 341–378
- Alfredo E. Zurita, Alfredo A. Carlini, David Gillette und Rodolfo Sánchez: Late Pliocene Glyptodontinae (Xenarthra, Cingulata, Glyptodontidae) of South and North America: Morphology and paleobiogeographical implications in the GABI. Journal of South American Earth Sciences 31, 2011, S. 178–185
- Cristian Oliva, Alfredo Eduardo Zurita, Alejandro Dondas und Gustavo Juan Scillato-Yané: Los Glyptodontinae (Xenarthra, Glyptodontidae) del Piso/Edad Chapadmalalense (Plioceno tardío): revisión y aportes a su conocimiento. Revista Mexicana de Ciencias Geológicas 27 (1), 2010, S. 112–120
- Gustavo J. Scillato-Yané, Flávio Góis, Alfredo E. Zurita, Alfredo A. Carlini Laureano R. Gonzáles Ruiz, Cecilia M. Krmpotic, Cristian Oliva und Martín Zamorano: Los Cingulata (Mammalia, Xenarthra) del “Conglomerado osífero” (Mioceno Tardío)de la Formación Ituzaingó de Entre Ríos, Argentina. In: Diego Brandoni und Jorge I. Noriega (Hrsg.): El Neógeno de la Mesopotamia argentina. Buenos Aires, 2013, S. 118–134
- Alizia Núñez-Blasco, Alfredo E. Zurita, Ángel R. Miño-Boilini, Ricardo A. Bonini und Francisco Cuadrelli: The glyptodont Eleutherocercus solidus from the late Neogene of North-Western Argentina: Morphology, chronology, and phylogeny. Acta Palaeontologica Polonica 66 (3), 2021, S. s079-s099, doi:10.4202/app.00824.2020
- Alfredo Eduardo Zurita, Alfredo Armando Carlini und Gustavo Juan Scillato-Yané: A new species of Neosclerocalyptus Paula Couto, 1957 (Xenarthra, Glyptodontidae, Hoplophorinae) from the middle Pleistocene of the Pampean region, Argentina. Geodiversitas 30 (4), 2008, S. 779–791
- Alfredo A. Carlini, Alfredo E. Zurita und Orangel A. Aguilera: North American Glyptodontines (Xenarthra, Mammalia) in the Upper Pleistocene of northern South America. Paläontologische Zeitschrift 82 (2), 2008, S. 125–138
- Alfredo Eduardo Zurita, Ángel Ramón Miño-Boilini, Esteban Soibelzon, Alfredo Armando Carlini und Freddy Paredes Ríos: The diversity of Glyptodontidae (Xenarthra, Cingulata) in the Tarija Valley (Bolivia): systematic, biostratigraphic and paleobiogeographic aspects of a particular assemblage. Neues Jahrbuch für Geologie und Paläontologie 251 (2), 2009, S. 225–237
- Francisco Cuadrelli, Alfredo E. Zurita, Pablo Toriño, ÁNgel R. Miño-Boilini, Santiago Rodríguez-Bualó, Daniel Perea und Gabriel E. Acuña Suárez: Late Pleistocene Glyptodontinae (Mammalia, Xenarthra, Glyptodontidae) from southern South America: a comprehensive review. Journal of Vertebrate Paleontology 38 (5), 2018, S. e1525390, doi:10.1080/02724634.2018.1525390
- Alfredo Eduardo Zurita, David D.Gillette, Francisco Cuadrelli und Alfredo Armando Carlini: A tale of two clades: Comparative study of Glyptodon Owen and Glyptotherium Osborn (Xenarthra, Cingulata, Glyptodontidae). Geobios 51 (3), 2018, S. 247–258, doi:10.1016/j.geobios.2018.04.004
- Alfredo E. Zurita, Matias Taglioretti, Martin Zamorano, Gustavo J. Scillato-Yané, Carlos Luna, Daniel Boh und Mariano Magnussen Saffer: A new species of Neosclerocalyptus Paula Couto (Mammalia: Xenarthra: Cingulata): the oldest record of the genus and morphological and phylogenetic aspects. Zootaxa 3721 (4), 2013, S. 387–398
- Juan Carlos Fernicola, Néstor Toledo, M. Susana Bargo und Sergio F. Vizcaíno: A neomorphic ossification of the nasal cartilages and the structure of paranasal sinus system of the glyptodont Neosclerocalyptus Paula Couto 1957 (Mammalia, Xenarthra). Palaeontologia Electronica 15 (3), 2012, 27A (online)
- Oscar Carranza-Castañeda und Wade E. Miller: Later Tertiary terrestrial mammals from Central Mexico and their relationship to South American immigrants. Revista Brasileira de Paleontologia 7 (2), 2004, S. 249–261
- H. Gregory McDonald: Paleoecology of extinct Xenarthrans and the Great American Biotic Interchange. Bulletin of the Florida Museum of Natural History 45 (4), 2005, S. 313–333
- Nicholas J. Czaplewski: A glyptodont (Mammalia: Xenarthra) from northern Oklahoma. Oklahoma Geology 64, 2004, S. 4–10
- Édison V. Oliveira, Recife, Kleberson O. Porpino, Mossoró und Alcina F. Barreto: On the presence of Glyptotherium in the Late Pleistocene of Northeastern Brazil, and the status of “Glyptodon” and “Chlamydotherium”. Paleobiogeographic implications. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 258 (3), 2010, S. 353–363
- Mário André Trindade Dantas, Lucas de Melo França, Mario Alberto Cozzuol und Ascanio Daniel Rincón: About the occurrence of Glyptodon sp. in the Brazilian intertropical region. Quaternary International 305, 2013, S. 206–208
- Anthony D. Barnosky und Emily L. Lindsey: Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. Quaternary International 217, 2010, S. 10–29
- Gustavo G. Politis und María A. Gutiérrez: Gliptodontes y cazadores-recolectores de la Región Pampeana (Argentina). Latin American Antiquity 9, 1998, S. 111–134
- Marína A. Gutiérrez und Gustavo A. Martínez: Trends in the faunal human exploitation during the Late Pleistocene and Early Holocene in the Pampean region (Argentina). Quaternary International 191, 2008, S. 53–68
- Thomas Falkner: A Description of Patagonia and the Adjoining Parts of South America. London, 1774, S. 1–144 (S. 54–55) ()
- Georges Cuvier: Recherches sur les ossemens fossiles, où l'on rétablit les caractères de plusieurs animaux dont les révolutions du globe ont détruit les espèces. Tome Cinquième. Paris, 1823, S. 1–405 (S. 174–192 online)
- Christian Samuel Weiss: Über das südliche Ende des Gebirgszuges von Brasilien in der Provinz S. Pedro do Sul und der Banda oriental oder dem Staate von Monte Video; nach den Sammlungen des Herrn Fr. Sellow. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin, 1827, S. 217–293 (S. 278–293) ()
- William Clift: Some account of the remains of the Megatherium sent to England from Buenos Ayres by Woodbine Parish. Transactions of the Geological Society 2 (3), 1835, S. 437–450 ()
- Joseph Eduard d'Alton: Über die von dem verstorbenen Herrn Sellow aus der Banda oriental mitgebrachten fossilen Panzerfragmente und die dazugehörigen Knochen-Überreste. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin, 1833, S. 369–424 ()
- Heinrich Georg Bronn: Lethaea geognostica: oder, Abbildungen und Beschreibung der für die Gebirgs-Formationen bezeichnendsten Versteinerungen, Band 2. Stuttgart, 1838, S. 1–1346 (S. 1256–1259) ()
- Fariña, Vizcaíno und de Iuliis 2013. S. 31–72
- Richard Owen: Note on the Glyptodon. In: Woodbine Parish (Hrsg.): Buenos Ayres, and the provinces of the Rio de La Plata: their present state, trade and dept; with some account from original documents of the progress of geographical discovery in those parts of South America during the last sixty years. London, 1839, S. 178b–178e ()
- Richard Owen: Description of a tooth an part of the skeleton of the Glyptodon clavipes, a large quadrupede of the Edentate order, to which belongs the tesselated bony armour described and figured by Mr. Clift in the former Volume of the Transactions of the Geological Society; with a consideration of the question whether the Megatherium possessed an analogous dermal armor. Transactions of the Geological Society 2 (6), 1841, S. 81–106 ()
- Juan Carlos Fernicola, Sergio F. Vizcaíno und Gerardo De Iuliis: The fossil mammals collected by Charles Darwin in South America during his travels on board the HMS Beagle. Revista de la Asociación Geológica Argentina 64 (1), 2009, S. 147–159
- Peter Wilhelm Lund: Blik paa Brasiliens Dyreverden för Sidste Jordomvaeltning. Anden Afhandling: Patteedyrene. Det Kongelige Danske Videnskabernes Selskbas Naturvidenskabelige og Matematiske Afhandlinger 8, 1838, S. 61–144 (S. 83) ()
- Cástor Cartelle: Peter W. Lund, a naturalist of several sciences. Lundiana 3 (2), 2002, S. 83–85
- Hermann Burmeister: Fauna argentina. I. Mamíferos fósiles. Lista de los mamíferos fósiles del terreno diluviano. Anales del Museo Publico de Buenos Aires 1, 1866–1867, S. 87–300 (S. 183–185) ()
- John Edward Gray: Catalogue of carnivorous, pachydermatous and edentate Mammalia in the British Museum. Taylor und Francis, London, 1869, S. 1–398 (S. 387) ()
- Barnum Brown: Brachyostracon, a New Genus of Glyptodonts from Mexico. American Museum of Natural History Bulletin 31 (17), 1912, S. 167–177
- Henry Fairfield Osborn: Glyptotherium texanum, a New Glyptodont, from the Lower Pleistocene of Texas. Bulletin of the American Museum of Natural History 19 (17), 1903, S. 491–494
- Florentino Ameghino: Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias de la República Argentina 6, 1889, S. 1–1027 (S. 758–852) ()
- Fariña, Vizcaíno und de Iuliis 2013. S. 183–186
Weblinks
- The Paleobiology Database Glyptodontidae