Pachyarmatheriidae
Die Pachyarmatheriidae sind eine ausgestorbene Familie aus der Ordnung der Gepanzerten Nebengelenktiere, zu denen auch die heute noch bestehenden Gürteltiere gehören. Sie kam vom Mittleren Miozän vor rund 14 Millionen Jahren bis zum ausgehenden Pleistozän vor rund 10.000 Jahren vor. Ihr Hauptverbreitungsgebiet umfasste den zentralen und nördlichen Teil Südamerikas und den südlichen Teil Nordamerikas und damit Landschaften unter warm-feuchten Klimabedingungen. Die Tiere ähnelten äußerlich heutigen Gürteltieren und besaßen auch in etwa deren Ausmaße. Ein wichtiger Unterschied ist die Gestaltung des Rückenpanzers. Dieser bestand aus zwei starren Teilen im Bereich des Schulter- und Beckengürtels, die sich etwa oberhalb der Körpermitte scharnierartig überlappten. Dadurch fehlten die für die Gürteltiere charakteristischen beweglichen Bänder zwischen den festen Panzerabschnitten. Typisch sind auch die sehr dicken Knochenplättchen, aus denen der Panzer besteht. Der Skelettbau verweist darauf, dass die Vertreter der Familie eher generalisierte Gräber darstellten, die vermutlich hauptsächlich Ameisen und Termiten fraßen. Die Familie wurde im Jahr 2018 wissenschaftlich eingeführt und besteht momentan aus zwei Gattungen. Vermutlich ist sie mit den Glyptodontidae und den Pampatheriidae näher verwandt.
| Pachyarmatheriidae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Zeitliches Auftreten | ||||||||||||
| Mittleres Miozän bis Unteres Holozän | ||||||||||||
| 14 Mio. Jahre bis 10.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Pachyarmatheriidae | ||||||||||||
| Fernicola, Rinderknecht, Jones, Vizcaíno & Porpino, 2018 | ||||||||||||
Merkmale
Allgemein und Körpergröße
Die Vertreter der Pachyarmatheriidae ähnelten im äußeren Erscheinungsbild möglicherweise eher den heutigen Gürteltieren als den Glyptodonten. Ihre Körpergröße entsprach weitgehend dem eines rezenten Langnasengürteltiers wie dem Neunbinden-Gürteltier. Die Kopf-Rumpf-Länge wird auf 50 bis 60 cm geschätzt, das Gewicht dürfte rund 10 bis 15 kg betragen haben.[1]
Skelettmerkmale
Skelettfunde von Pachyarmatherien sind bis heute relativ rar. Dies trifft auch auf den Schädel zu, von dem nur einzelne Bruchstücke als wahrscheinlich zu diesen Gepanzerten Nebengelenktieren gehörig betrachtet werden. Diese zeigen an, dass am Hinterhauptsbein offensichtlich ein mondsichelförmiger Wulst auftrat, der sich durch entsprechende Bildungen bei den Langnasengürteltieren wiederfindet. Der Unterkiefer besaß einen zierlichen und klingenförmigen Bau. Abweichend von allen anderen Vertretern der Gepanzerten Nebengelenktiere bestanden keine Alveolen für die Zähne, so dass er höchstwahrscheinlich zahnlos war. Dieses Merkmal ist typisch für die heutigen Ameisenbären als verwandte Gruppe. Die Symphyse am vorderen Ende zur Gelenkung der beiden Unterkieferhälften war nicht verwachsen und deutlich reduziert. Dies entspricht wiederum weitgehend den Langnasengürteltieren. Übereinstimmend mit diesen und im Unterschied zu den Ameisenbären stieg der Kronenfortsatz weit auf und lag deutlich oberhalb des Gelenkfortsatzes. Letzterer stand nur wenig oberhalb des horizontalen Knochenkörpers des Unterkiefers. Ameisenbären hingegen haben stark zurückgebildete Fortsätze des aufsteigenden Unterkieferastes.[2]
Das postcraniale Skelett ist ebenfalls nur bruchstückhaft überliefert. Die bekannten Brust- und Lendenwirbel waren mit wenigen Ausnahmen, die zu Neoglyptatelus gehören, frei und nicht verwachsen, fusionierte Wirbel hingegen sind ein Merkmal der Glyptodonten. Der Oberarmknochen war massiv und mit einer zwar kurzen, aber kräftigen deltopectoralen Leiste am Schaft ausgestattet. An der Elle trat ein extrem großer oberer Gelenkfortsatz auf (Olecranon), das fast die Länge des übrigen Knochens erreichte und entsprechende Bildungen bei den Gürteltieren übertrifft. Demnach besaß ein insgesamt 11,5 cm langes Exemplar ein 5,7 cm langes Olecranon. Den Oberschenkelknochen zeichnete ein deutlicher dritter Rollhügel etwa auf mittlerer Schafthöhe aus. Der Große Rollhügel überragte den Gelenkkopf, der wiederum auf einem nur schwach ausgeprägten Hals saß. Das untere Gelenkende wies keine seitlichen Verbreiterungen auf, übereinstimmend mit den Glyptodonten und Pampatherien. Schien- und Wadenbein waren an den Enden miteinander verwachsen. Außerdem war ein massiver äußerer Knöchel ausgebildet. Der vollständige bekannte Hinterfuß wies fünf Strahlen auf, von denen der innerste und äußerste am kürzesten, der zweite und dritte am längsten waren. Dadurch fehlt eine Hervorhebung des Mittelstrahls, der bei vielen grabenden Tieren wie bei einigen Gürteltieren besonders kräftig ausgebildet ist. Die Endglieder der Zehen sowie einzelne aufgefundene der Finger hatten alle eine gestreckte Form. Somit verweisen sie auf einst ausgebildete Krallen, die an den Händen möglicherweise schmaler als an den Füßen waren. Merkliche Größenunterschiede zwischen den einzelnen Strahlen traten nicht auf. Die Ausbildung von Krallen stimmt mit den Gürteltieren überein und weicht von den Glyptodonten mit ihren hufartigen Bildungen ab. Im Unterschied zu den Gürteltieren endeten die letzten Zehenglieder bei den Pachyarmatherien aber deutlich stumpfer.[2][3][4]
Panzer

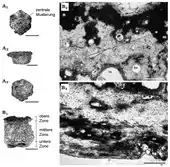
Wie alle Gepanzerten Nebengelenktiere zeichneten sich auch die Pachyarmatherien durch eine Panzerung des Rückens und zusätzlich auch des Schwanzes aus. Ein vollständig überlieferter Rückenpanzer von Neoglyptatelus war insgesamt 55 cm lang und über der Mitte 30 cm hoch. Am vorderen und hinteren Ende betrug die Höhe 12 beziehungsweise 14 cm. Der Rückenpanzer oder Carapax setzte sich aus einen jeweils festen Schulter- und Rückenschild zusammen, die sich in der Mitte leicht überlappten. Dies ist ein bedeutender Unterschied zu den Panzern der Gürteltiere und Pampatherien, bei denen zwischen den festen Panzerabschnitten eine unterschiedliche Anzahl an beweglichen Bändern ausgebildet ist, oder zu dem der Glyptodonten mit ihrem insgesamt starren Panzer. Aufgebaut war der Panzer aus einzelnen kleinen Knochenplättchen oder Osteodermen von häufig sechseckiger Gestalt. Ihre Größe variierte meist zwischen 0,8 und 3,2 cm in der Länge, zwischen 0,9 und 2,9 cm in der Breite und zwischen 0,5 und 1,8 cm in der Dicke. Die größten Osteoderme lagen im Bereich der Rückenlinie. Auf der Oberfläche war häufig eine zentrale Musterung von polygonaler bis rundlicher Gestalt ausgebildet, die etwa die Hälfte der Größe des gesamten Knochenplättchens einnahm und sich dezentral verschoben befand. Umgeben wurde diese zentrale Musterung von einer variierenden Anzahl an kleineren randlichen Mustern. Den unteren Rand des Schulter- und Beckenschildes bildeten eins bis zwei Reihen an Osteodermen, von denen die der äußersten einen fünfeckigen Umriss aufwiesen und keine Musterung trugen. Davon wichen die äußersten Reihen am hinteren Ende des Schulter- und am vorderen Ende des Beckenschildes ab, die die Artikulationsfläche der beiden Panzerteile bildeten. Diese verfügte einerseits über eine konkav-konvex gewellte Oberfläche der Unterseite des Schulterschildes, die zu einer ähnlich geformten Oberfläche der Oberseite des Beckenschildes korrespondiert. Dadurch konnte der Schulterschild den Beckenschild überlappen und es entstand so eine scharnierartige Kontaktzone zwischen beiden, die gewisse Bewegungen der Panzerabschnitte zueinander zuließ.[3][5][4]
Der Schwanz war ebenfalls von einem Panzer umhüllt. Im vorderen Bereich bestand er jeweils aus einzelnen Ringen aus zwei Reihen von Osteodermen. Jeder Ring umschloss einen Schwanzwirbel. Die Größe der Ringe und der eingebundenen Knochenplättchen nahm zum hinteren Schwanzbereich ab. An der Schwanzspitze waren keine Ringe mehr ausgebildet. Hier formten die Knochenplättchen eine Art Röhre. Alle Knochenplättchen des Schwanzpanzers wiesen eine glatte Oberfläche ohne Musterung auf.[6][4]
Im Querschnitt wies jedes Knochenplättchen analog zu den Bildungen bei den Gürteltieren, Glyptodonten und Pampatherien mehrere Lagen auf. Hierbei formten eine feste Knochenschicht den oberen und unteren Abschluss. In der oberen Knochenschicht ließen sich Wachstumszonen feststellen, auch durchzogen sie einzelne Kanälchen, in denen wohl Haarfollikel saßen. Die untere Knochenschicht setzte sich aus kompakten Kollagenbündeln zusammen. Zwischen beiden dehnte sich ein Bereich mit spongioser Struktur aus, die aber abweichend von den Glyptodonten eher schwach entwickelt war.[5][7]
Verbreitung
Die Pachyarmatheriidae sind sowohl aus Südamerika als auch aus Mittel- und Nordamerika überliefert. Die südamerikanischen Fundgebiete erstrecken sich von Uruguay im Süden nach Norden über Brasilien und Peru bis nach Kolumbien und Venezuela. In Mittelamerika stammen Nachweise aus Costa Rica, in Nordamerika wurden Reste der Tiere bisher vor allem im südöstlichen Teil des Kontinentes in Florida und in South Carolina entdeckt. Die zeitliche Tiefe reicht vom Mittleren Miozän vor rund 14 Millionen Jahren bis in den Übergang vom Pleistozän zum Holozän vor rund 10.000 Jahren.[4]
Paläobiologie

Neben der generell gürteltierartigen äußeren Erscheinung der Pachyarmatherien finden sich am Skelett einzelne Hinweise auf eine grabende Lebensweise. Der eindeutigste ist der ausgedehnte Fortsatz des oberen Gelenkendes (Olecranon) der Elle, der fast die Länge des übrigen Knochens einnimmt. Ähnliche Ausmaße weist das entsprechende Vorderbeinelement beim Sechsbinden-Gürteltier auf. In der Regel erhöht ein langes Olecranon die Hebelkraft des Unterarms. Dem gegenüber fehlen an Hand und Fuß ein deutlich verlängerter Mittelstrahl, der häufig bei grabenden Tieren vorkommt. Auch die zwar kräftigen, aber weitgehend stumpf endenden letzten Phalangen der Finger und Zehen sprechen gegen außerordentlich gut ausgebildete Grabeigenschaften. Vermutlich waren die Pachyarmatherien eher generalisierte Gräber, die überwiegend im Untergrund kratzten, vergleichbar dem heutigen Neunbinden-Gürteltier.[2] Die starren und sich überlappenden Panzerteile verhinderten eine deutliche Flexion des Körpers, anders als bei den Gürteltieren mit ihren beweglichen Bändern. Die scharnierartige Verbindung zwischen dem vorderen und hinteren Panzerabschnitt ließ eine gegengerichtete Bewegung auf 25 mm Länge zu. Noch stärker beschränkt waren Auf- und Abwärtsbewegungen des Körpers. Sie dürften aber nicht so deutlich begrenzt gewesen sein wie bei den Glyptodonten, deren fest verwachsener Panzer und die ebenso verwachsene Wirbelsäule kaum derartige Bewegungen unterstütze. Möglicherweise nahmen bei den Pachyarmatherien die Bewegungsbeschränkungen im Laufe der Stammesgeschichte zu, da frühe Vertreter wie Neoglyptatelus noch eine ausgedehntere Panzergelenkung zeigten als späte wie Pachyarmatherium.[4]
Der zahnlose, klingenförmige Unterkiefer unterscheidet sich deutlich von dem der Gürteltiere, die noch stiftartige Zähne aufweisen. In diesem Merkmal treten Übereinstimmungen zu den Ameisenbären auf, die eine stark auf Ameisen und Termiten spezialisierte Lebensweise haben (Myrmecophagie), wobei aufgrund fehlender Kauprozesse dieser besondere, zahnlose Unterkiefer entstand. Bei ihnen ist aber zusätzlich noch die Gelenkverbindung zum Schädel stark reduziert, was bei den Pachyarmatherien nicht der Fall war. Daraus kann geschlossen werden, dass die Vertreter der Pachyarmatheriidae eine deutlich myrmecophage Ernährungsweise verfolgten, die stärker entwickelt war als bei zahlreichen Gürteltieren, aber sich nicht so ausgeprägt erwies wie bei den Ameisenbären.[2]
An einzelnen Knochenplättchen der Pachyarmatherien lassen sich sekundäre Fraßspuren feststellen, die auf Parasitismus zurückgehen. Es handelt sich meist um löcherige Strukturen, wie sie heute bei den Gürteltieren durch Flöhe der Gattung Tsunga hervorgerufen werden. Andere strukturelle Oberflächenveränderungen sind auf das krankheitsbedingte Einwirken von Bakterien und Pilzen zurückzuführen. Ähnliches ist auch von ausgestorbenen Gürteltieren und den Glyptodonten belegt.[8][9]
Systematik
Äußere Systematik
Innere Systematik der Gepanzerten Nebengelenktiere nach Fernicola et al. 2018[4]
|
Die Pachyarmatheriidae stellen eine Familie innerhalb der Ordnung der Gepanzerten Nebengelenktiere (Cingulata) dar. Zu dieser einst formenreich in Süd- und Nordamerika verbreiteten Ordnung gehören des Weiteren auch die Glyptodontidae, Pampatheriidae, Peltephilidae und Palaeopeltidae, die allesamt ausgestorben sind. Als einzige rezente Vertreter haben bis heute die Gürteltiere (Dasypoda) überlebt. Die Gepanzerten Nebengelenktiere stehen wiederum in der Überordnung der Nebengelenktiere (Xenarthra), welche zusätzlich noch die Zahnarmen (Pilosa) mit den heutigen Faultieren (Folivora) und den Ameisenbären (Vermilingua) einschließt. Die Nebengelenktiere repräsentieren eine der vier Hauptlinien der Höheren Säugetiere, die den anderen drei (zusammengefasst als Epitheria) als Schwestergruppe gegenübersteht.[10] Eine generelle Gemeinsamkeit der Nebengelenktiere findet sich in den namengebenden xenarthrischen Gelenken (Nebengelenke, auch Xenarthrale) an den Gelenkfortsätzen der hinteren Brust- und der Lendenwirbel.[11] Der Ursprung der Nebengelenktiere ist bisher nicht geklärt. Fossil traten sie bereits im Paläozän vor mehr als 56 Millionen Jahren in Südamerika auf, die Funde werden den Gürteltieren zugerechnet.[12] Molekulargenetische Untersuchungen weisen die Abspaltung der Nebengelenktiere von den anderen Höheren Säugetieren bis in die ausgehende Unterkreide vor etwa 103 Millionen Jahren zurück. Die Gürteltiere als heute lebende, nächste Verwandte der Pachyarmatheriidae trennten sich von der gemeinsamen Linie mit den Zahnarmen zu Beginn des Paläozän vor etwa 65 Millionen Jahren ab.[13][14]
Die innere Gliederung der Gepanzerten Nebengelenktiere ist nicht vollständig geklärt. Den Pachyarmatherien kann aufgrund des Skelettbaus und der Panzergestaltung eine enge Beziehung zu den Glyptodonten und den Pampatherien zugesprochen werden, als deren Schwestergruppe sie anzunehmen sind. Die engen Verbindungen drücken sich unter anderem in einigen Einzelmerkmalen des Bewegungsapparates, etwa am Oberschenkelknochen, am Sprungbein und am Fersenbein aus. Hinzu kommen die fehlenden beweglichen Bänder und die dicken Knochenplättchen des Panzers. Allerdings ist das Fehlen beweglicher Panzerabschnitte nicht ganz eindeutig, da dies sowohl bei den Pachyarmatherien als auch bei den Glyptodonten ein abgeleitetes Merkmal sein kann. Die Glyptodonten und Pampatherien gelten innerhalb der Gepanzerten Nebengelenktiere als näher miteinander verwandt, dies wird unter anderem durch den Bau der Zähne und die Gestaltung des Ohres unterstützt. Beide werden daher zur übergeordneten Gruppe der Glyptodonta zusammengefasst. Nach schädelmorphologischen Untersuchungen wiederum sind die Glyptodonta tiefer in die Gürteltiere eingebettet und stehen in einer näheren Beziehung zu euphractinen Gürteltieren wie dem Sechsbinden-Gürteltier oder den Borstengürteltieren.[15][16] Die Einbettung in die Gürteltiere ließ sich auch molekulargenetisch bestätigen, im Unterschied zu den Ergebnissen der Skelettanatomie ergab sich hier jedoch eine engere Bindung an die tolypeutinen (Kugelgürteltiere und Nacktschwanzgürteltiere) und chlamyphorinen (Gürteltmulle) Gürteltiere.[17][18] Die Pachyarmatherien stünden somit als Schwesterlinie der Glyptodonta ebenfalls den Gürteltieren sehr nahe.[3][4]
Innere Systematik
Als eigenständige Familie wurden die Pachyarmatheriidae im Jahr 2018 von Juan C. Fernicola und Forscherkollegen eingeführt. Ihr gingen die Überlegungen voraus, dass sich Pachyarmatherium und Neoglyptatelus durch ihre typische Gestaltung der Osteoderme deutlich näher stehen als ursprünglich angenommen. Pachyarmatherium war 1995 durch Kevin F. Downing und Richard S. White anhand von Funden aus der Leisey Shell Pit im Hillsborough County und Fossillagerstätte Haile 16A im Alachua County, beide im US-Bundesstaat Florida gelegen, definiert worden. Die Autoren stellten ihre neue Gattung damals in die nähere Gürteltierverwandtschaft.[2] Die Benennung von Neoglyptatelus geht wiederum auf Alfredo A. Carlini und Kollegen zurück und basiert auf Fundmaterial aus La Venta in Kolumbien. Das Arbeitsteam um Carlini befürwortete eine sehr basale Stellung der neuen Form innerhalb der Glyptodonten und verband sie mit der urtümlichen Unterfamilie der Glyptatelinae, als deren stammesgeschichtlich jüngstes Mitglied Neoglyptatelus angesehen wurde.[19] Dass sowohl Pachyarmatherium als auch Neoglyptatelus eng verwandt sind, vermutete erstmals Sergio F. Vizcaíno im Jahr 2003,[20] ähnlich äußerte sich sechs Jahre später Kleberson de Oliveira Porpino.[3] Allerdings variierte in der Folgezeit vor allem für Pachyarmatherium die systematische Stellung je nach Autor zwischen den Gepanzerten Nebengelenktieren mit unsicherer Stellung,[3] den Glyptodonten[21][22] oder den Gürteltieren.[5] Fernicola und Kollegen vereinigten dann beide Gattungen unter der Familie der Pachyarmatheriidae. Der Familienname bezieht sich auf die Gattung Pachyarmatherium als Nominatform. Deren wissenschaftliche Bezeichnung geht wiederum auf die griechische Sprache zurück und bedeutet so viel wie παχύ- (pachy-) für „dick“, άρμα (arma) für „Streitwagen“ und θηρίον (thērion) für „Tier“. Der Name bezieht sich somit auf die dicke Panzerung.[2][4]
Die Familie setzt sich zur Zeit aus zwei Gattungen zusammen:[4]
- Familie: Pachyarmatheriidae Fernicola, Rinderknecht, Jones, Vizcaíno & Porpino, 2018
- Neoglyptatelus Carlini, Vizcaíno & Scillato-Yané , 1997
- Pachyarmatherium Downing & White, 1995
Es besteht die Überlegung, dass die als urtümliche Glyptodonten eingestuften übrigen Mitglieder der Glyptatelinae, namentlich Glyptatelus und Clypeotherium, ebenfalls zu den Pachyarmatherien gehören. Beide Taxa datieren in das Eozän und basieren auf nur einigen wenigen Osteodermen, die wie bei den Pachyarmatherien relativ dick sind und eine dezentrale Musterung aufweisen. Histologische Untersuchungen zeigen aber eine stärkere Ähnlichkeit zu den Panzerelementen der Glyptodonten.[4]
Stammesgeschichte
Der Ursprung der Pachyarmatheriidae ist weitgehend unbekannt. Erstmals fassbar ist die Gruppe im Mittleren Miozän vor rund 14 Millionen Jahren. Hierbei handelt es sich um Neoglyptatelus, das in La Venta aufgefunden wurde, einer bedeutenden Fossillagerstätte am Mittellauf des Río Magdalena in Kolumbien. Das hier geborgene Fossilmaterial umfasst zahlreiche einzelne Osteoderme und den Rest der Schwanzpanzerung.[19] Die Landschaft, in der dieser früheste Vertreter lebte, bestand aus Wäldern, die sich bis in Überschwemmungsebenene ausdehnten und unter dem Einfluss von warm-feuchten Klimaten wuchsen. Möglicherweise in das Obere Miozän gehören Teile der Panzerung des Rückens und des Schwanzes aus der Sincelejo-Formation in der Küstenregion von Kolumbien.[6] Bereits zu diesem Zeitpunkt hatte sich die Gattung sehr weit in den Süden des Kontinentes ausgebreitet und ist unter anderem wiederum mit Knochenplättchen aus erodierten Sanden der Camacho-Formation im Mündungsgebiet des Río de la Plata in Uruguay belegt. Die gleiche Gesteinseinheit brachte auch einen der bisher vollständigsten Funde zu Tage: ein schädelloses Teilskelett mit nahezu komplettem Rückenpanzer und der Schwanzpanzerung. Die Ablagerungen der Camacho-Formation entstanden in einer ähnlichen Landschaft wie sie für La Venta rekonstruiert wird. Ihr Alter wird mit 8 bis 6,8 Millionen Jahren angegeben.[20][4] Ebenso stammen Reste von Neoglyptatelus aus dem gleichen Zeitraum aus dem südwestlichen Amazonasbecken von Brasilien.[23]
Die Gattung Pachyarmatherium lässt sich erstmals im Übergang vom Pliozän zum Pleistozän in Südamerika belegen. Allein 184 Knochenplättchen wurden in den Asphaltgruben von El Breal de Orocual im nordöstlichen Venezuela aufgedeckt. Die einzelnen Gruben der Region entstanden zu unterschiedlichen Zeiten. Dadurch ist die Form dort auch noch bis in das ausgehende Pleistozän über mehr als drei Dutzend Osteoderme nachgewiesen.[24][25] Zusätzlich sind Fossilreste von Pachyarmatherium aus dem nördlichen Venezuela dokumentiert, so aus den Karsthöhlen von Cerro Misión im Bundesstaat Falcón,[26] ebenso wie aus dem nordöstlichen Peru.[27] Auf die jüngsten Funde der Gattung und gleichzeitig der gesamten Familie verweisen einige Lokalitäten im nordöstlichen Brasilien. Während die Osteoderme der Fazenda Nova im Bundesstaat Pernambuco laut radiometrischen Daten noch zwischen 59.000 und 64.000 Jahre alt sind,[22][5] gehören die Skelett- und Panzerreste von Lajedo da Escada im Bundesstaat Rio Grande do Norte wahrscheinlich in den Übergang vom Pleistozän zum Holozän.[3]
Wie viele andere, ursprünglich in Südamerika endemische Säugetiere gelangten auch die Pachyarmatheriidae im Zuge des Großen Amerikanischen Faunenaustausches nach Nordamerika. Dieser begann im Pliozän vor rund 3,5 Millionen Jahren, als sich am Isthmus von Panama eine Landbrücke zwischen den beiden Kontinentalmassen bildete. Ins Obere Pliozän und Untere Pleistozän gehören einzelne Knochenplättchen von Pachyarmatherium aus Costa Rica, etwa aus der Region um Buenos Aires del cantón de Palmares.[21][28][29] Das bedeutendste Material wurde aber im südöstlichen Teil der USA geborgen. Herausragend sind dabei die Leisey Shell Pit im Hillsborough County von Florida wie auch die Fundstelle Haile 16A im Alachua County, ebenfalls Florida. Beide Lokalitäten bargen jeweils mehrere hundert Osteoderme und Panzerreste, letztere enthielt zusätzlich noch einige Elemente des Bewegungsapparates. Alle Funde werden ebenfalls zu Pachyarmatherium gestellt. Hinzu kommen noch einzelne weitere kleinere Fundplätze wie etwa der Kissimmee River im Okeechobee County in Florida, der zur Leisey Shell Pit benachbarte Apollo Beach und mehrere Stücke aus dem Dorchester County in South Carolina. Für alle genannten Fundstellen besteht eine Alterseinstufung im Pliozän-Pleistozän-Übergangsbereich.[2]
Literatur
- Juan C. Fernicola, Andrés Rinderknecht, Washington Jones, Sergio F. Vizcaíno und Kleberson de Oliveira Porpino: A new species of Neoglyptatelus (Mammalia, Xenarthra, Cingulata) from the late Miocene of Uruguay provides new insights on the evolution of the dorsal armor in cingulates. Ameghiniana 55, 2018, S. 233–252
Einzelnachweise
- Darin A. Croft und Velizar Simeonovski: Horned armadillos and rafting monkeys. The fascinating fossil mammals of South Amerika. Indiana University Press, 2016, S. 1–304 (S. 180–181)
- Kevin F. Downing und Richard S. White: The cingulates (Xenarthra) of the Leisey Shell Pit local fauna (Irvingtonian), Hillborough County, Florida. Bulletin of the Florida Museum of Natural History 37 (Part II), 1995, S. 375–396
- Kleberson de Oliveira Porpino, Juan C. Fernicola und Lílian P. Bergqvist: A New Cingulate (Mammalia: Xenarthra), Pachyarmatherium brasiliensesp. nov., from the Late Pleistocene of Northeastern Brazil. Journal of Vertebrate Paleontology 29 (3), 2009, S. 881–893
- Juan C. Fernicola, Andrés Rinderknecht, Washington Jones, Sergio F. Vizcaíno und Kleberson de Oliveira Porpino: A new species of Neoglyptatelus (Mammalia, Xenarthra, Cingulata) from the late Miocene of Uruguay provides new insights on the evolution of the dorsal armor in cingulates. Ameghiniana 55, 2018, S. 233–252
- Édison Vicente Oliveira, Kleberson de Oliveira Porpino und Fabiana Marinho da Silva: New material of Pachyarmatherium from the late Pleistocene of northeastern Brazil: insights into its morphology and systematics. Paläontologische Zeitschrift 87, 2013, S. 505–513, doi:10.1007/s12542-013-0166-4
- Carlos Villarroel und Jairo Clavijo: Los mamíferos fósiles y las edades de las sedimentitas continentales del Neógeno de la Costa Caribe Colombiana. Revista de la Academia Colombiana de Ciencias 29 (112), 2005, S. 345–356
- Paulo Victor Luiz Gomes Da Costa Pereira, Gustavo Duarte Victer, Kleberson de Oliveira Porpino und Lílian Paglarelli Bergqvist: Osteoderm histology of Late Pleistocene cingulates from the intertropical region of Brazil. Acta Palaeontologica Polonica 59 (3), 2014, S. 543–552
- Rodrigo L. Tomassini, Claudia I. Montalvo und María C. Ezquiaga: The oldest record of flea/armadillos interaction as example of bioerosion on osteoderms from the late Miocene of the Argentine Pampas. International Journal of Paleopathology 15, 2016, S. 65–68, doi:10.1016/j.ijpp.2016.08.004
- Fábio Cunha Guimarães de Lima und Kleberson de Oliveira Porpino: Ectoparasitism and infections in the exoskeletons of large fossil cingulates. PLoS ONE 13 (10), 2018, S. e0205656, doi:10.1371/journal.pone.0205656
- Maureen A. O'Leary, Jonathan I. Bloch, John J. Flynn, Timothy J. Gaudin, Andres Giallombardo, Norberto P. Giannini, Suzann L. Goldberg, Brian P. Kraatz, Zhe-Xi Luo, Jin Meng, Xijun Ni, Michael J. Novacek, Fernando A. Perini, Zachary S. Randall, Guillermo W. Rougier, Eric J. Sargis, Mary T. Silcox, Nancy B. Simmons, Michelle Spaulding, Paúl M. Velazco, Marcelo Weksler, John R. Wible und Andrea L. Cirranello: The Placental Mammal Ancestor and the Post–K-Pg Radiation of Placentals. Science 339, 2013, S. 662–667, doi:10.1126/science.1229237
- Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, S. 1–431 (S. 200–204)
- Sergio F. Vizcaíno und W. J. Loughry: Xenarthran biology: Past, present and future. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 1–7
- Frédéric Delsuc, Michael J. Stanhope und Emmanuel J.P. Douzery: Molecular systematics of armadillos (Xenarthra, Dasypodidae): contribution of maximum likelihood and Bayesian analyses of mitochondrial and nuclear genes. Molecular Phylogenetics and Evolution 28, 2003, S. 261–275
- Frédéric Delsuc, Sergio F Vizcaíno und Emmanuel J. P. Douzery: Influence of Tertiary paleoenvironmental changes on the diversification of South American mammals: a relaxed molecular clock study within xenarthrans. BMC Evolutionary Biology 4 (11), 2004, S. 1–13
- Timothy J. Gaudin und John R. Wible: The Phylogeny of Living and Extinct Armadillos (Mammalia, Xenarthra, Cingulata): A Craniodental Analysis. In: M. T. Carrano, T. J. Gaudin, R. W. Blob und J. R. Wible (Hrsg.): Amniote Paleobiology. Chicago/London: University of Chicago Press, 2006, S. 153–198
- Guillaume Billet, Lionel Hautier, Christian de Muizon and Xavier Valentin: Oldest cingulate skulls provide congruence between morphological and molecular scenarios of armadillo evolution. Proceedings of the Royal Society B, 278, 2011, S. 2791–2797
- Frédéric Delsuc, Gillian C. Gibb, Melanie Kuch, Guillaume Billet, Lionel Hautier, John Southon, Jean-Marie Rouillard, Juan Carlos Fernicola, Sergio F. Vizcaíno, Ross D. E. MacPhee und Hendrik N. Poinar: The phylogenetic affinities of the extinct glyptodonts. Current Biology 26, 2016, S. R155–R156, doi:10.1016/j.cub.2016.01.039
- Kieren J. Mitchell, Agustin Scanferla, Esteban Soibelzon, Ricardo Bonini, Javier Ochoa und Alan Cooper: Ancient DNA from the extinct South American giant glyptodont Doedicurus sp. (Xenarthra: Glyptodontidae) reveals that glyptodonts evolved from Eocene armadillos. Molecular Ecology 25 (14), 2016, S. 3499–3508, doi:10.1111/mec.13695
- Alfredo A. Carlini, Sergio F. Vizcaíno und Gustavo J. Scillato-Yané: Armored xenarthrans: a unique taxonomic and ecologic assemblage. In: Richard F. Kay, Richard H. Madden, Richard L. Cifelli und John J. Flynn (Hrsg.): Vertebrate Paleontology in the Neotropis. The Miocene fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, 1997, S. 213–226
- Sergio F. Vizcaíno, André S. Rinderknecht und Ada Czerwonogora: An enigmatic Cingulata (Mammal ia: Xenarthra) from the Late Miocene of Uruguay. Journal of Vertebrate Paleontology 23 (4), 2003, S. 981–983
- César Laurito Mora, Ana Lucía Valerio Zamora und Eduardo Antonio Pérez Gamboa: Los xenarthras fósiles de la localidad de Buenos Aires de Palmares (Blancano tardío-Irvingtoniano temprano), provincia de Alajuela, Costa Rica. Revista Geológica de América Central 33, 2005, S. 83–90
- Édison Vicente Oliveira, Alcina M. Franca Barreto und Rosemberg da Silva Alves: Aspectos sistemáticos, paleobiogeográficos e paleoclimáticos dos mamíferos quaternários de Fazenda Nova, PE, nordeste do Brasil. Gaea Journal of Geoscience 5, 2009, S. 75–85
- Mario Alberto Cozzuol: The Acre vertebrate fauna: Age, diversity, and geography. Journal of South American Earth Sciences 21, 2006, S. 185–203
- Ascanio D. Rincón, Gilberto E. Parra, Francisco J. Prevosti, Maria Teresa Alberdi und Christopher J. Bell: A preliminary assessment of the mammalian fauna from the Pliocene-Pleistocene El Breal de Orocual locality, Monagas State, Venezuela. Museum of Northern Arizona Bulletin 65, 2009, S. 593–620
- Andrés Solórzano, Ascanio D. Rincón und H. Gregory McDonald: A New Mammal Assemblage from the Late Pleistocene El Breal de Orocual, Northeast of Venezuela. Science Series 42, 2015, S. 125–150
- Ascanio D. Rincón und Richard White: Los Xenarthra Cingulata del Pleistoceno tardío (Lujanense) de Cerro Misión, Estado Falcón, Venezuela. Boletín de la Sociedad Venezolana de Espeleología 41, 2007, S. 2–12
- Jean-Noël Martinez und Ascanio Daniel Rincón: Los Xenarthra Cingulata del noroeste del Perú. Resúmenes extendidos del XV Congreso Peruano de Geología, Publicación Especial de la Sociedad Geológica del Perú, Actas 9, 2010, S. 432–435
- Ana L. Valerio und César A. Laurito: Nuevos hallazgos de Mammalia, Xenarthra (Cingulata) y confirmación del registro de Pachyarmatherium leiseyi Downing & White, 1995 en la localidad de Buenos Aires de Palmares, provincia de Alajuela, Costa Rica. Revista Geológica de América Central 44, 1911, S. 131–139
- César A. Laurito und Ana L. Valerio: Paleobiogeografía del arribo de mamíferos suramericanos al sur de América Central de previo al Gran Intercambio Biótico Americano: un vistazo al GABI en América Central. Revista Geológica de América Central 46, 2012, S. 123–144