Neunbinden-Gürteltier
Das Neunbinden-Gürteltier (Dasypus novemcinctus) ist eine Säugetierart aus der Gruppe der Gürteltiere (Dasypoda). Es zählt zu den bekanntesten und weitestverbreiteten Vertretern dieser Gruppe, zudem ist es der am besten erforschte. Das Vorkommen dieser Gürteltierart beschränkt sich nicht nur auf weite Teile Südamerikas, als einzige heute noch lebende Art ist es auch in Nordamerika verbreitet, wo es erstmals in den 1850er Jahren beobachtet wurde. Die Lebensräume, die das Neunbinden-Gürteltier besiedelt, sind sehr vielfältig und umfassen hauptsächlich Wälder und teils offene Landschaften, die aber mehr oder weniger gut durchfeuchtet sein müssen; zudem gilt es als äußerst anpassungsfähig, jedoch kommt es mit längeren Kälteperioden nicht zurecht. Die Tiere leben einzelgängerisch und graben unterirdische Baue. Sie paaren sich einmal im Jahr. Die üblicherweise vier Neugeborenen stellen meist vier genetisch identische Jungtiere dar und verbleiben bis zu neun Monate beim Muttertier. Auch in der Ernährung ist die Gürteltierart sehr anpassungsfähig und verzehrt hauptsächlich Insekten, aber auch kleinere Wirbeltiere. Aufgrund der weiten Verbreitung werden bis zu sieben Unterarten unterschieden. Bedeutend in der Beziehung zum Menschen ist der Umstand, dass das Neunbinden-Gürteltier Erreger der Lepra und der Chagas-Krankheit trägt und so häufig in der Medizin als Labortier verwendet wird. In einigen Ländern seines Verbreitungsgebietes gilt es auch als Nahrungsressource. Das Neunbinden-Gürteltier wird nicht als gefährdet eingestuft.
| Neunbinden-Gürteltier | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Neunbinden-Gürteltier | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Dasypus novemcinctus | ||||||||||||
| Linnaeus, 1758 |
Merkmale
Habitus

Das Neunbinden-Gürteltier erreicht eine Kopf-Rumpf-Länge von 32 bis 57 cm, der Schwanz ist 21 bis 45 cm lang und ist damit in der Regel etwa so lang wie der Rumpf oder teilweise länger. Das Gewicht variiert von 3 bis maximal 9,8 kg, wobei aber 6,5 kg eher selten überschritten werden; Männchen sind insgesamt etwas schwerer als Weibchen. Die Gürteltierart hat einen bis zu 10 cm langen und schmalen, dreieckig geformten Kopf mit einer lang ausgezogenen Schnauze. Die Ohren sind mit bis zu 5,9 cm Länge im Vergleich zum Kopf groß, sehr beweglich und weisen gerundete Enden auf. Der typische Kopfschild besteht aus großen Knochenschildchen, die dicht auf dem Skelettunterbau liegen. Wie alle Gürteltiere ist das Neunbinden-Gürteltier durch die Rückenpanzerung gekennzeichnet, die drei getrennte Abschnitte umfasst: einen festen Schulter- und Beckenteil sowie einen Teil bestehend aus 8 bis 10, meist jedoch 9 beweglichen Bändern dazwischen, die durch Hautfalten miteinander verbunden sind. Die festen Panzerteile bestehen aus je etwa 18 bis 20 Reihen kleiner, rundlich gestalteter Knochenplättchen. Durch die Überdeckung der Knochenplättchen mit kleinen Hornschildchen wirken diese aber nicht immer symmetrisch angeordnet. Die Knochenplättchen der beweglichen Bänder sind dagegen viereckig geformt. Sie sind mit einer dreieckigen Musterung mit nach hinten zeigender Spitze versehen. Das vierte Band weist 54 bis 65 derartige knöcherne Bildungen auf. Zusätzlich ist der lange Schwanz in seinen ersten beiden Dritteln von 12 bis 15 Knochenringen umgeben. Der Panzer besitzt meist eine braune Färbung, wobei die unteren Partien etwas heller sind, ebenso wie die vorderen beweglichen Bänder. Der Kopfschild ist ebenfalls heller getönt, während die Ohren einen sehr dunklen Farbton aufweisen. Der spärlich behaarte Bauch hat eine rosafarbene Tönung. Die Beine sind kurz, die Vorderfüße tragen vier und die Hinterfüße fünf Zehen, die alle mit scharfen Krallen versehen sind. Jene der zwei mittleren Vorderfußzehen sind am längsten. Der Hinterfuß erreicht eine Länge von durchschnittlich 8,7 cm. Weibliche Tiere besitzen vier Milchdrüsen. Das Tier verfügt außerdem über eine stark schwankende Körperkerntemperatur von 30 bis 36 °C, wobei diese mit durchschnittlich 33,4 °C bei Männchen höher ist als bei Weibchen mit 31,7 °C.[1][2][3]
Schädel- und Skelettmerkmale

Der Schädel ist bis zu 9,9 cm lang und an den Jochbeinbögen gut 4,3 cm breit. Der knöcherne Teil der Schnauze, das Rostrum, besitzt eine Länge von etwa 5,9 cm, es ist dadurch deutlich verlängert und zusätzlich schmal geformt. Der Unterkiefer kann 7,8 cm lang werden. Der Zahnbau unterscheidet sich von dem anderer Säugetiere und folgt nicht der gängigen Gebissformel. Die Zähne sind einfach gebaut und ohne Zahnschmelz, besitzen nur eine Wurzel und haben eine stiftartige Form. Sowohl im Ober- als auch im Unterkiefer sitzen jeweils je Kieferhälfte 7 bis 9 Zähne, also insgesamt 28 bis 36, im Durchschnitt sind es aber je Kieferbogen 8, insgesamt also 32.[1] Allerdings scheint es innerhalb der einzelnen Unterarten des Neunbinden-Gürteltiers deutliche Unterschiede zu geben, da bei Populationen aus Nicaragua nur jeweils 7 Zähne je Kieferbogen beobachtet wurden. Die Gesamtlänge der unteren Zahnreihe beträgt 2,6, die der oberen 2,4 cm.[4] An den Vordergliedmaßen weist die Ulna ein besonders großes oberes Gelenk auf (Olecranon), dass bei einer Gesamtlänge des Knochens von 6,9 cm etwa 2,7 cm erreicht. Solche großen Gelenkenden an den Vordergliedmaßen sind typisch für Tiere mit grabender Lebensweise.[5]
Sinnesleitungen und Lautäußerungen
Das Neunbinden-Gürteltier hat einen eher schlechten Sehsinn, dafür aber einen ausgesprochen guten Geruchssinn, den es vor allem bei der Nahrungssuche einsetzt. Mehrere Lautrufe sind bekannt. Grabende und sich wehrende Tiere geben ein keuchendes Grunzen von sich, ängstliche ein Quieken. In der Paarungszeit sind auch niederfrequente Kontaktrufe nachgewiesen. Zudem ist die Gürteltierart ein sehr geräuschvolles Tier, welches ein beständiges Schnauben oder Grunzen bei der Suche nach Futter ausstößt. Auch sonst verursacht es viel Lärm durch Zerbrechen trockener Äste oder Rascheln im Laub und gilt als einer der geräuschintensivsten Waldbewohner.[1]
Verbreitung
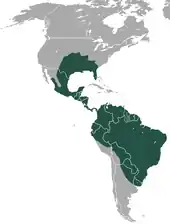
Allgemein
Das Verbreitungsgebiet des Neunbinden-Gürteltiers reicht von Nord- über Mittel- bis nach Südamerika und besitzt damit die größte Ausdehnung aller Gürteltierarten. Als einziger Vertreter der Gürteltiere ist es zudem in den USA anzutreffen, wo es vor allem den Süden und Südosten besiedelt. Die südliche Verbreitungsgrenze befindet sich im Norden Argentiniens und in Uruguay. Im Westen Südamerikas stellen die Hochflächen der Anden eine natürliche Grenze dar, vor allem in Peru und Bolivien. Darüber hinaus sind auch einzelne Populationen auf den Karibischen Inseln anzutreffen, namentlich Grenada und Trinidad und Tobago. Das gesamte bewohnte Gebiet umfasst 19,1 Millionen Quadratkilometer, die tatsächlich okkupierte Fläche ist aber unbekannt.[6][7][3]
Das Neunbinden-Gürteltier lebt von Meeresspiegelhöhe bis in Höhen um 2000 m. Es ist sehr anpassungsfähig und toleriert zahlreiche Habitate. So ist die Gürteltierart in feuchten Wäldern ebenso heimisch wie in offenem Grasland, auf landwirtschaftlich genutzten Flächen und in städtischen Gebieten. Generell ist eine Bevorzugung von warmen und feuchten Klimaten mit einem nur geringen Anteil an Frosttagen im Jahr zu beobachten. Bevorzugt bewohnte Landschaften sind hierbei Ufergebiete und feuchte Wälder, vor allem die atlantischen Küstenwälder (Mata Atlântica), aber auch Sümpfe. Es fehlt aber vor allem in den sehr trockenen Gebieten des Gran Chaco mit weniger als 380 mm Jahresniederschlag, die von anderen Gürteltierarten bewohnt werden. Möglicherweise ist das auch der Grund, warum das Neunbinden-Gürteltier nicht weiter südlich anzutreffen ist. Die Populationsdichte der einzelnen Regionen ist sehr unterschiedlich. So ist in der Cerrado-Region nur ein Individuum auf drei Quadratkilometern anzutreffen, in den küstennahen Gebieten von Texas kommen aber bis zu 50 Tiere auf einem Quadratkilometer vor. In geschützten Arealen der atlantischen Küstenwälder können es bis zu 24 Individuen sein, die die gleiche Fläche besetzen.[1][7][3]
Ausbreitung nach Nordamerika
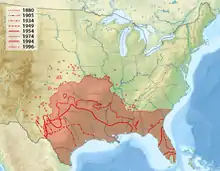
Die Ausbreitung nach Nordamerika, vor allem in die USA, die erst in jüngerer Zeit stattfand und immer noch anhält, ist gut untersucht. Die ersten Tiere wurden in südlichsten Texas am Rio Grande im Jahr 1854 beobachtet, bereits 1880 hatte die Gürteltierart den Nueces River im Südosten des US-Bundesstaates erreicht.[8] Die Westexpansion stoppte etwa 1905 am Pecos River in New Mexico, gegenwärtig sind nur selten vereinzelte Tiere westlich des Flusses zu finden.[9] Nach Osten hin erschien das Neunbinden-Gürteltier erstmals 1921 in Arkansas. Schon 1919 wurden mehrere Tiere in Florida durch Marinesoldaten freigelassen, gleichzeitig entwichen einige Exemplare aus einem Zoo, beide Gruppen begründeten die Population in diesem Bundesstaat. Im weiteren Verlauf der Ostausdehnung des Verbreitungsgebietes trat das Neunbinden-Gürteltier 1925 erstmals in Louisiana auf, den Mississippi überschritt es 1934, kam 1939 in Alabama an und 1942 in Georgia. Weiterhin erfolgte auch eine Nordausdehnung, so wird über das erste Auftreten der Art 1936 in Oklahoma, 1943 in Kansas, 1947 in Missouri, 1966 in Colorado, 1970 in Nebraska, 1974 in South Carolina und schließlich 1992 in Illinois berichtet. Gegenwärtig umfasst die Verbreitung den ganzen Südosten der USA, im Westen ist die Ausdehnung weitgehend durch die trockenen Wüstengebiete mit heftigen Wintereinbrüchen abgeschlossen, im Norden geht sie jedoch weiter und schreitet mit einer Geschwindigkeit von durchschnittlich 7,8 km pro Jahr voran, in den Großen Ebenen liegt sie sogar bei 11 km, an der Golfküste erreichte sie bis zu 17,2 km pro Jahr.[10] Allerdings wird die Expansion nach Norden durch die kalten Winter gebremst, da das Neunbinden-Gürteltier kaum kälteresistent ist. Ein weiterer limitierender Faktor ist das weniger reichhaltige Nahrungsangebot in der kalten Jahreszeit dort.[1][2]
Warum das Neunbinden-Gürteltier erst so spät in Nordamerika auftrat, ist unbekannt, da es schon wesentlich früher, seit einigen tausend Jahren, in Mexiko nachgewiesen ist und keine klimatischen Unterschiede zum südlichen Teil Nordamerikas bestehen oder geographische Barrieren vorliegen. Möglicherweise hat es mit der Größe und Dichte der Populationen in Mexiko zu tun oder aber mit dem Überwinden eines physiologischen Hemmfaktors. Letzteres wäre nur mit einer evolutiven Weiterentwicklung des Neunbinden-Gürteltiers zu erklären.[7]
Lebensweise
Territorialverhalten

Das Neunbinden-Gürteltier ist die am besten untersuchte und dokumentierte Gürteltierart, dementsprechend viel ist über ihre Lebensweise bekannt. Das Tier lebt einzelgängerisch und ist überwiegend nachtaktiv, allerdings ist es auch zeitweise am Tag sichtbar. Da es weniger kälteangepasst ist, sind die Aktivitäten in kühleren Phasen geringer, auch wird die Körpertemperatur um bis zu 2,5 °C abgesenkt. Die Vertreter der Gürteltierart sind nicht territorial, doch nutzen sie Aktionsräume (home ranges), in denen sie sich bevorzugt aufhalten und deren Größe mit dem Alter der einzelnen Individuen zunimmt. In optimalen Habitaten sind sie in der Regel zwischen 1,2 und 3,4 ha groß, in weniger gut geeigneten Landschaften können sie bis zu 15 ha groß werden. Unterschiede zwischen männlichen und weiblichen Tieren wurden dabei nicht beobachtet. Die Aktionsräume verschiedener Einzeltiere, sowohl der Männchen als auch der Weibchen, können sich überlappen, ohne dass es zu Aggressionen untereinander kommt. Lediglich in der Brunftzeit sind die Gebiete der männlichen Tiere strikt getrennt. Bei Kontakten von Individuen untereinander werden meist zuerst die Genitalien beschnüffelt. Während der täglichen Nahrungssuche legt ein einzelnes Tier bis zu 1,2 km zurück, die durchschnittlichen Distanzen liegen aber bei 80 bis 180 m. Zudem ist das Neunbinden-Gürteltier fähig, Gewässer zu überqueren, wobei es flache, schmale Gewässer einfach am Grund durchwandert, größere Gewässer aber schwimmend überquert; zu diesem Zweck pumpt es Luft in seinen Verdauungstrakt. Gelegentlich sieht man ein einzelnes Tier auch aktiv in einem kleinen Gewässer baden.[1][2][3]

Den Tag verbringt das Neunbinden-Gürteltier überwiegend in selbst gegrabenen Bauen, die es mit den kräftigen Vorderfüßen anlegt. im Durchschnitt besitzt ein Tier zwischen 4 und 8 Baue in seinem Aktionsgebiet. Diese sind 0,5 bis 6 m lang und befinden sich meist in Wäldern mit gut durchfeuchteten Böden und in der Nähe von Gewässern sowie überwiegend in Hanglage.[11] Sie reichen rund 50 cm schräg in den Untergrund, in Extremfällen sind sie bis zu 150 cm tief. Jeder Bau hat mehrere Eingänge von 17 bis 20 cm Durchmesser, die manchmal mit Pflanzen bedeckt sind. Die Eingänge sind nicht in eine bestimmte Richtung orientiert, im Gegensatz zu den Bauen von Gürteltierarten, die im Offenland leben und die diese deshalb windgeschützt anlegen. Der lange Gang endet in einer größeren, oft mit getrocknetem Gras oder Blättern gepolsterten Kammer von rund 34 cm Durchmesser. Das Material für das Nestpolster holt das Neunbinden-Gürteltier in mehreren Phasen aus der näheren Umgebung herbei. Im Bau herrscht häufig eine 3 bis 4° kühlere Temperatur als außen. Manchmal findet man mehrere Tiere in einem Bau, die aber in der Regel der gleichen Familie angehören und immer gleichen Geschlechts sind. In einigen Fällen teilt sich das Neunbinden-Gürteltier seinen Bau mit anderen Tierarten wie dem Gemeinen Schwarzleguan oder Skunks und Beutelratten. In Regionen, die regelmäßig überflutet werden, legt die Gürteltierart bauartige Nester aus Pflanzenmaterial auf dem Erdboden an, die etwa 20 mal 34 cm groß und 22 cm hoch sind und nur einen einzigen Eingang aufweisen.[12][1][3]
Ernährung

Das Neunbinden-Gürteltier ist in erster Linie ein Fleischfresser mit einer Spezialisierung auf Insekten, wobei vorzugsweise Käfer verspeist werden. Untersuchungen aus Nordamerika zeigen einen Anteil von 30 bis 45 % an Käfern und deren Larven in der Nahrung, hier vor allem Blatthornkäfer, Laufkäfer und Schnellkäfer. Des Weiteren sind Hautflügler, Heuschrecken und Schmetterlinge mit Anteilen von jeweils 4 bis 15 % vertreten, daneben wurden auch kleine Wirbeltiere wie Amphibien und Reptilien verzehrt. Dabei plündert die Gürteltierart gelegentlich auch Eigelege der Diamantschildkröte und Nester der Virginiawachtel. Beobachtet wurde auch, dass sie Fleisch von verendeten Kaninchen oder Vögeln, etwa der Wanderdrossel frisst. Eine eher untergeordnete Rolle spielen Früchte und Beeren, unter anderem solche von Ebenholzgewächsen, regional kann aber pflanzliche Nahrung auch bis zu 10 % des Nahrungsbedarfes einnehmen.[13][2] Ähnliche Untersuchungen aus der Cerrado-Region Brasiliens ergaben bei Käfern nur eine Häufigkeit von 1,1 %, die aber gut 71 % der gefressenen Biomasse ausmachten. Häufiger wurden Ameisen mit 32,6 % aufgenommen, der Biomasseanteil lag hier aber bei nur 16,5 %. Am höchsten jedoch war die verzehrte Menge von Termiten mit 66,3 %, die aber als reine Biomasse nur 12,2 % einnahmen. Besonders zahlreich wurden dabei Angehörige der Termitengattungen Velocitermes und Nasutitermes gefressen.[14][1] Auch im Nationalpark Serra da Capivara im nördlichen Brasilien konnten durch Untersuchungen an Kotresten und Mageninhalten überwiegend Käfer, Ameisen und Termiten nachgewiesen werden, zusätzlich aber auch Überreste von Leguanen.[15] Verschiedene analysierte Mageninhalte sowohl im süd- wie auch im nordamerikanischen Verbreitungsgebiet enthielten Schlangen als Nahrungsreste. Nach Beobachtungen im Nationalpark Santa Rosa in Costa Rica erlegte ein Neunbinden-Gürteltier eine giftige Korallenschlange durch Sprünge mit der Panzerrückseite auf das Tier. Der vollständige Verzehr der rund 1 m langen Schlange nahm etwa 30 Minuten in Anspruch.[16] Insgesamt zeigt sich das Neunbinden-Gürteltier als sehr opportunistisch in seinem Fressverhalten.[7][3]
Das Neunbinden-Gürteltier nutzt 77 bis 90 % seiner Wachphase mit der Nahrungssuche. Dabei läuft es behände hin und her, schnüffelt an allen potenziellen Fressstellen und gibt ein beständiges Schnauben von sich. Ein Tier benutzt meist häufig begangene Pfade und bewegt sich oft von tieferen Wäldern in eher offenes Land. Während der Suche legt es zahlreiche Löcher an, in einem Sumpfkiefer-Ökosystem in Georgia wurden im Durchschnitt 267 Löcher von 9 cm Tiefe auf einem Hektar aufgefunden.[17] Häufig ist auch zu beobachten, dass sich das Tier aufrichtet, wobei es sich mit dem Schwanz dabei abstützt, und in der Luft schnüffelt. Um einer Duftspur zu folgen, hält es die Nase bei der Futtersuche sehr dicht über dem Boden. Zudem stoppt die Atmung bis zu sechs Minuten lang, damit kein Staub eingeatmet wird. Bei der Nahrungssuche ist das Neunbinden-Gürteltier so konzentriert, dass es bisweilen gegen Hindernisse läuft.[1][2][3]
Fortpflanzung
Männliche Tiere sind mit etwa zwölf Monaten geschlechtsreif, weibliche mit 18. Eine Paarung findet nur einmal jährlich statt, die Brunft beim Weibchen dauert gut vier Tage. Die Paarungszeit ist aber aufgrund der weiten Verbreitung des Neunbinden-Gürteltiers regional sehr unterschiedlich und abhängig von den Jahreszeiten. In Nordamerika findet sie hauptsächlich von Juni bis November, in Bolivien von Oktober bis Dezember[18] und in Paraguay von August bis November statt. Nur während dieser Zeit kommt es zur Paarbildung zwischen männlichen und weiblichen Tieren. Auch finden teilweise Dominanzkämpfe zwischen den werbenden Männchen statt, die teils auf den Hinterfüßen stehend und mit den Krallen der Vorderfüße ausgetragen werden, aber auch indem ein Tier nach hinten ausschlägt. Aufgrund der Anatomie des Panzers findet die Befruchtung mit dem auf dem Rücken liegenden Weibchen statt.[1][2][3]
Die Tragzeit dauert etwa 8 bis 9 Monate, die eigentliche Entwicklung im Mutterleib benötigt aber durchschnittlich nur rund 140 Tage, es kommt jedoch aufgrund einer Diapause nach der Befruchtung und Einlagerung der Eizelle in die Gebärmutter zu einer Entwicklungsverzögerung des Embryos von bis zu 4 Monaten. Dies führt dazu, dass die Nachkommen in einer klimatisch günstigen Zeit zur Welt gebracht werden können. Es kann aber auch in Fällen von Stress zu einer verzögerten Implantation der Eizelle kommen, was bedingt, dass die Geburt erst bis zu 32 Monate nach der letzten Befruchtung stattfindet. Auch wurde beobachtet, dass Weibchen zweimal hintereinander gebaren, ohne dass Kontakt zu Männchen bestand, was ebenfalls auf die verzögerte Implantation zurückzuführen ist.[7][3]
Üblicherweise kommen vier Jungtiere zur Welt, es gibt aber auch Berichte über zwei oder acht Nachkommen. Aufgrund von Polyembryonie sind alle Jungtiere genetisch identisch und die Anzahl der Nachkommen dadurch fast immer geradzahlig, nur sehr selten wurde die Geburt von Drillingen beobachtet. Ein Junges wiegt etwa 85 bis 100 g. Es ähnelt bereits erwachsenen Tieren, allerdings sind der Panzer und die Krallen weicher und härten sich erst im Lauf der ersten Lebenswochen.[19] Weiterhin sind die Augen geöffnet und ein Neugeborenes kann bereits nach wenigen Stunden laufen. Die Saugphase dauert insgesamt etwa drei Monate und die Jungtiere nehmen anfänglich etwa 11 g je Tag an Gewicht zu. Erstmals das Nest verlassen sie nach rund 20 Tagen und gehen mit dem Muttertier auf Nahrungssuche, wobei sie auch zum ersten Mal Wasser trinken. Die ersten Insekten werden jedoch erst nach rund 70 Tagen gefressen. Nach der Entwöhnung bleibt die Familiengruppe noch etwa sechs weitere Monate zusammen, bis die Jungtiere herangewachsen sind und das Muttertier verlassen. Die Lebenserwartung liegt bei zwölf bis fünfzehn Jahren.[1][2][3]
Die Polyembryonie hat zur Folge, dass die genetische Diversität einer Gruppe in unmittelbarer Umgebung eines Mutternestes relativ gering ist, was Studien an mehreren hundert Individuen im Yazoo National Wildlife Reserve im US- Bundesstaat Mississippi erbrachten. Erst mit zunehmender Entfernung vom ursprünglichen Nest steigt auch die genetische Variabilität an. Dies hängt mit der Abwanderung der Jungen nach der Entwöhnung zusammen. Die zunehmende Vielfalt in der genetischen Struktur ist bei weiblichen Tieren höher als bei männlichen. Die an der Untersuchung beteiligten Wissenschaftler boten zwei Erklärungsmodelle für dieses Phänomen an: einerseits ließe sich dadurch auf eine häufigere Abwanderung der Weibchen und eine stärkere Ortstreue (Philopatrie) der Männchen schließen, andererseits könnte sich hier auch der unterschiedliche Paarungserfolg der Männchen bei polygyner Fortpflanzung widerspiegeln. Eine ausschließliche Abwanderung weiblicher Individuen ist aus anderen Bereichen des Verbreitungsgebietes bisher genetisch nur selten belegt.[20][21]
Beutegreifer und Feindverhalten

Üblicherweise flüchtet das Neunbinden-Gürteltier bei aufziehender Gefahr in den nächsten Bau und verstopft den Eingang korkenartig mit seinem Beckenpanzer, außerdem krümmt es den Rücken und verhakt sich mit den beweglichen Bändern in der umliegenden Erde, so dass es kaum aus seinem Bau herausgezogen werden kann. Beim Laufen ist ein Tier sehr schnell und kann Berichten zufolge auch jagenden Hunden entkommen. Zudem produziert das Neunbinden-Gürteltier ein Sekret, welches Übelkeit bei potentiellen Fressfeinden auslöst. Ein aufmerksames Tier schnüffelt auf den Hinterbeinen stehend, ein erschrockenes springt mit allen vieren in die Luft und beugt den Rückenpanzer. Zu den bedeutendsten Fressfeinden gehören der Jaguar und der Puma, jedoch ist der Druck, den beide Beutegreifer auf die Populationen ausüben regional unterschiedlich. In Belize konnte in 54 % von 183 untersuchten Kotresten des Jaguars das Neunbinden-Gürteltier nachgewiesen werden,[22] in den Mata-Atlântica-Küstenwäldern des südöstlichen Brasiliens waren es noch 15 %.[23] Im brasilianischen Bundesstaat São Paulo bestanden 8,3 % der Fäkalien vom Puma und 3,7 % jener vom Jaguar aus Resten der Gürteltierart.[24] In Nordamerika scheint sie aber kaum zum Beutespektrum des Pumas zu gehören, ebenso wenig zu dem des Kojoten, da bei 566 untersuchten Fäzes kein Nachweis erbracht werden konnte.[2] In einigen Fällen wurde auch vom Ozelot als Beutegreifer berichtet. Jungtiere können zudem der Harpyie oder dem Schwarzweißen Teju zum Opfer fallen.[1]
Parasiten
Trotz der großen Verbreitung des Neunbinden-Gürteltiers sind nur wenige Parasiten nachgewiesen, von denen es befallen wird. Zu den häufigen äußeren gehören Zecken der Gattung Amblyomma.[25] Flöhe sind unter anderem mit Tunga- und Polygenes-Arten vertreten. Innere Parasiten umfassen überwiegend Fadenwürmer, so beispielsweise Aspidodera und Moeniggia. Bedeutend ist die Gürteltierart vor allem als Träger von Mycobacterium leprae, welches die Lepra auch beim Menschen hervorrufen kann. Die Höhe des Übertragungsrisikos ist aber unklar. In Louisiana, wo die Krankheit seit Mitte des 19. Jahrhunderts endemisch ist, trat das Neunbinden-Gürteltier erst in den 1920er Jahren auf. Weiterhin ist Lepra allein in den USA deutlich weiter verbreitet als die Gürteltierart insgesamt, doch konnte 2011 in einer Studie der Übertragungsweg über die Tierart nachgewiesen werden. Die Krankheit selbst wurde erst mit der Besiedlung Amerikas durch die Europäer dort heimisch und kam vorher nicht vor.[26] Ebenso ist Trypanosoma cruzi beim Neunbinden-Gürteltier nachgewiesen. Dieser Einzeller verursacht die in Südamerika häufige Chagas-Krankheit.[7][1]
Systematik
Innere Systematik der Gürteltiere nach Gibb u. a. 2015[27]
|
Das Neunbinden-Gürteltier ist eine von sieben Arten, welche heute die Gattung der Langnasengürteltiere (Dasypus) formen. Die Langnasengürteltiere gehören wiederum zur Gruppe der Gürteltiere (Dasypoda) und bilden innerhalb dieser eine eigene Familie, die Dasypodidae. Dieser gehören auch zahlreiche ausgestorbene Gattungen an, wie beispielsweise Stegotherium und Propraopus. Dabei ist ersteres weitgehend aus dem Miozän bekannt und umfasste mehrere Arten,[28][29] letzteres dagegen stammt aus dem Pleistozän und trat gleichfalls mit mehreren Arten auf.[30] Laut molekulargenetischen Untersuchungen trennten sich die Dasypodidae bereits im Mittleren Eozän vor rund 45 Millionen Jahren von der Linie der anderen Gürteltiere ab. Diese werden heute zur Familie der Chlamyphoridae gestellt, die alle anderen rezenten Gattungen einschließt.[31][32][27]
Zusammen mit dem Siebenbinden- (Dasypus septemcinctus), dem Südlichen Siebenbinden- (Dasypus hybridus), dem Savannen- (Dasypus sabanicola) und dem Yungas-Gürteltier (Dasypus mazzai) bildet das Neunbinden-Gürteltier eine enger miteinander verwandte Gruppe, die der Untergattung Dasypus angehört. Innerhalb dieser steht es laut molekulargenetischen Untersuchungen aus dem Jahr 2015 wiederum mit dem Yungas- und dem Savannen-Gürteltier in näherer Beziehung. Das Kappler-Gürteltier (Dasypus kappleri) wird dagegen in eine eigene Untergattung, Hyperoambon, verwiesen, ebenso das Pelzgürteltier (Dasypus pilosus) in Cryptophractus. Ein weiteres Ergebnis der neueren molekulargenetischen Untersuchungen ist, dass eine stärkere Diversifizierung der Untergattung Dasypus vor etwa 5 Millionen Jahren begann, was dem Übergang vom Miozän zum Pliozän entspricht.[31][32][27]
Insgesamt werden heute sieben Unterarten unterschieden:[1]
- D. n. aequatorialis Lönnberg, 1913; Südamerika westlich der Anden
- D. n. davisi Russell, 1953; Mittelamerika vom Río Balsas bis Morelos
- D. n. fenestratus Peters, 1864; Mittelamerika von Oaxaca bis Panama
- D. n. hoplites Allen, 1911; Karibische Inseln und Costa Rica
- D. n. mexianae Hagmann, 1908; Südamerika im Mündungsbereich des Amazonas
- D. n. mexicanus Peters, 1864; Nordamerika und Mittelamerika bis südliches Mexiko
- D. n. novemcinctus Linnaeus, 1758; Südamerika östlich der Anden bis nach Argentinien
Untersuchungen zufolge besitzen die Populationen in Nordamerika vergleichsweise eine nur geringe genetische Variabilität, im Gegensatz zu denen in Südamerika. Erklärt wird dies mit einer nur kleinen Gründergruppe, die Nordamerika vor rund 160 Jahren erreichte.[33] Dabei ließ sich bei genetischen Analysen von rund 150 Tieren aus Mexiko zeigen, dass diese zwei unterschiedlichen Kladen angehören, die sich wahrscheinlich schon im Oberen Pleistozän in Südamerika herausdifferenziert haben. Die beiden Gruppen verteilen sich einmal eher im östlichen und einmal eher im westlichen Teil des Landes mit der Sierra Madre Oriental als mögliche natürliche Barriere. Dadurch wurde Nordamerika wohl zweimal unabhängig von unterschiedlichen Populationen des Neunbinden-Gürteltiers erreicht.[34]
Möglicherweise stellt eine Population aus Französisch-Guayana eine eigenständige Art dar, da diese sich nach den neueren genetischen Studien aus dem Jahr 2015 stärker unterscheidet und sich wohl schon vor etwa 3 Millionen Jahren von der Linie, die zu den heutigen Neunbinden-Gürteltieren führte, abgespalten hatte.[27] Bestätigt wird dies auch durch schädelanatomische Untersuchungen aus dem Jahr 2018. Diese ergaben darüber hinaus, dass sich das Neunbinden-Gürteltier in seinem übrigen Verbreitungsgebiet in drei bis vier morphologische Typen gliedern lässt. Den Autoren der Analyse zufolge könnten diese Typen eigene taxonomische Einheiten bilden, die aber nicht mit den bereits zuvor definierten Unterarten übereinstimmen.[35]

Fossilfunde des Neunbinden-Gürteltiers sind nur wenige bekannt, die Art ist fast vollständig auf das Holozän beschränkt. Aus der Medford-Höhle in Florida stammen einzelne Reste in Form von Knochenplättchen und Gliedmaßenresten, die genetischen Untersuchungen zufolge als Reste des Neunbinden-Gürteltiers anzusehen sind. Da das Alter der Funde auf etwa 10.000 bis 12.000 Jahre datiert wird und damit ins ausgehende Pleistozän gehört, ist die Anwesenheit dieser Art schon sehr früh in Nordamerika belegt ist.[36] Aus Südamerika liegen ebenfalls nur wenige Fossilien, zumeist Knochenschildchen vor, so aus den Höhlen Gruta dos Moura und Gruta do Urso im brasilianischen Bundesstaat Tocantins und aus der Gruta do Urso Fóssil im Ubajara-Nationalpark im brasilianischen Bundesstaat Ceará. Die zeitliche Reichweite der Funde umfasst das ausgehende Pleistozän und das frühe Holozän.[37][38] Der überwiegende Teil der Fossilfunde von Langnasengürteltieren des Pleistozäns steht mit Dasypus bellus, das „Schöne Gürteltier“, in Verbindung, das als nah verwandt mit dem Neunbinden-Gürteltier angesehen wird, aber mit einer Gesamtlänge von rund 1,2 m und einem Gewicht bis über 15 kg deutlich größere Ausmaße besaß.[39] Dieser nahen Verwandtschaft wurde in einer Studie aus dem Jahr 2008 aber widersprochen, in der Dasypus bellus in Beziehung mit dem Kappler-Gürteltier steht,[30][40] allerdings belegen phylogenetische Untersuchungen des Jahres 2015 die nahe Verwandtschaft mit dem Neunbinden-Gürteltier.[41]
Die erste Erwähnung des Neunbinden-Gürteltiers in Europa beruhte auf den spanischen Conquistador Francisco Hernández de Córdoba, der 1517 die Halbinsel Yucatán entdeckte und für das Tier in seinen Schriften das lokale aztekische Wort Azotochtli verwendete.[42] Azotochtli bedeutet dabei so viel wie „Schildkrötenhase“ und bezieht sich auf das Aussehen des Tieres. Die Erstbeschreibung des Neunbinden-Gürteltiers erfolgte 1758 durch Linnaeus, wobei er als Typusvorkommen „in America Meridionali“ angab. Er unterschied die Gürteltierart vom Siebenbinden-Gürteltier (Dasypus septemcinctus) anhand der Anzahl der beweglichen Bänder, allerdings nahm er für letzteres als Verbreitungsgebiet Indien an. Den Gattungsnamen Dasypus übersetzte Linnaeus vom Wort Azotochtli in die griechische Sprache, da er letzteres als ungeeignet für eine Latinisierung und damit für eine Verwendung in der Binären Nomenklatur befand, die von ihm eingeführt worden war. Dabei ist Dasypus als tautonyme Form von Azotochtli anzusehen und hat aus diesem Grund die gleiche Bedeutung.[43] Der Artname novemcinctus bezieht sich auf die Anzahl der frei beweglichen Bänder.[1][2]
Neunbinden-Gürteltier und der Mensch

Bedeutung
Als einzige Gürteltierart hat das Neunbinden-Gürteltier sein Verbreitungsgebiet seit der Ankunft der Europäer in Nordamerika drastisch ausweiten können. Das Verhältnis der Menschen zu diesen Tieren ist zwiespältig: einerseits werden sie ihres Fleisches wegen und als Insektenvertilger geschätzt. Andererseits werden sie verfolgt, da sie gelegentlich Felder umgraben und Vogeleier fressen, allerdings machen diese nur einen Bruchteil ihrer Nahrung aus. In der medizinischen Forschung spielen diese Tiere eine wichtige Rolle: Da die Gürteltierart sowohl das Leprabakterium Mycobacterium leprae als auch den die Chagas-Krankheit auslösenden Parasiten Trypanosoma cruzi in sich tragen kann, werden Behandlungsmethoden und Impfstoffe an ihnen getestet.[1][2]
Bedrohung und Schutz
Größere Bedrohungen für das Neunbinden-Gürteltier sind nicht bekannt, es wird aber innerhalb seines gesamten Verbreitungsgebietes gejagt. So ist es die Hauptnahrung des Indianervolkes der Aché in Paraguay, die laut einer Studie zwischen 1980 und 1996 insgesamt 1500 Individuen dieser Gürteltierart erlegten, was einem Gesamtgewicht von 5,7 t entsprach und in etwa 32 % ihrer benötigten Nahrung ausmachte. In anderen Regionen gilt es als beliebtes Wildgericht, das zum Teil im Panzer angerichtet wird. In Nordamerika werden Neunbinden-Gürteltiere häufig bei Autounfällen getötet, wobei ihre Angewohnheit, bei Erschrecken hochzuhüpfen ihnen hier zum Verhängnis wird. So stellt sie in den südlichen Großen Ebenen die dritthäufigste, durch Fahrzeuge getötete Säugetierart dar. Insgesamt ist das Neunbinden-Gürteltier weit verbreitet und wird von der IUCN als „nicht gefährdet“ (least concern) eingestuft, mit einer steigenden Populationsgröße. Es kommt in zahlreichen geschützten Gebieten innerhalb des gesamten Verbreitungsgebietes vor.[44][1]
Literatur
- Karen McBee, Robert J. Baker: Dasypus novemcinctus. In: Mammalian Species. 162, 1982, S. 1–9.
- C. M. McDonough und W. J. Laughry: Dasypodidae (Long-nosed armadillos). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 30–47 (S. 44–45) ISBN 978-84-16728-08-4.
Einzelnachweise
- Paul Smith: Nine-banded armadillo Dasypus novemcinctus Linnaeus 1758. In: Mammals of Paraguay. 8, 2008, S. 1–30.
- Karen McBee, Robert J. Baker: Dasypus novemcinctus. In: Mammalian Species. 162, 1982, S. 1–9.
- C. M. McDonough und W. J. Laughry: Dasypodidae (Long-nosed armadillos). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 30–47 (S. 44–45) ISBN 978-84-16728-08-4.
- Hugh H. Genoways, Robert M. Timm: The Xenarthrans of Nicaragua. In: Mastozoologia Neotropical. 10 (2), 2003, S. 231–253.
- S. F. Vizcaíno, N. Milne: Structure and function in armadillo limbs (Mammalia: Xenarthra: Dasypodidae). In: Journal of Zoology. 257, 2002, S. 117–127.
- Agustín M. Abba, M. Superina: Dasypus novemcinctus. In: Edentata. 11 (2), 2010, S. 160.
- Mariella Superina: Biologie und Haltung von Gürteltieren (Dasypodidae). Universität Zürich, 2000, OCLC 632874842.
- Edward Drinker Cope: On the zoological position of Texas. In: Bulletin of the United States National Museum. 17, 1880, S. 1–51.
- Jennifer K. Frey, James N. Stuart: Nine-Banded Armadillo (Dasypus novemcinctus) Records in New Mexico, USA. In: Edentata. 8–10, 2009, S. 54–55.
- S. Erich Eichler, Timothy J. Gaudin: New records of the nine-banded armadillo, Dasypus novemcinctus, in southeast Tennessee, and their implications. In: Edentata. 12, 2011, S. 7–13.
- Maria Clara Arteaga, Eduardo Martins Venticinque: Influence of topography on the location and density of armadillo burrows (Dasypodidae: Xenarthra) in the central Amazon, Brazil. In: Mammalian Biology. 73, 2008, S. 262–266.
- James N. Layne: Above-ground nests of the nine-banded armadillo in Florida. In: Florida Field Naturalist. 12, 1984, S. 58–61.
- Kent H. Redford: Dietary specialization and variation in two mammalian myrmecophages (variation in mammalian myrmecophagy). In: Revista Chilena de Historia Natural. 59, 1986, S. 201–208.
- Teresa Cristina da Silveira Anacleto: Food Habits of Four Armadillo Species in the Cerrado Area, Mato Grosso, Brazil. In: Zoological Studies. 46, 2007, S. 529–537.
- Vanderson Corrêa Vaz, Ricardo Tadeu Santori, Ana Maria Jansen, Ana Cláudia Delciellos, Paulo Sérgio D’Andrea: Notes on food habits of armadillos (Cingulata, Dasypodidae) and anteaters (Pilosa, Myrmecophagidae) at Serra da Capivara National Park (Piauí State, Brazil). In: Edentata. 13, 2012, S. 84–89.
- Eduardo Carrillo und Todd K. Fuller: Predation of a Central American coral snake (Micrurus nigrocinctus) by a nine-banded armadillo (Dasypus novemcinctus) in Santa Rosa National Park, Costa Rica. In: Edentata. 19, 2018, S. 67–69.
- Samuel M. Simkin, William K. Michener: Faunal Soil Disturbance Regime of a Longleaf Pine Ecosystem. In: Southeastern Naturalist. 4 (1), 2005, S. 133–152.
- Clara R. Rojas-Suárez, Leonardo Maffei: Reproducción de Dasypus novemcinctus en el Izozog, Santa Cruz, Bolivia. In: Edentata. 5, 2003, S. 47–54.
- Mariella Superina, W. J. Loughry: Life on the Half-Shell: Consequences of a Carapace in the Evolution of Armadillos (Xenarthra: Cingulata). In: Journal of Mammal Evolution. 19, 2012, S. 217–224.
- Silvia D. Frutos, Ronald A. van den Bussche: Genetic diversity and gene flow in Nine-banded armadillos in Paraguay. In: Journal of Mammalogy. 83 (3), 2002, S. 815–823.
- Loren A. Binns, W. J. Loughry, Colleen M. McDonough, Corey Devin Anderson: Spatial genetic structure within a population of nine-banded armadillos in western Mississippi. In: Journal of Mammalogy. 101 (1), 2020, S. 143–150, doi:10.1093/jmammal/gyz174.
- A. R. Rabinowitz, B. G. Nottingham: Ecology and Behaviour of the Jaguar Panthera onca in Belize, Central America. In: Journal of Zoology. 210, 1986, S. 149–159.
- Ricardo C. Garla, Eleonore Z. F. Setz, Nivar Gobbi: Jaguar (Panthera onca) Food Habits in Atlantic Rain Forest of Southeastern Brazil. In: Biotropica. 33 (4), 2001, S. 691–696.
- Rogério Martins, Juliana Quadros, Marcello Mazzolli: Hábito Alimentar e Interferência Antrópica na Atividade de Marcação Territorial do Puma concolor e Leopardus pardalis (Carnivora: Felidae) e Outros Carnívoros na Estação Ecológica de Juréia.Itatins, São Paulo, Brasil. In: Revista Brasileira de Zoologia. 25, 2008, S. 427–435.
- A.A. Guglielmone, A. Estrada-Peña, C. A. Luciani, A. J. Mangold, J. E. Kerans: Hosts and distribution of Amblyomma auricularium (Conil 1878) and Amblyomma pseudoconcolor Aragão, 1908 (Acari: Ixodidae). In: Experimental and Applied Acarology. 29, 2003, S. 131–139.
- Richard W. Truman, Pushpendra Singh, Rahul Sharma, Philippe Busso, Jacques Rougemont, Alberto Paniz-Mondolfi, Adamandia Kapopoulou, Sylvain Brisse, David M. Scollard, Thomas P. Gillis, Stewart T. Cole: Probable Zoonotic Leprosy in the Southern United States. In: The New England Journal of Medicine. 364 (17), 2011, S. 1626–1633.
- Gillian C. Gibb, Fabien L. Condamine, Melanie Kuch, Jacob Enk, Nadia Moraes-Barros, Mariella Superina, Hendrik N. Poinar, Frédéric Delsuc: Shotgun Mitogenomics Provides a Reference Phylogenetic Framework and Timescale for Living Xenarthrans. In: Molecular Biology and Evolution. 33 (3), 2015, S. 621–642.
- Timothy J. Gaudin, John R. Wible: The phylogeny of living and extinct armadillos (Mammalia, Xenarthra, Cingulata): a craniodental analysis. In: Matthew T. Carrano, Timothy J. Gaudin, Richard W. Blob, John R. Wible (Hrsg.): Amniote Paleobiology: Phylogenetic and Functional Perspectives on the Evolution of Mammals, Birds and Reptiles. University of Chicago Press, Chicago 2006, S. 153–198.
- Laureano Raúl González Ruiz, Gustavo Juan Scillato-Yané: A new Stegotheriini (Mammalia, Xenarthra, Dasypodidae) from the “Notohippidian” (early Miocene) of Patagonia, Argentina. In: Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 252 (1), 2009, S. 81–90.
- Ascanio D. Rincón, Richard S. White, H. Gregory Mcdonald: Late Pleistocene Cingulates (Mammalia: Xenarthra) from Mene De Inciarte Tar Pits, Sierra De Perijá, Western Venezuela. In: Journal of Vertebrate Paleontology. 28 (1), 2008, S. 197–207.
- Maren Möller-Krull, Frédéric Delsuc, Gennady Churakov, Claudia Marker, Mariella Superina, Jürgen Brosius, Emmanuel J. P. Douzery, Jürgen Schmitz: Retroposed Elements and Their Flanking Regions Resolve the Evolutionary History of Xenarthran Mammals (Armadillos, Anteaters and Sloths). In: Molecular Biology and Evolution. 24, 2007, S. 2573–2582.
- Frédéric Delsuc, Mariella Superina, Marie-Ka Tilak, Emmanuel J. P. Douzery, Alexandre Hassanin: Molecular phylogenetics unveils the ancient evolutionary origins of the enigmatic fairy armadillos. In: Molecular Phylogenetics and Evolution. 62, 2012, S. 673–680.
- Dorothée Huchon, Frédéric Delsuc, François M. Catzeflis, Emmanuel J. P. Douzery: Armadillos exhibit less genetic polymorphism in North America than in South America: nuclear and mitochondrial data confirm a founder effect in Dasypus novemcinctus (Xenarthra). In: Molecular Ecology. 8 (10), 1999, S. 1743–1748.
- Maria Clara Arteaga, Daniel Piñero, Luis E. Eguiarte, Jaime Gasca, Rodrigo A. Medellín: Genetic structure and diversity of the Nine-banded armadillo in Mexico. In: Journal of Mammalogy. 93 (2), 2012, S. 547–559.
- Lionel Hautier, Guillaume Billet, Benoit de Thoisy und Frédéric Delsuc: Beyond the carapace: skull shape variation and morphological systematics of long-nosed armadillos (genus Dasypus). In: PeerJ. 5, 2017, S. e3650, doi:10.7717/peerj.3650.
- Beth Shapiro, Russell W. Graham, Brandon Leets: A revised evolutionary history of armadillos (Dasypus) in North America based on ancient mitochondrial DNA. In: Boreas. 44 (1), 2015, S. 14–23, doi:10.1111/bor.12094.
- Esteban Soibelzon, L. S. Avilla, M. Castro: The cingulates (Mammalia: Xenarthra) from the late Quaternary of northern Brazil: Fossil records, paleoclimates and displacements in America. In: Quaternary International. 377, 2015, S. 118–125, doi:10.1016/j.quaint.2015.02.052.
- Paulo V. Oliveira, Ana Maria Ribeiro, Édison V. Oliveira, Maria Somália S. Viana: The Dasypodidae (Mammalia, Xenarthra) from the Urso Fóssil Cave (Quaternary), Parque Nacional de Ubajara, State of Ceará, Brazil: paleoecological and taxonomic aspects. In: Anais da Academia Brasileira de Ciências. 86 (1), 2014, S. 147–158.
- H. Gregory McDonald: Paleoecology of extinct Xenarthans and the Great American Biotic Interchange. In: Bulletin of the Florida Museum of Natural History. 45 (4), 2005, S. 313–333.
- Mariela C. Castro, Ana Maria Ribeiro, Jorge Ferigolo, Max C. Langer: Redescription of Dasypus punctatus Lund, 1840 and considerations on the genus Propraopus Ameghino, 1881 (Xenarthra, Cingulata). In: Journal of Vertebrate Paleontology. 33 (2), 2013, S. 434–447.
- Mariela C. Castro, Martín R. Ciancio, Victor Pacheco, Rodolfo M. salas-Gismondi, J. Enrique Bostelmann, Alfredo A. Carlini: Reassessment of the hairy long-nosed armadillo “Dasypus” pilosus (Xenarthra, Dasypodidae) and revalidation of the genus Cryptophractus Fitzinger, 1856. In: Zootaxa. 3947 (1), 2015, S. 30–48, doi:10.11646/zootaxa.3947.1.2.
- J. M. Anderson, K. Benirschke: The armadillo, Dasypus novemcinctus, in experimental biology. In: Laboratory Animal Care. 16, 1966, S. 202–216.
- Oldfield Thomas: The mammals of the tenth edition of Linnaeus; an attempt to fix the types of the genera and the exact bases and localities of the species. In: Proceedings of the Zoological Society of London. 1911, S. 120–158.
- Agustín M. Abba, Mariella Superina: Dasypus novemcinctus. In: IUCN 2012: IUCN Red List of Threatened Species. Version 2012.2. zuletzt abgerufen am 27. März 2013.
Weblinks
- Dasypus novemcinctus in der Roten Liste gefährdeter Arten der IUCN 2006. Eingestellt von: Abba & Superina, 2006. Abgerufen am 27. März 2013.