Liste von Subtypen des Influenza-A-Virus
Diese Liste von Subtypen des Influenza-A-Virus gibt einen Überblick über die Varianten der Viren der „echten Grippe“ (Influenza-A-Subtypen) und – soweit belegbar – über deren Pathogenität für Tiere (→ Geflügelpest) und den Menschen (→ Influenza).
Eine umfassendere Übersicht über bislang bekannte Subtypen ist abrufbar auf der UniProt-Datenbank.[1]
Subtypen

Das Genom der Influenzaviren ist segmentiert, weshalb bei einer gleichzeitigen Infektion zweier Influenzastämme einzelne Genom-Segmente zwischen Influenzaviren ausgetauscht werden können (eine Antigenshift zur Immunevasion). Epidemien mit Influenzaviren entstehen meistens durch eine mangelnde Herdenimmunität gegen ein neu rekombiniertes Virus (sowohl in Tieren als auch in Menschen). Die geringe Immunität gegen veränderte Influenzaviren liegt an der vergleichsweise großen Änderung der viralen Proteine nach einer Antigenshift. Die Ausbildung einer Immunantwort gegen diese veränderten Antigene muss dann erneut erfolgen, was jedoch drei bis sieben Tage dauert. Bis dorthin zeigen sich ausgeprägte Symptome einer Grippe. Vermutlich sind die meisten heute zirkulierenden Gensegmente des Influenza-A-Virus im Jahr 1872 aus einer H3N8-Pferdegrippe über Nutzgeflügel auf den Menschen übergegangen.[2]
Als besonders bedrohlich für die Gesundheit von Geflügel und Menschen bewertet die Weltgesundheitsorganisation (WHO) die Subtypen-Gruppen A/H5 und A/H9, da sie besonders rasch von niedrigpathogenen in hochpathogene Varianten übergehen können.[3]
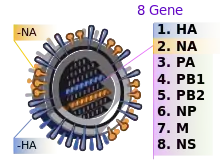
Die Abkürzung ‚H‘ steht für Hämagglutinin, die Abkürzung ‚N‘ steht für Neuraminidase. Bislang wurden 18 ‚H‘-Varianten und 11 ‚N‘-Varianten nachgewiesen.[4]
H1
A/H1N1
A/H1N1 ist ein häufig umlaufender Subtyp der Humaninfluenza. Er kann besonders leicht in menschliche Körperzellen eindringen und sein Erbgut einschleusen. Da sein erster Nachweis 1930 aus Schweinen erfolgt war,[5][6] werden durch diesen Subtyp verursachte Infektionen beim Schwein als Schweineinfluenza und beim Menschen – bezogen auf eine bis 2009 unbekannte Variante – umgangssprachlich als „Schweinegrippe“ bezeichnet.
Eine Variante von A/H1N1 konnte als Auslöser der so genannten Spanischen Grippe von 1918/1920 im Lungengewebe von Opfern nachgewiesen werden. 2005 gelang Jeffery Taubenberger eine Rekonstruktion des Erregers der Spanischen Grippe aus Genfragmenten. Im Jahr 2007 wurde durch Forscher des St. Jude Children’s Hospital, Memphis (Tennessee), bekannt, dass ein nur 90 Aminosäuren großes Virusprotein mit der Bezeichnung PB1-F2 verantwortlich für die ungewöhnlich hohe Letalität von A/H1N1 in den Jahren nach 1918 zu sein scheint.[7] Es bewirke besonders ausgeprägte Entzündungen bei den Infizierten. Testtiere erkrankten schon dann schwer, wenn nur ihre Nasenschleimhaut mit dem Protein in Berührung kam. Die heute noch kursierenden H1N1-Viren verfügen hingegen über ein verstümmeltes, nur 67 Aminosäuren umfassendes Protein: Dies sei die Folge davon, dass infolge einer Mutation ein Stopp-Signal ins PB1-F2-Gen eingefügt worden sei, so dass es nicht mehr vollständig abgelesen werde und das entstehende Protein daher minder pathogen sei.
Ein erneuter weltweiter Ausbruch – die so genannte Russische Grippe – ereignete sich 1977.[8]
Ende Januar 2008 wurde von norwegischen Ärzten bei normalen Grippepatienten ein gegen Oseltamivir resistenter Virusstamm (A/H1N1-H274Y) entdeckt,[9] der sich inzwischen weltweit verbreitet hat.[10] Mitte 2009 sind weltweit 96 %, in den entwickelten Ländern mit Benutzung von Oseltamivir praktisch alle[11] saisonalen A/H1N1-Viren resistent.[12]
Im April 2009 ereignete sich in Mexiko ein epidemieartiger Ausbruch einer bis dahin unbekannten Variante des H1N1-Subtyps, an dem zahlreiche Menschen erkrankten (siehe: Pandemie H1N1 2009/10). Einer 2016 publizierten Studie zufolge entstand die pandemische Variante des Virus (pdmH1N1) in Schweinen in Mexiko.[13]
Im Juni 2020 wurde mit „G4 EA H1N1“ eine Variante bekannt, die für Influenzaerkrankungen bei Schweinen in China in den Jahren zwischen 2011 und 2018 verantwortlich war; die Gene dieser Variante entstammen teils einer aus europäischen und asiatischen Vögeln bekannten Linie, teils dem pandemischen H1N1-Virus aus dem Jahr 2009 und teils einer aus Nordamerika bekannten Linie, die ihrerseits aus mehreren Herkünften zusammengesetzt ist.[14] Dieser Dreifachkombination wurde ein erhebliches „Pandemiepotenzial“[15] zugeschrieben. Laut einer im Juni 2020 im Fachblatt PNAS veröffentlichten Studie sind bereits mehr als 10 Prozent der Schweinehalter (35 von 338 untersuchten Personen) infiziert. Zudem wurde darauf hingewiesen, dass eine Infektion mit saisonaler Influenza keinen Schutz vor „G4 EA H1N1“ bietet.
A/H1N2
Der Subtyp A/H1N2 ist seit geraumer Zeit und von unterschiedlichen Kontinenten aus Geflügel und Schweinen bekannt,[16] ferner ist dieser Subtyp wiederholt – aber nur vereinzelt – von Schweinen auf Menschen übergegangen.[17] Erstmals nachgewiesen wurde A/H1N2 im Jahr 1980 in Japan in Schweinen; detaillierte Analysen ergaben, dass einige der umlaufenden Varianten von A/H1N2 als Reassortierungen von A/H1N1 und A/H3N2 waren.[18] Die Krankheitssymptome beim Menschen sind mild und vergleichbar mit anderen, saisonal auftretenden Subtypen.[19]
A/H1N3
Der Subtyp A/H1N3 entstand infolge einer Reassortierung auf der Basis von nicht-pandemischen A/H1N1- und A/H3N2-Viren; er ist seit 1998 in Nordamerika vereinzelt auch bei Menschen beobachtet worden, deren Krankheitsverläufe grippe-typisch, aber mild waren.[20][21] Bereits 1976 wurde beschrieben, dass der Subtyp in einem Wal nachgewiesen werden konnte.[22]
A/H1N8
Seroarchäologische Rekonstruktionen anhand von erhaltenen Antikörpern in konservierten Gewebeproben wurden dahingehend interpretiert, dass zwischen 1847 und 1889 sowie zwischen ca. 1900 und 1918 der Subtyp A/H1N8 umlief. Danach sei er verdrängt worden durch A/H1N1, den Erreger der Spanischen Grippe.[23]
H2
A/H2N1
Der älteste Nachweis des Subtyps A/H2N1 stammte aus dem Jahr 1957 und ist in den Influenza-Datenbanken unter der Bezeichnung A/JapanxBellamy/57(H2N1) registriert. Die UniProt-Datenbank weist zahlreiche jüngere Belege aus den USA nach, jedoch auch Virusfunde aus Deutschland – A/mallard/Potsdam/177-4/1983(H2N1), A/mallard/Stralsund/41-4/1981(H2N1) – sowie aus Schweden und Japan.[24]
A/H2N2
Ein weltweiter Ausbruch des Subtyps A/H2N2 war 1957 die Ursache einer Pandemie, die als Asiatische Grippe bezeichnet wurde.[25]
A/H2N3
Der Subtyp A/H2N3 ist weltweit unter frei lebenden Gänsen und Enten verbreitet.[26]
H3
A/H3N1
Der Subtyp A/H3N1 ist weltweit vor allem in Schweinen verbreitet und wurde in vielfältigen Reassortierungen nachgewiesen;[27] es gibt aber auch Nachweise aus der Geflügelhaltung.[28] Im Jahr 2003 wurde beispielsweise eine Reassortierung ausgehend von den Subtypen A/H3N2 (in Menschen umlaufend) und A/H1N1 (aus Schweinen) nachgewiesen.[29]
A/H3N2
Der Subtyp A/H3N2 ist in Europa und in den USA verbreitet. Ein weltweiter Ausbruch von A/H3N2 (A/Hong Kong/1/1968 (H3N2))[30] war 1968 die Ursache einer Pandemie beim Menschen, die als Hongkong-Grippe bezeichnet wurde. Belegt sind beispielsweise auch Übergänge von Schweinen auf den Menschen.[31] Die Influenza-Schutzimpfung umfasst daher regelmäßig auch einen Impfstoff gegen A/H3N2.[32] Durch eine Mutation im zirkulierenden Virus kam es zur Entwicklung eines Subclade (einer Untervariante), benannt 3C.2a1. Größte Ähnlichkeit mit dem veränderten Virus hat der Stamm A/Bolzano/7/2016 (H3N2). In der Grippesaison 2016/17 war daher für nur rund ein Viertel der Infektionen mit A/H3N2 der Stamm 3C.2a (ähnlich Hong Kong) verantwortlich, die anderen drei Viertel wurden durch den Stamm 3C.2a1 (ähnlich Bolzano) ausgelöst.[33]
Der Subtyp A/H3N2 wurde – wie auch A/H3N8 – wiederholt in Hunden nachgewiesen und wird dann als „Canine Influenza virus (CIV)“ bezeichnet.[34]
Im Jahr 2020 wurde festgestellt, dass dieser Subtyp durch eine Mutation die Fähigkeit erworben hat, der menschlichen Immunantwort und Impfmitteln viel leichter zu trotzen als zuvor.[35]
A/H3N3
Der Subtyp A/H3N3 wurde unter anderem 1992 aus Robben[36], 2007 aus Stockenten[37] und wiederholt aus Schweinen[38] isoliert.
A/H3N8
Der Subtyp A/H3N8 war ursprünglich bei Pferden verbreitet. Das Virus ging jedoch im frühen 21. Jahrhundert auf Hunde über und kommt bei dieser Art vor allem in den USA vor.[39] Seit 2011 ist eine Variante des Virus auch bei Seehunden nachgewiesen, unter denen es in Neuengland zu mehreren Dutzend Todesfällen kam.[40]
Infektionen beim Menschen sind bislang nicht durch direkten Virusnachweis belegt. Es gibt jedoch Hinweise auf ein Infektionsgeschehen unter Menschen im späten 19. Jahrhundert aufgrund von serologischen Befunden aus erhaltenen Gewebeproben.
H4
A/H4N1
Der Subtyp A/H4N1 wurde 1998 aus kanadischen Stockenten [A/mallard/Alberta/47/98(H4N1)] und 2009 in Zentralchina aus Schweinen isoliert, die Genomsequenz der chinesischen Isolate wurde im Dezember 2012 von chinesischen Forschern publiziert.[41]
H5
Von A/H5Nx sind neun Subtypen bekannt.[42]
A/H5N1
Der Subtyp A/H5N1 gilt als besonders aggressiv (HPAI, Highly Pathogenic Avian Influenza) und ist einer von mehreren Auslösern der Geflügelpest. Ein verändertes Nichtstruktur-Gen führt bei ihm dazu, dass bestimmte Botenstoffe des Immunsystems, welche normalerweise Viren abwehren, keine Wirkung mehr gegenüber dem A/H5N1-Subtyp erzielen. Eine Änderung im Hämagglutinin führt zu seiner verstärkten Aktivierung. Deshalb tötet der Subtyp befallene Vögel, die nicht zu seinem Virusreservoir gehören, vergleichsweise schnell und wird von Wissenschaftlern wegen seiner pathogenen Eigenschaften auf Interdependenzen mit anderen Stämmen und Überschreitungen der Artengrenze aufmerksam beobachtet.
Erstmals trat ein hoch pathogenes aviäres A/H5N1-Virus 1959 in Hühnervögeln in Schottland auf: A/chicken/Scotland/59 (H5N1).[43] Die heute zirkulierenden H5N1-Stämme tauchten 1997 in Nutzgeflügel in Hongkong auf. Diese Stämme erzeugten im Menschen eine ungewöhnlich hohe Letalität von über 50 %, verbreiteten sich jedoch nur schlecht im Menschen.
A/H5N2
Der Subtyp A/H5N2 ist weltweit verbreitet. So gab es beispielsweise in den Jahren 1983 und 1984 mehrere Ausbrüche in Geflügelfarmen der USA, als deren Folge 17 Millionen Tiere getötet wurden.[44] Zwischen 1992 und 1995 gab es zudem mehrere Ausbrüche in Mexiko.[45] 2015 trat das Virus in Kanada in einem 45.000 Tiere großen Jungputenbestand auf[46] und in Dutzenden großen Puten- und Hühnerbeständen insbesondere in den US-Bundesstaaten Iowa, Minnesota und Wisconsin, in denen insgesamt rund 30 Millionen Tiere starben oder getötet wurden;[47] diese 2015 in Kanada und den USA aufgetretene Variante des Subtyps hatte zuvor im Wege der Reassortierung Gene jener A/H5N8-Variante aufgenommen, die Ende 2014 u. a. in Deutschland in Tierhaltungen nachgewiesen worden war.
Im Sommer 2005 wurde A/H5N2 in Japan nachgewiesen, weswegen Presseberichten zufolge mehr als 1,5 Millionen Hühner und anderes Geflügel getötet wurden.[48] Im Frühjahr 2015 kam es auf Taiwan zu mehreren großen Ausbrüchen in Gänse- und Hühnerfarmen.[49][50]
Im Dezember 2008 wurde ein niedrig pathogenes A/H5N2 Virus in Belgien und Deutschland festgestellt. Dies führte zur Keulung von mehreren Geflügelbeständen in Niedersachsen.[51]
Eine Folge von Ausbrüchen ereignete sich ab Februar 2011 in Südafrika und wurde bis Sommer 2013 beendet.[52]
Anfang Dezember 2015 kam es in einem Geflügelbetrieb im bayrischen Roding zu einem H5N2-Ausbruch; 13.000 Hühner, Enten, Puten und Gänse wurden vorsorglich gekeult.[53] Im November 2016 war ein kleiner Geflügelbestand in Mesekenhagen (Mecklenburg-Vorpommern) betroffen.[54]
A/H5N3
Der Subtyp A/H5N3 verursachte im Jahr 1961 in Südafrika ein großes Sterben unter frei lebenden Seeschwalben. Bei ihnen gelang der erste Nachweis von Influenzaviren in einer Wildvogelpopulation.[55]
Im Oktober 2008 wurde ein niedrig pathogenes H5N3-Virus im Rahmen einer Routinekontrolle in einer Gans des Leipziger Zoos nachgewiesen. Im gleichen Jahr kam es zu Ausbrüchen in mehreren Nutzgeflügelbetrieben im Landkreis Cloppenburg, Niedersachsen.[56] Daraufhin wurden in dieser Region bis Ende Januar 2009 über 560.000 Vögel getötet.[57] Auch in den folgenden Jahren wurde der Subtyp wiederholt in Deutschland nachgewiesen, im Dezember 2016 beispielsweise in Nordrhein-Westfalen, Sachsen-Anhalt und Rheinland-Pfalz.[58]
A/H5N4
Der Subtyp A/H5N4 wurde laut Influenza Research Database des US-amerikanischen National Institute of Allergy and Infectious Diseases seit dem Jahr 2000 wiederholt in den USA sowie in Guatemala in Wildenten und Möwen nachgewiesen sowie im Jahr 2015 in einem Huhn in Irak.[59]
A/H5N5
Der Subtyp A/H5N5 wurde laut Influenza Research Database des US-amerikanischen National Institute of Allergy and Infectious Diseases in den USA und in China wiederholt in Stockenten und Gänsen nachgewiesen. Vereinzelt kamen auch hochpathogene Varianten des Subtyps vor.[60][61][62][63] Im November 2016 wurde er hochpathogen in den Niederlanden in einer frei lebenden Reiherente entdeckt,[64] ferner im Januar 2017 in einem Höckerschwan im Regionalbezirk Rodopi (Griechenland) und bei 20 Höckerschwänen in der Woiwodschaft Niederschlesien (Polen).[65][66] Im Januar 2017 wurde der Subtyp auch in einer Ente in Schleswig-Holstein[67] und zugleich erstmals in Europa in einer Geflügelzucht – in Süderau (Schleswig-Holstein) – nachgewiesen, wo 18.400 Puten getötet wurden.[68]
A/H5N6
Der Subtyp A/H5N6 wurde unter anderem bereits 1984 in Potsdam in Enten nachgewiesen (A/duck/Potsdam/2216-4/1984).[69] In China, Laos und Vietnam ist A/H5N6 unter Geflügel weit verbreitet.[70] Einen großen Ausbruch gab es Ende August 2014 in der Nähe von Harbin im Nordosten der Volksrepublik China, als dessen Folge mehr als 86.000 Stück Geflügel zu Tode kamen.[71] Im April 2015 wurden in Hongkong ein infizierter, wilder Wanderfalke[72] sowie eine gleichfalls wilde Dajaldrossel tot aufgefunden.[73]
Auf regionaler Ebene wurde 2020 für China berechnet, dass die Verbreitung von aviären Influenzaviren auch entlang der Handelswege von Geflügel erfolgt.[74]
Im Mai 2014 wurde erstmal ein Übergang auf den Menschen bekannt: Bei einem 49-Jährigen aus der Provinz Sichuan im Südwesten Chinas, der an den Folgen der Infektion verstorben war.[75] Eine zweite Infektion wurde im Dezember 2014 in der südchinesischen Provinz Guangdong nachgewiesen,[76] eine dritte im Februar 2015 in der Provinz Yunnan.[77] Zwei weitere Infektionen wurden in China im Dezember 2015 nachgewiesen.[78] Mit Stand vom 19. Dezember 2016 waren der WHO insgesamt 16 gesicherte Erkrankungen beim Menschen bekannt, sechs Erkrankte waren an den Folgen der Infektion verstorben.[79] Bis zum 1. November 2018 erhöhte sich die Anzahl der nachgewiesenen Erkrankungen auf 22 bei insgesamt unverändert sechs Todesfällen,[80] bis Ende September 2019 wurden insgesamt 24 Erkrankungen virologisch bestätigt.[81] Auch danach kam es (Stand: 19. November 2021) zu mehreren Dutzend Neuerkrankungen.[82]
A/H5N7
Der Subtyp A/H5N7 wurde erstmals im Jahr 2003 auf der dänischen Halbinsel Salling in einer Entenzucht nachgewiesen,[83][84] so dass 12.000 Tiere getötet wurden. Eine genaue Analyse des Erbguts ergab eine verwandtschaftliche Nähe zu den Subtypen A/H5N2 und A/H7N7.[85]
A/H5N8
Der erste wissenschaftlich dokumentierte Ausbruch von A/H5N8 geschah im November 1983 in einer Geflügelhaltung in Irland.[86] Damals wurden 8000 Puten und 28.000 Hühnerküken getötet, ferner 270.000 Enten, was 97 Prozent des zu diesem Zeitpunkt in Irland kommerziell gehaltenen Entenbestands entsprach.[87]
Seit 2014 kam es wiederholt zu größeren Ausbrüchen in Mitteleuropa. Im Februar 2021 wurde erstmals eine Übertragung von infiziertem Geflügel auf den Menschen bekannt.[88]
A/H5N9
Der Subtyp A/H5N9 wurde 1966 in Truthühnern in Kanada nachgewiesen[89] (A/Turkey/Ontario/7732/1966 H5N9).[90] Zwei Jahre später trat der Subtyp auch in den USA in Erscheinung.[91] Aus Deutschland kam im Herbst 2014 der Nachweis aus einer abgeschossenen Wildente.[92]
Im Jahr 2015 entdeckte man in der VR China eine reassortierte, hochpathogene Variante des Subtyps, deren Entstehen auf die Subtypen A/H5N1, A/H7N9 und A/H9N2 zurückgeführt werden konnte.[93] Der erste große Ausbruch in Europa ereignete sich im Spätherbst 2015 in mehreren Geflügelbeständen im französischen Département Landes und im Département Dordogne; mehr als 25.000 Stück Geflügel (Perlhühner, Hühner und Enten) wurden vorsorglich getötet.[94]
H6
A/H6N1
Der Subtyp A/H6 wurde erstmals 1965 in Truthühnern entdeckt und kommt heute in Südchina als gering pathogenes Virus häufig in Geflügel vor; beim Menschen wurde der Subtyp A/H6N1 erstmals aus einer im Mai 2013 an Grippe-artigen Symptomen erkrankten Patientin in Taiwan isoliert.[95][96] Molekularbiologische Analysen ergaben, dass es sich bei den aus der Patientin isolierten Viren – A/Taiwan/2/2013(H6N1) – um eine bis dahin aus Geflügel unbekannte Reassortierung handelte, deren Gene unter anderem vom Subtyp A/H5N2 abstammten.[97] Eine weitere molekularbiologische Analyse ergab, dass die Bindungseigenschaften dieser Subtyp-Variante wie bei den zuvor bekannten Varianten an Geflügel angepasst sind und nicht an die Zellen des Menschen.[98]
H7
Von A/H7Nx sind neun Subtypen bekannt.[42]
A/H7N1
Durch den Subtyp A/H7N1 kam es im März 1999 in Italien zu einer massiven Epidemie, in deren Folge bis Anfang 2000 mehr als 13 Millionen Tiere betroffen waren. Eine Übertragung auf den Menschen war nicht nachweisbar.[99] Im Oktober 2016 wurden in Algerien (Distrikt El Ménia) in Feuchtgebieten mehrere hundert tote Wildvögel – vor allem Rostgänse, Marmelenten, Stockenten und Teichrallen – entdeckt, die mit A/H7N1 infiziert waren.[100]
A/H7N2
Der Subtyp A/H7N2 wurde im Jahr 2002 im Verlauf eines Ausbruchs in Geflügelhaltungen in den USA auch auf einen Menschen in Virginia übertragen[101] sowie 2003 auf einen Menschen in New York.[102] 2007 gab es Ausbrüche in Geflügelhaltungen in Wales,[103] 2016 in einer Straußen-Farm in Südafrika.[104]
Mit besonderem Aufwand untersuchten die US-amerikanischen Gesundheitsbehörden im Dezember 2016 einen Ausbruch von A/H7N2 unter Hauskatzen und bei einem Mitarbeiter in einem Tierheim in New York.[105]
A/H7N3
Der Subtyp A/H7N3 wurde erstmals 1963 bei Truthähnen in Großbritannien nachgewiesen. Im April 2006 infizierte dieser Subtyp Puten und einen Mitarbeiter eines Zuchtbetriebs im britischen North Tuddenham in Norfolk.[106]
In Nordamerika wurde die Ausbreitung dieses Subtyps mehrmals bestätigt. Beispielsweise wurden im April 2004 18 Farmen in British Columbia unter Quarantäne gestellt und zwei Fälle von Übertragung auch auf Menschen dokumentiert;[107] beide überstanden die Infektion ohne Folgen. Die Symptome sind ähnlich der einer leichten Grippe.[108][109] Weitere Ausbrüche in Kanada gab es 2007/08 in Saskatchewan,[110] und im August 2014 wurde der Subtyp in den USA (Salem County, Salem, New Jersey) in einem Bestand von 44.000 Stockenten und 7.200 Fasanen nachgewiesen.[111]
2012/13 und erneut 2015 kam es zu Ausbrüchen unter Legehennen in Mexiko;[112][113] 2015 wurden zudem hochpathogene Viren im Nationalpark El Zapotal (Bundesstaat Chiapas) in mehreren Braunflügelguanen und Gilbdrosseln nachgewiesen.[114] Im Januar 2017 kamen 4000 Enten eines Freiland-Zuchtbetrieb in Kambodscha infolge eine LPAI-Infektion mit A/H7N3 zu Tode.[115]
A/H7N4
Der Subtyp A/H7N4 trat laut WHO erstmals 1997 in einer hochpathogenen Variante bei Hühnern in New South Wales (Australien) auf.[116] Dokumentiert sind ferner gleich alte Virusproben aus Emus: A/emu/New South Wales/775/1997(H7N4) und A/emu/NSW/1742/1997(H7N4).
Anfang 2018 wurde in der ostchinesischen Provionz Jiangsu bei einer 68-jährigen Frau erstmals der Übergang dieses Subtyps auf den Menschen nachgewiesen. Die Patientin war an einer schweren Lungenentzündung erkrankt, von der sie sich im Krankenhaus erholte. Bei 28 Kontaktpersonen wurden Abstriche der Rachenschleimhaut gewonnen, die in allen Fällen virusfrei waren.[117][118][119]
A/H7N5
Der Subtyp A/H7N3 wurde laut der Influenza-Datenbank der National Center for Biotechnology Information erstmals 1977 dokumentiert, das Virus wurde damals in Kanada aus Stockenten isoliert (Taxonomy ID in der Influenza Research Database (IRD): 286295), später trat es auch in den USA auf.[120] Übergänge auf den Menschen sind nicht dokumentiert.[121]
A/H7N6
Bereits 1981 wurde die Pathogenität von zwei Varianten des Subtyps A/H7N6 experimentell untersucht, die 1975 bzw. 1976 in Australien nachgewiesen worden waren (A/chicken/Victoria/75 und A/duck/Victoria/76).[122] In der OpenFlu database des Swiss Institute of Bioinformatics ist dieser Subtyp erstmals für das 2007 dokumentiert (A/duck/Yunnan/87/2007); eine enge Verwandtschaft besteht mit dem Subtyp A/H4N6.[123] Außer in China[124] gibt es Nachweise u. a. in Thailand, Südkorea, Japan, Kanada, den USA und in Chile. In Japan wurden beispielsweise im Jahr 2009 mehr als 1,5 Millionen Japanwachteln nach Ausbrüchen in mehreren Farmen getötet,[125] in Chile kamen bei einem großen Ausbruch um die Jahreswende 2016/17 mehr als 35.000 Truthühner zu Tode.[126]
A/H7N7
Der Subtyp A/H7N7 wurde erstmals 1956 in Prag in Pferden nachgewiesen,[127] weswegen die durch die Viren verursachte Erkrankung auch als Pferdegrippe bezeichnet wird.[128]
Diesen Subtyp zählte man zunächst nicht zu den beim Menschen eine Grippe auslösenden Influenza-A-Viren und sah ihn von daher für den Menschen als wenig bedrohlich an. 1996 kam es jedoch zu einer Infektion bei einem Menschen im Vereinigten Königreich, nach dem Aufenthalt in einem Entenstall, von der sich die infizierte Person wieder erholte.[101] 2003 wurden dann in den Niederlanden 89 Infektionen von Menschen mit diesem HPAI-Subtyp bestätigt. Ein Fall verlief tödlich; Opfer war ein Tierarzt, bei dem dieser Virussubtyp im Lungengewebe nachgewiesen werden konnte. Außerdem mussten 30.000 Nutzvögel getötet werden.[107] Ein weiterer Ausbruch in England, in dessen Verlauf mehr als 80.000 Hühner getötet wurden, ereignete sich 2015;[129] ebenfalls 2015 wurden mehr als 10.000 Legehennen im Emsland (Niedersachsen) getötet.[130] Im Juli 2020 wurden aus dem Südosten Australiens Ausbrüche in zwei Freiland-Legehennenbetrieben gemeldet, in denen mehr als 43.000 Tiere gehalten wurden.[131]
2013 wurde in China beim Versuch, die dem Ausbruch der Vogelgrippe H7N9 vorhergehende Reassortierung von Influenza-A-Viren zu rekonstruieren, eine bis dahin unbekannte Variante von A/H7N7 aus frei verkäuflichen Hühnern isoliert, die im Labor auch auf Frettchen übertragbar war; Frettchen gelten bezüglich Influenza als Modellorganismus für den Menschen. Die neue A/H7N7-Variante erwies sich als hoch ansteckend, und die Forscher befürchteten, dass sie auch auf den Menschen übertragbar sein könnte.[132]
A/H7N8
Der Subtyp A/H7N8 wurde wiederholt – als niedrig pathogene Variante – in den USA im Rahmen des Wildvogel-Monitorings nachgewiesen; zu einem Ausbruch in einer Geflügelhaltung kam es erstmals Anfang 2016 im US-Bundesstaat Indiana.[133] Betroffen von der hoch pathogenen Variante[134] war ein Bestand von Truthühnern, mehr als 40.000 Tiere wurden getötet.[135] Bei der anschließenden Untersuchung weiterer Bestände wurde eine fast identische aber niedrig pathogene Variante in mehreren nahegelegenen (klinisch symptomlosen) Beständen gefunden und weitere mehr als 150.000 Tiere getötet.[136]
A/H7N9
Vermutlich nach Kontakt mit infiziertem Geflügel kam es im Februar 2013 erstmals beim Menschen zu Infektionen mit dem Influenza-A-Virus H7N9 und als deren Folgen zu Todesfällen durch die sogenannte Vogelgrippe H7N9 aufgrund schwerer Pneumonien durch eine bis dahin unbekannte, reassortierte Variante des Virus A/H7N9,[137] und zwar in Shanghai und in den chinesischen Provinzen Anhui und Zhejiang.[138][139] Das Virus war bis dahin nur in wenigen Einzelfällen bei symptomfreien Tieren nachgewiesen worden.
Bei der Suche nach einer Infektionsquelle wurden in Shanghai auch Tauben gefunden, die das Virus in sich trugen;[140] Tauben gelten als wenig empfänglich für Erkrankungen durch Influenzaviren.[141]
In seltenen Fällen ist eine Übertragung von Mensch zu Mensch möglich. Allerdings ist der Erreger nur begrenzt übertragbar und es fand bislang keine anhaltende Mensch-zu-Mensch-Übertragung statt.[142][143]
H8
A/H8N1
Der Subtyp A/H8N1 wurde Ende der 1990er-Jahre im Rahmen einer Suche nach potentiell pandemischen Influenzaviren in Sibirien und Japan aus Enten isoliert.[144] 2011 wurde der Subtyp in Mississippi (USA) aus einer Löffelente isoliert – A/northern shoveler/Mississippi/11OS5900/2011(H8N1) – und in der Virendatenbank des National Institute of Allergy and Infectious Diseases der USA unter der Taxon-ID 1460845 registriert.[145]
H9
Von A/H9Nx sind neun Subtypen bekannt (N1 – N9).[42]
A/H9N2
Der Subtyp A/H9N2 ist weltweit verbreitet und hat wiederholt zu großen Ausbrüchen in Geflügelzuchten geführt;[146] auch in Schweinen wurde er nachgewiesen.[147]
In China wird der Subtyp sporadisch beim Menschen nachgewiesen,[148] bislang jedoch nur in einer minder pathogenen Form (LPAI, Lowly Pathogenic Avian Influenza).[149] Bei drei Fällen in Hongkong und China (1999, 2003) erholten sich die Patienten von dieser influenza-ähnlichen Infektion.[101] Auch aus Ägypten[150] und Bangladesch[151] wurden mild verlaufende Erkrankungen bekannt. 2016 wurde erstmals der Verdacht auf einen Todesfall infolge einer Infektion mit A/H9N2 bekannt.[152]
Nachdem sechs Gene für interne Virusproteine von A/H9N2 auf den Subtyp A/H7N9 übergegangen waren, entwickelte A/H7N9 ein stark erhöhtes Gefährdungspotential für den Menschen (→ Vogelgrippe H7N9).[153]
H10
A/H10N3
Der Subtyp A/H10N3 wurde laut Influenza Research Database erstmals 1978 in Kanada bei einer Blauflügelente (A/blue-winged teal/ALB/778/1978) und seitdem wiederholt und vor allem in Vögeln aus China, Thailand und den USA nachgewiesen. Im April 2021 wurde erstmals eine Infektion beim Menschen dokumentiert, und zwar bei einem Mann aus der Stadt Zhenjiang im Osten der Volksrepublik China. Nach Angaben der chinesischen Nationalen Gesundheitskommission vom 1. Juni 2021 war er ins Krankenhaus eingeliefert worden, nachdem er Fieber entwickelt und sich sein Gesundheitszustand verschlechtert hatte. Übertragungen des Virus auf Kontaktpersonen wurden nicht entdeckt, der Mann erholte sich von der Infektion. Auch bei Tieren verursacht der Subtyp der chinesischen Behörde zufolge nur milde Verläufe.[154][155]
A/H10N7
Der Subtyp A/H10N7 wurde erstmals 1949 in Deutschland isoliert (A/chicken/Germany/N/1949(H10N7)). In jüngerer Zeit wurde dieser Subtyp u. a. in den USA bei Truthühnern und Emus nachgewiesen, ferner bei Pekingenten in Südafrika und bei Hühnern in Kanada. 2010 kam es zu einem Ausbruch auf einer Hühnerfarm in Australien, in dessen Folge auch mehrere Farmarbeiter mit diesem LPAI-Virus infiziert wurden und erkrankten.[156]
Im Oktober 2014 wurden an der dänischen Nordseeküste mehrere Dutzend verendete Seehunde angetrieben, die an den Folgen einer H10N7-Infektion verstorben waren.[157] Kurz darauf wurden auch an der deutschen Nordseeküste, vor allem an den Stränden auf Sylt, Helgoland, Amrum und Föhr, mehrere hundert tote Seehunde aufgefunden, die an einer H10N7-Infektion verstorben waren.[158][159]
A/H10N8
Der Subtyp A/H10N8 rief erstmals Ende Dezember 2013 weltweite Aufmerksamkeit hervor, nachdem in Südchina eine 74-jährige Frau an den Folgen einer Infektion mit A/H10N8 verstorben war; wenig später wurde aus dem gleichen Gebiet eine zweite Infektion[160] und später noch eine dritte bekannt; zwei der drei Erkrankten starben.[161] Zuvor war dieser Subtyp vor allem in südchinesischem Geflügel und in Zugvögeln – auch in den USA und in Europa – nachgewiesen worden.[162] Bereits im Juli 2012 war die Sequenzierung des Genoms aus einer chinesischen Virusprobe von A/H10N8 publiziert worden.[163] Sechs seiner Gene sind weitgehend identisch mit denen von A/H9N2, die aus chinesischen Geflügelbeständen bekannt sind, während das Hämagglutinin der Hülle in der menschlichen Lunge aufgrund einer Mutation Bindungseigenschaften aufweise, die es für Menschen vergleichbar infektiös wie A/H5N1 mache;[164][165] gleichwohl sind die Bindungseigenschaften dieser bei den Erkrankten isolierten Subtyp-Varianten – wie die zuvor bekannten Varianten – primär an Geflügel angepasst.[166]
H11
A/H11N2
Der Subtyp A/H11N2 wurde erstmals im Mai 2014 wissenschaftlich beschrieben. Er war in der Antarktis bei Adeliepinguinen nachgewiesen worden, die keine Krankheitssymptome zeigten. Das Genom wies eine verwandtschaftliche Nähe zum Subtyp A/H3N8 aus infizierten Pferden der 1960er-Jahre auf.[167]
H12
H13
A/H13N2
Der Subtyp A/H13N2 (A/Whale/Maine/2/84) wurde 1986 aus der Lunge und gemeinsam mit A/H13N9 aus einem Lymphknoten eines zwei Jahre zuvor an der Küste von Massachusetts, USA, gestrandeten Grindwals isoliert.[172] Die Hämagglutinin-Variante des Subtyps erwies sich als eng verwandt mit der erstmals 1982 beschriebenen H13-Variante[173] aus Möwen, und auch die Neuraminidase-Variante ähnelte dokumentierten Varianten aus diversen Vogelarten. A/H13N2 wurde später auch in Vögeln nachgewiesen.[174]
A/H13N9
Der Subtyp A/H13N9 (A/Whale/Maine/1/84) wurde 1986 gemeinsam mit A/H13N2 aus einem Lymphknoten eines zwei Jahre zuvor an der Küste von Massachusetts, USA, gestrandeten Grindwals isoliert.[172] Die Hämagglutinin-Variante des Subtyps erwies sich als eng verwandt mit der erstmals 1982 beschriebenen H13-Variante[173] aus Möwen, und auch die Neuraminidase-Variante ähnelte dokumentierten Varianten aus diversen Vogelarten. A/H13N9 wurde später auch in Wasservögeln nachgewiesen.[175]
H14 bis H18
A/H14Nx
Die ersten Subtypen von A/H14 – A/H14N5 und A/H14N6 – wurden 1982 im Gebiet von Astrachan am Kaspischen Meer aus Silbermöwen und Stockenten isoliert. Im Jahr 2010 wurden erstmals außerhalb dieser Region – in Wisconsin, USA – Varianten beider Subtypen und zusätzlich die Subtypen A/H14N4 und A/H14N8 nachgewiesen.[176] Auch in den Folgejahren gab es in Nord- und Südamerika weitere Funde dieser Subtypen, in Guatemala wurde zudem der Subtyp A/H14N3 entdeckt.[177]
A/H15Nx
Der erste Nachweis des Subtyps A/H15 wurde im Jahr 1996 publiziert, anhand von zwei Belegen aus Australien, die bereits 1979 (A/H15N9) und 1983 (A/H15N8) aus einer Ente und aus einem zweiten Küstenvogel gewonnen worden waren; das A/H15-Hämagglutinin ist am engsten verwandt mit dem A/H7-Hämagglutinin.[178]
Aus einer im März 2013 publizierten Studie geht hervor, dass bis dahin auch die Subtypen A/H15N2 und A/H15N4 bekannt waren.[179]
A/H16N3
Der erste Nachweis des Subtyps A/H16 wurde im Jahr 2005 publiziert, anhand von Belegen aus Schweden, die 1999 aus Lachmöwen gewonnen worden waren; das A/H16-Hämagglutinin ist entfernt verwandt mit dem A/H13-Hämagglutinin.[180] Der Erstnachweis wurde A/black-headed gull/Sweden/2/99 benannt und als A/H16N3 identifiziert; A/H16N3 wurde laut Influenza Research Database in den folgenden Jahren wiederholt auch in den Niederlanden und in Alaska nachgewiesen und ist auch aus Asien belegt.[181]
A/H17N10
Der Subtyp A/H17N10 wurde erstmals 2012 wissenschaftlich beschrieben, nachdem er in Guatemala aus Fruchtfledermäusen der Art Sturnira lilium isoliert worden war.[182] Genetische Experimente mit künstlich veränderten Viren wurden im Jahr 2014 dahingehend interpretiert, dass ein Übergang von H17N10-Viren auf den Menschen zwar nicht auszuschließen, aber unwahrscheinlich sei,[183][184] weil A/H17N10 – im Unterschied zu allen zuvor bekannten Subtypen – nicht an Sialinsäuren binde.[185] 2019 wurde jedoch nachgewiesen, dass H17N10-Viren MHC-II-Moleküle als „Eintrittspforten“ in Zellen von Menschen, Hühnern, Schweinen und Mäusen nutzen können.[186] Siehe auch A. Banerjee et al. (2020).[187]
A/H18N11
Der Subtyp A/H18N11 wurde erstmals im Oktober 2013 wissenschaftlich beschrieben, nachdem die genomische Sequenz des Virus in Peru aus Fruchtfledermäusen der Art Artibeus planirostris isoliert worden war.[188] In einer im Jahr 2019 veröffentlichen Screening-Studie wurden in der verwandten Art Artibeus lituratus aus Brasilien ebenfalls genomische Sequenzen von A/H18N11 isoliert.[189] A/H18N11-Viren nutzen, genauso wie der A/H17N10-Subtyp, MHC-II-Moleküle verschiedenster Organismen, um eine Zelle zu infizieren.[190] Die experimentelle Charakterisierung des A/H18N11 Subtyps legt nahe, dass dessen Fähigkeit, andere Arten als Fledermäuse zu infizieren, als eher gering einzustufen ist.[191] Siehe auch A. Banerjee et al. (2020).[187]
Siehe auch
Literatur
- David E. Swayne (Hrsg.): Avian Influenza. Blackwell Publishing, 2008, ISBN 978-0-8138-2047-7.
- Robert G. Webster et al.: Evolution and ecology of influenza A viruses. In: Microbiology and Molecular Biology Reviews. Band 58, Nr. 1, 1992, S. 152–179, Volltext (PDF).
Weblinks
- Influenza Research Database (engl.)
- Avian Influenza auf den Webseiten der Weltorganisation für Tiergesundheit (engl.)
- RespVir – Netzwerk für respiratorische Viren und Bakterien mit Angaben zur Häufigkeitsverteilung und aktueller epidemiologischer Lage in Deutschland
Einzelnachweise
- UniProt-Datenbank: Taxonomy – Influenza A virus (SPECIES).
- Michael Worobey et al.: A synchronized global sweep of the internal genes of modern avian influenza virus. In: Nature. Band 508, 2014, S. 254–257, ISSN 1476-4687. doi:10.1038/nature13016. PMID 24531761.
Study on flu evolution may change textbooks, history books. Auf: eurekalert.org vom 16. Februar 2014 - Warning signals from the volatile world of influenza viruses. Auf: who.int vom Februar 2015.
- G. Koch (Wageningen Bioveterinary Research, Februar 2014): Epidemiology of avian influenza. (Memento vom 28. Februar 2018 im Internet Archive)
- Richard E. Shope: Swine Influenza: III. Filtration and Ion Experiments and Etiology. In: Journal of Experimental Medicine. Band 54, 1931, S. 373–385.
- What Is Swine Flu? (Memento vom 4. März 2010 im Internet Archive) Suburban Emergency Management Project: Biot Report 162 vom 9. Januar 2005
- Julie L. McAuley et al.: Expression of the 1918 Influenza A Virus PB1-F2 Enhances the Pathogenesis of Viral and Secondary Bacterial Pneumonia. In: Cell Host & Microbe. Band 2, Nr. 4, 2007, S. 240–249, doi:10.1016/j.chom.2007.09.001
- Vogelgrippe (aviäre Influenza). Informationsschrift, erstellt in Zusammenarbeit mit Prof. Hans-Peter Seelig. Stand: Oktober 2005 Auf: labor-clotten.de. Abgerufen am 5. Dezember 2018.
- Resistance to Oseltamivir (Tamiflu) found in some European influenza virus samples. European Centre for Disease Prevention and Control (ECDC), 20. September 2008
- WHO: Influenza A(H1N1) virus resistance to oseltamivir - 2008 influenza season, southern hemisphere. (Memento vom 9. August 2009 im Internet Archive) (PDF; 29 kB) 18. Juli 2008
- Prevalence of Oseltamivir-resistant H1N1 viruses. Last quarter 2008 – First quarter 2009. (Memento vom 9. August 2009 im Internet Archive) Grafik der WHO.
- WHO, 4. Juni 2009: Influenza A virus resistance to oseltamivir and other antiviral medicines.; vergl. dazu: WHO Guidelines for Pharmacological Management of Pandemic Influenza A(H1N1) 2009 and other Influenza Viruses (PDF; 553 kB), Februar 2010, S. 8/unten
- Ignacio Mena et al.: Origins of the 2009 H1N1 influenza pandemic in swine in Mexico. In: eLife. Online-Veröffentlichung vom 28. Juni 2016, doi:10.7554/eLife.16777
- Honglei Sun et al.: Prevalent Eurasian avian-like H1N1 swine influenza virus with 2009 pandemic viral genes facilitating human infection. In: PNAS. Online-Vorabveröffentlichung vom 29. Juni 2020, doi:10.1073/pnas.1921186117.
DER SPIEGEL: Neuer Schweinegrippe-Typ in China entdeckt - DER SPIEGEL - Wissenschaft. Abgerufen am 30. Juni 2020. - Neue Art der Schweinegrippe mit Pandemiepotenzial. Auf: science.orf.at vom 30. Juni 2020.
- H1N2 Variant Virus Detected in Minnesota. Auf: cdc.gov vom 7. September 2012
- Tapasi Roy Mukherjee et al.: Full genomic analysis of an influenza A (H1N2) virus identified during 2009 pandemic in Eastern India: evidence of reassortment event between co-circulating A(H1N1)pdm09 and A/Brisbane/10/2007-like H3N2 strains. In: Virology Journal. Band 9, 2012, S. 233, doi:10.1186/1743-422X-9-233
- Naomi Komadina et al.: A Historical Perspective of Influenza A(H1N2) Virus. In: Emerging Infectious Diseases. Band 20, Nr. 1, 2014, doi:10.3201/eid2001.121848, Volltext
- Influenza at the human-animal interface. Auf: who.int vom 19. Dezember 2016, S. 5
- Hintergrundinformation zur Schweineinfluenza. (Memento vom 27. Dezember 2016 im Internet Archive) Auf: virologie.uniklinikum-jena.de
- Supassama Chaiyawong et al.: Genetic characterization of influenza A virus subtypes H1N3 and H1N9 isolated from free-grazing ducks in Thailand. In: Archives of Virology. Band 161, Nr. 10, 2016, S. 2819–2824, doi: 10.1007/s00705-016-2962-0
- Taxonomy: Influenza A virus, strain A/Whale/Pacific ocean/19/1976 H1N3 Auf: uniprot.org, eingesehen am 27. Dezember 2016
- Michael Worobey et al.: Genesis and pathogenesis of the 1918 pandemic H1N1. In: PNAS. Band 111, Nr. 22, 2014, S. 8107–8112, 2014, doi:10.1073/pnas.1324197111, Volltext
- UniProt-Datenbank: H2N1 subtype.
- Pandemic-causing 'Asian flu' accidentally released. In: New Scientist vom 13. April 2005.
- UniProt-Datenbank: H2N3 subtype.
- Daniela S. Rajãoa et al.: Novel Reassortant Human-Like H3N2 and H3N1 Influenza A Viruses Detected in Pigs Are Virulent and Antigenically Distinct from Swine Viruses Endemic to the United States. In: Journal of Virology. Band 89, Nr. 22, 2015, S. 11213–11222, doi:10.1128/JVI.01675-15
Jin-Young Shin et al.: Isolation and Characterization of Novel H3N1 Swine Influenza Viruses from Pigs with Respiratory Diseases in Korea. In: Journal of Clinical Microbiology. Band 44, Nr. 11, 2006, S. 3923–3927, doi:10.1128/JCM.00904-06
Ana Moreno et al.: Novel swine influenza virus subtype H3N1 in Italy. In: Veterinary Microbiology. Band 138, Nr. 3–4, 2009, S. 361–367, doi:10.1016/j.vetmic.2009.04.007 - N. Siddique et al.: Isolation, identification, and phylogenetic analysis of reassortant low-pathogenic avian influenza virus H3N1 from Pakistan. In: Poultry Science. Band 91, 2012, S. 129–138, doi:10.3382/ps.2011-01530
- Ching-Ping Tsai und Ming-Jeng Pan: New H1N2 and H3N1 influenza viruses in Taiwanese pig herds. In: The Veterinary Record. Band 153, Nr. 13, 2003, S. 408.
- H3N2 in der Influenza Research Database.
- Increase in Influenza A H3N2v Virus Infections in Three U.S. States. (Memento vom 1. April 2014 im Internet Archive) Centers for Disease Control and Prevention: Official CDC Health Advisory vom 3. August 2012.
- Charakterisierung der zirkulierenden Viren und Übereinstimmung mit den im Impfstoff enthaltenen Stämmen. Auf: rki.de, eingesehen am 29. Januar 2017
- Charakterisierung der Influenzaviren in: Robert Koch-Institut: Influenza-Wochenbericht der AGI-Studiengruppe, Kalenderwoche 9 (25.02. bis 03.03.2017), Seite 4, (PDF; 167 kB)
- FAQ about the H3N2 strain of canine influenza. (Memento vom 16. Juni 2016 im Internet Archive) Publiziert auf dem Webserver der Cornell University, Stand: April 2015
- Harrison Powell und Andrew Pekosz: Neuraminidase antigenic drift of H3N2 clade 3c.2a viruses alters virus replication, enzymatic activity and inhibitory antibody binding. In: PLoS Pathogens. Band 16, Nr. 6, 2020, e1008411. doi:10.1371/journal.ppat.1008411.
Fast-Spreading Mutation Helps Common Flu Subtype Evade Immune Response and Vaccines. Auf: SciTechDaily.com vom 10. Juli 2020. - Influenza A virus (A/seal/MA/3984/1992(H3N3)). Auf: uniprot.org, zuletzt abgerufen am 23. September 2015.
- Influenza A virus (A/mallard duck/Minnesota/Sg-00100/2007(H3N3)). Auf: uniprot.org, zuletzt abgerufen am 23. September 2015.
- Alexander I. Karasin et al.: Characterization of Avian H3N3 and H1N1 Influenza A Viruses Isolated from Pigs in Canada. In: Journal of Clinical Microbiology. Band 42, Nr. 9, 2004, S. 4349–4354, doi:10.1128/JCM.42.9.4349-4354.2004.
- T. Jamanaka et al.: Interspecies transmission of equine influenza virus (H3N8) to dogs by close contact with experimentally infected horses. In: Veterinary Microbiology. Band 139, Nr. 3–4, 2009, S. 351–355, doi:10.1016/j.vetmic.2009.06.015, PMID 19596528
- S. J. Anthony et al.: Emergence of Fatal Avian Influenza in New England Harbor Seals. In: mBio. Band 3, Nr. 4, 2012, e00166-12, doi:10.1128/mBio.00166-12
- Yong Hu et al.: Complete genome sequence of a novel H4N1 influenza virus isolated from a pig in central China. In: Journal of Virology. Band 86, Nr. 24, 2012, S. 13879, doi:10.1128/JVI.02726-12, Volltext (PDF)
- Influenza Type A Viruses. Auf: cdc.gov, Stand 19. April 2017, eingesehen am 26. Februar 2018
- Dennis J. Alexander: A review of avian influenza in different bird species. In: Veterinary Microbiology. Band 74, 2000, S. 3–13, Volltext (PDF) (Memento vom 26. Juni 2013 im Internet Archive)
- Y. Kawaoka et al.: Is virulence of H5N2 influenza viruses in chickens associated with loss of carbohydrate from the hemagglutinin? In: Virology. Band 139, Nr. 2, 1984, S. 303–316, doi:10.1016/0042-6822(84)90376-3.
- T. Horimoto et al.: Origin and molecular changes associated with emergence of a highly pathogenic H5N2 influenza virus in Mexico. In: Virology. Band 213, Nr. 1, 1995, S. 223–230, doi:10.1006/viro.1995.1562
- Immediate notification report: CAN-2015-NAI-002 REF OIE 17483. (Memento vom 10. April 2015 im Internet Archive) Auf: oie.int vom 8. April 2015.
- Mara Hvistendahl: Enigmatic bird flu strain races across the U.S. Midwest. In: Science. Band 348, Nr. 6236, 2015, S. 741–742, doi:10.1126/science.348.6236.741
- Dozens In Japan May Have Mild Bird Flu. Auf: cbsnews.com vom 10. Januar 2006.
- Immediate notification report: REF OIE 16936, Report Date: 12/01/2015, Country: Chinese Taipei. (Memento vom 13. Januar 2015 im Internet Archive) Auf: oie.int vom 12. Januar 2015.
- Immediate notification report: reference 15-1 H5N2 LPAI REF OIE 17494, Report Date: 22/04/2015, Country: Chinese Taipei (Memento vom 23. September 2015 im Internet Archive)
- Wildvögel sind unschuldig: Ausbreitung der Vogelgrippe durch Menschen oder Putenküken! (Memento vom 17. Dezember 2014 im Internet Archive) Pressemitteilung der Arbeitsgemeinschaft für artgerechte Nutztierhaltung e. V. vom 12. Dezember 2008. Abgerufen am 23. September 2015
- Follow-up report No. 15 (Final report), Country: South Africa. (Memento vom 29. März 2015 im Internet Archive)
- Immediate notification report: REF OIE 19292, Report Date: 08/12/2015. (Memento vom 9. Dezember 2015 im Internet Archive) Auf: oie.int vom 8. Dezember 2015
Geflügelpest-Verdacht: Erregertyp steht fest. Freie Presse, 6. Dezember 2015, abgerufen am 6. Dezember 2015. - Immediate notification report: LPAI H5 REF OIE 21648. (Memento vom 25. November 2016 im Internet Archive) Auf: oie.int vom 23. November 2016
- W. B. Becker: The isolation and classification of tern virus influenza virus A/Tern/South Africa/1961. In: Journal of Hygiene. Band 64, 1966, S. 309
- Geflügelpest im Landkreis Cloppenburg weitet sich aus. (Memento vom 20. Dezember 2013 im Internet Archive) Ärzte Zeitung online, 15. Dezember 2008.
- Vogelgrippe: Fragen an die Bundesregierung. Auf: hamburger-fortbildungstage.de, unter Bezug auf den Landkreis Cloppenburg. Pressemitteilung vom 20. Januar 2009
- OIE: Immediate notification report. LPAI H5N3 REF OIE 22550, Report Date: 24/01/2017, Country: Germany. (Memento vom 30. Januar 2017 im Internet Archive) Auf: oie.int vom 24. Januar 2017
- H5N4 in der „Influenza Research Database“.
- Min Gu et al.: Novel Reassortant Highly Pathogenic Avian Influenza (H5N5) Viruses in Domestic Ducks, China. In: Emerging Infectious Diseases. Band 17, Nr. 6, 2011, doi:10.3201/eid1706.101406
- Wei Zou et al.: Complete Genome Sequence of a Novel Natural Recombinant H5N5 Influenza Virus from Ducks in Central China. In: Journal of Virology. Band 86, Nr. 24, 2012, doi:10.1128/JVI.02725-12
- Kunkun Zhao et al.: Characterization of three H5N5 and one H5N8 highly pathogenic avian influenza viruses in China. In: Veterinary Microbiology. Band 163, Nr. 3–4, 2013, S. 351–357, doi:10.1016/j.vetmic.2012.12.025
- CG Liu et al.: Emerging multiple reassortant H5N5 avian influenza viruses in ducks, China, 2008. In: Veterinary Microbiology. Band 167, Nr. 3–4, 2013, S. 296–306, doi:10.1016/j.vetmic.2013.09.004
- OIE: Immediate notification report. HPAI 2016/1 H5N5 REF OIE 21909, Report Date:14/12/2016, Country: Netherlands (Memento vom 20. Dezember 2016 im Internet Archive) vom 14. Dezember 2016
- OIE: Immediate notification report. REF OIE 22606, Report Date: 27/01/2017, Country: Greece. (Memento vom 30. Januar 2017 im Internet Archive) Auf: oie.int vom 27. Januar 2017
- OIE: Immediate notification report. REF OIE 22678, Report Date: 03/02/2017, Country: Poland. (Memento vom 3. Februar 2017 im Internet Archive) Auf: oie.int vom 3. Februar 2017
- OIE: Immediate notification report. REF OIE 22560, Report Date: 24/01/2017, Country: Germany. (Memento vom 30. Januar 2017 im Internet Archive) Auf: oie.int vom 24. Januar 2017
- H5N5 Geflügelpest: Es ist noch nicht vorbei. Auf: kn-online.de vom 23. Januar 2017
- John C. Obenauer et al.: Large-Scale Sequence Analysis of Avian Influenza Isolates. In: Science. Band 311, Nr. 5767, 2006, S. 1576–1580, doi:10.1126/science.1121586
- Weltgesundheitsorganisation: Influenza at the human-animal interface. Summary and assessment as of 2 October 2014.
- Information received on 02/09/2014 from Dr Zhang Zhongqui, Director General, China Animal Disease Control Centre, Veterinary Bureau, Ministry of Agriculture, Beijing, China. (Memento vom 23. September 2015 im Internet Archive) Mitteilung der World Organisation for Animal Health (OIE) vom 1. September 2014
- OIE: Immediate notification report (Memento vom 17. April 2015 im Internet Archive) vom 16. April 2015, Report reference: REF OIE 17512.
- OIE: Immediate notification report (Memento vom 5. Mai 2015 im Internet Archive) vom 5. Mai 2015, Report reference: REF OIE 17648.
- Qiqi Yang, Xiang Zhao et al.: Assessing the role of live poultry trade in community-structured transmission of avian influenza in China. In: PNAS. Online-Vorabveröffentlichung vom 2. März 2020, doi:10.1073/pnas.1906954117.
Avian influenza and live poultry trade in China. Auf: eurekalert.org vom 2. März 2020. - Sichuan man dies in first human case of H5N6 bird flu. Auf: South China Morning Post vom 7. Mai 2014.
- Weltgesundheitsorganisation: Influenza at the human-animal interface. Summary and assessment as of 6 January 2015.
- Influenza at the human-animal interface. Summary and assessment as of 3 March 2015. Auf: who.int vom 3. März 2015.
- Human infection with avian influenza A(H5N6) virus – China. Auf: who.int vom 4. Januar 2016
- Influenza at the human-animal interface. Auf: who.int vom 19. Dezember 2016
- Influenza at the human – animal interface. Auf: who.int vom 25. Januar 2018
Influenza at the human – animal interface. Auf: who.int vom 1. November 2018 - Influenza at the human-animal interface. Summary and assessment, from 25 June 2019 to 27 September 2019.
- Assessment of risk associated with influenza A(H5N6) virus. Auf: who.int vom 19. November 2021.
- Anders Fomsgaard et al.: New avian influenza virus A H5N7 identified in ducks in Denmark. In: Eurosurveillance. Band 7, Nr. 39, Artikel 3, 2003, Volltext
- New influenza virus identified for the first time in the world at Statens Serum Institut. (Memento vom 26. Februar 2004 im Internet Archive) Publiziert am 19. September 2003
- K. Bragstad et al.: New avian influenza A virus subtype combination H5N7 identified in Danish mallard ducks. In: Virus Research. Band 109, Nr. 2, 2005, S. 181–190, doi:10.1016/j.virusres.2004.12.004
- Influenza Strain Details for A/turkey/Ireland/?/1983(H5N8). Auf: fludb.org - Influenza Research Database, aufgerufen am 6. November 2014.
- Dennis J. Alexander et al.: Highly pathogenic avian influenza outbreaks in Europe, Asia, and Africa since 1959, excluding the Asian H5N1 virus outbreaks. In: David E. Swayne (Hrsg.): Avian Influenza. Blackwell Publishing, 2008, S. 223, ISBN 978-0-8138-2047-7
- Virus H5N8 in Russland: Vogelgrippe erstmals auf Menschen übertragen. Auf: tagesschau.de vom 20. Februar 2021.
- Avian influenza A(H5N1) – update 31: Situation (poultry) in Asia: need for a long-term response, comparison with previous outbreaks. Auf: who.int vom 2. März 2004
- Taxonomy – Influenza A virus (strain A/Turkey/Ontario/7732/1966 H5N9). Auf: uniprot.org, eingesehen am 10. Dezember 2015
- Influenza Strain Details for A/turkey/Wisconsin/1/1968(H5N9). Auf: fludb.org, eingesehen am 10. Dezember 2015
- NRW-Ministerium hebt Stallpflicht auf – Geflügelhalter werden um Vorsicht gebeten. Auf: lokalkompass.de vom März 2015
- Yang Yu et al.: Newly-emergent highly pathogenic H5N9 subtype avian influenza A virus. In: Journal of Virology. Online-Vorabveröffentlichung vom 17. Juni 2015, doi:10.1128/JVI.00653-15
- Immediate notification report. REF OIE 19296, Report Date: 08/12/2015, Country: France. (Memento vom 10. Dezember 2015 im Internet Archive) Auf: oie.int vom 8. Dezember 2015
- Sung-Hsi Wei et al.: Human infection with avian influenza A H6N1 virus: an epidemiological analysis. In: The Lancet. Band 1, Nr. 10, 2013, S. 771–778, doi:10.1016/S2213-2600(13)70221-2
- Novel H6N1 bird flu jumps to humans. Auf: clinicaladvisor.com vom 15. November 2013
- Jian Yuan et al.: Origin and Molecular Characteristics of a Novel 2013 Avian Influenza A(H6N1) Virus Causing Human Infection in Taiwan. In: Clinical Infectious Diseases. Band 57, Nr. 9, 2013, S. 1367–1368, doi: 10.1093/cid/cit479
- Netanel Tzarum et al.: Structure and Receptor Binding of the Hemagglutinin from a Human H6N1 Influenza Virus. In: Cell Host & Microbe. Band 17, Nr. 3, 2015, S. 369–376, doi:10.1016/j.chom.2015.02.005
- Ilaria Capua et al.: The 1999–2000 avian influenza (H7N1) epidemic in Italy: veterinary and human health implications. In: Acta Tropica. Band 83, Nr. 1, 2002, S. 7–11, doi:10.1016/S0001-706X(02)00057-8
- OIE: Immediate notification report. REF OIE 21433, Report Date: 02/11/2016, Country: Algeria (Memento vom 22. November 2016 im Internet Archive)
- Avian Influenza A Virus Infections of Humans / Instances of Avian Influenza A Virus Infections of Humans. Auf: cdc.gov vom 23. Mai 2008
- H7N2 in New York, 2003. Auf: cdc.gov vom 17. Februar 2006
- Avian influenza A/(H7N2) in the United Kingdom. Auf: who.int vom 29. Mai 2007
- OIE: Immediate notification report (Memento vom 28. Oktober 2016 im Internet Archive) vom 24. Oktober 2016, Report reference: LPAI_H7N2_2016 REF OIE 21341
- Avian Influenza A (H7N2) in Cats in Animal Shelters in NY; One Human Infection. Auf: cdc.gov vom 22. Dezember 2016
- Poultry workers free of bird flu. Auf: news.bbc.co.uk vom 30. April 2006
- Für zahlreiche Verweise siehe: Scott Krauss, Robert G. Webster: Predicting the Next Influenza Virus. In: Science. Band 337, Nr. 6095, 2012, S. 644, doi:10.1126/science.337.6095.644-a
- S. Aleina Tweed, Danuta M. Skowronski, Samara T. David, Andrew Larder, Martin Petric, Wayne Lees et al.: Human Illness from Avian Influenza H7N3, British Columbia. In: Emerging Infectious Diseases. Band 10, Nr. 12, November 2004, ISSN 1080-6059 (Volltext [abgerufen am 9. Januar 2013]).
- Martin Hirst, Caroline R. Astell, Comments Malachi Griffith, Shaun M. Coughlin, Michelle Moksa, Thomas Zeng et al.: Novel Avian Influenza H7N3 Strain Outbreak, British Columbia. In: Emerging Infectious Diseases. Band 10, Nr. 12, November 2004, PMC 3323367 (freier Volltext).
- Informe de seguimiento nº: 4. Referencia del informe: CAN-AI-2007-01, OIE Ref: 6690. (PDF; 67 kB) Auf: oie.int vom 18. April 2008
- OIE: Low pathogenic avian influenza (poultry), United States of America. (Memento vom 18. September 2014 im Internet Archive) Information received on 05/09/2014 from Dr John Clifford, Deputy Administrator, Animal and Plant Health Inspection Service, United States Department of Agriculture, Washington, United States of America
- fao.org (PDF; 926 kB): Empress Watch. Band 26, August 2012: Highly Pathogenic Avian Influenza in Mexico (H7N3).
OIE: Immediate notification report. Report reference: REF OIE 12839. (PDF; 49 kB) (Memento vom 23. Januar 2013 im Internet Archive) Auf: oie.int vom 8. Januar 2013 - OIE: Immediate notification report. Report reference: REF OIE 17489. (Memento vom 10. April 2015 im Internet Archive) Auf: oie.int vom 8. April 2015
- OIE: Immediate notification report. Report reference: REF OIE 17686, Report Date: 08/05/2015, Country: Mexico. (Memento vom 12. Mai 2015 im Internet Archive) Auf: oie.int vom 8. Mai 2015
- OIE: Immediate notification report. REF OIE 22599, Report Date: 27/01/2017, Country: Cambodia. (Memento vom 30. Januar 2017 im Internet Archive) Auf: oie.int vom 27. Januar 2017
- Avian influenza A(H5N1)- update 31: Situation (poultry) in Asia: need for a long-term response, comparison with previous outbreaks. Auf: who.int vom 2. März 2004.
- Human infection with avian influenza A(H7N4) virus – China. Auf: who.int vom 22. Februar 2018
- Kwok-Yung Yuen: Another avian influenza A subtype jumping into human: this time is H7N4. In: Science Bulletin. Band 63, Nr. 16, 2018, S. 1025–1026, doi:10.1016/j.scib.2018.08.002
- Xiang Huo, Lun-biao Cui, Cong Chen et al.: Severe human infection with a novel avian-origin influenza A(H7N4) virus. In: Science Bulletin. Band 63, Nr. 16, 2018, S. 1043–1050, doi:10.1016/j.scib.2018.07.003
- A/H7N5 in der Influenza Research Database.
- cdc.gov: Influenza Type A Viruses and Subtypes. Stand vom 2. April 2013, eingesehen am 20. Januar 2015.
- H. A. Westbury, A. J. Turner und L. Amon: Transmissibility of two avian influenza a viruses (H7 N6) between chickens. In: Avian Pathology. Band 10, Nr. 4, 1981, S. 481–487, doi:10.1080/03079458108418498, Volltext (PDF)
- Robert J. Dusek et al.: North Atlantic Migratory Bird Flyways Provide Routes for Intercontinental Movement of Avian Influenza Viruses. In: PLoS ONE. Band 9, Nr. 3, 2014, e92075, doi:10.1371/journal.pone.0092075, Abb. 18, Volltext
- Haibo Wu et al.: Molecular characterization of a novel reassortant H7N6 subtype avian influenza virus from poultry in Eastern China, in 2016. In: Archives of Virology. Band 162, Nr. 5, 2017, S. 1341–1347, doi:10.1007/s00705-017-3219-2
- Katsuaki Sugiura et al.: An outbreak of H7N6 low pathogenic avian influenza in quails in Japan. In: Veterinaria Italiana. Band 45, Nr. 4, 2009, S. 481–489, Volltext
- Low pathogenic avian influenza (poultry), Chile. (Memento vom 26. Februar 2018 im Internet Archive) Mitteilung der División de Protección Pecuaria, Servicio Agrícola y Ganadero (SAG), Ministerio de Agricultura, Santiago, Chile, an die OIE, 18. Januar 2017
- O. Sovinova et al.: Isolation of a virus causing respiratory disease in horses. In: Acta Virol. Bd. 2, 1958, S. 52–61.
- Scott Krauss, Robert G. Webster: Predicting the next Influenza Virus. In: Science. Band 337, Nr. 6095, 2012, S. 644, doi:10.1126/science.337.6095.644-a
- OIE: Immediate notification report. (PDF) (Memento vom 15. Juli 2015 im Internet Archive) Report reference: AIV 2015/02 REF OIE 18116, Report Date: 13/07/2015, Country: United Kingdom
- OIE: Immediate notification report. (PDF) (Memento vom 28. Juli 2015 im Internet Archive) Report reference: 15-015-00006 REF OIE 18234, Report Date: 27/07/2015, Country: Germany
- Highly pathogenic avian influenza, Australia. (Memento vom 31. Juli 2020 im Internet Archive) Mitteilung des Department of Agriculture, Water and the Environment, Australian Government, vom 31. Juli 2020 an die OIE.
- Tommy Tsan-Yuk Lam: The genesis and source of the H7N9 influenza viruses causing human infections in China. In: Nature. Band 502, 2013, S. 241–244, doi:10.1038/nature12515
H7N7-Viren: Die neue Vogelgrippe hat einen potenziell gefährlichen Verwandten. Auf: zeit.de vom 21. August 2013, abgerufen am 25. September 2015. - Avian Influenza H7N8 Update. Auf: cdc.gov vom 19. Januar 2016
- Outbreaks of H7N8 avian influenza in poultry in the USA. Auf: gov.uk vom 19. Januar 2016
- Report reference: REF OIE 19540, Report Date: 15/01/2016, Country: United States of America. (Memento vom 22. Januar 2016 im Internet Archive)
- OIE: Immediate notification report. (PDF) (Memento vom 22. Januar 2016 im Internet Archive) Report reference: REF OIE 19561, Report Date: 19/01/2016, Country: United States of America
- OIE: Low pathogenic avian influenza (poultry), China (People's Rep. of). (Memento vom 2. Februar 2014 im Internet Archive) Information received on 04/04/2013 from Dr Zhang Zhongqui, Director General, China Animal Disease Control Centre, Veterinary Bureau, Ministry of Agriculture, Beijing, China (People's Rep. of). Auf: oie.int vom 4. April 2013
- H7N9 avian influenza human infections in China. Auf: who.int vom 1. April 2013
Human infection with influenza A(H7N9) virus in China - update. Auf: who.int vom 4. April 2012
Neuer Vogelgrippe-Typ ruft WHO auf den Plan. Auf: sueddeutsche.de vom 2. April 2013
straitstimes.com (Memento vom 4. April 2013 im Internet Archive) vom 3. April 2013: Man dies of H7N9 bird flu in China, third fatality from lesser-known strain. - China steps up monitoring after more H7N9 bird flu cases. Auf: news.xinhuanet.com vom 4. Februar 2013, zuletzt abgerufen am 25. September 2015
- OIE: Low pathogenic avian influenza (poultry), China (People's Rep. of). (Memento vom 2. Februar 2014 im Internet Archive) Auf: oie.int vom 4. April 2013
H7N9-Virus: Neue Grippe-Todesfälle – China keult Geflügel. Auf: spiegel.de vom 5. April 2013 - Untersuchungen zur Empfänglichkeit von Tauben für Aviäre Influenza. (Memento vom 22. Februar 2014 im Internet Archive) Friedrich-Loeffler-Institut, abgerufen am 25. September 2015
- Xian Qi, Yan-Hua Qian, Chang-Jun Bao u. a.: Probable person to person transmission of novel avian influenza A (H7N9) virus in Eastern China, 2013: epidemiological investigation. In: British Medical Journal. (BMJ) 06 August 2013, Band 347, f 4752, doi:10.1136/bmj.f4752
- Das RKI zu humanen Erkrankungsfällen mit aviärer Influenza A(H7N9). Stand: 24. Mai 2018. Auf: rki.de; zuletzt abgerufen am 5. Dezember 2018.
- K. Okazaki et al.: Precursor genes of future pandemic influenza viruses are perpetuated in ducks nesting in Siberia. In: Arch virol. Band 145, 2000, S. 885–893, doi:10.1007/s007050050681
- Eintrag von A/H8N1 in der Virendatenbank des National Institute of Allergy and Infectious Diseases der USA
- Rui Wu et al.: Generation and evaluation of an H9N1 influenza vaccine derived by reverse genetics that allows utilization of a DIVA strategy for control of H9N2 avian influenza. In: Archives of Virology. Band 154, Nr. 8, 2009, S. 1203–1210, doi:10.1007/s00705-009-0425-6
- Yan L. Cong et al.: Antigenic and genetic characterization of H9N2 swine influenza viruses in China. In: Journal of General Virology. Band 88, 2007, S. 2035–2041, doi:10.1099/vir.0.82783-0
- Hong Kong sees first case of H9N2 avian flu in four years. South China Morning Post, 30. Dezember 2013
- Malik Peiris et al.: Human infection with influenza H9N2. In: The Lancet. Band 354, Nr. 9182, 1999, S. 916–917, doi:10.1016/S0140-6736(99)03311-5
- Influenza at the human-animal interface. Summary and assessment as of 23 June 2015. Auf: who.int vom 23. Juni 2015 (PDF)
- Influenza at the human-animal interface. Summary and assessment as of 4 September 2015. Auf: who.int vom 4. September 2015
- Influenza at the human-animal interface. Summary and assessment, 20 July to 3 October 2016. Auf: who.int vom 3. Oktober 2016
- Juan Pu et al.: Evolution of the H9N2 influenza genotype that facilitated the genesis of the novel H7N9 virus. In: PNAS. Online-Vorabveröffentlichung vom 29. Dezember 2014, doi:10.1073/pnas.1422456112
- China weist erstmals Infektion mit Vogelgrippe-Variante H10N3 nach. Auf: aerzteblatt.de vom 1. Juni 2021.
- Mitteilung der Nationalen Gesundheitskommission der VR China vom 1. Juni 2021. (chinesisch)
- George G. Arzey et al.: Influenza Virus A (H10N7) in Chickens and Poultry Abattoir Workers, Australia. In: Emerging Infectious Deseases. Band 18, Nr. 5, 2012, doi:10.3201/eid1805.111852
- Forscher lösen Rätsel um Seehundsterben. Auf: ndr.de vom 20. Oktober 2014
- Seehunde im Wattenmeer sterben an der Vogelgrippe. Auf: welt.de vom 24. Oktober 2014
- Nordsee: Vogelgrippe-Variante für Seehundsterben verantwortlich. Auf: spiegel-online.de vom 24. Oktober 2014
- Avian influenza A (H10N8) who.int vom 30. Januar 2014.
- Sebastien G. Vachieri et al.: Receptor binding by H10 influenza viruses. In: Nature. Band 511, Nr. 7510, 2014, S. 475–477, doi:10.1038/nature13443
- CDC issues alert after H10N8 bird flu found in China. In: Taipei Times vom 27. Januar 2014.
- Peirong Jiao et al.: Complete Genome Sequence of an H10N8 Avian Influenza Virus Isolated from a Live Bird Market in Southern China. In: Journal of Virology. Band 86, Nr. 14, 2012, S. 7716, doi:10.1128/JVI.00959-12
- HaiYing Chen et al.: Clinical and epidemiological characteristics of a fatal case of avian influenza A H10N8 virus infection: a descriptive study. In: The Lancet. Band 383, Nr. 9918, 2014, S. 714–721, doi:10.1016/S0140-6736(14)60111-2
Chinese scientists report first human death associated with new bird flu virus. Auf: eurekalert.org vom 4. Februar 2014 - Sebastien G. Vachieri et al.: Receptor binding by H10 influenza viruses. In: Nature. Band 511, 2014, S. 475–477, doi:10.1038/nature13443
- Heng Zhang et al.: A Human-Infecting H10N8 Influenza Virus Retains a Strong Preference for Avian-type Receptors. In: Cell Host & Microbe. Band 17, Nr. 3, 2015, S. 377–384, doi:/10.1016/j.chom.2015.02.006
- Aeron C. Hurt et al.: Detection of Evolutionarily Distinct Avian Influenza A Viruses in Antarctica. In: mBio. Band 5, Nr. 3, 2014, e01098-14, doi:10.1128/mBio.01098-14
- James S. Gill et al.: Avian Influenza among Waterfowl Hunters and Wildlife Professionals. In: Emerging Infectious Deseases. Band 12, Nr. 8, 2006, doi:10.3201/eid1208.060492
- Ye Zhang et al.: Detection of reassortant avian influenza A (H11N9) virus in environmental samples from live poultry markets in China. In: Infectious Diseases of Poverty. Online-Publikation, 2016, 5:59, doi:10.1186/s40249-016-0149-2
- A/H12N1 in der Influenza Research Database
- Manoosak Wongphatcharachai et al.: Genetic characterization of influenza A virus subtype H12N1 isolated from a watercock and lesser whistling ducks in Thailand. In: Archives of Virology. Band 57, Nr. 6, 2012, S. 1123–1130, doi: 10.1007/s00705-012-1260-8
- Virginia S. Hinshaw et al.: Characterization of two influenza A viruses from a pilot whale. In: Journal of Virology. Band 58, Nr. 2, 1986, S. 655–656, Volltext (PDF)
- Virginia S. Hinshaw et al.: Antigenic and genetic characterization of a novel hemagglutinin subtype of influenza A viruses from gulls. In: Journal of Virology. Band 42, Nr. 3, 1982, S. 865–872, Volltext (PDF)
- V. Sivanandan et al.: Isolation of H13N2 Influenza A Virus from Turkeys and Surface Water. In: Avian Diseases. Band 35, Nr. 4, 1991, S. 974–977, doi:10.2307/1591638
- Ariel J. Pereda et al.: Avian Influenza Virus Isolated in Wild Waterfowl in Argentina: Evidence of a potentially unique phylogenetic lineage in South America. In: Virology. Band 378, Nr. 2, 2008, S. 363–370, doi:10.1016/j.virol.2008.06.010
- Jacqueline Nolting et al.: Recovery of H14 influenza A virus isolates from sea ducks in the Western Hemisphere. In: PLoS Currents. Band 4: RRN1290, 2012, doi: 10.1371/currents.RRN1290
- Andrew M. Ramey et al.: Genomic Characterization of H14 Subtype Influenza A Viruses in New World Waterfowl and Experimental Infectivity in Mallards (Anas platyrhynchos). In: PLoS ONE. 9(5): e95620, 2012, doi:10.1371/journal.pone.0095620
- Carolin Röhm et al.: Characterization of a Novel Influenza Hemagglutinin, H15: Criteria for Determination of Influenza A Subtypes. In: Virology. Band 217, Nr. 2, 1996, S. 508–516, doi:10.1006/viro.1996.0145
- Mariya V. Sivay et al.: Influenza A (H15N4) Virus Isolation in Western Siberia, Russia. In: Journal of Virology. Band 87, Nr. 6, 2013, S. 3578–3582, doi:10.1128/JVI.02521-12
- Ron A. Fouchier et al.: Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. In: Journal of Virology. Band 79, Nr. 5, 2005, S. 2814–2822, doi:10.1128/JVI.79.5.2814-2822.2005.
- Yulei Li, Minghui Li, Jingman Tian et al.: Characteristics of the first H16N3 subtype influenza A viruses isolated in western China. In: Transboundary and Emerging Diseases. Online-Vorabveröffentlichung vom 7. April 2020, doi:10.1111/tbed.13511.
- Suxiang Tong et al.: A distinct lineage of influenza A virus from bats. In: PNAS. Band 109, Nr. 11, 2014, S. 4269–4274, doi:10.1073/pnas.1116200109
- Mindaugas Juozapaitis et al.: An infectious bat-derived chimeric influenza virus harbouring the entry machinery of an influenza A virus. In: Nature Communications. Band 5, 2014, Artikelnummer: 4448, doi:10.1038/ncomms5448
- Bin Zhou et al.: Characterization of Uncultivable Bat Influenza Virus Using a Replicative Synthetic Virus. In: PLoS Pathogens. Band 10, Nr. 10, 2014: e1004420, doi:10.1371/journal.ppat.1004420
- Xueyong Zhu et al.: Hemagglutinin homologue from H17N10 bat influenza virus exhibits divergent receptor-binding and pH-dependent fusion activities. In: PNAS. Band 110, Nr. 4, 2013, S. 1458–1463, doi:10.1073/pnas.1218509110
- Umut Karakus, Thiprampai Thamamongood et al.: MHC class II proteins mediate cross-species entry of bat influenza viruses. In: Nature. Online-Vorabveröffentlichung vom 20. Februar 2019, doi:10.1038/s41586-019-0955-3
Neue Eintrittspforte für Influenza-Viren entdeckt. Auf: idw-online.de vom 20. Februar 2019 - Arinjay Banerjee et al.: Bat Influenza Viruses: Making a Double Agent of MHC Class II, in: Cell Press TIMI 1823, S. 1–3
- Suxiang Tong et al.: New World Bats Harbor Diverse Influenza A Viruses. In: PLoS Pathogens. Band 9, Nr. 10, 2013, e1003657, doi:10.1371/journal.ppat.1003657
- Campos ACA et al.: Bat Influenza A(HL18NL11) Virus in Fruit Bats, Brazil. In: Emerging Infectious Diseases. Band 25, Nr. 2, 2019, doi:10.3201/eid2502.181246
- Umut Karakus, Thiprampai Thamamongood, Kevin Ciminski, Wei Ran, Sira C. Günther: MHC class II proteins mediate cross-species entry of bat influenza viruses. In: Nature. Band 567, Nr. 7746, März 2019, ISSN 1476-4687, S. 109–112, doi:10.1038/s41586-019-0955-3 (nature.com [abgerufen am 24. Oktober 2019]).
- Kevin Ciminski et al.: Bat influenza viruses transmit among bats but are poorly adapted to non-bat species. In: Nature Microbiology. 2019, doi:10.1038/s41564-019-0556-9.
{kind=link}