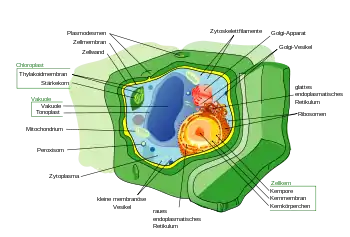
Zellwand
Eine Zellwand ist eine aus Polymeren aufgebaute Hülle, die die Zellen von Pflanzen, Bakterien, Pilzen, Algen und manchen Archaeen umgibt. Tiere und Protozoen haben keine Zellwände. Die Zellwand liegt außerhalb der Zellmembran, die ihrerseits das Zellinnere enthält. Sie wird als Abscheidungsprodukt lebender Zellen gebildet. Die Zellwand bietet Struktur und Schutz und wirkt zudem als Filter. Eine Hauptfunktion der Zellwand ist es, als Druckbehälter zu wirken; sie verhindert ein Platzen der Zelle, wenn aufgrund des osmotischen Gradienten Wasser eindringen sollte.
| Übergeordnet |
| Zellperipherie |
| Untergeordnet |
| Zellwand bei Bakterien/Pilzen/Pflanzen/Sporen Mikrofibrille Lignin/Suberin/Hemicellulose-Netzwerk Endosporenhülle Exosporium |
| Gene Ontology |
|---|
| QuickGO |
Eigenschaften
Die Zellwände verschiedener Organismen dienen ähnlichen Zwecken. Die Wand gibt Zellen Starrheit und Festigkeit, was Schutz gegen mechanische Beanspruchung bietet. Vielzelligen Organismen ermöglicht sie es, eine Form aufzubauen und zu erhalten (Morphogenese). Die Zellwand begrenzt auch den Eintritt großer, potentiell toxischer Moleküle. Sie erlaubt des Weiteren, ein osmotisch stabiles Milieu zu erzeugen, da übermäßige Wasseraufnahme aus der Umgebung verhindert wird, was ein Platzen der Zelle zur Folge hätte, und die Zellwand Wasser speichern kann. Zusammensetzung, Eigenschaften und Form der Zellwand können sich während des Zellzyklus ändern und sind abhängig von Wachstumsbedingungen.
Festigkeit der Zellwände
Die Festigkeit der Zellwände wird oft überschätzt. In den meisten Zellen ist die Zellwand flexibel, sie wird sich eher verbiegen, als eine bestimmte Form zu halten, besitzt dafür aber eine erhebliche Zugfestigkeit. Die Stabilität entsteht aus einem Zusammenspiel zwischen Turgor und Zugfestigkeit der Zellwand. Sobald der Turgor durch Wassermangel (welken) nachlässt, beginnen Blätter und Stängel in unverholzten (nicht lignifizierten) Pflanzen zu hängen. Nach John Howland ist ein guter Vergleich für die Zellwand ein Weidenkorb, in dem ein Ballon aufgeblasen ist (die Plasmamembran), der von innen Druck ausübt (den Turgor). Ein solcher Korb ist sehr fest und widerstandsfähig gegenüber mechanischen Beschädigungen.[1]
Pflanzliche Zellwände
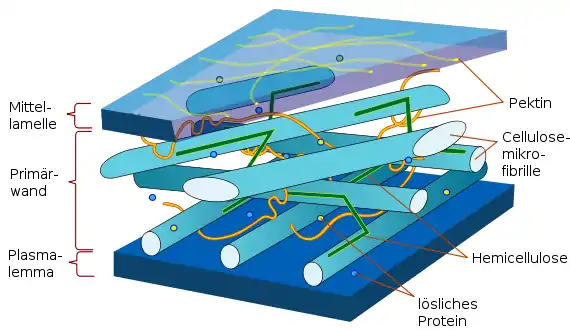
Pflanzliche Zellwände haben zwei Hauptfunktionen: Sie dienen einerseits der Zelle als formgebendes Element und sorgen für Stabilität. Außerdem hält die Zellwand dem Turgordruck stand, der den Protoplasten mit circa 0,5–1 MPa gegen die Zellwand drückt.[2] Sie umschließen die Zelle komplett und schützen sie. Pflanzliche Zellwände bestehen aus Zellulosefibrillen, die in eine Matrix aus Pektinen, Hemizellulosen, Proteinen und zum Teil auch Lignin eingebunden sind. Durch die Zellwände hindurch sind die einzelnen Zellen über Plasmodesmen verbunden.
Die Gesamtheit aller Zellwände und der Zellzwischenräume wird Apoplast genannt und entsteht durch Abgabe von Stoffen aus dem lebenden Teil der Zelle.
Aufbau der pflanzlichen Zellwand
von außen nach innen:
- Mittellamelle
- Primärwand
- Sekundärwand
- Tertiärwand
Mittellamelle
Die Mittellamelle besteht größtenteils aus Pektinen, die in die noch flüssige Zellplatte im Bereich des Phragmoplasten eingelagert werden. Sie hat Gel-Charakter und ist von geringer Ausdehnung.
Primärwand
Die Primärwand besteht aus Pektinen, Zellulose, Hemizellulose und Proteinen (vor allem aus Glykoproteinen wie Extensine). Nach der Zellteilung wird Zellulose in Form von Mikrofibrillen auf die Mittellamelle aufgelagert und es bildet sich eine Primärwand. Die Fibrillen bilden dabei keine Struktur. Daher ist die Primärwand elastisch, wodurch die Pflanzenzelle nicht in ihrem Wachstum eingeschränkt ist. Im Kollenchymgewebe kommt es zur teilweisen Verdickung der Primärwand. Die Zelle bleibt jedoch nach wie vor lebensfähig.
Cellulose ist in der primären (und noch wachsenden) Zellwand mit einem Anteil von 8–14 % vertreten. Sie liegt in Form von Mikrofibrillen vor, die wirr durcheinander angeordnet sind (als Streuungstextur). Diese Fibrillen sind in eine Matrix eingelagert, die hauptsächlich aus Hemicellulose und Pektin besteht. Die häufigste Hemizellulose in der primären Zellwand ist Xyloglucan. Die Hemicellulosestränge sind mit jeweils mehreren Cellulosefibrillen verbunden, sowie die Cellulosefibrillen untereinander, beide jeweils über Wasserstoffbrückenbindungen. Dadurch ergibt sich ein Netzwerk. In dieses sind die restlichen Komponenten als Unterstruktur eingebunden: über Ca2+- und Mg2+-Ionen vernetzte Pektine und über Isodityrosin-Brücken vernetzte Extensine. Die gelartigen Matrixpolimere und die darin eingelagerten, gerüstbildenden Cellulosefibrillen führen zu einem sehr reißfesten und trotzdem plastisch verformbaren Verbundmaterial. Auch durch die Reversibilität der vernetzenden Wasserstoffbrückenbindungen ist ein Umbau des Netzes möglich.[3]
Der Aufbau der Zellwände der Gräser (Poales) weicht massiv von dem hier beschriebenen ab, Xyloglucan und Pektin kommen seltener vor und sind teilweise durch Glucuronarabinoxylan, eine Hemizellulose, ersetzt. Sie stehen damit als Typ-II-Zellwand im Gegensatz zur Typ-I-Zellwand, die bei typischen Dikotyledonen vorkommen, sowie den meisten Monokotylen und Gymnospermen.[4]
Während des Wachstums erweitert sich die primäre Zellwand nach einer Ansäuerung durch Auxin durch eine Turgor-angetriebene Bewegung der festen Cellulosemikrofibrillen innerhalb der schwächeren Hemizellulose/Pektin-Matrix, katalysiert durch Expansine. Der stabile Endzustand der primären Zellwand wird als Sakkoderm bezeichnet.
Sekundärwand
Die Sekundärwand wird erst gebildet, wenn die Zelle ihr Wachstum beendet hat. Sie besteht größtenteils aus dichtgepackten Zellulosemikrofibrillen und Hemizellulosen, die in Mineralsubstanzen und vor allem in Lignine eingepackt sind. Dabei werden die Mikrofibrillen parallel zueinander aufgelagert. Mehrere Schichten überkreuzen sich dabei. In der Sekundärwand gibt es Aussparungen (Tüpfel), die die Verbindung zwischen einzelnen Zellen erlauben.
Kommt es zur Verholzung der Sekundärwand, so stirbt die Zelle ab (Bildung von Sklerenchym). Sekundäre Zellwände sind typisch für das Xylem. Es gibt auch abdichtende Sekundärwandschichten, z. B. bei Epidermiszellen die Cuticula, die ein Austrocknen der Pflanze verhindern. Sie enthalten Cutin, Wachse, oder im Falle von Kork auch Suberin.
Sekundäre Zellwände enthalten eine breite Palette zusätzlicher Verbindungen, die ihre mechanischen Eigenschaften und Durchlässigkeit verändern. So sind die Wände der Korkzellen in der Rinde der Bäume mit Suberin imprägniert, ebenso ist der äußere Teil der primären Zellwand der Pflanzenepidermis in der Regel mit Cutin und Wachs imprägniert und bildet eine Permeabilitätsbarriere. Suberin bildet auch die Permeabilitätsbarriere in primären Wurzeln, bekannt als Casparischer Streifen. Sekundäre Wände – vor allem in Gräsern – können auch mikroskopische Siliciumdioxid-Kristalle enthalten, die die Wand stärken und vor Pflanzenfressern schützen.
Die Zellwände einiger Pflanzengewebe funktionieren auch als Lager für Kohlenhydrate, die wieder monomerisiert und aufgenommen werden können, um Stoffwechsel und Wachstum zu unterstützen. Beispielsweise sind die Zellwände des Endosperms in den Samen von Getreide und Gräsern reich an Glucanen und anderen Polysacchariden, die leicht durch Enzyme während der Keimung der Samen zu einfachen Zuckern abgebaut werden können, um den wachsenden Embryo zu ernähren. Cellulose-Mikrofibrillen können hingegen nicht ohne weiteres von den Pflanzen verdaut werden.
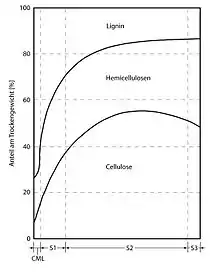
Die wichtigsten Polymere, aus denen Holz besteht (überwiegend sekundäre Zellwand), sind:
- Cellulose, 35–50 %
- Xylan, 20–35 %, eine Hemizellulose
- Lignin, 10–25 %, ein komplexes Phenolpolymer, das Zwischenräume in der Zellwand zwischen Cellulose, Hemizellulose und Pektin ausfüllt und sie hydrophober und fester macht
Tertiärwand
Die innerste Schicht wird auch „tertiäre Zellwand“ genannt. Sie ist dünn aber besonders resistent und deckt die Zellwand nach innen ab; sie hat eine warzige Oberfläche und ist reich an Pektinen und Hemizellulosen. Sie unterscheidet sich von der Sekundärwand in Zusammensetzung und Textur.[3][4]
Entstehung der pflanzlichen Zellwand
Nur während einer Zellteilung werden neue Zellwände gebildet. Dabei entsteht zuerst in der Äquatorialebene der Phragmoplast und aus diesem die Zellwandplatte. Diese entsteht durch das Zusammenfließen vieler Golgi-Vesikel und bleibt nach der Fertigstellung der Zellwand als Mittellamelle erhalten. Nun werden von beiden Seiten Mikrofibrillen in einer Streuungstextur regellos aufgelagert und bilden so die Primärwände. Die einzelnen Fibrillen sind über Wasserstoffbrücken miteinander verbunden. Da die Zelle noch wächst, kommt es zum Flächenwachstum der Zellwand. Die Dehnungsfähigkeit der Zellwand ist in der Streustruktur der Fibrillen begründet. Durch die Dehnung kommt es zur Wandverdünnung, was mit der Auftragung weiterer Fibrillen ausgeglichen wird. Das Wachstum der Primärwand endet mit der maximalen Ausdehnung der Zelle. Nach dem Flächenwachstum der Zellwand setzt nun das Dickenwachstum ein. Es werden Mikrofibrillen parallel und schichtweise aufgetragen, wobei die Fibrillen anliegender Schichten sich meist kreuzen (Paralleltextur). Die so entstehende Sekundärwand macht den Großteil der Zellwand aus und gibt ihr Stabilität. Sie ist jedoch nicht, wie die Primärwand, dehnungsfähig. Gegen Ende des Wachstums der Zellwand wird eine letzte Schicht, die Abschlusslamelle oder Tertiärwand, aufgetragen. Diese besteht aus Hemicellulose und Protopektin.
Chemische Zusammensetzung
Am stärksten am Aufbau der pflanzlichen Zellwand beteiligt sind die Kohlenhydrate Cellulose, Hemicellulose und Pektin. Deren Anteile unterscheiden sich zum Teil stark. Diese drei Stoffe sind Fraktionen, sie umfassen also zahlreiche verschiedene Polysaccharide. Im Wesentlichen bestehen sie nur aus sieben verschiedenen, glycosidisch verknüpften Monosacchariden: D-Glucose D-Galactose, D-Galacturonsäure, L-Rhamnose, L-Fucose, D-Xylose, L-Arabinose. Weitere Stoffe wie Lignin und Suberin können hinzu kommen, einen geringen Teil machen außerdem Polypeptide aus (5–10 %).
Die Zellwand besteht also hauptsächlich aus den folgenden Stoffgruppen:
Pektin
Pektine lassen sich mit relativ milden Medien aus der Zellwand lösen, z. B. mit heißem Wasser und Komplexbildnern oder mit Kaliumchlorat und Salpetersäure (Schulzesches Gemisch) oder nach einer Einwirkung spezifischer Enzyme (Pektinasen). Die cellulosehaltigen Zellwände widerstehen dieser Behandlung, nicht jedoch die pektinhaltige Mittellamelle, sodass sich die Zellen auch voneinander trennen (Mazeration). Chemisch gesehen handelt es sich bei Pektin um eine heterogene Fraktion, nach einer Extraktion kann man unterscheiden zwischen: Homogalacturonanen (1,4-α-D-Galacturonane, Polygalacturonsäure), Rhamnogalacturonanen (verzweigte Mischpolymere aus Galacturonsäure und Rhamnose mit verschiedenen zusätzlichen Zuckerresten), Arabinanen (1,5-α-L-Arabinosylketten) und Galactanen (1,4-β-D-Galactosylketten). Die Carboxygruppen des Pektins sind durch Ca2+ und Mg2+ über Salzbrücken miteinander verknüpft. Einige Carboxygruppen liegen jedoch als Ester mit Methanol vor und können somit keine Salzbrücken mehr bilden. Diese Salzbrücken sind als Bindung relativ reversibel, sodass Pektin elastisch und leicht veränderlich ist. Es wird zwischen Protopektin unterschieden, welches den größten Teil des Mittellamelle ausmacht und hauptsächlich aus Galacturonsäure und Rhamnose besteht sowie Pektin, welches aus hochmethyliertem Galacturonan besteht und in den Zellwänden vieler Früchte in größeren Mengen vorkommt.[3][4]
Hemicellulose
Hemicellulosen lassen sich aus der Zellwand durch eine alkalische Behandlung lösen. Ihr Name kommt daher, dass man sie früher fälschlicherweise für ein Zwischenprodukt der Cellulose-Synthese gehalten hat. Hemicellulose macht den Hauptteil der Matrix in der Primärwand aus. Bei Hemicellulosen handelt es sich ebenso wie beim Pektin um ein heterogenes Gemisch verschiedener Polysaccharide, dessen Zusammensetzung stark variieren kann, Hauptbestandteil sind meist Xyloglucane, es kommen aber auch Arabinogalactane und Glucomannane vor. Zusammengesetzt sind sie aus Pentosen wie D-Xylose und L-Arabinose und Hexosen wie D-Glucose, D-Mannose und D-Galactose.[3][4]
Cellulose
Cellulose ist ein lineares, unverzweigtes Polymer aus β-D-Glucose-Monomeren. Der Cellulose-Anteil beträgt in Primärwänden ca. 10 % und kann in Sekundärwänden über 90 % betragen. Cellulosemoleküle liegen in parallelen Bündeln vor, den Elementarfibrillen (bzw. Micellarsträngen), die zu Mikrofibrillen geordnet sind, die wiederum zu Makrofibrillen gebündelt sein können. Durch Wasserstoffbrückenbindungen werden die Fibrillen fest zusammen gehalten und können einen hohen Anteil parakristalliner Regionen besitzen. Durch die Parakristallinität gewinnen die Fibrillen eine Reißfestigkeit, die mit der von Stahl vergleichbar ist. Ebenfalls durch die Parakristallinität sind sie, anders als Matrixmaterial, kaum hydratisiert. In der Primärwand sind die Cellulose-Fibrillen zufällig in der Ebene angeordnet, dies nennt sich Streutextur. In der Sekundärwand hingegen werden die Fibrillen schichtweise aufgetragen, parallel und zur jeweils nächsten Schicht in der Ausrichtung ein wenig gedreht. Dies nennt sich Paralleltextur.[5][4]
Cellulose ist wasserunlöslich und kann somit nicht im Golgi-Apparat synthetisiert werden und durch Vesikel in die Zellwand transportiert werden, wie Pektine, Hemicellulosen und Zellwandproteine. Stattdessen wird Cellulose durch den Enzymkomplex Cellulose-Synthase direkt als Elementarfibrille in die Zellwand synthetisiert. Cellulose-Synthase ist ein Transmembranprotein, das als hexamerer Rosettenkomplex auftritt, meist in Rosettenfeldern. Die für die Synthese nötige Glucose wird in Form von Uridindiphosphoglucose (UDPG) im Cytoplasma geliefert. Die Ausrichtung der Cellulose-Fibrillen, wichtig für Zellwachstum und -differenzierung, erfolgt durch das Cytoskelett, an dem sich die Cellulose-Synthase wie auf Schienen bewegt. Pflanzen selbst können Cellulose nicht abbauen, Pflanzenfresser und manche Pilze jedoch besitzen das dafür nötige Enzym Cellulase. Technisch wird Cellulose durch Schwefelsäure wieder in Glucose aufgespalten (Holzverzuckerung).[3][4]
Zellwandproteine
Neben zahlreichen Enzymen, wie z. B. Hydrolasen, Esterasen, Peroxidasen und Transglycosylasen, die am Auf- und Umbau der Zellwand beteiligt sind (besonders in Primärwänden), machen Strukturproteine (1–5 %) den Hauptteil der Zellwandproteine aus. Es wird unterschieden zwischen glycinreichen Proteinen (GRP), prolinreichen Proteinen (PRP), Arabinogalactan-Proteinen (AGP) und hydroxyprolinreichen Glykoproteinen (HRGP). Die hydroxyprolinreichen Glykoproteine (HRGP) sind wohl am weitesten verbreitet und am besten untersucht. Jede Klasse von Glykoproteinen ist durch eine charakteristische, stark repetitive Proteinsequenz definiert. Die meisten sind glykosyliert, enthalten Hydroxyprolin (Hyp) und werden in der Zellwand vernetzt. [5] Der relative Anteil von Kohlenhydraten, sekundären Verbindungen und Proteinen variiert zwischen verschiedenen Pflanzen, Zelltyp und Alter.[6][7]
Durchlässigkeit
Die primäre Zellwand der meisten Pflanzenzellen ist semipermeabel und erlaubt den Durchtritt von kleinen Molekülen und Proteinen; durch Gel-Permeations-Chromatographie wurde die maximale Größe auf 30–60 kDa geschätzt. Vor allem Wasser und Kohlendioxid werden in der gesamten Pflanze von Zellwand zu Zellwand durch apoplastischen Transport verteilt. Der pH-Wert ist ein wichtiger Faktor für den Transport von Molekülen durch Zellwände.[8]
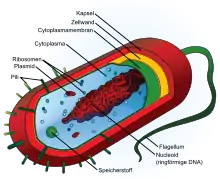
Bakterielle Zellwände
Die bakterielle Zellwand trennt die eigentliche Zelle mit ihrer umgebenden Zellmembran von der Umgebung. Die Zellwand ist einerseits robust genug, um die Zellgeometrie aufrechtzuerhalten, um so als Schutz vor widrigen Umweltbedingungen zu dienen. Andererseits ist sie aber auch flexibel genug, um Zellwachstum, Zellteilung und Transportvorgänge in die Zelle hinein und aus der Zelle heraus nicht zu behindern.
Durch die hohe Konzentration löslicher Stoffe im Cytoplasma entsteht in der Zelle ein osmotischer Druck von bis zu 1,5 MPa, welcher von der Zellwand kompensiert werden muss.
Außerdem dient die Zellwand dem Schutz vor Phagen und bei pathogenen Bakterien gegen das Immunsystem ihrer Wirte und muss aggressiven Metaboliten konkurrierender Mikroorganismen standhalten.
Bakterien können mit der sogenannten Gramfärbung grob nach ihrem Zellwandaufbau klassifiziert werden. Der Farbstoff Gentianaviolett ist bei grampositiven Bakterien aufgrund ihrer vielschichtigen Zellwand nicht auswaschbar, daher erscheinen diese Bakterien blau, wohingegen die gramnegativen Bakterien mit sehr dünner Zellwand aufgrund der Auswaschung des Farbstoffes gar nicht gefärbt sind. Diese können dann mit dem Farbstoff Fuchsin rötlich eingefärbt werden. Grampositive und gramnegative Bakterien unterscheiden sich im Aufbau ihrer Zellwände. Bei grampositiven besteht sie aus vielen Schichten des sogenannten Mureins (Peptidoglycan), in welches (Lipo)teichonsäuren und Proteine eingelagert sind.
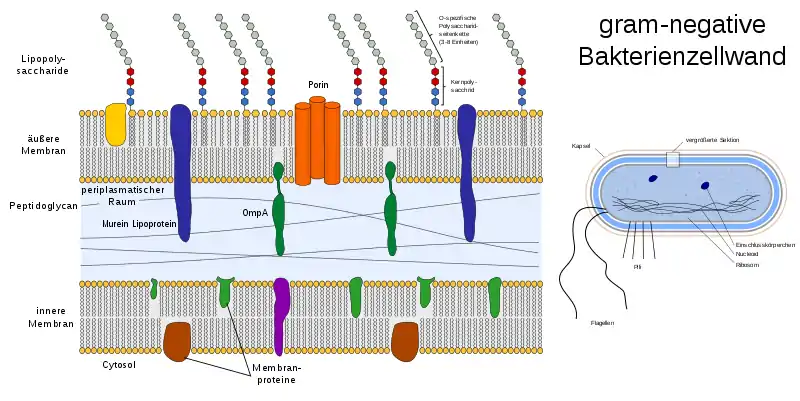
Bei gramnegativen Bakterien liegt der Zellmembran (innere Membran) nur eine dünne Peptidoglykanschicht auf, auf der eine zweite, äußere Zellmembran aufgelagert ist, die sich in Chemie und Aufbau von der inneren Zellmembran unterscheidet. Diese äußere Membran durchziehen Proteine, wie Porine, und sie weist außen Lipopolysaccharide (LPS) auf, weswegen sie auch als Lipopolysaccharidschicht bezeichnet wird. Das Lipid A der LPS kann als Endotoxin wirken und ist ein Virulenzfaktor pathogener Bakterien.
Je nach Spezies werden zusätzliche Proteinschichten (siehe S-Layer bei Archaea), Kapseln oder Schleimschichten ausgebildet.
Zellwände bei Pilzen
Die Zellwände, die bei manchen Pilzen die Zellen umgeben, bestehen aus Chitin, aus welchem auch das Exoskelett von Insekten aufgebaut ist. Ähnlich wie bei Pflanzen dienen die Zellwände auch hier der Versteifung, damit die Zellen ihre Form halten können.
Literatur
- Andreas Bresinsky, Christian Körner, Joachim W. Kadereit, Gunther Neuhaus, Uwe Sonnewald: Strasburger – Lehrbuch der Botanik. Begründet von E. Strasburger. Spektrum Akademischer Verlag, Heidelberg 2008 (36. Aufl.) ISBN 978-3827414557
- Biologie, Neil A. Campbell/ Jane B. Reece, Spektrum Akademischer Verlag
Weblinks
- www.biologie.uni-hamburg.de
- Max-Planck-Gesellschaft: Enzyme für Zellwandsynthese sind über Artgrenzen hinweg konserviert, 11. Juli 2011
Einzelnachweise
- John L. Howland: The Surprising Archaea: Discovering Another Domain of Life. Oxford University Press, Oxford 2000, ISBN 0-19-511183-4, S. 69–71.
- Strasburger, S. 91.
- Weiler, Elmar; Lutz Nover (2008): Allgemeine und molekulare Botanik, Thieme Verlag. S. 94–98.
- Peter Schopfer; Axel Brennicke (2006): Pflanzenphysiologie, Elsevier, München 2006, ISBN 978-3-8274-1561-5. S. 23–30.
- Weiler, Elmar; Lutz Nover (2008): Allgemeine und molekulare Botanik, Thieme Verlag. S. 240
- Peter Schopfer; Axel Brennicke (2006): Pflanzenphysiologie, Elsevier, München 2006, ISBN 978-3-8274-1561-5, S. 240
- Laurence Moire, Alain Schmutz, Antony Buchala, Bin Yan, Ruth E. Stark, and Ulrich Ryser: Glycerol Is a Suberin Monomer. New Experimental Evidence for an Old Hypothesis. In: Plant Physiol. 119, Nr. 3, 1999, S. 1137–1146. doi:10.1104/pp.119.3.1137. PMID 10069853. PMC 32096 (freier Volltext).
- C. Michael Hogan. 2010. Abiotic factor. Encyclopedia of Earth. eds Emily Monosson and C. Cleveland. National Council for Science and the Environment. Washington DC.