Tyrannosaurus
Tyrannosaurus (Latinisierung von altgriechisch τύραννος týrannos bzw. lat. tyrannus [„König, Despot“] sowie σαῦρος sauros [„Echse“])[2][3] ist eine Gattung bipeder, fleischfressender Dinosaurier aus der Gruppe der Theropoda.
| Tyrannosaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Das Tyrannosaurus-Exemplar „Stan“ im Manchester Museum | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (Oberes Maastrichtium)[1] | ||||||||||||
| 68 bis 66 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Tyrannosaurus | ||||||||||||
| Osborn, 1905 | ||||||||||||
| Arten | ||||||||||||
| ||||||||||||
Die einzige allgemein anerkannte Art ist Tyrannosaurus rex, häufig mit T. rex, umgangssprachlich auch T-rex, abgekürzt. Diese Art ist als bekanntester Dinosaurier ein fester Bestandteil der Populärkultur. Einige Paläontologen halten jedoch Tarbosaurus bataar aus Asien für eine zweite Art. Verschiedene andere Gattungen nordamerikanischer Tyrannosauriden wurden ebenfalls als Synonyme des Tyrannosaurus beschrieben. Eine 2022 veröffentlichte und in Fachkreisen kontrovers diskutierte Studie stellt aufgrund der hohen anatomischen Variabilität von Tyrannosaurus-Funden die zwei weiteren Arten Tyrannosaurus imperator und Tyrannosaurus regina auf.[4]
Fossilien des Tyrannosaurus fanden sich in verschiedenen Gesteinsformationen im westlichen Nordamerika, die auf die letzten drei Millionen Jahre der Kreidezeit datiert werden, also vor etwa 68 bis 66 Millionen Jahren (spätes Maastrichtium). Tyrannosaurus gehörte damit zu den letzten Nichtvogel-Dinosauriern, die bis zum Kreide-Tertiär-Massenaussterben existierten.
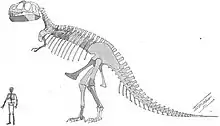
Wie andere Tyrannosauriden auch war Tyrannosaurus ein bipeder (zweibeiniger) Fleischfresser mit einem massigen Schädel, welcher durch einen langen, schweren Schwanz ausbalanciert wurde. Im Verhältnis zu den langen und kräftigen Hinterbeinen waren die Arme von Tyrannosaurus rex klein, aber für ihre Größe ungewöhnlich kräftig. Sie zeigten lediglich zwei Finger, obwohl über einen möglichen rudimentären (zurückgebildeten) dritten Finger berichtet wurde. Mit einer Länge von bis zu 13 Metern, einer Hüfthöhe von vier Metern und einem Gewicht von bis zu 9 Tonnen war er der größte bekannte Tyrannosauride und einer der größten landlebenden Fleischfresser überhaupt, obwohl einige andere Theropoden ähnlich groß oder noch größer (allerdings zumeist graziler und leichter) waren.[5][6][7] Als der bei weitem größte Fleischfresser seines Lebensraums stand Tyrannosaurus vielleicht als Spitzenprädator an der Spitze der Nahrungskette und jagte Hadrosaurier und Ceratopsier; allerdings vermuten einige Experten, dass es sich primär um einen Aasfresser handelte.
Bisher wurden mehr als 30 Exemplare von Tyrannosaurus identifiziert, darunter sind einige fast vollständige Skelette. Weiches Gewebe und Proteine wurden von mindestens einem dieser Funde beschrieben. Die vergleichsweise zahlreichen Funde erlaubten Studien über die Biologie dieses Tieres, einschließlich der Individualentwicklung und der Biomechanik. Zur Diskussion stehen des Weiteren die Fressgewohnheiten, die Physiologie und die mögliche Geschwindigkeit von Tyrannosaurus. Die Gesamtzahl aller jemals gelebten Exemplare wird auf 2,5 Milliarden geschätzt.[8]
Merkmale

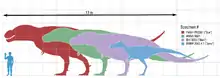
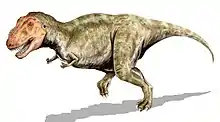
Tyrannosaurus gehörte zu den größten landlebenden Fleischfressern aller Zeiten. Das größte und schwerste bekannte Exemplar ist der kanadische Fund „Scotty“ (Katalognummer RSM P2523.8), der Ausmaße von gut 13 Metern und ein Gewicht von knapp 9 Tonnen erreichte.[9] Damit übertrifft der Fund das bisher größte nahezu vollständige Exemplar „Sue“ (Katalognummer FMNH PR2081), welches in der Länge zwischen 12,29[10] und 12,4[11] Meter misst und eine Hüfthöhe von 4 Metern hat.[6] Gewichtsschätzungen verschiedener Wissenschaftler variieren beträchtlich und reichen von weniger als 4,5[12][13] bis über 7,2 Tonnen,[14] wobei die neuesten Schätzungen zwischen 5,4 und 6,8 Tonnen liegen.[7][15][16][17] Eine neue Methode hat jedoch das Volumen einiger Individuen gemessen und führte zu einem Minimum von 9,5 t für „Sue“.[10] Greg Paul schätzt „Sue“ auf 6,1 t.[18]
Tyrannosaurus rex war größer als der gut bekannte Allosaurus aus dem Oberjura und etwas kleiner als Spinosaurus aus der frühen Oberkreide.[19][20] Sein Gewicht übertrifft das aller anderen terrestrischen Theropoden – nur Spinosaurus könnte ein ähnliches Gewicht erreicht haben.
Der „S“-förmig gebogene Hals von Tyrannosaurus war kurz und muskulös, um den schweren Kopf zu unterstützen. Die Beine gehörten zu den im Verhältnis zum Körper längsten aller Theropoden und stehen im Kontrast zu den winzigen, aber kräftigen Armen. Lange wurde angenommen, dass die Arme lediglich zwei Finger besaßen – in einem noch nicht veröffentlichten Bericht wird allerdings ein dritter, rudimentärer Finger beschrieben.[21] Der Schwanz war schwer und lang, um den massiven Rumpf und Kopf auszubalancieren, und zeigte manchmal über vierzig Wirbel. Um die enorme Masse des Tieres zu kompensieren, waren viele Knochen hohl.[5]
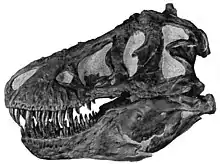
Die größten bekannten Tyrannosaurus-Schädel waren bis zu 1,5 Meter lang.[22] Im Gegensatz zu den Schädeln anderer nicht-tyrannosauroider Theropoden war der hintere Teil des Schädels extrem breit, während die Schnauze schmal war. Durch diese Anpassung waren die Augen mehr nach vorne gerichtet, was ein ungewöhnlich gutes räumliches Sehen ermöglichte.[23][24] Die Schädelknochen waren massiv und einige Knochen, wie das paarige Nasenbein, waren verschmolzen, was den Schädel stabilisierte. Viele Knochen waren jedoch pneumatisch, zeigten also Aushöhlungen, welche die Knochen flexibler aber auch leichter machten. Diese und andere Merkmale, die den Schädel verstärkten, gehörten zu einem Trend innerhalb der Tyrannosauroiden, der zu einem zunehmend kräftigen Biss führte, der andere Nicht-Tyrannosauriden bei weitem übertraf.[25][26][27] Die Oberkiefer waren von oben betrachtet „U“-förmig angeordnet, und nicht „V“-förmig, wie bei den meisten anderen nicht-tyrannosauroiden Theropoden. Dies vergrößerte zwar die Menge an Gewebe, die ein Tyrannosaurier mit einem Biss aus der Beute herausreißen konnte, steigerte allerdings auch die Belastung der vorderen Zähne.[28][29]
Die Zähne von Tyrannosaurus und anderen Tyrannosauroiden zeigten eine Heterodontie (Unterschiede in der Form).[5][30] So waren die Zähne des Zwischenkieferbeins am vorderen Ende des Oberkiefers eng gepackt, im Querschnitt „D“-förmig, hatten verstärkende Kämme auf der Rückseite, waren incisiform (die Spitzen waren meißelähnlich geformt) und nach hinten gekrümmt. Der „D“-förmige Querschnitt, die verstärkenden Kämme und die Krümmung nach hinten verminderten das Risiko, dass die Zähne während des Bisses umknickten. Die restlichen Zähne waren robust, bananenförmig und hatten mehr Abstand zueinander; auch sie hatten verstärkende Kämme.[31] Die Zähne des Oberkiefers waren größer als die Zähne des Unterkiefers, mit Ausnahme der Zähne am hinteren Ende des Unterkiefers. Der größte gefundene Zahn wird inklusive Zahnwurzel auf 30 cm Länge geschätzt; damit ist er der größte Zahn, der bisher von einem fleischfressenden Dinosaurier bekannt ist.[6]
Systematik
Tyrannosaurus ist die Typgattung der Überfamilie Tyrannosauroidea, der Familie Tyrannosauridae sowie der Unterfamilie Tyrannosaurinae – mit anderen Worten ist sie der Standard, an welchem Paläontologen entscheiden, ob andere Spezies in derselben Gruppe eingeordnet werden. Andere Mitglieder der Unterfamilie Tyrannosaurinae schließen den nordamerikanischen Daspletosaurus und den asiatischen Tarbosaurus mit ein, die beide in der Vergangenheit von manchen Forschern als Synonyme von Tyrannosaurus betrachtet wurden.[29][32][33] Früher wurde angenommen, dass die Tyrannosauriden die Nachfahren früherer Gruppen großer Theropoden waren, wie die Megalosaurier und die Carnosaurier. Heute werden sie innerhalb der Coelurosauria eingeordnet, deren Mitglieder sich meist durch eine geringe Körpergröße auszeichneten.[28]
Im Jahr 1955 benannte der sowjetische Paläontologe Evgeny Maleev eine neue Art, Tyrannosaurus bataar, die in der Mongolei entdeckt wurde.[34] Im Jahr 1965 wurde sie in Tarbosaurus bataar umbenannt und somit einer eigenen Gattung zugeschrieben.[35] Trotz dieser Umbenennung kamen viele phylogenetische Analysen zu dem Ergebnis, dass Tarbosaurus bataar das Schwestertaxon von Tyrannosaurus rex war,[33] weswegen er häufig als eine asiatische Tyrannosaurus-Art geführt wurde.[28][36][37] Eine Neubeschreibung des Schädels von Tarbosaurus bataar zeigte jedoch, dass dieser viel schmaler als der von Tyrannosaurus rex war und dass die Kräfte während eines Bisses ganz anders auf die Schädelknochen verteilt wurden. Die Kräfteverteilung im Schädel von Tarbosaurus bataar ähnelte vielmehr der von Alioramus, einem weiteren asiatischen Tyrannosaurier.[38] Eine kladistische Analyse kam zu dem Schluss, dass Alioramus, und nicht Tyrannosaurus, das Schwestertaxon von Tarbosaurus war – falls dies bestätigt werden kann, würde es bedeuten, dass Tarbosaurus und Tyrannosaurus separat geführt werden sollten.[32]
Andere Tyrannosauriden-Überreste, die in den gleichen geologischen Formationen wie Tyrannosaurus rex gefunden wurden, galten ursprünglich als separate Taxa – darunter sind Aublysodon und Albertosaurus megagracilis,[29] wobei letzterer im Jahr 1995 in Dinotyrannus megagracilis umbenannt wurde.[39] Heute werden diese Funde als juvenile Exemplare von Tyrannosaurus rex geführt.[40]
Eine 2022 veröffentlichte Studie des amerikanischen Paläontologen Gregory S. Paul und seinen Kollegen W. Scott Persons IV. und Jay Van Raalte[4] kommt aufgrund der hohen Variabilität des Skelettaufbaus von Tyrannosaurus-Fossilien (bezüglich des Verhältnisses von Länge und Umfang des Femurs, den generellen Körperproportionen sowie der Anzahl der Zähne in den Zahnfächern) zu dem Ergebnis, dass es zusätzlich zu Tyrannosaurus rex noch zwei weitere Arten gegeben hat. Bei den stratigraphisch ältesten Tyrannosaurus-Funden handelt es sich um besonders robust gebaute Exemplare mit kleinem Verhältnis von Femurlänge zum -umfang, deren Kiefer zwei kleine Eckzähne beinhalteten. Diese Exemplare benannten sie als Tyrannosaurus imperator („Kaiser“). Jüngere Funde konnten in zwei Morphotypen unterteilt werden: Tyrannosaurus rex („König“) ist durch einen robusten Körperbau mit moderatem Verhältnis von Femurlänge zum -umfang und einem Eckzahn gekennzeichnet. Der gleichzeitig mit Tyrannosaurus rex lebende Morphotyp mit grazilem Körperbau, einem hohen Verhältnis von Femurlänge zu dessen Umfang und einem Eckzahn wurde Tyrannosaurus regina („Königin“) genannt. Die Forscher schlossen Ontogenese und Geschlechtsdimorphismus als Erklärungen für die Variabilität aus. Die Variabilität der Exemplare, die bisher der einen Art Tyrannosaurus rex zugeordnet wurden, sei größer als bei anderen Gattungen von Theropoden, die als unterschiedliche Arten interpretiert wurden. Die Forscher interpretieren Tyrannosaurus imperator als eine Chronospezies, die sich später in die beiden Arten Tyrannosaurus rex und Tyrannosaurus regina aufspaltete.
Von anderen Paläontologen wurden die Ergebnisse von Paul und Kollegen gemischt aufgenommen. Während einige Forscher wie der amerikanische, auf Tyrannosaurier spezialisierte Paläontologe Thomas Holtz die Studie für insgesamt plausibel halten, jedoch mehr Untersuchungen fordern, die die Aufspaltung in drei Arten stützen, wird von anderen (beispielsweise der amerikanischen Paläontologin Jingmai O'Connor) insbesondere das Fehlen eindeutiger Unterschiede kritisiert. Die zur Unterscheidung herangezogenen Merkmale wie das Länge-Umfang-Verhältnis des Femurs sowie die Zeiträume, in denen die Arten lebten, überschneiden sich und einige Exemplare konnten keiner der drei Arten sicher zugeordnet werden. Laut dem englischen Paläontologen David Hone habe insbesondere die hohe Popularität von Tyrannosaurus zur Folge, dass Belege für gravierende Änderungen wie die Aufspaltung von einer in mehrere Arten einen höheren Standard erfüllen müssen, als es bei weniger bekannten und gut untersuchten Gattungen der Fall wäre.[41]
Nanotyrannus
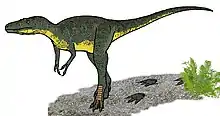
Einen in Montana entdeckten, nur 60 cm langen Schädel beschrieb Charles W. Gilmore 1946 ursprünglich als Gorgosaurus lancensis,[42] später wurde er jedoch einer neuen Gattung zugeordnet, Nanotyrannus.[43] Im Jahr 2001 entdeckte eine Forschergruppe des Burpee Museum of Natural History ein weiteres, zu 50 % erhaltenes Skelett, das auf den Spitznamen „Jane“ getauft wurde (Katalognummer BMRP 2002.4.1.).[44] Die Meinungen bezüglich der Gültigkeit von N. lancensis sind geteilt. Viele Paläontologen schreiben die Funde einem juvenilen T. rex zu[45]. Eine 2020 in Science Advances veröffentlichte Studie stützt diese Annahme: Eine detaillierte Analyse von Knochenquerschnitten juveniler Tyrannosaurus legt nahe, dass die Wachstumsraten von Tyrannosaurus mit zunehmendem Alter variierten und dass die Art ihr Wachstum scheinbar verlangsamen konnte, wenn die Nahrung knapp war, was ihnen möglicherweise einen evolutionären Vorteil verschafft haben könnte. Einige der so untersuchten Knochen von Nanotyrannus bestätigten, dass es sich tatsächlich um Jungtiere handelt. Nanotyrannus war also entweder größer als bisher angenommen, oder – was die Forscher für viel wahrscheinlicher halten – die Fossilien gehören zu jungen Tyrannosaurus.[46] Das Taxon Nanotyrannus ist demnach wahrscheinlich ungültig; inzwischen ist die Betrachtung von Nanotyrannus als juveniles Synonym von Tyrannosaurus weitgehend wissenschaftlicher Konsens.
Manospondylus

Der erste Fund, der Tyrannosaurus rex zugeschrieben werden kann, besteht aus zwei teilweisen Wirbeln (wovon einer verloren gegangen ist), die von Edward Drinker Cope im Jahr 1892 gefunden und als Manospondylus gigas beschrieben wurden. Osborn erkannte die Ähnlichkeit zwischen M. gigas und T. rex bereits 1917, konnte beide Gattungen jedoch nicht für identisch erklären, da die Manospondylus-Wirbel zu fragmentarisch waren.[47]
Im Juni 2000 lokalisierte das Black Hills Institute die Fundstelle von M. gigas in South Dakota und brachte weitere Tyrannosaurierknochen zutage. Diese wurden als weitere Überreste desselben Individuums beschrieben und als identisch mit Tyrannosaurus rex erkannt. Nach den Internationalen Regeln für die Zoologische Nomenklatur (ICZN), nach denen die Benennung von Tieren international geregelt wird, sollte Manospondylus gigas gegenüber Tyrannosaurus rex Priorität haben, weil dieser vor T. rex benannt wurde. Jedoch besagt die vierte Ausgabe der ICZN, die am 1. Januar 2000 in Kraft trat, dass der vorherrschende Name beibehalten werden muss, wenn das früher benannte Synonym seit 1899 nicht mehr als gültiger Name verwendet und das später benannte Synonym als gültiger Name in mindestens 25 Arbeiten geführt wurde, die von mindestens zehn Autoren in den vorhergegangenen 50 Jahren veröffentlicht wurden.[48] Tyrannosaurus rex erfüllt diese Bedingungen und würde wahrscheinlich, falls er jemals herausgefordert werden sollte, als Nomen protectum („geschützter Name“) geführt werden.[49]
Paläobiologie
Entwicklungs- und Populationsbiologie
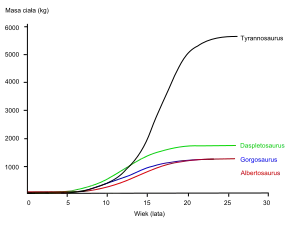
Durch die Entdeckung einiger juveniler Tyrannosaurus rex-Exemplare konnten Wissenschaftler Rückschlüsse auf die Individualentwicklung (Ontogenese), Lebenserwartung und Wachstumsraten ziehen. Das kleinste bekannte Individuum (LACM 28471, der „Jordan-Theropode“) wird auf ein Gewicht von nur 29,9 kg geschätzt, während die größten Exemplare, wie RSM P2523.8 ("Scotty"), wahrscheinlich über 8800 kg gewogen haben. Histologische Analysen kamen zu dem Ergebnis, dass LACM 28471 lediglich zwei Jahre alt war, als es starb, während „Scotty“ 30 Jahre alt wurde und damit vielleicht nahe am Maximalalter der Spezies lag.[9]
Weitere Exemplare wurden ebenfalls knochenhistologisch untersucht. So konnten Wachstumskurven entwickelt werden, welche das Lebensalter eines Individuums mit seiner Größe in Bezug setzen. Die Wachstumskurve von T. rex ist „S“-förmig gekrümmt und zeigt, dass Jungtiere ein Gewicht von 1800 kg nicht überschritten, bis sie etwa 14 Jahre alt waren, wo eine rasante Wachstumsphase einsetzte. Während dieser vier Jahre währenden Wachstumsphase hätte ein junger T. rex pro Jahr durchschnittlich 600 kg Gewicht zugelegt. Mit 18 Jahren flacht sich die Kurve wieder ab, was ein dramatisches Verlangsamen des Wachstums innerhalb von kurzer Zeit anzeigt. Beispielsweise liegen nur 600 kg zwischen dem 28 Jahre alten Exemplar „Sue“ und dem 22 Jahre alten kanadischen Exemplar (RTMP 81.12.1).[7] Eine weitere, 2004 veröffentlichte Studie unterstützt diese Ergebnisse und kam zu dem Ergebnis, dass sich das rapide Wachstum bereits mit 16 Jahren zu verlangsamen begann.[50] Diese plötzliche Verlangsamung des Wachstums könnte auf das Erreichen der Geschlechtsreife hindeuten. Diese Hypothese wird durch die Entdeckung von einer kalziumreichen Knochensubstanz im Oberschenkelknochen (Femur) eines 16 bis 20 Jahre alten T. rex aus Montana (MOR 1125, auch als „B-rex“ bekannt) unterstützt – diese Knochensubstanz ist ansonsten lediglich von weiblichen Vögeln bekannt, die vor der Eiablage stehen und das Kalzium zur Bildung der Eierschalen benötigten. Dies deutet darauf hin, dass „B-rex“ geschlechtsreif war.[51] Eine neue Studie gibt ein Alter von 18 Jahren für dieses Exemplar an.[52] Andere Tyrannosauriden zeigen sehr ähnliche Wachstumskurven, hatten aber niedrigere Wachstumsraten, weil sie nicht so groß wurden wie T. rex.[53]
Über die Hälfte der bekannten T. rex-Exemplare ist innerhalb von sechs Jahren nach dem Erreichen der Geschlechtsreife gestorben; ein Muster, das sich auch bei anderen Tyrannosauriern und bei einigen heutigen großen Vögeln und Säugetieren findet. Diese Spezies sind durch eine hohe Sterberate als „Babys“ gekennzeichnet, die von einer relativ geringen Sterberate als Jungtiere gefolgt wird. Nach der Geschlechtsreife steigt die Sterberate wieder an, zum Teil wegen der Belastungen, die die Fortpflanzung mit sich bringt. Eine Studie vermutet, dass die Seltenheit juveniler T. rex-Fossilien teilweise an einer geringen Sterberate von Jungtieren liegt; die Jungtiere sind nicht in großer Zahl gestorben und somit nur selten fossiliert. Diese Seltenheit kann allerdings auch andere Ursachen haben, wie eine unvollständige Fossilüberlieferung oder die Bevorzugung großer, spektakulärer Funde gegenüber kleinen Funden durch die Fossiliensammler.[53]
Ein Forschungsteam der University of California in Berkeley schätzte die Häufigkeit von Tyrannosaurus zu jeder Zeit auf 20.000 Individuen. Bei geschätzten 127.000 Generationen entspricht das einer Gesamtzahl von 2,5 Milliarden Tieren, die jemals existierten. Allerdings ist das nur eine Näherung und es kann mit hohen Abweichungen gerechnet werden.[8]
Geschlechtsdimorphismus

Anhand von Unterschieden im Bau der Skelette wurden zwei unterschiedliche Typen beschrieben, oder „Morphen“, ähnlich wie bei einigen anderen Theropoden. Der sogenannte „robuste Morph“ war schwerer gebaut als der andere, als „graziler Morph“ bezeichnete Typ. Für gewöhnlich werden diese morphologischen Unterschiede als Geschlechtsdimorphismus gedeutet, wobei die „robusten“ Individuen meistens als Weibchen interpretiert werden. Beispielsweise scheint das Becken einiger „robuster“ Skelette weiter zu sein, vielleicht um die Passage der Eier zu ermöglichen.[54] Es wurde auch angenommen, dass der robuste Morph mit einer Verkleinerung des Chevron-Knochens am ersten Schwanzwirbel in Verbindung steht, was ebenfalls die Passage der Eier ermöglicht haben sollte – ähnliches wurde fälschlicherweise bei Krokodilen beschrieben.[55]
In den letzten Jahren kamen Zweifel auf, ob es sich bei diesen Merkmalen tatsächlich um Geschlechtsdimorphismus handelte. Wie eine Studie aus dem Jahre 2005 zeigt, zeigen Krokodile keinen Geschlechtsdimorphismus an den Chevron-Knochen, obwohl dies früher vermutet wurde. Somit erscheint auch der vermutete Geschlechtsdimorphismus an den Chevron-Knochen des T. rex fragwürdig.[56] Der gut entwickelte Chevron-Knochen des ersten Schwanzwirbels von „Sue“, einem sehr robusten Individuum, zeigt letztendlich, dass dieses Merkmal nicht zur Unterscheidung der beiden Morphen herangezogen werden kann. Da Skelette von T. rex von Saskatchewan (Kanada) bis New Mexico gefunden wurden, könnten diese Unterschiede zwischen Individuen auch geografische Variationen sein, und kein Geschlechtsdimorphismus. Auch könnten die Unterschiede mit dem Alter zusammen hängen, wobei robuste Individuen ältere Tiere dargestellt haben könnten.[5]
Lediglich ein einziges T. rex-Skelett konnte schlüssig einem Geschlecht zugeordnet werden. In einigen Knochen von „B-rex“ ist weiches Gewebe erhalten geblieben, wobei auch kalziumreiches Gewebe entdeckt wurde, das ansonsten nur bei heutigen weiblichen Vögeln während der Eiablage wächst und für die Bildung der Eierschalen benötigt wird. Dieses Gewebe fehlt bei männlichen Vögeln, obwohl sie es produzieren können, wenn ihnen Sexualhormone wie Estrogen verabreicht werden. Dies lässt den Schluss zu, dass es sich bei „B-rex“ um ein Weibchen handelt.[51] Weitere Forschungen haben ergeben, dass dieses kalziumreiche Gewebe bei den Krokodilen fehlt, welche als die – zusammen mit den Vögeln – nächsten lebenden Verwandten der Dinosaurier gelten. Dies ist ein weiterer Hinweis auf die enge Verwandtschaft zwischen Vögeln und Dinosauriern.[57]
Körperhaltung

_2012-05-11_04.jpg.webp)
Wie viele zweibeinig laufende (bipede) Dinosaurier wurde auch Tyrannosaurus rex in der Vergangenheit oft in aufrechter Haltung dargestellt, wobei der Schwanz wie ein drittes Bein auf dem Boden auflag und die Körperachse von der vertikalen Lage aus um 45 Grad oder weniger geneigt war, ähnlich wie bei einem Känguru. Dieses Konzept rührt von Joseph Leidys Rekonstruktion eines Hadrosaurus aus dem Jahr 1865 her, der ersten Rekonstruktion eines Dinosauriers in bipeder Haltung.[58] Henry Fairfield Osborn, der ehemalige Präsident des American Museum of Natural History (AMNH) in New York, enthüllte 1915 eine Montage des ersten vollständigen T. rex-Skeletts in aufrechter Haltung, was die Hypothese einer aufrechten Haltung unterstützte. Das Skelett stand in dieser aufrechten Position für beinahe ein Jahrhundert, bis es im Jahr 1992 demontiert wurde.[59] Um das Jahr 1970 hatten Wissenschaftler bemerkt, dass diese Haltung inkorrekt war; so hätte eine aufrechte Haltung zur Verlagerung (Dislokation) verschiedener Gelenke geführt, wie den Hüften und der Verbindung zwischen Kopf und Wirbelsäule.[60] Trotz dieser Unstimmigkeiten inspirierte das Skelett im AMNH ähnliche Darstellungen in vielen Filmen und Gemälden (wie z. B. Rudolph Zallingers berühmtes Wandgemälde „The Age Of Reptiles“ im Peabody Museum of Natural History[61]) bis in die 1990er Jahre hinein, als Filme wie Jurassic Park eine korrektere Haltung populär machten. Moderne Darstellungen zeigen T. rex, wie er seinen Körper annähernd parallel zum Boden hielt, während der Schwanz den Kopf ausbalancierte.[29]
Arme
.jpg.webp)
Anfangs war von den Armen lediglich der Oberarmknochen bekannt,[62] weshalb Osborn für das erste montierte, ab 1915 ausgestellte Tyrannosaurus-Skelett längere, dreifingrige Arme modellierte, die denen von Allosaurus ähnelten.[47] Schon 1914 beschrieb Lawrence Lambe jedoch die kurzen, zweifingrigen Arme des nahe verwandten Gorgosaurus, was ähnliche Arme auch bei T. rex vermuten ließ.[63] Diese Annahme konnte erst 1989 bestätigt werden, als mit MOR 555 (dem „Wankel rex“) vollständige Arme gefunden wurden. Auch das Skelett von „Sue“ zeigt vollständige Arme.[5] Die Arme von T. rex waren mit einer Länge von nur einem Meter im Verhältnis zur Körpergröße sehr klein. Dennoch waren sie nicht zurückgebildet (rudimentär), sondern besaßen große Ansatzflächen für Muskeln, was zeigt, dass sie sehr kräftig waren. Dies bemerkte bereits Osborn im Jahr 1906, welcher spekulierte, die Arme könnten dem Festhalten des Partners während der Kopulation gedient haben.[64] Ein anderer Autor vermutet, die Arme könnten das Tier beim Erheben aus der Bauchlage heraus unterstützt haben.[60] Eine weitere Möglichkeit bestand im Festhalten von Beutetieren, damit diese von den enormen Kiefern des Tyrannosauriers erledigt werden konnten. Diese Hypothese könnte von biomechanischen Studien unterstützt werden: So zeigen die Armknochen eine extrem dicke Kortikalis (der harte Außenknochen), was zeigt, dass sie starken Belastungen widerstehen konnten. Der Biceps brachii eines ausgewachsenen Tyrannosaurus rex konnte alleine ca. 200 kg heben, im Zusammenspiel mit anderen Muskeln wäre er noch stärker. Die Schulter- und Ellenbogengelenke erlaubten jedoch nur Bewegungen von 40 bis 45 Grad – im Kontrast dazu konnte Deinonychus seine Arme um 88 bis 130 Grad bewegen, während sich ein menschlicher Arm an der Schulter um 360 Grad und am Ellenbogengelenk um 165 Grad bewegen kann. Der schwere Bau der Armknochen, die extreme Stärke der Muskeln und die eingeschränkten Bewegungsmöglichkeiten weisen nach Ansicht einiger Forscher auf ein System hin, das für das Festhalten von Beutetieren ausgelegt war.[65]
Weiches Gewebe
Im März 2005 machten Forscher um Mary Schweitzer bekannt, dass in der Markhöhle eines 68 Millionen Jahre alten Beinknochens von Tyrannosaurus weiches Gewebe entdeckt wurde. Der Knochen musste für einen Transport zerbrochen werden und wurde dann nicht der üblichen Prozedur zur Erhaltung unterzogen, weil Schweitzer hoffte, ihn auf weiches Gewebe testen zu können.[66] Der Knochen (MOR 1125) vom Museum of the Rockies wurde zuvor in der Hell-Creek-Formation ausgegraben. Die Forscher fanden flexible, sich gabelnde Blutgefäße und faseriges, aber elastisches Knochengewebe. In diesen Gefäßen und Geweben konnten außerdem Mikrostrukturen festgestellt werden, die Blutzellen ähneln, wobei die Strukturen Ähnlichkeiten mit den Blutzellen und Gefäßen von Straußen haben. Ob das Material durch einen unbekannten Prozess erhalten geblieben ist, der sich von der normalen Fossilisation unterscheidet, oder ob es sich um originales Material handelt, ist unbekannt.[67] Falls es sich um originales Material handelt, könnte mithilfe erhalten gebliebener Proteine indirekt auf einen Teil der DNA-Sequenz geschlossen werden, da jedes Protein von einem besonderen Gen hergestellt wird. Dass weiches Gewebe nicht schon früher in den Knochen von Dinosauriern entdeckt wurde, könnte daran liegen, dass Forscher bis heute überliefertes weiches Gewebe für ausgeschlossen hielten und somit nicht danach suchten. Seit diesem Fund wurden gewebeähnliche Strukturen bei zwei weiteren Tyrannosauriern und einem Hadrosaurier nachgewiesen.[66] Eine spätere Studie fand Spuren des Strukturproteins Kollagen in einem gereinigten Tyrannosaurus-Knochen.[68] Diese Untersuchungen an überliefertem Gewebe von Tyrannosauriern bestätigen, dass Vögel enger mit Tyrannosauriern als mit anderen modernen Tieren verwandt sind.[69]
In der 2007 erschienenen Studie von Chris Organ, Biologe an der Harvard University, und John Asara vom „Beth Israel Deaconess Medical Centre“ konnten sie mittels erhaltenen Kollagens in dem von Mary Schweitzer gefundenen Oberbeinknochen anhand von sechs Peptiden mit 89 Aminosäuren relativ sicher eine phylogenetische Beziehung zwischen Alligatoren und Hühnern und Straußen nachweisen. Demnach sind die erste Abspaltung der Hühner die nächsten noch lebenden Verwandten des T-Rex. Überprüft wurde diese Methode anhand der Verwandtschaft von Mastodon und Elefant.[70]
Thomas Kaye und Kollegen bezweifeln diese Ergebnisse in einer 2008 erschienenen Veröffentlichung und meinen, es handle sich bei den vermeintlichen Geweberesten um bakterielle Biofilme. Die Bakterien besiedelten einst die Hohlräume des Knochens, die zuvor von echten Blutgefäßen und Zellen eingenommen wurden.[71] Die von Schweitzer aufgrund ihrer Eisenhaltigkeit als Blutgefäße interpretierten Strukturen deuten die Forscher zudem als Framboide – rundliche, mikroskopisch kleine Mineralstrukturen. Die Forscher fanden derartige Strukturen in weiteren Fossilien, unter anderem in einem Ammoniten an einem Ort, wo das gefundene Eisen nichts mit Blut zu tun gehabt haben konnte.[72]
Haut und Federn
Im Jahre 2004 wurde der frühe Tyrannosauroide Dilong paradoxus aus der berühmten Yixian-Formation von China beschrieben. Wie bei vielen anderen Theropoden dieser Formation sind auf dem Skelett von Dilong faserartige Strukturen sichtbar, die für gewöhnlich als die Vorläufer von Federn interpretiert werden. Es gibt Vermutungen, dass auch Tyrannosaurus und andere Tyrannosauriden solche Protofedern gehabt haben könnten – seltene Hautabdrücke von erwachsenen Tyrannosauriden aus Kanada und der Mongolei zeigen jedoch Schuppen, wie sie für andere Dinosaurier typisch sind.[73] Zwar ist es möglich, dass Protofedern die Teile des Körpers bedeckten, die nicht durch Hautabdrücke überliefert sind. Das Fehlen einer isolierenden Körperbedeckung zeigt sich jedoch auch bei heutigen Großsäugern wie Elefanten, Flusspferden und Nashörnern. Große Körper können wesentlich besser Wärme speichern als kleine, da das Volumen mit steigender Größe schneller zunimmt als die Hautoberfläche. Somit stellt ein isolierendes Fell oder ein Federkleid für große Tiere in warmen Klimazonen keinen Vorteil mehr dar und kann gar zum Nachteil werden, da die Gefahr der Überhitzung besteht. Wahrscheinlich sind Protofedern während der Evolution großer Tyrannosauriden wie Tyrannosaurus verloren gegangen.[74]
Geschwindigkeit
Eine Studie niederländischer Wissenschaftler ermittelte anhand einer als Eigenfrequenzmethode bezeichneten Simulation der Beinmuskulatur und der Schwingung des Schwanzes die normale Ganggeschwindigkeit von Tyrannosaurus. Heraus kam eine gemächliche Geschwindigkeit von etwa 0,8 bis 1,64 m/s (2,9 bis 5,9 km/h), bei der das Tier vermutlich einen geringen Energieverbrauch hatte. Die maximale Geschwindigkeit ausgewachsener Tiere beträgt etwa 20–30 km/h und die schmaler gebauten Jungtiere erreichten wahrscheinlich noch sehr viel höhere Geschwindigkeiten.[75]
Ernährung
Wie bei vielen anderen großen fleischfressenden Dinosauriern herrscht Uneinigkeit darüber, ob Tyrannosaurus ein aktiver Jäger oder ein Aasfresser war.
Verfügbare Bissspuren an Fossilienknochen (teilweise sogar an anderen Tyrannosaurus-Knochen selbst), seine riesigen Zähne und großer Rachen scheinen für die Rolle eines Jägers zu sprechen. David Burnham von der Universität Kansas begründet dies in einer aktuellen Studie aus dem Jahr 2013: Mit seinen Mitarbeitern fand er bei Ausgrabungen einen abgebrochenen Tyrannosaurus-Zahn zwischen den Schwanzwirbeln des Hadrosauriers Edmontosaurus annectens. Da die Wirbel um den Zahn herum zusammengewachsen waren, ist seiner Meinung nach davon auszugehen, dass der T. rex die fliehende Beute verfolgt hat und, wie dies heute Raubtiere auch machen, zu Fall bringen wollte, indem es das Tier mit großen Wunden schwächte.[76]
Die Anordnung der Augen im Schädel spricht dafür, dass Tyrannosaurier mit beiden Augen ein Ziel fixieren konnten. Diese Eigenschaft ist typisch für jagende Tiere. Der Fund eines gut erhaltenen Schädels erlaubte zudem eine Rekonstruktion des Tyrannosaurusgehirns. Dabei wurde festgestellt, dass das Riechzentrum verhältnismäßig groß war. Vor einigen Jahren gemachte Funde von anderen großen, teilweise auch nahe mit Tyrannosaurus verwandten Theropoden deuten zudem darauf hin, dass diese in Familienverbänden gelebt haben, was ebenfalls ein starkes Indiz für ein spezialisiertes Jagdverhalten ist. Auch bei „Sue“, einem erwachsenen Exemplar eines Tyrannosaurus rex, wurden die Skelette eines Jungtiers und eines Halbwüchsigen gefunden.
Andere Wissenschaftler widersprechen hingegen der Rolle des aktiven Jägers. Ihrer Meinung nach sei Tyrannosaurus ein Aasfresser gewesen, der sich von bereits verendeten Tieren ernährt hat. Seine Größe und Kraft hätten es ihm ferner erlaubt, kleineren Fleischfressern ihre Beute abzujagen. Einer der bekanntesten Vertreter dieser Theorie ist der Paläontologe Jack Horner, der auch in den 1990er Jahren Steven Spielberg bei dessen Film Jurassic Park als Berater zur Seite stand. Er stützt sich dabei auf die Anatomie des Tyrannosaurus rex:
- Die Arme des Tyrannosaurus rex seien für einen Jäger ungeeignet. Wäre er beim Laufen einmal gestürzt, hätten ihn die kleinen Arme nicht abfangen können und auch das Wiederaufstehen hätte enorm viel Kraft gekostet.
- Die Zähne des Tyrannosaurus rex sind nicht wie bei Raubsauriern klingenförmig, sondern abgerundet. Dies deutet laut Horner darauf hin, dass die Zähne eher zum Zermahlen von Knochen und Knorpeln geeignet waren.
- Das Gehirn des Tyrannosaurus rex sei nicht so ausgeprägt wie das eines Räubers gewesen. Das Sehzentrum sei nicht für eine gute optische Erfassung von Beutetieren optimiert gewesen, stattdessen sei das Riechzentrum wie bei Aasfressern überproportional groß gewesen. Das einzige Tier, welches ein proportional noch größeres Riechzentrum habe, sei der Aasgeier. Tyrannosaurus rex konnte laut Horner Aas vermutlich auf eine Entfernung von bis zu 40 km riechen.
- Die Beine des Tyrannosaurus rex seien für kurze Sprints bei der Jagd ungeeignet. Das Verhältnis von Ober- zu Unterschenkelknochen ist nicht wie bei Sprintern. Bei einem Sprinter sind die Unterschenkelknochen länger als die Oberschenkelknochen, bei Tyrannosaurus rex ist das Verhältnis jedoch umgekehrt. Tyrannosaurus rex’ Beine seien demnach nicht für eine Verfolgung beziehungsweise das schnelle Attackieren von Beutetieren ausgelegt, sondern für lange Wanderungen zu Aas, das er auch aus großer Entfernung wahrnehmen konnte.[77][78]
Reine Aasfresser sind in der Tierwelt jedoch selten vertreten; ausschließlich von Aas leben zum Beispiel die Geier der Gattung Gyps, die mit minimalem Energieaufwand große Gebiete absuchen können, was einem bodenbewohnenden Tier nicht möglich wäre. Die meisten rezenten karnivoren Säugetiere und Vögel sowie viele Schuppenkriechtiere fressen auch frische Kadaver oder nutzen Gelegenheiten, einem kleineren Beutegreifer die Beute abzujagen.


Ähnlich kontrovers diskutiert wie die Aasfressertheorie ist die Frage, ob Tyrannosaurus rex ein Einzelgänger oder ein Rudeltier war. Zu den Vertretern der Rudeltiertheorie zählt ebenfalls Jack Horner. Die Theorie wird vor allem auf Fundstellen gestützt, an denen die Überreste mehrerer Exemplare gefunden wurden. Da Überreste anderer Saurier dort nicht gefunden wurden, ist die Erklärung ausgeschlossen, es handele sich um Orte, an denen lediglich aufgrund besonderer Gefahren viele Tiere verendeten, wie dies etwa bei Sümpfen, Mooren, den Füßen hoher Klippen oder Asphaltseen der Fall sein könnte. Ähnliche Fundstellen gibt es auch mit den Überresten anderer großer fleischfressender Saurier, beispielsweise Mapusaurus roseae, die aufgrund dieser Funde gemeinhin als Rudeltier betrachtet werden.[79][80] Neuere Funde solcher Fossillagerstätten mit Fossilien mehrerer Individuen stützen die Ansicht, dass Tyrannosaurus in parasozialen Familienverbänden lebte; die schnelleren Jungtiere könnten Beutetiere auf die adulten Tyrannosaurier zugetrieben haben, woraufhin diese die Beutetiere aus dem Hinterhalt mit einem Biss töteten.[81]
Nach einer Langzeitstudie der Universitäten Liverpool und Manchester erreichte Tyrannosaurus rex eine errechnete Beißkraft von 35.000 bis 57.000 Newton.[82] Diese Kraft wurde bei den Landraubtieren nur von den Urzeitkrokodilen, darunter Deinosuchus, übertroffen. In einer 2021 veröffentlichten Studie wurde für juvenile Tiere eine Beißkraft von 5600 Newton ermittelt. Dazu verwendeten die Forscher Bissspuren auf einem Edmontosaurus-Wirbel, die einem etwa 13 Jahre alten Tyrannosaurus zugeschrieben wurden, sowie einen Oberkieferzahn des ursprünglich als Nanotyrannus klassifizierten Exemplars „Jane“ (Katalognummer BMRP 2002.4.1.).[83]
Parasiten

Fossilien von Tyrannosaurus und anderen Tyrannosauriden zeigen oft Veränderungen an einer Seite oder beiden Seiten des Unterkiefers, glattrandige, erosive Läsionen, die sich durch die gesamte Dicke des Knochens ziehen. Bei „Sue“ werden diese Knochenveränderungen als Bissspuren von innerartlichen Kämpfen oder als Aktinomykose (eine bakterielle Infektion) gedeutet. Untersuchungen an zehn Individuen mit vergleichbaren Läsionen veranlassten Wolff et al. 2009 zu der alternativen Hypothese, dass eine mögliche Ursache auch eine Infektionskrankheit ähnlich der Trichomonose sein könnte, da die Läsionen mit denen bei heutigen Vögeln, insbesondere Greifvögeln, übereinstimmen, die mit dem entsprechenden Krankheitserreger (Trichomonas) infiziert sind. Im fortgeschrittenen Stadium seien die krankhaften Veränderungen und die damit einhergehenden funktionellen Einschränkungen an Maul, Rachen und Speiseröhre so erheblich gewesen, dass sie sehr wahrscheinlich zum Tod durch Verhungern geführt haben.[84]
Entdeckungsgeschichte
Henry Fairfield Osborn, damals Präsident des American Museum of Natural History, beschrieb und benannte Tyrannosaurus rex im Jahr 1905. Der Gattungsname leitet sich aus den griechischen Wörtern τυραννος (tyrannos – „König, Tyrann, Despot“) und σαυρος (sauros – „Echse“) ab. Als Epitheton wählte Osborn das lateinische Wort rex, was so viel wie „König“ bedeutet. Somit lässt sich Tyrannosaurus rex mit „König Königsechse“ übersetzen, was auf die enorme Körpergröße und die Dominanz dieses Tieres über andere Spezies seiner Zeit hinweist.[62]
Früheste Funde

Zähne, die heute Tyrannosaurus rex zugeschrieben werden, wurden bereits 1874 von A. Lakes nahe Golden in Colorado entdeckt. In den frühen 1890er Jahren sammelte J. B. Hatcher postkraniale Überreste im östlichen Wyoming, die anfangs einer großen Ornithomimus-Spezies zugeschrieben wurden (Ornithomimus grandis), aber heute Tyrannosaurus rex zugeschrieben werden. Fragmente der Wirbel, die Edward Drinker Cope 1892 im westlichen South Dakota fand und Manospondylus gigas nannte, wurden ebenfalls als Tyrannosaurus rex neu klassifiziert.[21]
Barnum Brown entdeckte das erste teilweise Skelett eines Tyrannosaurus rex im Jahr 1900 im östlichen Wyoming. Ein weiteres Skelett fand Brown im Jahr 1902 in der Hell Creek Formation in Montana. Henry Fairfield Osborn beschrieb beide Skelette im Jahr 1905 in ein und derselben Veröffentlichung. Während er das zweite, 1902 entdeckte Exemplar als Tyrannosaurus rex beschrieb, ordnete er das andere, 1900 entdeckte Skelett einer anderen neuen Art und Gattung zu, die er Dynamosaurus imperiosus nannte.[62] 1906 erkannte Osborn, dass beide Skelette zur selben Art gehörten, und wählte Tyrannosaurus als gültigen Namen.[64] Die Dynamosaurus-Knochen befinden sich heute in der Sammlung des Natural History Museums in London.[86]
Insgesamt entdeckte Brown fünf Teilskelette von Tyrannosaurus. Im Jahr 1941 wurde Browns Fund aus 1902, das Holotyp-Exemplar, an das Carnegie Museum of Natural History in Pittsburgh (Pennsylvania) verkauft. Browns vierter und größter Fund, ebenfalls aus der Hell-Creek-Formation, ist im American Museum of Natural History in New York zu besichtigen.[87]
Jüngere Funde
Obwohl zahlreiche Skelette entdeckt wurden, ist nur ein einziger Fußabdruck dokumentiert, der sich in der Philmont Scout Ranch im nordöstlichen New Mexico befindet. Dieser Abdruck wurde 1983 entdeckt und 1994 identifiziert und dokumentiert.[88]
Im Jahre 1990 wurde mit „Sue“ das bislang besterhaltene Skelett eines Tyrannosaurus rex gefunden.
Das 1987 gefundene Skelett von „Stan“ wurde 2020 für 27,5 Millionen Dollar versteigert.[89]
Ein 2012 in Montana gemachter Fund enthielt den bislang am besten erhaltenen Schädel. Die auf den Namen „Tristan Otto“ getauften Überreste werden seit Juli 2015 im Museum für Naturkunde in Berlin untersucht und sind seit Dezember 2015 dort ausgestellt.
Im Jahr 2019 wurde eine Studie zu einem bereits 1991 im kanadischen Saskatchewan entdeckten Skelett publik gemacht. Es war in so massiven Sandstein eingebettet, dass die Freilegung mehr als 10 Jahre dauerte. RSM P2523.8 („Scotty“) bekam seinen Namen durch die Feier beim Skelettfund, bei der auf „Scotty“ mit einer Flasche Scotch angestoßen wurde. „Scotty“ ist zu 65 % erhalten und übertrifft den bisherigen Größenrekordhalter „Sue“. Mit einer Länge von mindestens 13 Metern und einem geschätzten Gewicht von über 8,8 Tonnen ist „Scotty“ der größte und schwerste bisher entdeckte Tyrannosaurus weltweit.[9]
Popularität


Tyrannosaurier allgemein und die Art Tyrannosaurus rex im Besonderen gehören zu den weltweit populärsten Dinosauriern. Auf viele Menschen trifft wohl zu, dass Tyrannosaurus rex der einzige Saurier ist, dessen vollständigen wissenschaftlichen Namen sie kennen. In der modernen Popkultur ist Tyrannosaurus rex noch immer der Inbegriff der „tödlichen Fressmaschine“ und eine popkulturelle Ikone, ein Inbegriff für Stärke und Überlegenheit. Daran konnten bisher auch die neueren Theorien und Erkenntnisse über das mögliche Leben als Aasfresser nicht viel ändern.
Nachhaltig geprägt wurde das Image des Tyrannosaurus rex auch durch die tricktechnisch revolutionäre Darstellung im Film Jurassic Park (1993). Hier wurde er als enorm schnelle, übermächtige Kreatur charakterisiert, vor der die Menschen nur fliehen konnten. Auch schufen die Sounddesigner des Films ein weithin bekanntes, charakteristisches Brüllen für das Tier.
Literatur
- John R. Horner, Don Lessem: The Complete T. Rex. Simon & Schuster, New York NY u. a. 1993, ISBN 0-671-74185-3.
- William L. Abler: The Teeth of the Tyrannosaurs. In: Gregory S. Paul (Hrsg.): The Scientific American book of Dinosaurs. St. Martin’s Griffin, New York NY 2000, ISBN 0-312-26226-4, S. 276–278.
- Gregory M. Erickson: Breathing Life into Tyrannosaurus Rex. In: Gregory S. Paul (Hrsg.): The Scientific American book of Dinosaurs. St. Martin’s Griffin, New York NY 2000, ISBN 0-312-26226-4, S. 267–275.
- Rebecca R. Hanna: Dinosaurs Got Hurt Too. In: Gregory S. Paul (Hrsg.): The Scientific American book of Dinosaurs. St. Martin’s Griffin, New York NY 2000, ISBN 0-312-26226-4, S. 119–126.
- David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2. Ausgabe. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2.
Weblinks
- Tyrannosaurus rex „Sue“ (Memento vom 13. Juli 2004 im Internet Archive) im Field Museum von Chicago (englisch)
- Ulrich Dewald: Schlechte Nachrichten für den T. rex. In: wissenschaft.de. 9. Februar 2006.
- Cornelia Pfaff: Neue Studie: Es gab genug Aas für T. Rex. In: wissenschaft.de. 13. Februar 2003.
Einzelnachweise
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs. Princeton University Press, Princeton NJ u. a. 2010, ISBN 978-0-691-13720-9, S. 69, Online.
- Wilhelm Gemoll: Griechisch-Deutsches Schul- und Handwörterbuch. 9. Auflage, durchgesehene und erweiterte von Karl Vretska. Mit einer Einführung in die Sprachgeschichte von Heinz Kronasser. Freytag u. a., München u. a. 1965.
- Erich Pertsch: Langenscheidts Großes Schulwörterbuch. Lateinisch-Deutsch. 3. Auflage. Langenscheidt, Berlin u. a. 1978, ISBN 3-468-07201-5.
- Gregory S. Paul, W. Scott Persons, Jay Van Raalte: The Tyrant Lizard King, Queen and Emperor: Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus. In: Evolutionary Biology. 1. März 2022, ISSN 1934-2845, doi:10.1007/s11692-022-09561-5.
- Christopher A. Brochu, Richard A. Ketcham: Osteology of Tyrannosaurus rex. Insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull (= Society of Vertebrate Paleontology. Memoir. 7, ISSN 1062-161X = Journal of Vertebrate Paleontology. Bd. 22, Nr. 4, Supplement). Society of Vertebrate Paleontology, Northbrook IL 2002.
- Sue’s vital statistics. In: Sue at the Field Museum. Field Museum of Natural History, abgerufen am 17. Dezember 2010.
- Gregory M. Erickson, Peter J. Makovicky, Philip J. Currie, Mark A. Norell, Scott A. Yerby, Christopher A. Brochu: Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. In: Nature. Bd. 430, Nr. 7001, 2004, S. 772–775, doi:10.1038/nature02699.
- Charles R. Marshall, Daniel V. Latorre, Connor J. Wilson, Tanner M. Frank, Katherine M. Magoulick: Absolute abundance and preservation rate of Tyrannosaurus rex. In: Science. Band 372, Nr. 6539, 16. April 2021, S. 284–287, doi:10.1126/science.abc8300 (sciencemag.org [abgerufen am 22. April 2021]).
- W. Scott Persons, Philip J. Currie, Gregory M. Erickson: An Older and Exceptionally Large Adult Specimen of Tyrannosaurus rex. In: The Anatomical Record. Band 0, ja, doi:10.1002/ar.24118 (wiley.com [abgerufen am 26. März 2019]).
- John R. Hutchinson, Karl T. Bates, Julia Molnar, Vivian Allen, Peter J. Makovicky: A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth. In: PLoS ONE. Bd. 6, Nr. 10, 2011, e26037, doi:10.1371/journal.pone.0026037. Dazu: Corrections. In: PLoS ONE. Bd. 9, Nr. 5, 2014, e97055, doi:10.1371/journal.pone.0097055.
- Thomas R. Holtz Jr.: Winter 2011 Appendix. 2012. zu: Thomas R. Holtz jr.: Dinosaurs. The most complete, up-to-date Encyclopedia for Dinosaur Lovers of all Ages. Random House, New York NY 2007, ISBN 978-0-375-82419-7 (PDF; 704 kB).
- John F. Anderson, Anthony Hall-Martin, Dale A. Russell: Long-bone circumference and weight in mammals, birds and dinosaurs. In: Journal of Zoology. Bd. 207, Nr. 1, 1985, S. 53–61, doi:10.1111/j.1469-7998.1985.tb04915.x.
- Robert T. Bakker: The dinosaur Heresies. New Theories unlocking the Mystery of the Dinosaurs and their Extinction. Morrow, New York NY 1986, ISBN 0-688-04287-2.
- Donald M. Henderson: Estimating the masses and centers of mass of extinct animals by 3-D mathematical slicing Paleobiology. In: Paleobiology. Bd. 25, Nr. 1, 1999, ISSN 0094-8373, S. 88–106, Abstract.
- James O. Farlow, Matt B. Smith, John M. Robinson: Body mass, bone „strength indicator“, and cursorial potential of Tyrannosaurus rex. In: Journal of Vertebrate Paleontology. Bd. 15, Nr. 4, 1995, S. 713–725, doi:10.1080/02724634.1995.10011257.
- Frank Seebacher: A new method to calculate allometric length-mass relationships of dinosaurs. In: Journal of Vertebrate Paleontology. Bd. 21, Nr. 1, 2001, S. 51–60, doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2.
- Per Christiansen, Richard A. Fariña: Mass prediction in theropod dinosaurs. In: Historical Biology. Bd. 16, Nr. 2/4, 2004, S. 85–92, doi:10.1080/08912960412331284313.
- Gregory S. Paul: Tyrannosaurus, the Lean Killing Machine. 2011, online.
- Cristiano Dal Sasso, Simone Maganuco, Eric Buffetaut, Marco A. Mendez: New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its size and affinities. In: Journal of Vertebrate Paleontology. Bd. 25, Nr. 4, 2005, S. 888–896, doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2, online.
- Jorge O. Calvo, Rodolfo Coria: New specimen of Giganotosaurus carolinii (Coria & Salgado, 1995), supports it as the as the largest theropod ever found. In: Gaia. Revista de Geociências. Bd. 15, 1998, S. 117–122, Digitalisat (PDF; 389 KB) (Memento vom 9. März 2012 im Internet Archive).
- Brent H. Breithaupt, Elizabeth H. Southwell, Neffra A. Matthews: In Celebration of 100 years of Tyrannosaurus Rex: Manospondylus Gigas, Ornithomimus Grandis, and Dynamosaurus Imperiosus, the Earliest Discoveries of Tyrannosaurus Rex in the West. (2005 Salt Lake City Annual Meeting (October 16–19, 2005)). In: The Geological Society of America. Abstracts with Programs. Bd. 37, Nr. 7, S. 406, online.
- Museum unveils world’s largest T-rex skull. In: Montana State University. Archiviert vom Original am 14. April 2006; abgerufen am 13. September 2008.
- Kent A. Stevens: Binocular vision in theropod dinosaurs. In: Journal of Vertebrate Paleontology. Bd. 26, Nr. 2, 2006, S. 321–330, doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2, Digitalisat (PDF; 1,2 MB) (Memento vom 4. März 2009 im Internet Archive).
- Eric Jaffe: Sight for 'saur eyes: T. rex vision was among nature’s best. In: Science News. Bd. 170, Nr. 1, 2006, S. 3–4, doi:10.2307/4017288.
- Eric Snively, Donald M. Henderson, Doug S. Phillips: Fused and vaulted nasals of tyrannosaurid dinosaurs: Implications for cranial strength and feeding mechanics. In: Acta Palaeontologica Polonica. bd. 51, Nr. 3, 2006, S. 435–454, (PDF; 683 KB).
- Gregory M. Erickson, Samuel D. Van Kirk, Jinntung Su, Marc E. Levenston, William E. Caler, Dennis R. Carter: Bite-force estimation for Tyrannosaurus rex from tooth-marked bones. In: Nature. Bd. 382, Nr. 6593, 1996, S. 706–708, doi:10.1038/382706a0.
- Mason B. Meers: Maximum bite force and prey size of Tyrannosaurus rex and their relationships to the inference of feeding behavior. In: Historical Biology. Bd. 16, Nr. 1, 2002, S. 1–12, doi:10.1080/0891296021000050755, Abstract.
- Thomas R. Holtz jr.: The Phylogenetic Position of the Tyrannosauridae: Implications for Theropod Systematics. In: Journal of Paleontology. Bd. 68, Nr. 5, 1994, ISSN 0022-3360, S. 1100–1117.
- Gregory S. Paul: Predatory Dinosaurs of the World. A complete and illustrated Guide. Simon & Schuster, New York NY u. a. 1988, ISBN 0-671-61946-2.
- Joshua B. Smith: Heterodonty in Tyrannosaurus rex: implications for the taxonomic and systematic utility of theropod dentitions. In: Journal of Vertebrate Paleontology. Bd. 25, Nr. 4, 2005, S. 865–887, doi:10.1671/0272-4634(2005)025[0865:HITRIF]2.0.CO;2, Digitalisat (PDF; 1,2 MB) (Memento vom 4. März 2009 im Internet Archive).
- Kate Douglas, Stephen Young: The dinosaur detectives. In: New Scientist. Bd. 158, Nr. 2130, 18. April 1998, S. 24, online, One palaeontologist memorably described the huge, curved teeth of T. rex as ‚lethal bananas‘.
- Philip J. Currie, Jørn H. Hurum, Karol Sabath: Skull structure and evolution in tyrannosaurid dinosaurs. In: Acta Palaeontologica Polonica. Bd. 48, Nr. 2, 2003, S. 227–234, (PDF; 137 kB).
- Thomas R. Holtz jr.: Tyrannosauroidea. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 111–136.
- Евгений А. Малеев: Гигантские хищные динозавры Монголии. In: Доклады Академии наук СССР. Bd. 104, Nr. 4, 1955, ISSN 0002-3264, S. 634–637.
- Анатолий К. Рождественский: Возрастная изменчивость и некоторые вопросы систематики динозавров Азии. In: Палеонтологический Журнал. Nr. 3, 1965, ISSN 0031-031X, S. 95–109.
- Ken Carpenter: Tyrannosaurids (Dinosauria) of Asia and North America. In: Niall J. Mateer, Chen Pei-ji (Hrsg.): Aspects of Nonmarine Cretaceous Geology. China Ocean Press, Beijing 1992, ISBN 7-5027-1463-4, S. 250–268.
- Thomas D. Carr, Thomas E. Williamson, David R. Schwimmer: A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama. In: Journal of Vertebrate Paleontology. Bd. 25, Nr. 1, 2005, S. 119–143, doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2.
- Jørn H. Hurum, Karol Sabath: Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared. In: Acta Palaeontologica Polonica. Bd. 48, Nr. 2, 2003, S. 161–190.
- George Olshevsky, Tracy L. Ford: The origin and evolution of the tyrannosaurids, part 1. In: Kyoryugaku Saizensen. = Dinosaur Frontline. Bd. 9, 1995, S. 92–119 (japanisch).
- Thomas D. Carr, Thomas E. Williamson: Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America. In: Zoological Journal of the Linnean Society. Bd. 142, Nr. 4, 2004, S. 479–523, doi:10.1111/j.1096-3642.2004.00130.x.
- Asher Elbein: They Want to Break T. Rex Into 3 Species. Other Paleontologists Aren’t Pleased. In: The New York Times. 1. März 2022, ISSN 0362-4331 (nytimes.com [abgerufen am 1. März 2022]).
- Charles W. Gilmore: A new carnivorous dinosaur from the Lance Formation of Montana (= Smithsonian Miscellaneous Collections. Bd. 106, Nr. 13, ISSN 0096-8749 = Smithsonian Institution. Publication. 3857). Smithsonian Institution, Washington DC 1946, Digitalisat.
- Robert T. Bakker, Michael Williams, Philip J. Currie: Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana. In: Hunteria. Bd. 1, Nr. 5, 1988, ZDB-ID 1251702-1, S. 1–30.
- Philip J. Currie, Jørn H. Hurum, Karol Sabath: Skull structure and evolution in tyrannosaurid dinosaurs. In: Acta Palaeontologica Polonica. Bd. 48, Nr. 2, 2003. S. 227–234. Abgerufen am 28. März 2020.
- Thomas D. Carr: Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria). In: Journal of Vertebrate Paleontology. Bd. 19, Nr. 3, 1999, S. 497–520, doi:10.1080/02724634.1999.10011161.
- Holly N. Woodward, Katie Tremaine, Scott A. Williams, Lindsay E. Zanno, John R. Horner: Growing up Tyrannosaurus rex: Osteohistology refutes the pygmy “Nanotyrannus” and supports ontogenetic niche partitioning in juvenile Tyrannosaurus. In: Science Advances. Band 6, Nr. 1, 1. Januar 2020, S. eaax6250, doi:10.1126/sciadv.aax6250 (sciencemag.org [abgerufen am 28. März 2020]).
- Henry Fairfield Osborn: Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus. In: Bulletin of the American Museum of Natural History. Bd. 35, Artikel 43, 1917, S. 733–771, Digitalisat.
- International Union of Biological Sciences (Hrsg.): International code of zoological nomenclature. 4th edition. International Trust for Zoological Nomenclature, London 1999, ISBN 0-85301-006-4, Article 23.9: Reversal of Precedence, online.
- Mike Taylor: So why hasn’t Tyrannosaurus been renamed Manospondylus?. In: The Dinosaur FAQ. 27. August 2002. Abgerufen am 8. Oktober 2008.
- John R. Horner, Kevin Padian: Age and growth dynamics of Tyrannosaurus rex. In: Proceedings of the Royal Society of London. Series B: Biological Sciences. Band 271, Nummer 1551, September 2004, S. 1875–1880, doi:10.1098/rspb.2004.2829, PMID 15347508, PMC 1691809 (freier Volltext).
- Mary H. Schweitzer, Jennifer L. Wittmeyer, John R. Horner: Gender-Specific Reproductive Tissue in Ratites and Tyrannosaurus rex. In: Science. Bd. 308, Nr. 5727, 2005, S. 1456–1460, doi:10.1126/science.1112158, PMID 15933198.
- Andrew H. Lee, Sarah Werning: Sexual maturity in growing dinosaurs does not fit reptilian growth models. In: Proceedings of the National Academy of Sciences. Bd. 105, Nr. 2, 2008, S. 582–587, doi:10.1073/pnas.0708903105, PMID 18195356.
- Gregory M. Erickson, Philip J. Currie, Brian D. Inouye, Alice A. Winn: Tyrannosaur Life Tables: An Example of Nonavian Dinosaur Population Biology. In: Science. Bd. 313, Nr. 5784, 2006, S. 213–217, doi:10.1126/science.1125721, PMID 16840697.
- Kenneth Carpenter: Variation in Tyrannosaurus rex. In: Kenneth Carpenter, Philip J. Currie (Hrsg.): Dinosaur Systematics. Approaches and Perspectives. Cambridge University Press, Cambridge u. a. 1990, ISBN 0-521-43810-1, S. 141–145.
- Peter L. Larson: Tyrannosaurus sex. In: Gary D. Rosenberg, Donald L. Wolberg (Hrsg.): Dino Fest (= Paleontological Society. Special Publication. Bd. 7, ZDB-ID 2002145-8). Proceedings of a Conference for the general Public held March 24–26, 1994 at Indianapolis, Indiana. University of Tennessee, Knoxville TN 1994, S. 139–155.
- Gregory M. Erickson, A. Kristopher Lappin, Peter Larson: Androgynous rex – The utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs. In: Zoology. Bd. 108, Nr. 4, 2005, S. 277–286, doi:10.1016/j.zool.2005.08.001.
- Mary H. Schweitzer, Ruth M. Elsey, Christopher G. Dacke, John R. Horner, Ellen-Therese Lamm: Do egg-laying crocodilian (Alligator mississippiensis) archosaurs form medullary bone? In: Bone. Bd. 40, Nr. 4, 2007, S. 1152–1158, doi:10.1016/j.bone.2006.10.029.
- Joseph Leidy: Cretaceous reptiles of the United States (= Smithsonian Contributions to Knowledge. Bd. 14, Nr. 6 = 192, ISSN 0096-9354). Smithsonian Institution, Washington DC 1865.
- Tyrannosaurus. American Museum of Natural History, archiviert vom Original am 30. Juni 2009; abgerufen am 27. Juli 2014.
- B. H. Newman: Stance and gait in the flesh-eating dinosaur Tyrannosaurus. In: Biological Journal of the Linnean Society. Bd. 2, Nr. 2, 1970, S. 119–123, doi:10.1111/j.1095-8312.1970.tb01707.x.
- The Age of Reptiles Mural. Yale University. 2008. Abgerufen am 27. Juli 2014.
- Henry Fairfield Osborn: Tyrannosaurus and other Cretaceous carnivorous dinosaurs. In: Bulletin of the American Museum of Natural History. Bd. 21, Article 14, 1905, S. 259–265, Digitalisat.
- Lawrence M. Lambe: On a new Genus and Species of Carnivorous Dinosaur from the Belly River Formation of Alberta, with a Description of the Skull of Stephanosaurus Marginatus from the same Horizon. In: The Ottawa Naturalist. Bd. 28, Nr. 1, 1914, S. 13–20, Digitalisat.
- Henry Fairfield Osborn: Tyrannosaurus, Upper Cretaceous carnivorous dinosaur. (Second Communication.) In: Bulletin of the American Museum of Natural History. Bd. 22, Article 16, 1906, S. 281–296, Digitalisat.
- Kenneth Carpenter, Matt Smith: Forelimb Osteology and Biomechanics of Tyrannosaurus rex. In: Darren Tanke, Kenneth Carpenter (Hrsg.): Mesozoic vertebrate life. Indiana University Press u. a., Bloomington IN u. a. 2001, ISBN 0-253-33907-3, S. 90–116.
- Helen Fields: Dinosaur Shocker. In: Smithsonian Magazine. Mai 2006. Abgerufen am 27. Juli 2014.
- Mary H. Schweitzer, Jennifer L. Wittmeyer, John R. Horner, Jan K. Toporski: Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex. In: Science. Bd. 307, Nr. 5717, 2005, S. 1952–1955, doi:10.1126/science.1108397, PMID 15790853.
- John M. Asara, Mary H. Schweitzer, Lisa M. Freimark, Matthew Phillips, Lewis C. Cantley: Protein Sequences from Mastodon and Tyrannosaurus Rex Revealed by Mass Spectrometry. In: Science. Bd. 316, Nr. 5822, 2007, S. 280–285, doi:10.1126/science.1137614, PMID 17431180.
- Paul Rincon: Protein links T. rex to chickens. In: BBC News. 12. April 2007, abgerufen am 27. Juli 2014.
- C. L. Organ, M. H. Schweitzer, W. Zheng, L. M. Freimark, L. C. Cantley, J. M. Asara: Molecular phylogenetics of mastodon and Tyrannosaurus rex. Science. 25. April 2008; Studie: doi:10.1038/4541035a; PMID 18436782.
- Thomas G. Kaye, Gary Gaugler, Zbigniew Sawlowicz: Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms. In: PLoS ONE. Bd. 3, Nr. 7, 2008, e2808, doi:10.1371/journal.pone.0002808.
- New Research Challenges Notion That Dinosaur Soft Tissues Still Survive. Newswise. 27. Juli 2014. Abgerufen am 27. Juli 2014.
- Darren Tanke Tanke: Tyrannosaur Skin Impression Found In Alberta. 25. März 1996. Archiviert vom Original am 7. September 2008. Abgerufen am 27. Juli.
- Xing Xu, Mark A. Norell, Xuewen Kuang, Xiaolin Wang, Qi Zhao, Chengkai Jia: Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. In: Nature. Bd. 431, Nr. 7009, 2004, S. 680–684, doi:10.1038/nature02855, PMID 15470426.
- Pasha A. van Bijlert, A. J. ‘Knoek’ van Soest, Anne S. Schulp: Natural Frequency Method: estimating the preferred walking speed of Tyrannosaurus rex based on tail natural frequency. In: Royal Society Open Science. Band 8, Nr. 4, S. 201441, doi:10.1098/rsos.201441 (royalsocietypublishing.org [abgerufen am 22. April 2021]).
- Robert A. DePalma II, David A. Burnham, Larry D. Martin, Bruce M. Rothschild, Peter L. Larson: Physical evidence of predatory behavior in Tyrannosaurus rex. In: Proceedings of the National Academy of Sciences. Bd. 110, Nr. 31, 2013, S. 12560–12564, doi:10.1073/pnas.1216534110, Digitalisat (PDF; 404 KB).
- John R. Horner: Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger). In: Gary D. Rosenberg, Donald L. Wolberg (Hrsg.): Dino Fest (= Paleontological Society. Special Publication. Bd. 7). Proceedings of a Conference for the general Public held March 24–26, 1994 at Indianapolis, Indiana. University of Tennessee, Knoxville TN 1994, S. 157–164.
- War der Tyrannosaurus Rex nur ein Aasfresser? In: Fundgroup.de. Das Fun-Stuff Verzeichnis.
- Rodolfo A. Coria, Philip J. Currie: A new carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina. In: Geodiversitas. Bd. 28, Nr. 1, 2006, ISSN 1280-9659, S. 71–118, (PDF; 2,9 MB).
- Gerd Braune: Konkurrenz für die Tyrannosaurier. In: Berliner Zeitung, 10. Mai 2006.
- Alan L. Titus, Katja Knoll, Joseph J.W. Sertich, Daigo Yamamura, Celina A. Suarez: Geology and taphonomy of a unique tyrannosaurid bonebed from the upper Campanian Kaiparowits Formation of southern Utah: implications for tyrannosaurid gregariousness. In: PeerJ. Band 9, 19. April 2021, S. e11013, doi:10.7717/peerj.11013 (peerj.com [abgerufen am 22. April 2021]).
- Karl T. Bates, Peter L. Falkingham: Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics. In: Biology Letters. Bd. 8, Nr. 4, 2012, S. 660–664, doi:10.1098/rsbl.2012.0056.
- Joseph E. Peterson, Z. Jack Tseng, Shannon Brink: Bite force estimates in juvenile Tyrannosaurus rex based on simulated puncture marks. In: PeerJ. Band 9, 2. Juni 2021, ISSN 2167-8359, S. e11450, doi:10.7717/peerj.11450 (peerj.com [abgerufen am 7. Juni 2021]).
- Ewan D. S. Wolff, Steven W. Salisbury, John R. Horner, David J. Varricchio: Common Avian Infection Plagued the Tyrant Dinosaurs. In: PLoS ONE. Bd. 4, Nr. 9, 2009, e7288, doi:10.1371/journal.pone.0007288.
- The First Tyrannosaurus Skeleton, 1905. Linda Hall Library of Science, Engineering and Technology. Archiviert vom Original am 28. September 2006. Abgerufen am 3. August 2008.
- Brent H. Breithaupt, Elizabeth H. Southwell, Neffra A. Matthews: Dynamosaurus Imperiosus and the earliest discoveries of Tyrannosaurus Rex in Wyoming and the West. In: New Mexico Museum of Natural History and Science. Bulletin. 35, 2006, S. 257–258, online.
- John R. Horner, Don Lessem: The complete T. rex. Simon & Schuster, New York NY u. a. 1993, ISBN 0-671-74185-3.
- Footprint of a Giant. In: Online guide to the continental Cretaceous-Tertiary boundary in the Raton basin, Colorado and New Mexico. United States Geological Survey. Archiviert vom Original am 29. Februar 2012. Abgerufen am 9. Oktober 2008.
- T-Rex-Skelett erzielt Rekordsumme