Bärtierchen
Bärtierchen (Tardigrada) – auch Wasserbären genannt – bilden einen Tierstamm innerhalb der Häutungstiere (Ecdysozoa). Die meistens weniger als einen Millimeter großen achtbeinigen Tiere erinnern durch ihr Aussehen und ihre tapsig wirkende Fortbewegungsweise etwas an Bären, was zu ihrer Bezeichnung im deutschen Sprachraum führte. Auch ihr wissenschaftlicher Name (Zusammensetzung aus lateinisch tardus ‚langsam‘ und gradus ‚Schritt‘) geht auf die langsame Fortbewegung zurück.
| Bärtierchen | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Das Bärtierchen Milnesium tardigradum | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Tardigrada | ||||||||||||
| Spallanzani, 1777 | ||||||||||||
| Klassen | ||||||||||||
|
Sie leben weltweit im Meer, Süßwasser oder in feuchten Lebensräumen an Land; besonders häufig findet man sie dort in Mooskissen. Eine Eigenschaft der Tiere ist die Kryptobiose, ein todesähnlicher Zustand, in dem sie extreme Umweltbedingungen überdauern können. Bärtierchen können sich sowohl vom Inhalt von Pflanzenzellen ernähren als auch räuberisch von kleinen Tieren wie Fadenwürmern (Nematoda) oder Rädertierchen (Rotifera), die sie dazu anstechen und aussaugen. Bärtierchen pflanzen sich meistens geschlechtlich fort. Manche Arten vermehren sich aber auch parthenogenetisch, das heißt ohne Beteiligung von Männchen; die Eier der Weibchen entwickeln sich in diesem Fall ohne Befruchtung.
Die nächsten rezenten Verwandten der Bärtierchen sind vermutlich Glieder- (Arthropoda) und Stummelfüßer (Onychophora), mit denen sie das Taxon Panarthropoda bilden.
Aufbau
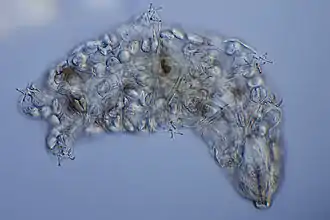
Die Größe von Bärtierchen liegt zwischen 50 Mikrometern bei einigen Jungtieren und 1,5 Millimetern bei einer Art der Gattung Macrobiotus, beträgt aber meist etwa 100 bis 500 Mikrometer, so dass die Tiere zur Meiofauna gezählt werden. Sie haben einen relativ plumpen, zylindrisch geformten Körper, der bauchseitig abgeflacht ist und oberflächlich gesehen aus vier Körpersegmenten mit je einem Beinpaar und einem Kopfsegment zu bestehen scheint. Dieser äußere, durch Querfalten in der äußersten Hautschicht hervorgerufene Eindruck ist allerdings irreführend: Der nur wenig vom Rest des Körpers abgesetzte und differenzierte Kopf besteht wahrscheinlich nicht aus einem, sondern aus drei miteinander verwachsenen Segmenten, der Körper könnte dagegen nicht aus vier, sondern möglicherweise aus fünf Teilen zusammengesetzt sein.
Meereslebende Arten sind meistens farblos oder unauffällig weiß bis grau gefärbt, während sich bei land- und süßwasserlebenden Formen oft Farben wie Rot, Gelb, Grün, Rosa, Lila oder Schwarz finden. Die Färbung wird entweder durch Pigmente in der Außenhaut, der Cuticula, oder durch den farbigen Inhalt der Leibeshöhle, des Hämocoeloms, oder des Darms hervorgerufen. Oft wandern auch Farbstoffe aus dem Darm in das Hämocoelom und werden von dort aus in der Cuticula abgelagert.
Charakteristischerweise sind viele Gewebe der Bärtierchen eutelisch, das heißt, die Zahl der Zellen, aus denen sie bestehen, ist genetisch festgelegt. Größenwachstum kann daher nicht durch eine Vermehrung der Zellenanzahl, sondern nur durch ein Wachstum der individuellen Zellen selbst stattfinden. Wird (durch experimentellen Eingriff) nach der ersten Zellteilung eine der beiden Tochterzellen abgetötet, entwickelt sich dennoch ein anatomisch vollständiges und lebensfähiges Tier, das dann nur halb so viele Zellen enthält wie normal.[1]
Extremitäten
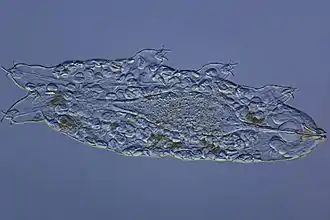
An vier Rumpfsegmenten entspringt am linken und rechten Rand der Bauchseite (ventrolateral) je ein „Stummelbein“, eine gelenklose Ausstülpung des Rumpfes, wie sie sich auch bei der verwandten Gruppe der Stummelfüßer findet. Gelegentlich werden einzelne Regionen des Beines unterschieden, die man dann als Coxa ("Hüfte"), Femur ("Oberschenkel"), Tibia ("Schienbein") oder Tarsus ("Fuß") bezeichnet. Da die Beine nur wenig differenziert sind, ist die anatomische Grundlage dieser Begriffe fraglich; in jedem Fall sind sie jedoch rein deskriptiv zu verstehen, deuten also nicht auf eine evolutionäre Verwandtschaft mit den entsprechenden Beinbestandteilen der Gliederfüßer hin.
Viele im Meer lebende Arten sind in der Lage, ihre Beine teleskopartig ineinanderzuschieben; dabei helfen ihnen die bei allen Bärtierchen innerhalb des Beines verlaufenden kurzen Muskeln. Am Beinende sitzen meistens vier bis acht, seltener bis zu dreizehn einziehbare Klauen, die manchmal an dünnen Beinauswüchsen, den „Zehen“, sitzen und von speziellen Drüsen, den „Klauendrüsen“, sezerniert werden. Bei einigen Arten sind die Klauen sekundär zu Haftscheiben abgewandelt; bei der Gattung Apodibius sind keine Klauen vorhanden. Oft zeigen die Klauen des vierten Beinpaares in die umgekehrte Richtung wie diejenigen der ersten drei Paare.
Haut

Nach außen wird der Körper durch eine nicht-zellige Außenschicht, die Cuticula, und eine darunterliegende, aus individuellen Zellen bestehende Innenschicht, die Epidermis, begrenzt. Begeißelte Zellen treten in der Bärtierchen-Haut grundsätzlich nicht auf.
Die Cuticula ist 0,5 Mikrometer dick und besteht aus drei Schichten, der äußeren Epicuticula mit einer Stärke von etwa 0,1 Mikrometern, einer darunterliegenden Intracuticula mit einer Dicke von ungefähr 0,2 Mikrometern und einer abschließenden Procuticula, die eine Stärke von etwa 0,25 Mikrometer aufweist. Sie setzt sich aus Chitin, verschiedenen Glykoproteinen, Polysacchariden, Mucopolysacchariden, Lipiden und Lipoproteinen zusammen und enthält bei vielen land- und süßwasserlebenden Arten auch Pigmente. Oft ist sie durch Höcker, Dornen oder Grübchen ornamentiert und bei manchen Arten rückseitig (dorsal), seltener seitlich (lateral), sehr selten auch bauchseitig (ventral) zu dicken Panzerplatten, den Skleriten verhärtet; diese sind dann wie bei den Gliederfüßern gelenkig verbunden. Die Cuticula wird zusammen mit den Beinklauen und der Auskleidung des Vorder- und Hinterdarms einschließlich der in Ersterem enthaltenen Mundwerkzeuge, der Stilette, regelmäßig gehäutet.
Die Epidermis besteht aus einer eutelischen, also innerhalb einer Art immer aus der gleichen Zellzahl bestehenden, einfachen Zellschicht, welche die darüberliegende Cuticula sezerniert. Sie ist von der Leibeshöhle nur durch eine dünne extrazelluläre Trennschicht, die sogenannte Basallamina, abgeteilt.
Hämocoelom
Die Leibeshöhle ist kein echtes Coelom, das heißt, sie ist nicht von einem auf embryonales Mesoderm zurückführbaren Gewebe ausgekleidet. Alle Organe sind von ihr lediglich durch die dünne Basallamina getrennt. Man bezeichnet die Leibeshöhle daher als Pseudo- oder, da sie mit „Blut“ gefüllt ist, als Hämocoelom. Ein echtes Coelom findet sich nur um die einzige Keimdrüse (Gonade) herum.
Das Blut ist farblos und dient nicht dem Sauerstofftransport. Es zirkuliert durch Körperbewegungen; ein eigentlicher Blutkreislauf existiert aber ebenso wenig wie separate Blutgefäße oder ein Herz. Dafür ist es mit bis zu 200 frei schwebenden oder an der Basallamina verankerten Zellen, den Hämozyten, gefüllt, deren erste Aufgabe die Nährstoffspeicherung zu sein scheint; möglicherweise spielen sie auch bei der Bakterienbekämpfung eine Rolle.
Muskulatur
Anders als bei den nahe verwandten Stummelfüßern treten Muskeln bei den Bärtierchen niemals in dicken Muskelschichten auf. Stattdessen besteht die Muskulatur des Rumpfs und der Gliedmaßen aus vereinzelten, dünnen, an der Cuticula befestigten Längsmuskeln, die aus nur wenigen, manchmal sogar nur aus einer einzigen Muskelzelle bestehen. Sie arbeiten entweder gegen das Hämocoel, das wie ein flüssigkeitsgefüllter Ballon als Hydroskelett wirkt, oder antagonistisch gegeneinander wie die Armbeuge- und -streckmuskulatur des Menschen. Ringmuskulatur findet man bei Bärtierchen nicht. Die Muskelzellen sind meistens glatt, selten quergestreift, wobei Letzteres vermutlich der ursprüngliche Zustand ist; glatte Muskulatur ist bei vielen Arten nach einer verbreiteten Hypothese erst sekundär als Anpassung an das Landleben entstanden.
Atmung
Muskeln benötigen für ihre Arbeit Sauerstoff, dessen durchschnittlicher Konsum bei Bärtierchen auf zwischen 0,05 und 0,1 Kubikmillimeter pro Stunde und Milligramm Körpergewicht geschätzt wird. Aufgrund des sehr günstigen Verhältnisses von Körperoberfläche und -volumen sind jedoch keine separaten Atmungsorgane notwendig und dementsprechend auch nicht vorhanden. Der gesamte Gasaustausch kann durch einfache Diffusion über die Haut stattfinden, die dazu allerdings von einem dünnen Wasserfilm bedeckt sein muss.
Verdauungsorgane
Der Verdauungstrakt besteht aus einem langen, von der Vorder- zur Hinterseite des Körpers laufenden Schlauch, der sich in Mundhöhle, Schlund (Pharynx), Speiseröhre, Mitteldarm und Enddarm (Rektum) gliedern lässt. Die ersten drei Abschnitte werden als Vorderdarm (Oesophagus) zusammengefasst, weil ihre aus Cuticula bestehende Auskleidung zusammen mit der cuticulären Hinterdarmauskleidung bei jeder Häutung abgestoßen wird.
Die Mundöffnung befindet sich bei fleisch- oder allesfressenden Arten (Carnivoren und Omnivoren) meistens endständig am Vorderende des Körpers (terminal), bei Arten, die sich von Pflanzen oder organischem Abfall ernähren (Herbivoren und Detritivoren), dagegen oft etwas hinter dem Vorderende auf der Bauchseite (ventral-subterminal). Sie ist vielfach von einem Kranz aus 10 bis 30 harten, quaderförmigen Vorsprüngen, den sogenannten Lamellen (Lamellae) umgeben und sitzt manchmal auf einem Mundkegel, der teleskopartig ausgefahren werden kann.
In die sich anschließende röhrenförmige Mundhöhle mündet links und rechts je eine Speicheldrüse, die nicht nur Verdauungssekrete absondert, sondern auch die für Bärtierchen charakteristischen Stilette synthetisiert. Dieses sind harte, meistens gerade, gelegentlich aber auch gekrümmte Nadeln, die normalerweise im Innenraum (Lumen) der jeweiligen Speicheldrüse liegen, aber mit ihrer scharfen Spitze in die Mundhöhle vorragen können. Durch Pro- und Retraktormuskeln, die an ihrem verdickten Hinterende ansetzen, können sie vorgestreckt oder eingezogen werden. Querverlaufende Stiletthalter verbinden sie mit der Mundhöhle, während ihr Vorderende im eingezogenen Zustand in eigenen Stilettscheiden ruht. Bei carnivoren Arten sind sie meistens etwas kräftiger ausgeprägt als bei herbivoren Formen.
Der Schlund besteht aus radial nach außen laufenden quergestreiften Epithelmuskelzellen, die von der Leibeshöhle durch eine Basallamina abgetrennt sind. Der Schlundinnenraum, das Lumen, ist dagegen von Cuticula ausgekleidet und durch seinen Y-förmigen, triradiaten Querschnitt ideal zum effizienten Erzeugen von Unterdruck geeignet, der dazu genutzt wird, flüssige Nahrung in den Darm einzusaugen. Dieses Pumpenprinzip hat sich unabhängig auch bei anderen, nicht näher verwandten Tieren wie etwa den Rädertierchen entwickelt.
Eine aus würfelförmigen Zellen bestehende kurze Speiseröhre, die vermutlich Schleim sezerniert, stellt die Verbindung zum Mitteldarm her, der aus einschichtigem Epithelgewebe besteht, das manchmal seitliche Ausstülpungen (Mikrovilli) besitzt, die vermutlich dazu dienen, die Oberfläche zu vergrößern. Beim Eintritt in den Darm wird die Nahrung oft durch eine peritrophe Membran eingehüllt, deren Funktion es ist, das empfindliche Darmgewebe vor der Beschädigung durch Fremdkörper zu schützen. Der Mitteldarm sezerniert hydrolytische Enzyme, nimmt Nährstoffe aus der Nahrung auf, speichert diese in Form von Fetten oder Polysacchariden und hat vermutlich auch eine Funktion bei der Ausscheidung von Abfallstoffen. Bei vielen Arten findet sich hier eine umfangreiche Bakterienflora, die vermutlich kommensal, also ohne Beeinflussung ihres Wirts, lebt, teils aber wohl auch als Nahrungsquelle dient.
In einem kurzen, von Cuticula ausgekleideten Enddarm werden die Nahrungsreste gesammelt, möglicherweise auch noch einmal modifiziert und dann durch den bauchseitig, etwas vor dem letzten Beinpaar auf der Mittellinie (medial-ventral) gelegenen Anus an die Außenwelt abgegeben. Bei vielen Arten münden nicht nur die nachfolgend näher beschriebenen Malpighischen Drüsen in den Enddarm ein, sondern auch der Eileiter der Weibchen; man bezeichnet den letzten Darmabschnitt in diesem Fall als Kloake.
Ausscheidungsorgane
Unverdauliche Reste, Abfallprodukte des Stoffwechsels oder Schadstoffe können auf verschiedenem Wege an die Außenwelt abgegeben werden. Zum einen werden von der Epidermis viele Schadstoffe in die darüberliegende Cuticula eingebaut und dann bei der nächsten Häutung zusammen mit dieser abgestoßen. Daneben übernehmen die Speicheldrüsen während der Häutung anscheinend eine Ausscheidungsfunktion. Auch der Darm nimmt nicht nur Nährstoffe auf, sondern gibt wohl auch Schadstoffe aus der Leibeshöhle an das Darmlumen, den Darminnenraum, ab.
Viele Arten, die der Klasse Eutardigrada zugeordnet werden, verfügen daneben noch über drei, sehr selten vier spezialisierte Organe, die man als Malpighische Drüsen bezeichnet. Zwei von ihnen liegen seitlich (lateral), eine liegt rückseitig (dorsal) des Darms. Sie münden an der Verbindungsstelle von Mittel- und Enddarm und dienen sehr wahrscheinlich der Ausscheidung stickstoffhaltiger Abfallstoffe. Obwohl sie strukturell den gleichnamigen Drüsen der Insekten ähneln, handelt es sich wahrscheinlich nicht um homologe Organe, das heißt, sie gehen evolutionsgeschichtlich nicht auf eine gemeinsame Vorgängerstruktur zurück. Da sie bei den als ursprünglich geltenden meereslebenden Arten der zweiten großen Klasse, der Heterotardigrada, nicht vorhanden sind, gelten sie als evolutionäre Anpassung an das Leben im Süßwasser und an Land.
Einige landlebende Formen der Heterotardigrada verfügen an Stelle von Malpighischen Drüsen über spezielle bauchseitig gelegene Organe, die am Ansatz des zweiten und dritten Beinpaares münden und vermutlich eine Ausscheidungsfunktion innehaben. Echte Nephridien, hochentwickelte Ausscheidungsorgane, kommen dagegen bei Bärtierchen grundsätzlich nicht vor.
Nervensystem
Das Nervensystem der Bärtierchen besteht aus einem um den Vorderdarm laufenden Nervenring im Kopf und zwei paarig auf der Bauchseite nach hinten verlaufenden Nervensträngen, die durch Querverbindungen in jedem Rumpfsegment ein sogenanntes Strickleiternervensystem bilden.
Der vordere Nervenring besteht aus einem oberhalb der Mundhöhle gelegenen Oberschlundganglion und einem unterhalb derselben befindlichen Unterschlundganglion, zwei Ansammlungen von Nervenzellen, die durch seitlich des Verdauungstrakts verlaufende Nervenbänder miteinander zu einem Ring verbunden sind und als primitives „Gehirn“ angesehen werden können. Das Oberschlundganglion besteht aus zwei Paaren rückseitig gelegener und zum Hinterende (caudal) ausgerichteter Gehirnlappen, einem inneren und einem äußeren, sowie einem weiteren seitlich der Stilette gelegenen Paar. Das äußere rückseitige Paar innerviert, falls diese vorhanden sind, die Augen und versorgt auch weitere am Kopf befindliche Sinnesorgane, die Cirri und Clavae, mit Nerven. Es ist außerdem in ungewöhnlicher Weise mit den Ganglien des ersten Rumpfsegments verbunden. Diese sind jedoch wie auch bei den verwandten Gliederfüßern in erster Linie durch breite Nervenbänder an das Unterschlundganglion angeschlossen.
Das Bärtierchengehirn entsteht aus der Fusion mehrerer, ursprünglich unabhängiger Ganglien, was als Hinweis auf die Entstehung des Kopfes aus der Verschmelzung mindestens dreier Segmente gewertet werden kann. Auch das Gehirn der eng verwandten Gliederfüßer setzt sich aus mehreren Ganglien zusammen, welche dort die drei Gehirnregionen Proto-, Deuto- und Tritocerebrum bilden. Deswegen wird manchmal vermutet, dass es sich bei den entsprechenden Strukturen um Homologien handelt, also Gewebe, die auf gemeinsame Vorfahren von Bärtierchen und Gliederfüßern zurückgehen. Neuere elektronenmikroskopische Untersuchungen widersprechen dieser Ansicht und kommen stattdessen zu dem Ergebnis, dass das gesamte Gehirn der Bärtierchen evolutionsgeschichtlich dem Protocerebrum der Gliederfüßer entspricht.
Zwei bauchseitig verlaufende Nervenstränge bilden das Rumpfnervensystem. Sie entspringen am Unterschlundganglion und verlaufen parallel zueinander zum Hinterende des Tieres. Jeder Nervenstrang weist vier oder fünf Ganglien auf, die den vier beintragenden Segmenten und vielleicht einem weiteren, beinlosen Genitalsegment entsprechen. Die zwei Ganglien eines Beinsegments sind miteinander durch querlaufende Nervenverbindungen verknüpft.
Sinnesorgane
Viele, aber nicht alle Bärtierchen verfügen über punktförmige, entweder rot oder schwarz gefärbte Augen. Sie sind als sogenannte Pigmentbecherocelli ausgeführt, das heißt, jedes Auge besteht aus einer von zwei Photorezeptorzellen umschlossenen, becherförmigen Pigmentzelle, deren konkave Seite der Körperoberfläche und damit dem Licht zugewandt ist. Sie werden durch die äußeren, rückseitigen Gehirnlappen mit Nerven versorgt und sind in ihrer speziellen Form nur bei Bärtierchen anzutreffen.
Daneben finden sich bei manchen Arten auf den Rumpfsegmenten borstenförmige Sensillen, die vermutlich auf chemische oder Berührungsreize reagieren. Fadenförmige Sensillen, die Cirri, befinden sich besonders bei vielen meereslebenden Arten auf der Kopfrückseite und sind wahrscheinlich Tastsinnesorgane, während Clavae, etwas dickere und von innen hohle Fäden, vermutlich Chemorezeptoren darstellen. Bei vielen Arten finden sich um die Mundöffnung herum angeordnete warzenförmige Erhebungen, die Papillen, die wahrscheinlich ebenfalls eine Funktion bei der Wahrnehmung der Umgebung innehaben.
Fortpflanzungsorgane
Bärtierchen besitzen grundsätzlich nur eine, unpaarig angelegte und von echtem Coelomgewebe umgebene Keimdrüse (Gonade), die oberhalb des Verdauungstrakts gelegen und durch Bänder am Vorderende der rückseitigen Körperwand befestigt ist.
Im Hoden der Männchen werden die begeißelten Spermien gebildet. Von ihm gehen zwei Spermienleiter aus, die sich bauchseitig an der vor dem Anus auf der Körpermittellinie gelegenen, oft röhrenartig vorstehenden Geschlechtsöffnung, der Gonopore, vereinigen und zur Umwelt nach außen öffnen. Die funktionell nicht erklärbare Dopplung der Spermienleiter wird als Hinweis auf den evolutionsgeschichtlichen Verlust einer Keimdrüse gewertet.
Der Eierstock der Weibchen besitzt hingegen nur einen Eileiter, der je nach Art entweder rechts oder links vom Darm liegt. Seine Mündung liegt bei den Arten einer Klasse, den Heterotardigrada, in einer separaten, meistens vor dem Anus gelegenen, Gonopore, bei den Arten der anderen Klasse, den Eutardigrada, dagegen im Hinterdarm, der damit zur Kloake wird. Viele Weibchen besitzen ein bis zwei Samenbläschen, die bei einer Kopulation die Spermien der Männchen aufnehmen und bis zur Eiablage speichern.
Zwittrige Individuen besitzen eine als Ovotestis bezeichnete Keimdrüse, in der sowohl Spermien- als auch Eizellen heranreifen, die durch einen gemeinsamen Ei- bzw. Samenleiter freigesetzt werden können.
Verbreitung
Bärtierchen leben weltweit auf allen Kontinenten einschließlich Antarktika und in allen Ozeanen. Sie finden sich sowohl in mitteleuropäischen Regenrinnen als auch in regelmäßig vereisten arktischen Tümpeln oder tropischen Regenwäldern, in mehr als 6000 Metern Höhe im Himalaja-Gebirge, auf abgelegenen Inseln wie den Südsandwich-Inseln, in der 4690 Meter tief gelegenen abyssalen Zone auf dem Boden des Indischen Ozeans oder mitten im Atlantik auf treibenden Braunalgen. Obwohl sie in allen Klimazonen vorkommen, besteht ein Verbreitungsschwerpunkt in polaren und gemäßigten Breiten.
Erst auf Familien- und Gattungsebene lässt sich eine biogeografische Struktur erkennen, die mit der Trennung des Urkontinents Pangaea in Gondwana und Laurasia in Verbindung gebracht werden kann. Insgesamt zehn Gattungen und 22 Arten sind aber selbst Kosmopoliten, das heißt, auf der ganzen Welt zu finden. Sie gelten als Überbleibsel einer vor der erdgeschichtlichen Epoche der Trias bestehenden Pangaea-Fauna. Die meisten anderen Arten besitzen ein räumlich eingeschränktes Verbreitungsgebiet.
Lebensraum

Nach ihrem Lebensraum (Habitat) lassen sich Bärtierchen grundsätzlich als meeres- (marin), süßwasser- (limnisch) oder landlebend (terrestrisch) beschreiben, wobei die Trennung zwischen den letzten beiden Kategorien nur unscharf ist, weshalb auch der zusammenfassende Begriff limnoterrestrisch häufig Verwendung findet. Alle Bärtierchen sind, obwohl teilweise hochgradig austrocknungsresistent, zum aktiven Leben auf einen dünnen Wasserfilm angewiesen.
Lebensraum Meer
Die marinen Arten leben sowohl in Salz- als auch in Brackwasser und finden sich von der Gezeitenzone hinab über die Flachwasserzone bis in die abyssalen Tiefebenen der Ozeane; mindestens eine Art ist in der Lage, den Wasserdruck zu überstehen, der auf dem Boden des Marianengrabens herrscht. Ist der Meeresgrund schlammig ausgebildet, haben sich die dortigen benthischen Bärtierchen meistens durch einen starken wurmförmigen Körper mit verkürzten Extremitäten an ihre Umgebung angepasst; in sandigem oder gerölligem Untergrund, der Spalten und Ritzen bietet, finden sich dagegen eher Tiere mit kräftig ausgebildeten Stummelbeinen. Eine Art hat sich anscheinend auf Manganknollen als Lebensraum spezialisiert, während andere in ausgedehnten Algenteppichen oder auf Tieren wie Steinkorallen (Scleractinia), Moostierchen (Bryozoa), Muscheln (Bivalvia), Asseln (Isopoda), Rankenfußkrebsen (Cirripedia), Seeigeln (Echinoidea) oder Seewalzen (Holothuroidea) leben, teils kommensal, also ohne Beeinflussung des Wirts, teils aber auch parasitär. Mindestens eine Art lebt auf treibenden Sargassum-Algen mitten auf dem offenen Ozean in der Sargassosee. Zum Land hin findet sich an fast allen Stränden eine ausgeprägte Sandlückenfauna in den obersten Zentimetern des Bodens, wo die Tiere zwischen einzelnen Sandkörnern leben. Sind mehrere Arten vorhanden, verteilen sie sich meistens auf unterschiedliche Mikrolebensräume, die sich durch Feuchtigkeits- oder Temperaturunterschiede voneinander abgrenzen lassen. Den Übergang zu den terrestrischen Formen bilden diejenigen Bärtierchen, die in marinen Flechten leben, welche sich oberhalb des bei Flut gewöhnlich erreichten Wasserspiegels auf Felsgeröll angesiedelt haben und normalerweise nur von der salzigen Gischt erreicht werden.
Lebensraum Süßwasser

Die im eigentlichen Sinne limnischen Arten leben sowohl in Fließgewässern als auch in Seen, Teichen, Tümpeln oder auch einzelnen Pfützen. Innerhalb eines Sees bilden Bärtierchen meistens einen Bestandteil der bodenlebenden Sandlückenfauna; Funde aus bis zu 150 Metern Tiefe sind bekannt. Die Tiere leben meistens in den obersten, sauerstoffhaltigen Zentimetern des Bodens, finden sich zum Seeufer hin, wo lockerer Sand die Tiere ernsthaft verletzen könnte, aber meistens etwas tiefer. Daneben werden auch Algen oder Wasserpflanzen besiedelt, an der Seeoberfläche finden sich Bärtierchen gelegentlich in Seerosen.
Oft lassen sich einzelne Individuen in Regenrinnen aufspüren; die Tiere werden vermutlich von Moosen des Dachs gewaschen, sodass sie als terrestrisch gelten. Schließlich bilden auch heiße Quellen einen von manchen Arten besiedelten Süßwasser-Lebensraum.
Lebensraum Land

Die wichtigsten terrestrischen Habitate sind Moosrasen; die dort lebenden Arten werden als moosliebend oder bryophil bezeichnet. Weil Moose in den Zwischenräumen Wasser speichern, können sie allerdings auch als aquatische Lebensräume angesehen werden. Oft finden sich unterschiedliche Arten in den verschiedenen Zonen des Mooses; die Bärtierchen-Faunen der Rhizoidschicht, mit der die Moose im Boden verwurzelt sind, und der photosynthetisch aktiven, aber austrocknungsgefährdeten Außenschicht sind zum Beispiel nicht identisch. Daneben finden sich die Tiere auch in Flechten oder geeigneten Blütenpflanzen wie etwa Bromelien (Bromeliaceae), Kannenpflanzen (Nepenthaceae), in Steinbrech (Saxifraga), Mannsschild (Androsace) oder Karden (Dipsacus). Bei all diesen Pflanzen sammelt sich etwa in den Blattansätzen oder anderen becherförmigen Pflanzenteilen Wasser; sie werden damit für Bärtierchen zu einem Miniaturlebensraum.
Daneben finden sich Bärtierchen häufig in der Laubstreu von Wäldern oder im Boden selbst, wobei Buchenwälder anscheinend besonders beliebt sind. Ein etwas ausgefalleneres Habitat sind die Gletscher der Hochgebirge: Dort können dunkle Ablagerungen von Staub oder feinkörnigem Geröll tagsüber zur Wärmeabsorption und damit zum vorübergehenden Antauen der Oberfläche führen; in der dabei entstehenden, wässrigen Kryokonit-Schicht lassen sich ebenfalls Bärtierchen finden. Städtische Lebensräume wurden bisher noch kaum untersucht.
Bärtierchen sind nur dann aktiv, wenn sie selbst in ihrem jeweiligen Substrat zumindest von einem dünnen Wasserfilm bedeckt sind, so dass sie in ariden Gebieten wie etwa Wüsten nicht leben können. Viele terrestrische Arten sind allerdings extrem austrocknungsresistent und können daher regelmäßige Trockenperioden und auch Temperaturextreme gut überstehen. Diesen grundlegenden ökologischen Vorteil, den sie mit manchen Rädertierchen (Rotifera) teilen, können sie gegenüber konkurrierenden Tieren wie etwa Fadenwürmern (Nematoda) dort am besten ausspielen, wo der Feuchtigkeitsgehalt der Umgebung starken Schwankungen unterworfen ist, was zumindest teilweise die große Vorliebe vieler Bärtierchenarten für Moose und Flechten erklärt, die in sehr kurzer Zeit austrocknen oder mit Wasser geflutet werden können. Die Besiedlung dieser Lebensräume, zu denen wenige andere Tiere Zugang haben, gilt als ein wichtiger Grund für den großen evolutionären Erfolg der Bärtierchen.
Populationsdichten
Genaue Angaben über Populationsdichten liegen nur für wenige Arten und auch dort bisher nur in Stichproben vor: An Sandstränden fanden sich so pro Kubikzentimeter Sand bis zu 35 Individuen, in Böden wurden bis zu 30 Individuen pro Quadratzentimeter Oberfläche gezählt, während Mooskissen mit etwa 200 Individuen pro Quadratzentimeter erwartungsgemäß einen sehr eng besiedelten Lebensraum darstellen. In der Regel liegen die Populationsdichten jedoch deutlich unter diesem Höchstwert. Soweit es sich aufgrund der bisher noch unzureichenden Datenlage sagen lässt, sind die Individuenzahlen meereslebender Arten meistens wesentlich kleiner als diejenigen der land- oder süßwasserlebenden Formen.
Populationsdichten können durch Temperatur und Feuchtigkeit, das Nahrungsangebot, die Zahl der Parasiten und Fressfeinde oder durch Umweltschadstoffe beeinflusst sein und schwanken bei den limnoterrestrischen Arten meistens jahreszeitbedingt, wobei sich oft im Frühjahr und Frühsommer ein erster und im Herbst ein zweiter Höhepunkt feststellen lässt.
Transport und Fortbewegung
Die Verbreitung von Bärtierchen erfolgt nur in sehr untergeordnetem Maße durch aktive Fortbewegung; die meisten Tiere werden stattdessen durch Wind, Wasser oder Tiere in neue potentielle Lebensräume verbracht. Dieser passive Transport betrifft in erster Linie die Eier der Tiere sowie Zysten und Tönnchen – gegenüber Umweltextremen in größerem oder extremem Maße unabhängige Lebensstadien.
Marine Arten lassen sich in Ozeanströmungen treiben, wobei ihnen vermutlich spezielle segelartige Auswölbungen ihrer Außenhaut helfen. Limnoterrestrische Arten werden auf dieselbe Weise manchmal von über die Ufer tretenden Fließgewässern oder von Schmelzwasser transportiert. Zeitweilig ausgetrocknete Habitate erlauben eine Verbreitung von Eiern oder Cysten mit dem Wind (Anemochorie), während kleinere Strecken in Wasserspritzern überbrückt werden können. Während eines Unwetters vor Grönland konnten sogar ausgewachsene Tiere in fallenden Regentropfen nachgewiesen werden, die wahrscheinlich zuvor vom Sturm aufgewirbelt wurden. Vermutlich bringen auch Insekten oder Vögel, an denen die Eier oder Zysten haften bleiben, Bärtierchen in neue Habitate (Zoochorie).
Zur aktiven Fortbewegung benötigen alle Arten einen dünnen umgebenden Wasserfilm. Sie nutzen dann die Beine der ersten drei Rumpfsegmente, die im Gegensatz zu den verwandten Stummelfüßern nicht nur paarweise, sondern auch einzeln bewegt werden können, um etwa über Sandkörner zu kraxeln oder in Mooskissen herumzuklettern. Die an den Beinen sitzenden Klauen oder Haftscheibchen werden dazu eingesetzt, das jeweilige Substrat zu ergreifen. Anders als die ersten drei Beinpaare dienen die hinteren beiden Gliedmaßen dazu, sich am Untergrund festzuhalten, bei manchen Arten auch zur Rückwärtsbewegung; an diese abgewandelte Funktion sind sie durch die unterschiedliche Klauenausrichtung angepasst. Die größte experimentell gemessene Laufgeschwindigkeit liegt bei 17,7 Zentimetern pro Stunde; der Wert lässt den lateinischen Namen der Gruppe, Tardigrada, der sich aus dem Lateinischen tardus ‚langsam‘ und gradus ‚Schritt‘ ableitet, also „Langsamschreiter“ bedeutet, angebracht erscheinen.
Phototaxis, also die Hin- oder Wegbewegung zu und von Lichtquellen, ist noch sehr unzureichend untersucht. Jungtiere reagieren anscheinend negativ photokinetisch, das heißt, sie reagieren auf Lichteinstrahlung mit schnelleren Bewegungen und spontanen Richtungsänderungen, ohne dass sich eine gezielte Vermeidungsreaktion feststellen ließe. Da Lichteinstrahlung oft mit Wärmestrahlung und nachfolgender Wasserverdunstung verbunden ist, hängt dieses Verhalten vielleicht mit der für Jungtiere bedrohlicheren Austrocknungsgefahr zusammen.
Eine marine Art ist in der Lage, aktiv zu schwimmen; ihre Cuticula ist glockenförmig ausgedehnt und kann durch Kontraktion einen gerichteten Wasserstrahl ausstoßen; die Tiere bewegen sich also wie Quallen nach dem Rückstoßprinzip.
Ernährung

Die meisten Bärtierchen ernähren sich vegetarisch, hauptsächlich von Algenzellen, die sie entweder freilebend oder in Flechten finden. Die pflanzlichen Zellen der Moose gehören dagegen selbst bei den permanent dort lebenden Tieren nur selten zum Nahrungsspektrum. Bodenbewohnende Arten nehmen neben Algen zusätzlich auch organische Abfälle mitsamt den darin enthaltenen Bakterien und Pilzsporen auf; manche Formen leben aber auch ganz oder teilweise räuberisch. Zu ihrem Beutespektrum gehören Protozoen, Rädertierchen (Rotifera) und Fadenwürmer (Nematoda), aber auch andere Bärtierchen. Einige marine Arten leben als Ectoparasiten auf der Haut von Seewalzen oder Rankenfußkrebsen. An Land sind Bärtierchen wegen der beständig existierenden Austrocknungsgefahr an eine parasitische Lebensweise nur schlecht angepasst; eine einzige Art ist hier bekannt, die möglicherweise endoparasitisch in Landlungenschnecken lebt.
Zum Fressen pressen Bärtierchen ihren Mundkegel gegen die betroffene Pflanzenzelle, die Haut ihrer Beute oder die Körperwandung ihres Wirts. Durch Vorschieben der nadelscharfen Stilette werden diese dann angestochen oder durchbohrt und mitsamt dem gesamten Zell- oder Körperinhalt ausgesaugt. Besonders große Arten können dagegen nicht nur flüssige Nahrung aufnehmen, sondern ihre Beute auch als Ganzes einsaugen; davon sind insbesondere kleinere Räder- und Bärtierchen betroffen.
Fressfeinde, Parasiten und Kommensalen
Die wichtigsten Fressfeinde von Bärtierchen sind andere Bärtierchenarten, Rädertierchen (Rotatoria) und Fadenwürmer, daneben auch Milben (Acari), Spinnen (Araneae), Springschwänze (Collembola), verschiedene Insektenlarven, aber auch in unspezifischer Weise „grasende“ Organismen wie Regenwürmer (Lumbricidae) oder diverse Süßwasserkrebse.
Einige Bärtierchen fallen fleischfressenden Pilzen zum Opfer, deren feine Zellfäden (Hyphen) zu Schlingen verflochten sind, in denen sich ihre Beute verfängt, und die daraufhin in die gefangenen Tiere einwachsen. Andere Pilze wie etwa Harposporium, das ungeschlechtliche Stadium der Schlauchpilzgattung Atricordyceps, geben Konidien, asexuelle Sporen, ab, die vermutlich, sobald sie von Bärtierchen gefressen werden, im Darm auskeimen und ihre Opfer von innen verdauen. Möglicherweise penetriert der Pilz aber auch von außen die Cuticula der Tiere. Weitere wichtige Pilzparasiten von Bärtierchen sind die Töpfchenpilzart Sorochytrium milnesiophthora und die Jochpilze Ballocephala sphaerospora und Ballocephala verrucospora.
Vermutlich gibt es eine ganze Reihe von Parasiten unter den Protozoen; näher untersucht wurde bisher aber erst das Wimpertierchen Pyxidium tardigradum, das bevorzugt Bärtierchen befällt. Es lebt vermutlich als Symphoriont, das heißt, es lässt sich von seinem Wirt nur verbreiten, wird ihm aber selbst anscheinend nicht gefährlich.
In marinen Bärtierchen finden sich oft zahlreiche symbiotische Bakterien; auch die landlebenden Formen besitzen meistens eine reiche bakterielle Darmflora. Das Proteobakterium Xanthomonas campestris, ein wichtiger Pflanzenschädling, wird vermutlich von Bärtierchen übertragen.
Häutung
Größenwachstum ist bei Bärtierchen nur durch regelmäßige Häutungen möglich. Dabei wird die nicht-zellige Außenhaut (Cuticula) mitsamt den Stiletten, der Auskleidung des Vorder- und Hinterdarms und den Beinklauen abgestoßen. Die Häutung beginnt immer am Vorderende der Tiere, und zwar damit, dass die alten Stilette und die Cuticula von Mundhöhle und Vorderdarm ausgestoßen werden. Dadurch, dass sich der Körper zeitweilig zusammenzieht, löst er sich von der alten Haut, die dann abgestreift und als leeres Häutungshemd (Exuvium) zurückgelassen wird. Während dieses normalerweise etwa 5 bis 10 Tage dauernden Vorgangs befinden sich die Tiere im stilettlosen Simplexstadium, in dem sie keine Nahrung aufnehmen können. Bereits während der Häutung wird neue Cuticula von der unterliegenden Epidermis gebildet, während die neuen Stilette in den Speicheldrüsen synthetisiert und die Klauen von speziellen Klauendrüsen aufgebaut werden.
Bei einzelnen Individuen konnten bis zu 13 Häutungen im Laufe ihres Lebens nachgewiesen werden; sie dienen, abgesehen davon, dass sie lebenslanges Wachstum erst möglich machen, auch dazu, den Körperinnendruck zu verringern, in der Cuticula gespeicherte Abfallstoffe abzustoßen oder Parasiten zu entfernen. Viele Arten legen ihre Eier in die Häutungshemden ab.
Resistenzstadien
Viele Bärtierchen haben einzigartige Anpassungen entwickelt, um Trockenheitsperioden, Kälteeinbrüche, starke Schwankungen im Salzgehalt des Wassers oder Sauerstoffmangel überstehen zu können. Ein Beitrag zur Robustheit ist ihre Eutelie: Nach der Embryonalentwicklung findet Zellteilung mit ihren empfindlichen Phasen kaum mehr statt (außer in der Keimbahn). Daraus folgt bereits eine sehr hohe Strahlenresistenz (abgesehen von Sterilität),[2] zum Vergleich siehe die Tabelle in →Strahlenschäden. Zudem passen sich einige Arten den Jahreszeiten durch morphologische Umstellungen an, andere können dickwandige Zysten bilden. Die extreme Form der Anpassung ist jedoch die sogenannte Kryptobiose, bei der die Tiere in einen todesnahen Zustand übergehen, in dem sich keinerlei Stoffwechselaktivität mehr registrieren lässt. Alle Resistenzstadien dienen dem Überstehen widriger Umweltbedingungen und verschaffen Bärtierchen dadurch einen evolutionären Wettbewerbsvorteil gegenüber anderen Tiergruppen. Sie spielen daneben auch eine Rolle beim passiven Transport der Tiere in neue potentielle Lebensräume. Das Bärtierchen ist auch das erste Tier, von dem bekannt wurde, dass es im Weltall überleben kann.[3][4]
Cyclomorphose
Als Cyclomorphose bezeichnet man die regelmäßige Änderung der Körperform in Reaktion auf zyklisch auftretende Veränderungen der Umweltbedingungen. Meistens wandeln sich die als Morphe bezeichneten Lebensstadien der Tiere jahreszeitlich bedingt ineinander um. Cyclomorphose ist bis jetzt nur von meereslebenden Bärtierchen aus den Gattungen Halobiotus, Amphibolus und Hypsibius bekannt. Das bisher am besten untersuchte Beispiel findet sich bei der Art Halobiotus crispae, die in der Gezeitenzone Grönlands lebt: Hier lässt sich eine Pseudosimplex genannte Wintermorphe von einer Sommermorphe unterscheiden. Erstere ist in der Lage, Temperaturen nahe dem Gefrierpunkt zu überstehen, und bleibt auch bei niedrigen Temperaturen beweglich, sie ist dafür aber steril; nur die weitaus weniger resistente Sommermorphe ist fortpflanzungsfähig.
Zysten
Besonders in Süßwasser lebende Arten, aber auch solche, die Moose oder Laubstreu besiedeln, sind in der Lage, als Zysten bezeichnete Resistenzstadien zu bilden. Dazu ziehen sich die Tiere auf 20 bis 50 Prozent ihrer ehemaligen Körpergröße zusammen, reduzieren ihren Stoffwechsel und bauen teilweise auch ihre inneren Organe ab. Dieser Vorgang wird von bis zu drei unvollständigen, unmittelbar aufeinanderfolgenden Häutungen begleitet, an deren Ende das Tier von einer mehrwandigen Umhüllung aus Cuticula-Schichten umgeben ist. In diesem Zustand können die Tiere mehr als ein Jahr überleben. Sobald sich die Umweltbedingungen geändert haben, können sich die Tiere innerhalb von 6 bis 48 Stunden wieder aus ihrer Umhüllung befreien.
Zysten bilden sich grundsätzlich nur im Wasser; sie sind bei weitem nicht so widerstandsfähig wie die weiter unten erwähnten Tönnchen und im Gegensatz zu diesen aufgrund ihres Wassergehalts auch nicht hitzeresistent.
Anoxybiose
Als Anoxybiose bezeichnet man die Toleranz gegenüber Sauerstoffmangel. Wenn die Konzentration dieses Atemgases zu niedrig liegt, bricht die Osmoregulation zusammen, die Tiere nehmen Wasser auf und schwellen deutlich an. Der gesamte Stoffwechsel muss nun anaerob stattfinden, giftige Abfallprodukte sammeln sich in der Leibeshöhle an. Die meisten Bärtierchen sind dennoch in der Lage, einen solchen Zustand für 3 bis 5 Tage zu überstehen; es wird sogar berichtet, dass einzelne Individuen aus der Gattung Echiniscoides in der Lage waren, bis zu 6 Monate in einem verschlossenen Reagenzglas mit verwesenden Rankenfußkrebsen zu überleben. Die Rückkehr in den Normalzustand dauert abhängig von der Zeitdauer des anoxybiotischen Stadiums einige Minuten bis wenige Stunden.
Anoxybiose ist besonders in Lebensräumen wichtig, in denen die Sauerstoffkonzentration zeitweilig stark abfallen kann, etwa in großen statischen Wassermassen. Bärtierchen, die in der Gezeitenzone auf Algenteppichen leben, sind bei Ebbe ebenfalls extremem Sauerstoffmangel ausgesetzt und gehörten vielleicht evolutionsgeschichtlich zu den ersten, die diese Anpassung erworben haben.
Osmobiose
Osmobiose ist die Fähigkeit, Schwankungen im Salzgehalt des Wassers zu tolerieren. Bärtierchen sind wie die meisten Tiere nur dann lebensfähig, wenn die Ionenkonzentration des Wassers innerhalb gewisser Grenzen liegt. Dennoch können vor allem solche Arten, die in der Gezeitenzone leben, für einen kurzen Zeitraum erstaunlich große Schwankungen in der Salzkonzentration überleben. Viele Arten bilden in sehr salzigem Wasser die unten näher beschriebenen Tönnchen; dieses ist jedoch ein Sonderfall der unten erläuterten Anhydrobiose und eine direkte Antwort auf die mit dem hohen Salzgehalt der Umgebung einhergehende innere Austrocknung. Tönnchenbildung wird daher formal nicht als eigentlich osmobiotisches Phänomen angesehen.
Kryobiose
Als Kryobiose bezeichnet man die Fähigkeit, niedrige Temperaturen überstehen zu können. Sie ist eine Erscheinungsform der Kryptobiose, die durch eine nahezu totale Beendigung des Stoffwechsels charakterisiert ist. Es tritt ein sogenannter Dormanzzustand ein. Die unten näher beschriebene Bildung von Tönnchen ist dabei möglich, aber nicht zwingend.
Bei einem langsamen Abfall der Temperatur findet eine allmähliche Umstellung des Stoffwechsels statt, weil die meisten Enzyme ihre Aktivität verlieren und stattdessen tieftemperaturaktive Katalysatoren wirksam werden, die den Zucker Trehalose und andere kryoprotektive Substanzen synthetisieren: Diese schützen die empfindlichen Biomembranen und ersetzen das an die lebenswichtigen Moleküle gebundene Wasser. Besonders in der extrazellulären Körperflüssigkeit wird durch bislang unidentifizierte mittelschwere Moleküle, die als Gefrierkeime wirken, ein kontrolliertes Wachstum von Eiskristallen angeregt, die durch Antigefrierproteine bei geringer Größe stabilisiert werden.
Auf diese Weise ist es den Tieren möglich, Temperaturen bis weit unterhalb des Gefrierpunkts zu überstehen. Diese Fähigkeit erlaubt erst eine Besiedelung der Polarregionen und Hochgebirgsgletscher, wo die Umgebungstemperaturen regelmäßig unter den Gefrierpunkt fallen. Die Unterart Echiniscus sigismundi groenlandicus überdauert zum Beispiel 6 bis 8 Monate eingefroren im Wintereis und toleriert in der Übergangszeit regelmäßiges Tauen und Gefrieren im Wechsel der Gezeiten.
Bärtierchen überstanden sogar zehn Tage im freien All. Mit dem Satelliten FOTON-M3 hatte die ESA 2007 mehrere Proben mit Bärtierchen im All dem Vakuum, der Kälte und UV-Strahlung ausgesetzt. Nach ihrer Rückkehr fanden die Wissenschaftler selbst unter denjenigen Bärtierchen Überlebende, die den extremsten Bedingungen ausgesetzt waren.[5]
Weil Bärtierchen bei genügend langsamem Abkühlen Extremtemperaturen von −273 Grad Celsius[6] überstehen können, wurde vereinzelt vermutet, dass sie außerirdische Lebensformen seien. Solche Temperaturen wurden in ihrem natürlichen Lebensraum niemals auch nur annähernd erreicht, und die Fähigkeit könne somit nicht auf der Erde durch natürliche Selektion entstanden sein. Gegen diese Auffassung spricht, dass ein Lebewesen, das auf der Erde Minusgrade überdauert, indem es Wasser in seinem Gewebe durch schützende Substanzen wie Trehalose ersetzt und den Gefrierprozess selbst kontrolliert, ohne weitere evolutionäre Anpassungen auch Temperaturen von −273 Grad Celsius potentiell überdauern kann. Bärtierchen sind multizelluläre Polyextremophile.[7]
Anhydrobiose
Anhydrobiose ist die Fähigkeit, eine Austrocknung des Körpers durch starke Wasserverluste überstehen zu können. Sie findet sich bei fast allen landlebenden Arten und ist mit der Bildung walzenförmiger, unbeweglicher Resistenzstadien, der Tönnchen, verbunden. Weil im anhydrobiotischen Zustand bei Tönnchen kein Stoffwechsel mehr nachweisbar ist, fällt auch die Anhydrobiose unter den Oberbegriff Kryptobiose.
Bei manchen Arten sammeln sich zahlreiche Individuen vor der eigentlichen Tönnchenbildung und bilden ein loses Knäuel. Diese Aggregation wird als Verhaltensanpassung gewertet und schirmt wohl insbesondere die innenliegenden Tiere etwas stärker von Umwelteinflüssen ab, so dass der Austrocknungsprozess bei ihnen etwas langsamer vor sich geht – eine zu schnelle Dehydrierung kann auch bei Bärtierchen zum Tod führen.
Die Tönnchenbildung beginnt mit morphologischen Reaktionen: Die Beine werden eingezogen und die Körperoberfläche insgesamt stark verkleinert. Durch Poren in der Cuticula werden zunehmend Lipide, fettlösliche Substanzen, abgegeben, die unter anderem vor Pilzangriffen schützen sollen. Durch einen Phasenwechsel dieser Lipide wird die Cuticula zu einem spezifischen Zeitpunkt abrupt wasserundurchlässig. Dieses verringert die Wasserverdunstung und erlaubt eine erneute Verlängerung der Vorbereitungszeit auf den anhydrobiotischen Zustand, während deren schützende Verbindungen synthetisiert werden müssen.
Das Hauptproblem bei der Anhydrobiose besteht darin, dass die strukturelle Integrität von wichtigen Makromolekülen wie Proteinen, Phospholipiden oder Nukleinsäuren um jeden Preis erhalten bleiben muss, da ansonsten in den Zellen irreversible Schäden entstehen würden. Die meisten dieser Verbindungen sind von lose angebundenen Wassermolekülen umgeben, deren Verlust unkontrollierte Reaktionen zwischen ihnen auslösen würde. Die von den Bärtierchen im Verlauf ihrer Evolutionsgeschichte gefundene Lösung des Problems besteht darin, das gebundene Wasser während einer Dehydrierung (Austrocknung) durch andere Verbindungen zu ersetzen, die bei einer Rehydrierung (Benetzung durch Wasser) leicht wieder abgebaut werden können. Die wichtigste dieser Verbindungen ist der Zucker Trehalose, der während der Vorbereitung auf den anhydrobiotischen Zustand in großen Mengen produziert und manchmal um den Faktor 23 gegenüber dem Ausgangszustand angereichert wird. Er schützt nicht nur die Biomembranen und unterbindet Reaktionen zwischen den entwässerten Proteinen und anderen Zellbestandteilen wie Kohlenhydraten, sondern verhindert auch unkontrollierte Oxidationen, die ebenfalls wichtige Makromoleküle zerstören könnten. Neue Forschungsergebnisse deuten darauf hin, dass Bärtierchen einen neuen Typ ungeordneter Proteine besitzen, die bei Austrocknung vermehrt produziert werden und vitrifizieren, das heißt ein glasähnliches, schützendes Material bilden.[8] Neben dieser Proteinfamilie und Trehalose spielt vermutlich auch Glycerin eine Rolle bei diesen Vorgängen. Lipidtröpfchen fällt unterdessen die Aufgabe zu, Körpergewebe voneinander getrennt zu halten, die im normalen Zustand nicht Gefahr laufen, sich zu nahe zu kommen, im Tönnchenzustand jedoch in Kontakt geraten und unerwünschte Vernetzungsreaktionen auslösen könnten. Bis die Tiere aus dem Normalzustand ein stabiles Tönnchenstadium erreicht haben, dauert es etwa 5 bis 7 Stunden.
Sobald die Tiere den anhydrobiotischen Zustand erreicht haben, ist zumindest in trockener Umgebung keinerlei Stoffwechselaktivität mehr nachweisbar; insbesondere fällt der Sauerstoffverbrauch auf Null. In diesem Stadium können die Tiere extreme Austrocknung, das Einbringen in Salzlake, Äther, reines Ethanol oder flüssiges Helium, Temperaturen zwischen −196 °Celsius und +90 °Celsius[9], großen hydrostatischen Druck[10] und Vakuum überstehen.
Sobald wieder genügend Wasser vorhanden ist, findet die Rücktransformation in den Normalzustand, die Restitution, statt. Dazu wird die Trehalose aerob, also unter Nutzung von Luftsauerstoff, abgebaut und wieder durch Wasser ersetzt. Die Erholungszeit ist abhängig von der Temperatur, dem Sauerstoffgehalt und dem pH-Wert der Umgebung sowie von der im kryptobiotischen Zustand verbrachten Zeit und schwankt darüber hinaus auch von Art zu Art etwas. In den meisten Fällen liegt sie zwischen zehn Minuten und mehreren Stunden, bei sehr lange andauerndem Tönnchenzustand auch bei einigen Tagen.
Ob die Rückkehr in ein aktives Lebensstadium erfolgreich ist, hängt in erster Linie vom korrekten Ablauf der Tönnchenbildung und vom Ernährungszustand der Tiere ab. Besonders wichtig sind in diesem Zusammenhang die Lipidreserven, die nicht nur wie oben erwähnt selbst eine Rolle im kryptobiotischen Zustand spielen, sondern auch in Glycerin und Trehalose umgewandelt werden können und darüber hinaus auch Energie für den Restitutionsvorgang bereitstellen. Solange die Tiere die Möglichkeit haben, diese Reserven in regelmäßigen Abständen aufzufrischen und darüber hinaus jede Austrocknung nur langsam stattfindet, können sie in regelmäßigen Abständen aus dem aktiven in den anhydrobiotischen Zustand übergehen und umgekehrt. Individuen aus der Gattung Echiniscoides, die in der Gezeitenzone leben, können zum Beispiel experimentell in einen Zyklus mit sechsstündlicher Periode versetzt werden.
Bärtierchen im kryptobiotischen Zustand werfen Fragen nach der Definition von Leben auf: So werden als charakteristische Merkmale eines lebenden Organismus häufig Stoffwechsel, Wachstum und Fortpflanzung angeführt. Keine dieser Eigenschaften findet sich jedoch im Tönnchenstadium, so dass man die Rückkehr in den aktiven Zustand mit den Worten von Lazzaro Spallanzani, einem italienischen Naturforscher des 18. Jahrhunderts, als „Wiederauferstehung von den Toten“ bezeichnen könnte. Andererseits kehren zwar viele, aber nicht alle Bärtierchen aus dem anhydrobiotischen Stadium wieder in einen aktiven Zustand zurück, was den Tardigradologen John H. Crowe 1975 zu der Frage veranlasste:
- Does this then mean that they "died" while they were "dead"?
- Bedeutet das dann, dass sie „gestorben“ sind, während sie „tot“ waren?
Nach Crowes Auffassung kann die Definition von Leben nicht auf einzelne Merkmale reduziert werden; er sieht stattdessen das Fortbestehen der strukturellen Kontinuität und Integrität eines lebenden Systems als im Wortsinne „lebensnotwendig“ an.
Fortpflanzung und Lebenszyklus
Bärtierchen können sich sowohl ungeschlechtlich als auch auf geschlechtliche Weise vermehren. Die meisten Arten pflanzen sich aber ausschließlich sexuell fort.
Asexuelle Fortpflanzung
Asexuelle Fortpflanzung ist nur durch Parthenogenese möglich, also die Reifung unbefruchteter Eier, die sich anschließend zu Weibchen entwickeln. Sie kommt bei einer Reihe von Arten, unter anderem aus den Gattungen Echiniscus und Pseudoechiniscus vor. Männliche Tiere sind oft nicht bekannt, auch wenn in manchen Fällen nachträglich Zwergmännchen beschrieben werden konnten. Bei diesen und anderen Arten lässt sich ein Nebeneinander von parthenogenetischer und sexueller Fortpflanzung feststellen; die Männchen sind dabei grundsätzlich in der Lage, geeignete, sexuell aktive, von ungeeigneten, sich parthenogenetisch reproduzierenden Weibchen zu unterscheiden. Ein wichtiger Vorteil, der mit Parthenogenese einhergeht, ist der fehlende Aufwand für die Partnersuche, ein gravierender Nachteil die Verringerung der genetischen Vielfalt. Die negativen Folgen davon werden allerdings teilweise durch die Fähigkeit, widrigen Umwelteinflüssen durch Kryptobiose einfach auszuweichen, kompensiert.
Sexuelle Fortpflanzung
Sexuelle Fortpflanzung bedingt bei Bärtierchen meistens, allerdings nicht immer, die Existenz zweier getrennter Geschlechter. Einige hermaphroditische (zwittrige) Arten sind bekannt, die zur Selbstbefruchtung in der Lage sind; Ei- und Samenzellen reifen bei ihnen in derselben Keimdrüse heran. Auch sie haben den Vorteil, keine Energie auf die Partnersuche aufwenden zu müssen, unterscheiden sich aber von parthenogenetischen Individuen, die sich letztlich klonen, durch die Möglichkeit genetischer Rekombination.
Für die überwiegende Mehrzahl der Arten existieren hingegen getrennte Geschlechter, die sich allerdings äußerlich nicht immer leicht unterscheiden lassen. Wo detaillierte Untersuchungen zur Populationsstruktur vorliegen, lässt sich fast immer ein deutliches Übermaß an Weibchen konstatieren.
Die Befruchtung kann sowohl außer- als auch innerhalb des Körpers der Weibchen stattfinden, die Eiablage ist für die Weibchen meistens mit einer Häutung verbunden. Die Einzelheiten beider Vorgänge hängen unter anderem vom Lebensraum der Tiere ab.
Bei marinen Arten werden die Spermien der Männchen meistens in den Samenbläschen der Weibchen deponiert, die ihre Eier dann frei ablegen und am Substrat wie zum Beispiel Algenzellen anheften. Daneben kommt es aber auch vor, dass das Weibchen seine unbefruchteten Eier unmittelbar nach der Häutung in die abgestoßene Cuticula legt, wo sie dann extern von den Männchen befruchtet werden.
Bei limnoterrestrischen Arten werden die Spermien gelegentlich noch vor oder während der stattfindenden Häutung des Weibchens in den entstehenden Spalt zwischen der alten und neuen Haut injiziert; sobald das Weibchen seine Eier in das abgelegte Häutungshemd legt, findet die Befruchtung statt. Besonders bei landlebenden Arten kommt es häufig zur Kopulation, wozu sich das Männchen an einem Weibchen festklammert; eine solche Verbindung wird auch bei äußeren Störungen des Geschlechtsakts nicht aufgegeben. Das Männchen führt nun sein Sperma in den Geschlechtstrakt des Weibchens ein und zwar, bevor dieses seine Häutung abgeschlossen hat. Alternativ können die Spermien auch in Samenbläschen deponiert oder auch direkt durch die Außenhaut hindurch in die Leibeshöhle injiziert werden; in letzterem Fall findet die Befruchtung in der weiblichen Keimdrüse statt.
Die Zahl der abgelegten Eier schwankt je nach Art zwischen 1 und 35, wobei sich mit zunehmendem Lebensalter eine stetige Zunahme dieser Zahl beobachten lässt. Über die gesamte Lebenszeit gesehen können einzelne Weibchen über 100 Eier produzieren. Bei meereslebenden Arten fallen sie je nach Umgebungsbedingungen dünn- oder dickschalig aus, während sich bei landlebenden Arten unterschiedliche Eiformen beobachten lassen: Glatte Eier werden meistens in den Häutungshemden der Weibchen deponiert, während Eier mit dicker, aufwendig ornamentierter und vermutlich austrocknungsresistenter Schale frei an Moospflanzen oder Rindenstückchen angeklebt werden. Einige süßwasserlebende Arten nutzen die abgestoßenen Exoskelette von Insekten oder anderen Gliederfüßern als Ablageort.
Besondere Verhaltensanpassungen bei der Fortpflanzung finden sich nur in seltenen Fällen: So wurde bei einigen Arten ein primitives „Paarungsritual“ beobachtet. Das Männchen streichelt dazu sein Weibchen mit den am Kopf befindlichen Cirri. Das Weibchen legt, in dieser Weise stimuliert, nach einiger Zeit seine Eier auf einem Sandkorn ab, auf dem das Männchen dann sein Sperma deponiert. Die Tatsache, dass manche Weibchen die Häutungshemden, in denen sie ihre Eier abgelegt haben, für eine Weile mit sich schleppen, wird gelegentlich als einfacher Fall von „Brutpflege“ interpretiert.
Bei günstigen Bedingungen schlüpfen die Jungtiere nach etwa 5 bis 40 Tagen. Sie absorbieren dazu aus der Umgebung solange Flüssigkeit, bis ihr sich ausdehnender Körper die Eischale sprengt. Manche Arten setzen auch ihre Mundstilette zum Aufbrechen der Eihülle ein. Der gesamte Vorgang dauert meistens nur wenige Minuten.
Die Jungtiere sind meistens ungefärbt und besitzen weniger Borsten, Cirri oder Klauen als die erwachsenen Tiere. Ansonsten sind sie diesen aber schon sehr ähnlich, so dass ihre Entwicklung direkt, also ohne Larvenstadium verläuft. Bei einigen Arten aus der Klasse Heterotardigrada bilden sich der Anus und die Geschlechtsöffnung erst nach ein beziehungsweise zwei Häutungen; dies wird gelegentlich als Hinweis auf eine indirekte Entwicklung gedeutet. In den meisten Fällen wachsen die Jungtiere nur durch eine Vergrößerung des individuellen Zellvolumens, nicht aber durch eine Vermehrung der Zellenanzahl, so dass viele Gewebe bereits nach dem Schlüpfen die endgültige Zellenzahl der Erwachsenenform aufweisen. Allerdings lassen sich auch in späteren Stadien gelegentlich noch Mitosen (Zellteilungen) nachweisen, die vermutlich dem Zweck dienen, abgestorbene Zellen zu ersetzen. Die Geschlechtsreife wird in jedem Fall erst nach mehreren Häutungen erreicht.
Die normale Lebensdauer von Bärtierchen liegt zwischen drei Monaten und zweieinhalb Jahren; sie entspricht der tatsächlichen Lebenszeit der meisten marinen Arten. Bei den limnoterrestrischen Arten wird das Leben der Tiere jedoch manchmal oder oft durch kryptobiotische Zustände unterbrochen, während deren die betroffenen Individuen nicht altern. Mooslebende Arten erreichen auf diese Weise häufig eine reale Lebensdauer von vier Jahren oder mehr; in Einzelfällen können sie auch Jahrzehnte überdauern. In ausgetrocknetem Moos eines botanischen Museums wurde ein Bärtierchen entdeckt, das nach 120 Jahren im anhydrobiotischen Zustand „wiederbelebt“ werden konnte.
Bärtierchen und der Mensch
Bärtierchen finden sich zwar in fast allen menschlichen Lebensräumen, fallen aber wegen ihrer geringen Größe und ausgefallenen Lebensweise kaum auf. Da sie zudem keine unmittelbare wirtschaftliche, medizinische oder tiermedizinische Bedeutung besitzen, sind sie den meisten Menschen unbekannt. Jene, die sie das erste Mal sehen, beschreiben sie oft als „süß“, ein Adjektiv, das sich selbst in seriösen zoologischen Publikationen finden lässt und wohl nicht nur durch die bärenähnliche Körperform, sondern auch durch die tapsige Fortbewegungsweise der Tiere inspiriert ist. Eine Haltung in monoxenischer Kultur, also zusammen mit mindestens einer weiteren Art, ist möglich.
Angaben zur Gefährdung liegen nicht vor; eine Art, Thermozodium esakii, ist allerdings möglicherweise ausgestorben. Da Bärtierchen sehr empfindlich auf Umweltgifte, insbesondere Schwefeltrioxid, reagieren, gibt es Ideen, die Tiere als Indikatoren für die Umweltqualität eines Standorts einzusetzen. Untersuchungen zur Schädigung von Tardigrada durch Schwermetallbelastung von Moosen wurden von ungarischen Forschern vorgelegt.[11] Untersuchungen zu städtischen Lebensräumen liegen aber noch nicht vor.
Stammesgeschichte
Moderne Formen
| Vertreter verwandter Taxa |
|---|
 Vierfleck-Kreuzspinne |
.jpg.webp) Vierfleck (Libelle) |

Die engsten Verwandten der Bärtierchen finden sich in zwei sehr unterschiedlichen Gruppen: Die Gliederfüßer (Arthropoda), die unter anderem Krebstiere (Crustacea), Spinnentiere (Arachnida), Tausendfüßer (Myriapoda) und Insekten (Insecta) umfassen, sind der umfangreichste Tierstamm überhaupt, während die Stummelfüßer (Onychophora), die sich als Würmer mit Beinen beschreiben lassen, ein eher obskures Taxon bilden. Bärtierchen, Glieder- und Stummelfüßer bilden sehr wahrscheinlich zusammen eine natürliche Verwandtschaftsgruppe, ein sogenanntes monophyletisches Taxon, das als Panarthropoda bezeichnet wird. Als gemeinsames abgeleitetes Merkmal kann die bei allen Tieren dieser Gruppe auftretende Segmentierung des Körpers sowie das Auftreten paariger Körperanhänge angesehen werden; auch der Aufbau der Cuticula wird von allen drei Taxa geteilt.
Innerhalb der Panarthropoda werden Bärtierchen traditionell mit den Stummelfüßern als Protoarthropoda zusammengefasst:
| Panarthropoda |
| ||||||||||||
Dabei spielte ursprünglich die Vorstellung eine Rolle, dass Stummelfüßer und Bärtierchen noch nicht die volle Organisationshöhe der Gliederfüßer erreicht haben. Evolutionstheoretisch gilt die Unterscheidung mehr oder weniger hoch entwickelter Formen jedoch mittlerweile als veraltet; moderne Klassifikationen sollen ausschließlich die tatsächlichen stammesgeschichtlichen Beziehungen der Taxa zueinander wiedergeben.
Als gemeinsame Merkmale der Protoarthropoda werden dann die sowohl bei Stummelfüßern als auch bei Bärtierchen zu findenden sackartigen Körperanhänge, die Stummelbeine, angeführt. Daneben gibt es eine Reihe weiterer Homologien, also Merkmale, die sich auf eine gemeinsame Vorläuferstruktur zurückführen lassen. Insbesondere entspricht der vermutlich aus drei Segmenten bestehende Bärtierchen-Kopf sehr wahrscheinlich den ersten drei Einheiten des Stummelfüßer-Körpers. Ihre Stilette können dementsprechend als stark abgewandelte Körperanhänge des zweiten Segments angesehen werden und wären somit den Kiefern der Stummelfüßer homolog. Auch die Klauenstruktur ist bei Bärtierchen und Stummelfüßern sehr ähnlich. Trotz dieser ins Auge springenden Gemeinsamkeiten ist es umstritten, ob die Protoarthropoda eine natürliche Verwandtschaftsgruppe bilden, da vermutlich alle angeführten Merkmale Symplesiomorphien sind, das heißt, schon bei den Vorfahren aller Panarthropoda zu finden waren. Zu Beginn des 21. Jahrhunderts hat sich daher stattdessen eine Präferenz für eine Schwestergruppenbeziehung zwischen Bärtierchen und Gliederfüßern herauskristallisiert:
| Panarthropoda |
| ||||||||||||
Die weitere Verwandtschaft der Bärtierchen wird traditionell in den Ringelwürmern (Annelida) gesehen. Mit ihnen teilen sie unter anderem weiche, nicht gelenkige Körperanhänge mit abschließenden verhärteten „Klauen“ und eine terminale, den Körper nach vorne abschließende Mundstellung. Diese auf den französischen Naturforscher Georges Cuvier zurückgehende Articulata-Hypothese hat auch zu Anbeginn des 21. Jahrhunderts noch viele Anhänger, wurde aber gegen Ende der 1990er Jahre aufgrund morphologischer und molekulargenetischer Erkenntnisse zunehmend in Frage gestellt. An ihre Stelle trat ein Konzept, das die nächsten Verwandten der Panarthropoda in einer Gruppe wurmartiger Tiere, der Cycloneuralia, sieht: Zu diesem Taxon zählt man Faden- (Nematoda) und Saitenwürmer (Nematomorpha), aber auch drei eher obskure Tiergruppen, Priapswürmer (Priapulida), Hakenrüssler (Kinorhyncha) und Korsetttierchen (Loricifera). Alle diese Taxa zeichnen sich wie auch die Panarthropoda dadurch aus, dass sie ihre nicht-zellige Außenhaut oder Cuticula zumindest während einzelner Stadien ihres Lebenszyklus abstoßen; sie werden daher als Häutungstiere (Ecdysozoa) bezeichnet:
| Häutungstiere (Ecdysozoa) |
| ||||||
Ausgestorbene Formen
Bärtierchen-Fossilien sind ausgesprochen rar und tragen daher nur wenig zum Verständnis der Entwicklung dieser Tiergruppe bei. Neben einigen Funden aus dem frühen Erdaltertum sind nur einige in Bernstein erhaltene Individuen aus dem späten Erdmittelalter bekannt.
Als mögliche Stammlinienvertreter der Tactopoda, des Taxons aus Gliederfüßern und Bärtierchen, werden zunehmend die Lobopoden gesehen, eine Gruppe wurmähnlicher Tiere aus den erdgeschichtlichen Epochen des Kambriums und Ordoviziums, die ihren Ursprung allerdings wohl schon im vorhergehenden Ediacarium hatte. Sie liefen ähnlich wie die Stummelfüßer auf nicht-gelenkigen, sackartigen Stummelbeinen und werden daher traditionell diesem Tierstamm zugeordnet. Einige moderne kladistische Untersuchungen halten diese Einschätzung für unbegründet und sehen die Lobopoden stattdessen als nicht-natürliche Verwandtschaftsgruppe, aus der sowohl Stummelfüßer als auch die Tactopoda hervorgegangen sind. Eine vorgeschlagene Variante, die diese Alternativsicht zum Ausdruck bringt, ist in dem folgenden Diagramm dargestellt:
| Panarthropoda |
| |||||||||||||||||||||||||||||||||||||||
Die Gattung Aysheaia, die noch sehr viele ursprüngliche Merkmale wie eine einfache Reihung unspezialisierter langer Beine oder eine feine, nicht mit der unauffälligen Körpersegmentierung übereinstimmende oberflächliche Ringung aufweist, bildet demnach die evolutionäre Schwestergruppe aller anderen Panarthropoda, die sich sodann in die Stummelfüßer und alle weiteren Gruppen aufteilen. Eine Reihe von Lobopoden wie die schwer gepanzerten Gattungen Cardiodictyon, Hallucigenia oder Paucipodia bildet vermutlich einen ausgestorbenen Seitenast, welcher der nicht zugeordneten Gattung Luolishania einerseits und einem unbenannten Taxon aus der Art Kerygmachela kierkegaardi und den Tactopoda andererseits gegenübersteht.
Kerygmachela kierkegaardi, nach dieser Hypothese die unmittelbare Schwestergruppe der Tactopoda, ist aus dem frühen Kambrium Nordgrönland bekannt und ähnlich wie Bärtierchen und Gliederfüßer auch äußerlich deutlich segmentiert. Ob auch die mysteriösen Anomalocaris-Fossilien in die Stammlinie der Tactopoda gehören oder wie oben dargestellt näher mit den Gliederfüßern als mit den Bärtierchen verwandt sind, ist unklar.
Die ersten eindeutig den Bärtierchen zuzuordnenden Fossilien entstammen der sibirischen Kuonamka-Formation. In 530 Millionen Jahre altem Kalkstein aus dem mittleren Kambrium haben sich dort vier Individuen erhalten, die nach Körperform und -größe als Bärtierchen identifizierbar sind. Die noch unbeschriebene Art verfügte anscheinend über drei, vielleicht auch vier Beinpaare, die in einem Paar ungleicher Klauen abschlossen und besaß eine Cuticula-Struktur, die jener der heutigen Tiere schon sehr ähnlich war. Die bereits sehr stark spezialisierten Tiere können womöglich sogar einer der modernen Klassen, den Heterotardigrada, zugeordnet werden.[12]
Wie sich der Übergang von den robusten Lobopoden zu den nur submillimetergroßen Bärtierchen vollzog, ist unbekannt. Ein möglicher Mechanismus wäre Progenese, ein Vorgang, bei dem sich die Keimdrüsen in der Embryonalentwicklung vorzeitig ausbilden und die Geschlechtsreife daher im Vergleich mit dem Ausgangszustand früher eintritt. Diese arrestiert nun die weitere Entwicklung und Differenzierung des Körpers, so dass Larven- oder Jungtiermerkmale wie eine wesentlich geringe Größe beim erwachsenen Tier auftreten, ein Phänomen, dass man als Pädomorphose bezeichnet.
Auch wann Bärtierchen erstmals terrestrische Lebensräume erobert haben, lässt sich mangels Fossilfunden nicht mit Gewissheit sagen. Da Bärtierchen heute häufig in Mooskissen zu finden sind, Moose sehr wahrscheinlich die ersten Pflanzen waren, die das Land besiedelten und dabei hinsichtlich der Austrocknungsgefahr ähnlichen Herausforderungen ausgesetzt waren wie die Bärtierchen selbst, ist es sehr gut möglich, dass Moose und Bärtierchen den Schritt an Land zusammen mit den dazu notwendigen Anpassungen gemeinsam vollzogen haben.
Spätestens zu diesem Zeitpunkt muss sich bei den Tieren die einzigartige Anpassung an Trockenheitsperioden, die Kryptobiose, herausgebildet haben. Sie erlaubte ihnen, widrigen Umweltbedingungen durch vorübergehendes Abschalten des Stoffwechsels einfach auszuweichen. Dieses hat zur Folge, dass zumindest die umweltbedingte Selektion nur verhältnismäßig schwach ist, und erklärt so die sehr langsame, bradytelische Evolutionsrate der körperlichen Erscheinungsform, des Phänotyps.
Es überrascht daher nicht, dass die späteren, aus der Kreidezeit erhaltenen Bärtierchen-Fossilien gegenüber den heutigen Arten kaum eine Änderung der Körperform erkennen lassen. Beorn leggi etwa, eine aus kanadischem Bernstein erhaltene Art, lässt sich bereits der Klasse Eutardigrada zuordnen; ein nur sehr schlecht erhaltenes unbenanntes Jungtier vom selben Fundort gehört möglicherweise in die Klasse Heterotardigrada. Aus dem US-amerikanischen Bundesstaat New Jersey sind weitere Bernsteinfossilien bekannt, die vermutlich aus der Turonian genannten Epoche der späten Kreidezeit stammen. Auch sie sind vermutlich bereits den Eutardigrada zuzuordnen und lassen keine weitergehenden Schlüsse zu, als dass diese Entwicklungslinie schon seit mehr als 65 Millionen Jahren existiert.
Die einzigen weiteren Bärtierchenfossilien sind etwas mehr als 7000 Jahre alte Eier, die sich in subantarktischen Torfmooren erhalten haben. Da verschiedene Bärtierchenarten unterschiedliche Temperatur- und Feuchtigkeitsvorlieben haben, gibt es Überlegungen, die in den verschiedenen Moorschichten auftretenden Eier ähnlich wie Pflanzenpollen zur Bestimmung des damaligen Klimas heranzuziehen.
Systematik
Es besteht kein ernsthafter Zweifel daran, dass Bärtierchen ein monophyletisches Taxon bilden, also auf eine gemeinsame Stammart zurückgehen und alle Nachfahren dieser Art umfassen. Wichtige Synapomorphien, gemeinsame, abgeleitete Merkmale, sind etwa die teleskopartig einziehbaren Beine und die Mundstilette.
Bis zum Jahre 2005 wurden etwa 930 Arten beschrieben, darunter 160 marine Formen. Die tatsächliche Artenzahl ist naturgemäß unbekannt, wird aber auf etwa 10.000 geschätzt. Hinzu kommt, dass sich vermutlich hinter vielen nach morphologischen Kriterien abgegrenzten „Arten“ stattdessen Gruppen kryptischer Arten verbergen, die nur molekulargenetisch auseinanderzuhalten sind, was die Biodiversität des Taxons nochmals erhöhen würde.
Man unterscheidet drei verschiedene Klassen, deren Verwandtschaftsverhältnisse zueinander noch unklar sind:
- Als Heterotardigrada bezeichnet man die „gepanzerten“ Bärtierchen, auch wenn nicht alle Arten tatsächlich eine rückseitig verhärtete und in einzelne Panzerplatten (Skleriten) geteilte Cuticula besitzen. Bei vielen Formen lassen sich auffällige Kopfanhänge wie Cirri und Clavae beobachten; die Beine können sowohl in Klauen als auch in Haftscheiben enden. Die Geschlechtsöffnung liegt immer direkt auf der Körperoberfläche, Malpighische Drüsen zur Ausscheidung und Osmoregulation treten nicht auf. Heterotardigrada finden sich sowohl in marinen als auch in limnoterrestrischen Lebensräumen.
- Als Eutardigrada bezeichnet man die „nackten“ Bärtierchen, ihre Außenhaut ist dünn und nicht verhärtet. Auffällige Sinneshärchen am Kopf finden sich in dieser Gruppe nie; die Beine enden grundsätzlich in Klauen. Anders als bei den Heterotardigrada mündet der Eileiter in den Enddarm ein, der dadurch zur Kloake wird; zur Ausscheidung dienen die spezialisierten Malpighischen Drüsen. Die meisten Eutardigrada leben im Süßwasser oder an Land, obwohl auch einige marine Arten existieren.
- Die Mesotardigrada sind nur durch eine einzige, verschollene Art, Thermozodium esakii bekannt, die formell in eine Familie Thermozodiidae gestellt wird. Der Beschreibung nach besitzt sie am Kopf je einen seitlichen Cirrus, aber keine Clavae; der Mund ist von vier warzenähnlichen Vorsprüngen (Papillen) umgeben. Auch am Beinansatz befinden sich demnach Papillen, während das Beinende in 6 bis 10 einfache Klauen übergeht; Malpighische Drüsen sind vorhanden. Die Art wurde auf Algenpolstern in einer nahe der japanischen Stadt Nagasaki gelegenen heißen Quelle, der Typ-Lokalität, gefunden, die jedoch nach dem Zweiten Weltkrieg durch ein Erdbeben zerstört wurde. Da sich auch kein Typmaterial erhalten hat und die Art bis heute nicht mehr wieder aufgefunden werden konnte, gilt ihre Existenz heute als zweifelhaft und ihr Name entsprechend als Nomen dubium.
Oft wird angenommen, dass eine Ordnung der Heterotardigrada, die mehrheitlich marinen Arthrotardigrada, die ursprünglichsten Bärtierchenarten umfasst, aus denen sich dann zunächst die Formen der anderen Heterotardigrada-Ordnung Echiniscoidea und dort insbesondere der limnoterrestrischen Familie Echiniscidae entwickelt haben, bevor diese wiederum die andere Bärtierchenklasse Eutardigrada hervorbrachte, deren Arten sich hauptsächlich im Süßwasser und an Land finden:
| Bärtierchen |
| ||||||||||||||||||||||||
Diese Hypothese konnte jedoch bislang nicht bestätigt werden; vorläufige molekulargenetische Daten sprechen dafür, dass sowohl Hetero- als auch Eutardigrada natürliche Verwandtschaftsgruppen bilden:
| Bärtierchen |
| ||||||||||||
Forschungsgeschichte
Die Tardigradologie oder Bärtierchenforschung reicht in ihren Anfängen bis ins 18. Jahrhundert zurück. Der deutsche Naturforscher Johann Conrad Eichhorn war vermutlich am 10. Juni 1767 der erste Mensch, der die Tiere beobachtete. Da er seine Entdeckung in seinem Werk Beyträge zur Naturgeschichte der kleinsten Wasserthierchen die mit blossem Auge nicht gesehen werden und die sich in den Gewässern in und um Danzig befinden erst 1775 und damit Jahre später veröffentlichte, gilt aber heute zumeist der Quedlinburger Pastor Johann August Ephraim Goeze als Entdecker der Gruppe. Er konnte nach eigenen Angaben seine erste Beobachtung am 10. Dezember 1772 machen und veröffentlichte seine Beschreibung der Tiere in einem selbst verfassten Anhang der von ihm aus dem Französischen übersetzten und im Jahre 1773 erschienenen Schrift Herrn Karl Bonnets Abhandlungen aus der Insektologie. Dort schrieb er unter anderem:
„Seltsam ist dieses Thierchen, weil der ganze Bau seines Körpers außerordentlich und seltsam ist, und weil es in seiner äusserlichen Gestalt, dem ersten Anblicke nach, die größte Aehnlichkeit mit einem Bäre im Kleinen hat. Dies hat mich auch bewogen ihm den Namen des kleinen Wasserbärs zu geben. […] Man fürchte sich indessen nicht, auch diese Raubthiere der unsichtbaren Welt zu betrachten.“[13]
Bereits ein Jahr später wurde erstmals die Rückkehr aus dem anhydrobiotischen Zustand beobachtet, die der italienische Naturforscher Lazzaro Spallanzani kurz darauf als „Wiederauferstehung von den Toten“ beschrieb. Er war es auch, der im zweiten Band seines 1776 in Modena erschienenen Buchs Opuscoli di Fisica animale e vegetabile der Gruppe den Namen Il Tardigrada gab. 1790 wurden die Tiere in das Werk Systema Naturae des schwedischen Naturforschers und Systematikers Carl von Linné aufgenommen.
Die erste wissenschaftliche Monographie erschien im Jahre 1840, neun Jahre später wurde das erste marine Bärtierchen entdeckt; die wissenschaftliche Beschreibung der ersten fossilen Art, Beorn leggi, musste hingegen bis 1964 noch mehr als ein Jahrhundert warten.
Die systematische Stellung der Tiere war von Anbeginn unklar; während des gesamten 19. Jahrhunderts wurden sie taxonomisch wahlweise mit den Rädertierchen (Rotifera) oder Asselspinnen (Pycnogonida) gruppiert oder zu den Gliederfüßern (Arthropoda) gestellt. Auch in der ersten Hälfte des 20. Jahrhunderts blieben ihre genauen stammesgeschichtlichen Beziehungen umstritten, so dass schließlich der italienische Tardigradologe Giuseppe Ramazzotti die Gruppe in den Rang eines eigenen Tierstamms erhob. Die weitere Klassifikation des Taxons geht auf den deutschen Biologen Ernst Marcus zurück, der 1929 alle Bärtierchen in die Untergruppen der Hetero- und Eutardigrada einteilte; 1937 wurden durch Gilbert Rahm die Mesotardigrada als dritte Gruppe eingeführt.
Diverses
- Durch die missglückte Landung des israelischen Mondlandegerätes Beresheet wurden 2019 einige Tausend Bärtierchen verstreut. Da sie ohne Sauerstoff auskommen und sich nach dem Aufwärmen aus dem tiefgekühlten Zustand wieder zum Leben erwecken lassen,[14] wurden sie für diese Mondmission ausgewählt. „Wie totes Material können sie jahrelang im ausgetrockneten Zustand überdauern. Auch den Absturz auf dem Mond dürften sie überstanden haben“, versicherte Nova Spivack dem US-Magazin Wired. Nova Spivack ist Leiter der Stiftung „Arch Mission“.
Literatur
- E. E. Ruppert, R. S. Fox, R. D. Barnes: Invertebrate Zoology. A functional evolutionary approach. 7. Auflage. Brooks/Cole, London 2004, ISBN 0-03-025982-7, S. 510.
- R. C. Brusca, G. J. Brusca: Invertebrates. 2. Auflage. Sinauer Associates, Sunderland Mass 2002, ISBN 0-87893-097-3, S. 469.
- I. M. Kinchin: The biology of tardigrades. Portland Press, London 1994, ISBN 1-85578-043-7, S. 186.
- M. Blaxter, B. Elsworth, J. Daub: DNA taxonomy of a neglected animal phylum: an unexpected diversity of tardigrades. In: Proceedings of the Royal Society of London. Series B. London 271.2004, S. 189. ISSN 0375-0434
- G. E. Budd: The morphology of "Opabinia regalis" and the reconstruction of the arthropod stem group. In: Lethaia. Taylor & Francis, Oslo 29.1996, ISSN 0024-1164, S. 1.
- R. A. Dewel, W. C. Dewel: The place of tardigrades in arthropod evolution. In: R. A. Fortey, R. H. Thomas (Hrsg.): Arthropod Relationships. (= The Systematics Association Spec. Band 55). Chapmann and Hall, London 1998, ISBN 0-412-75420-7, S. 109.
- R. A. Dewel, D. R. Nelson, W. C. Dewel: The brain of "Echiniscus veridissimus" Peterfi, 1956 (Heterotardigrada). A key to understanding the phylogenetic position and the evolution of the arthropod head. In: Zoological Journal of the Linnean Society. Blackwell, Oxford 116.1996, ISSN 1096-3642, S. 35.
- J. R. Garey, D. R. Nelson, L. Y. Mackey, L. Li: Tardigrade phylogeny, Congruency of morphological and molecular evidence. In: Zoologischer Anzeiger. Elsevier, Jena 238.1999, ISSN 0044-5231, S. 205.
- A. Hejnol, R. Schnabel: The eutardigrade Thulinia stephaniae has an indeterminate development and the potential to regulate early blastomere ablations. In: Development. 132.2005, S. 1349.
- A. Jörgensen, R. Kristensen: Molecular Phylogeny of Tardigrada - investigation of the monophyly of Heterotardigrada. In: Molecular Phylogenetics and evolution. Elsevier, Amsterdam 32.2004, 2, ISSN 1055-7903, S. 666.
- D. R. Nelson: Tardigrada. In: J. H. Thorp, A. P. Covich (Hrsg.): Ecology and Classification of North American Freshwater Invertebrates. Academic Press, San Diego Ca 2001, ISBN 0-12-690647-5, S. 527.
- D. R. Nelson: Current Status of the Tardigrade. Evolution and Ecology. In: Integrative and Comparative Biology. Lawrence 42.2002, ISSN 1540-7063, S. 652.
- D. R. Nelson, N. J. Marley: The biology and ecology of lotic Tardigrada. In: Freshwater Biology. Blackwell, Oxford 44.2000, ISSN 0046-5070, S. 93.
- Joh. August Ephraim Goeze: Herrn Karl Bonnets Abhandlungen aus der Insektologie. (PDF; 31,2 MB). Supplement by the translator J.A.E. Goeze, Halle 1773. (Originaltext: Seltsam ist dieses Thierchen, weil der ganze Bau seines Körpers ausserordentlich und seltsam ist, und weil es in seiner äusserlichen Gestalt, dem ersten Anblicke nach, die grösste Ähnlichkeit mit einem Bäre im Kleinen hat. S. 367–375)
Weblinks
- Tardigrada Astronomy Picture of the Day der NASA vom 6. März 2013 (englisch)
- Tardigrada Newsletter (engl.)
- Bärtierchen: Fotos und Videos (engl.)
- Umfangreiche Sachtexte, Fotogalerien und Kurzfilme zu Bärtierchen
- Taxonomie bis auf Gattungsebene
- Microscopy UK (engl.)
- Forschung mit Bärtierchen im Weltall
- Schweizer Forschungsstelle für Ökologie, Physiologie und Evolutionsbiologie von Tardigraden
- A recent claim that tardigrades got a sixth of their DNA from microbes is starting to unravel (engl.)
- Claudia Ruby: Bärtierchen - Die knuffigen Überlebenskünstler Bayern 2 Radiowissen. Ausstrahlung am 25. Juli 2019 (Podcast)
- Doch nicht unverwüstlich – das ist der Schwachpunkt von Bärtierchen
- Lars Fischer: Blaues Leuchten schützt Bärtierchen vor Strahlung, auf: spektrum.de vom 15. Oktober 2020
- Peter Dockrill: We Just Found Another Trick Tardigrades Use to Be Basically Indestructible, auf: sciencealert vom 14. Oktober 2020 (engl.)
Einzelnachweise
- Andreas Hejnol, Ralf Schnabel: The eutardigrade Thulinia stephaniae has an indeterminate development and the potential to regulate early blastomere ablations. In: Development. 132, 2005, S. 1349–1361. doi:10.1242/dev.01701
- Daiki Horikawa et al.: Radiation tolerance in the tardigrade Milnesium tardigradum. Int. J. Radiat. Biol. 82, 2006, doi:10.1080/09553000600972956 (freier Volltext).
- Emma Brennand: Tardigrades: Water bears in space. Hrsg.: BBC. 17. Mai 2011 (bbc.co.uk [abgerufen am 30. November 2019]).
- Peter Reuell: Harvard study suggests asteroids might play key role in spreading life. In: Harvard Gazette. 8. Juli 2019, abgerufen am 30. November 2019 (amerikanisches Englisch).
- Lorena Rebecchi et al.: Resistance of the anhydrobiotic eutardigrade Paramacrobiotus richtersi to space flight (LIFE-TARSE mission on FOTON-M3), in: Journal of Zoological Systematics and Evolutionary Research 49, 21. April 2011, S. 98–103, doi:10.1111/j.1439-0469.2010.00606.x.
- Neil A. Campbell et al.: Biologie. 8., aktualisierte Auflage. 2011, ISBN 978-3-8273-7287-1.
- D. Schulze-Makuch et al.: Tardigrades: An Example of Multicellular Extremophiles. In: Joseph Seckbach u. a.: Polyextremophiles - life under multiple forms of stress. Springer, Dordrecht 2013, ISBN 978-94-007-6487-3, S. 597–607.
- Thomas C. Boothby, Hugo Tapia, Alexandra H. Brozena, Samantha Piszkiewicz, Austin E. Smith: Tardigrades Use Intrinsically Disordered Proteins to Survive Desiccation. In: Molecular Cell. Band 65, Nr. 6, 16. März 2017, ISSN 1097-2765, S. 975–984.e5, doi:10.1016/j.molcel.2017.02.018, PMID 28306513 (cell.com [abgerufen am 20. Juni 2017]).
- D. D. Horikawa, T. Kunieda et al.: Establishment of a rearing system of the extremotolerant tardigrade Ramazzottius varieornatus: a new model animal for astrobiology. In: Astrobiology. Band 8, Nummer 3, Juni 2008, S. 549–556, doi:10.1089/ast.2007.0139, PMID 18554084.
- Fumihisa Ono et al.: Effect of ultra-high pressure on small animals, tardigrades and Artemia.Cogent Physics 3, 2016, doi:10.1080/23311940.2016.1167575 (freier Volltext).
- B. Vargha, E. Otvös, Z. Tuba: Investigations on ecological effects of heavy metal pollution in Hungary by moss-dwelling water bears (Tardigrada), as bioindicators. In: Ann Agric Environ Med. 2002;9(2), S. 141–146. PMID 12498580
- Klaus J. Müller, Dieter Waloszek, Arcady Zakharov: 'Orsten' type phosphatized soft-integument preservation and a new record from the Middle Cambrian Kuonamka Formation in Siberia. In: Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen 197(1), 1995, S. 101–118.
- J. A. E. Goeze: Herrn Karl Bonnets Abhandlungen aus der Insektologie. J. J. Gebauer, Halle, 1773. Digitalisat abgerufen 9. September 2016. Zitiert nach www.baertierchen.de.
- Patrick Illinger: Bärtierchen sind die ersten Mondbewohner. In: Süddeutsche Zeitung, 7. August 2019