Bilateria
Die Bilateria sind die bilateralsymmetrisch gebauten dreikeimblättrigen Gewebetiere (Eumetazoa). Der wissenschaftliche Name des Taxons ist von lateinisch bis und latus abgeleitet und bedeutet „Zweiseitige“ oder „Zweiseitentiere“.
| Zweiseitentiere | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Zweiseitentiere (Bilateria) | ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Bilateria | ||||||||||
| Haeckel, 1874 |
Begriff
Im Jahr 1841 schrieb der deutsche Geowissenschaftler Heinrich Georg Bronn, dass die Körpergestalt vieler Tiere zu einer Art Keil vereinfacht werden könnte.[1] Einundzwanzig Jahre später führte der kanadische Geowissenschaftler John William Dawson das Wort Bilaterata ein, um die Gruppe der Tiere mit bilateralsymmetrischem Körperbau zu bezeichnen.[2] Im darauffolgenden Jahrzehnt prägte der deutsche Zoologe Ernst Haeckel für diese Tiere schließlich den Ausdruck Bilateria. Er stellte dem Begriff zudem das Wort Sphenota als Synonym zur Seite. Letztere Wortprägung sollte „Keiltiere“ bedeuten. Sie bezog sich auf die gewisse Keilförmigkeit der Zweitenseitentiere, die von Heinrich Georg Bronn herausgestellt worden war.[3] In aktuelleren biologischen Veröffentlichungen taucht Dawsons Wort Bilaterata nur sehr selten auf, obwohl es sich um die ältere Bezeichnung handelt.[4] Bereits seit ihrer Begriffsprägung fand Haeckels Wortalternative Sphenota kaum Beachtung. Andererseits setzte sich seine zweite Prägung Bilateria allgemein durch.[5]
Neben der Bilateralsymmetrie gehört auch die embryonale Anlage von drei Keimblättern zu den grundlegenden Merkmalen aller Zweiseitentiere. Aus diesem Umstand wurden eigene Gruppenbezeichnungen abgeleitet, die als Synonyme für den Ausdruck Bilateria gelten. Im Jahr 1873 stellte der britische Zoologe Edwin Ray Lankester das Wort Triploblastica vor.[6][7] Es wird bis heute benutzt.[8] Allerdings wird ähnlich häufig auch die abgeschliffene Ausdrucksvariante Triploblasta geschrieben.[9] Sie kann bis in die 1970er Jahre zurückverfolgt werden[10] und stellt eine latinisierte Version des englischen Fachausdrucks „Triploblasts“[11] für die „Triploblasten“[12] dar.
Merkmale
Die Bilateria zeigen wenigstens im Larvenstadium Bilateralsymmetrie, das heißt, es existiert eine linke und eine rechte Körperhälfte, die jeweils spiegelbildlich zueinander aufgebaut sind. Es sind keine weiteren Symmetrieebenen vorhanden. Im Erwachsenenstadium können einzelne Organe nicht symmetrisch ausgebildet sein, zum Beispiel die Leber des Menschen oder die Vorderscheren der Winkerkrabben. Die erwachsenen Stachelhäuter sind sekundär radiärsymmetrisch.
Alle Bilateria verfügen ursprünglich über ein Vorder- und ein Hinterende sowie einen Mund. Im Verlauf der stammesgeschichtlichen Entwicklung können diese Merkmale aber reduziert worden sein, beispielsweise bei Parasiten. Die Hauptachse des Körpers verläuft durch das Vorder- und das Hinterende. Bei einigen Gruppen der Bilateria ist diese ursprüngliche Bewegungsrichtung jedoch abgeändert, zum Beispiel schwimmen Tintenfische morphologisch gesehen nach oben oder unten.

Die erwachsenen Bilateria leben überwiegend solitär und frei beweglich. Die Hauptachse liegt meist in der Fortbewegungsrichtung. Sie suchen meist aktiv nach Nahrung, wobei die Entwicklung eines Kopfabschnittes mit einem übergeordneten Teil des Nervensystems, also einem Cerebralganglion oder Gehirn eine wesentliche Rolle spielt. Dieses als Cephalisation bezeichnete Ereignis ist während der Evolution in verschiedenen Linien der Bilateria aufgetreten.
Meist ist während der Bewegung eine bestimmte Körperseite zum Boden gerichtet. Diese wird als Bauch- oder Ventralseite bezeichnet. Sie weist entsprechende Strukturen zur Bewegung des Körpers, wie Kriechsohlen oder Extremitäten auf. Der Ventralseite entgegengesetzt liegt die Rücken- oder Dorsalseite. Die imaginäre Trennfläche zwischen Dorsal- und Ventralseite wird Dorsoventralebene genannt. Im rechten Winkel auf der Dorsoventralebene steht die Symmetrieebene des Körpers der Bilateria, die Sagittalebene. Sie trennt die rechte von der linken Körperhälfte.
Sessile und in Kolonien lebende Lebensformen sind in wenigen Gruppen der Bilateria vertreten, zum Beispiel bei Kamptozoa und Bryozoa. Diese haben zudem eine mit dieser Lebensweise verbundene filtrierende Nahrungsaufnahme.
Ein weiteres, für alle Bilateria charakteristisches Merkmal ist die Dreikeimblättrigkeit oder Triploblastie, d. h. die Existenz des Mesoderms, eines dritten Keimblattes, das während der Gastrulation angelegt wird und sich zwischen die beiden primären Keimblätter Ektoderm und Entoderm schiebt. Es ist meist mesentodermaler Herkunft, es wird also vom Entoderm gebildet. Aus dem Mesoderm wird die Muskulatur, das Bindegewebe und das Epithel um die sekundäre Leibeshöhle Coelom gebildet.
Systematik
Das Schwestertaxon der Zweiseitentiere (Bilateria) besteht entweder aus den Hohltieren (Coelenterata) oder aus den Rippenquallen (Ctenophora) oder aus den Nesseltieren (Cnidaria). Zusammen mit Rippenquallen und Nesseltieren bilden die Zweiseitentiere möglicherweise eine gemeinsame Abstammungsgemeinschaft namens Gewebetiere (Eumetazoa). Hier bestehen aber noch immer Unsicherheiten und Uneinigkeiten.[13] Verschiedene taxonomische Arbeitsgruppen führen Argumente für drei unterschiedliche Konzepte an.[14] Das Coelenterata-Konzept[15] steht neben dem Acrosomata-Konzept[16] und dem ParaHoxozoa/Planulozoa-Konzept.[17] Je nach verwendeter Systematik werden auch die beiden weiteren Großgruppen der Schwämme (Porifera)[18] und der Scheibentiere (Placozoa) an verschiedenen Positionen in der Systematik der Tiere (Metazoa) verortet.
Konzepte zur Systematik der Metazoa einschließlich der Bilateria | ||
| Coelenterata-Konzept | Acrosomata-Konzept | ParaHoxozoa/Planulozoa-Konzept |
|---|---|---|
|
|
|
Die Zweiseitentiere (Bilateria) spalten sich in die Linien der Xenacoelomorpha und der Nierentiere (Nephrozoa).[19] Während sich erstere in Xenoturbellida und Acoelomorpha gliedern,[20] wird bei letzteren zwischen Urmundtieren (Protostomia) und Neumundtieren (Deuterostomia) unterschieden. Die Urmundtiere vereinigen in sich die Taxa der Häutungstiere (Ecdysozoa) und der Spiralfurcher (Spiralia).[21] Die Neumundtiere umfassen die Gruppen der Ambulacraria und der Rückensaitentiere (Chordata).[22]
| Innere Systematik der Bilateria | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||
Evolution
Die Population der letzten gemeinsamen Vorfahren aller Zweiseitentiere wird Urbilateria genannt.[23] Die Tiere hatten eine Hauptbewegungsrichtung nach vorne evolviert. Im Zuge dessen hatten sich eine Kopfregion und eine bilateralsymmetrische Körpergestalt herausgebildet.[24] Es besteht nach wie vor kein evolutionsbiologischer Konsens darüber, ob schon die Urbilateria ein Zentralnervensystem besaßen.[25][26][27] Ungeachtet dessen konnten sie ihre kleinen wurmförmigen Körper aktiv mit Hilfe von Muskeln bewegen. Die Muskelgewebe stammten von Zellen eines embryonal angelegten Mesoderms.[28]
Eine darüber hinausgehende Rekonstruktion der Urbilateria hängt davon ab, welcher Abstammungshypothese gefolgt wird. Gemäß der Nephrozoa-Hypothese[29] spaltete sich schließlich die Gruppe der Nierentiere (Nephrozoa) von morphologisch ursprünglicheren Bilateria ab.[30] Nachfahren jener ursprünglicheren Zweiseitentiere überdauerten allerdings ebenfalls bis heute. Sie werden als Gruppe der Xenacoelomorpha zusammengefasst und den Nephrozoa gegenübergestellt. Rezente Vertreter der Xenacoelomorpha können noch immer viele der ursprünglicheren Bilateria-Merkmale zeigen. Demzufolge wird aus morphologischen Studien der Xenacoelomorpha abgeleitet, dass die Urbilateria noch keine sekundäre Leibeshöhle bildeten. Sie besaßen einen Verdauungstrakt, die mit Epithelgewebe ausgekleidet war, aber als Gastralraum nur über eine Öffnung nach außen verfügte.[31] Falls die Nephrozoa-Hypothese zutreffen sollte, evolvierten die Nierentiere aus den eben beschriebenen, acoelomorpha-ähnlichen Tieren. Erst die Nephrozoa haben dann eine sekundäre Leibeshöhle sowie einen durchgehenden Verdauungstrakt mit Mund und After evolviert.[32] Allerdings ist die Nephrozoa-Hypothese nicht ganz unumstritten.[33]
Vor ungefähr 800 Millionen Jahren spalteten sich die Zweiseitentiere von den übrigen Tieren ab. Dies legen phylogenomische Studien mit molekularer Uhr nahe. Die Spaltung fiel zeitlich in die neoproterozoische Periode des Toniums.[34] Danach evolvierten die Bilateria weiter. Gemäß der Nephrozoa-Hypothese trennten sich Xenacoelomorpha und Nephrozoa. Letztere gliederten sich schließlich vor etwa 680 Millionen Jahren während der Sturtischen Eiszeit in Urmundtiere (Protostomia) und Neumundtiere (Deuterostomia).[35]
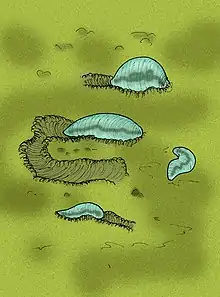
Die ältesten Fossil-Belege für das Vorhandensein von Zweiseitentieren sind allerdings mindestens 250 Millionen Jahre jünger als ihr errechnetes erstes Auftreten.[36] Sie stammen nicht aus dem Tonium, sondern erst aus der letzten Phase des Ediacariums. Dementsprechend werden zum Beispiel Fossilien der Gattung Kimberella auf 555 Millionen Jahre datiert.[37] Überreste dieser Tiere waren das erste Mal im Jahr 1959 bekannt gemacht worden[38] und ihre sorgfältige naturwissenschaftliche Erstbeschreibung erfolgte 1966. Damals wurde Kimberella noch für ein Hohltier gehalten.[39] Ab 1997 legten allerdings neue Fossilfunde nahe, dass die Gattung zu den Zweiseitentieren gehörte.[40] Im März 2020 wurde die Entdeckung eines ähnlich alten Fossils, eines Wurms, der die Bezeichnung Ikaria wariootia erhielt, veröffentlicht. Das etwa reiskorngroße Fossil wurde in Südaustralien entdeckt und auf ein Alter von 571 bis 539 Millionen Jahre datiert.[41]
Literatur
- Rüdiger Wehner, Walter Jakob Gehring: Zoologie. Georg Thieme Verlag, New York 2007, ISBN 978-3-13-367424-9.
- Wilfried Westheide, Reinhard Rieger: Spezielle Zoologie. Teil 1: Einzeller und Wirbellose Tiere. Spektrum Akademischer Verlag, Heidelberg 2007, ISBN 978-3-8274-1575-2.
Weblinks
Einzelnachweise
- Heinrich Georg Bronn: Handbuch einer Geschichte der Natur. Erster Band. In: Gottlieb Wilhelm Bischoff, Johann Reinhard Blum, Heinrich Georg Bronn, Karl Caesar von Leonhard, Friedrich Sigismund Leuckart, Friedrich Siegmund Voigt (Hg.): Naturgeschichte der drei Reiche. Band 13, 1841, S. 4.
- John William Dawson: Zoological Classification; or Cœlenterata and Protozoa, versus Radiata. In: The Canadian Naturalist and Geologist. Band 07, 1862, S. 440.
- Ernst Haeckel: Die Gastraea-Theorie, die phylogenetische Classification des Thierreichs und die Homologie der Keimblätter. In: Jenaische Zeitschrift für Naturwissenschaft. Band 08, 1874, S. 35.
- zum Beispiel Wolfgang Hennig: Genetik. Springer-Verlag, Berlin/Heidelberg, 2002, S. 17.
- zum Beispiel Joanna Fietz, Jutta Schmid: Prüfungen erfolgreich bestehen im Fach Zoologie. Verlag Eugen Ulmer, Stuttgart, 2019, S. 64, 262, 265, 266.
- Edwin Ray Lankester: On the primitive cell-layers of the embryo as the basis of genealogical classification of animals, and on the origin of vascular and lymph systems. In: The Annals and magazine of natural history; zoology, botany, and geology. Band 11, 1873, S. 330.
- Ernst Haeckel: Die Gastraea-Theorie, die phylogenetische Classification des Thierreichs und die Homologie der Keimblätter. In: Jenaische Zeitschrift für Naturwissenschaft. Band 08, 1874, S. 32.
- zum Beispiel Vitaly V. Kozin, Roman P. Kostyuchenko: Evolutionary Conservation and Variability of the Mesoderm Development in Spiralia: A Peculiar Pattern of Nereid Polychaetes. In: Biology Bulletin. Band 43, 2016, S. 216.
- zum Beispiel Roberto Ligrone: Biological Innovations that Built the World. Springer International Publishing, Cham, 2019, S. 322.
- Alice Levine Baxter: Edmund B. Wilson as a Preformationist: Some Reasons for His Acceptance of the Chromosome Theory. In: Journal of the History of Biology. Band 09, 1976, S. 39.
- George L. Purser: Preliminary notes on some Problems connected with Respiration in Insects generally and in Aquatic forms in particular. In: Proceedings of the Cambridge Philosophical Society. Band 18, 1916, S. 63.
- Adolf Lendl: Hypothese über die Entstehung von Soma- und Propagationszellen. Verlag R. Friedländer und Sohn, Berlin, 1889, S. 34.
- Bernd Schierwater, Peter W.H. Holland, David J. Miller, Peter F.Stadler, Brian M. Wiegmann, Gert Wörheide, Gregory A. Wray, Rob DeSalle: Never Ending Analysis of a Century Old Evolutionary Debate: “Unringing” the Urmetazoon Bell. In: Frontiers in Ecology and Evolution. Band 4, 2016, Artikel 5, S. 2, 9.
- Martin Dohrmann, Gert Wörheide: Novel Scenarios of Early Animal Evolution—Is It Time to Rewrite Textbooks?. In: Integrative and Comparative Biology. Band 53, 2013, S. 504, 507–508.
- Walker Pett, Marcin Adamski, Maja Adamska, Warren R. Francis, Michael Eitel, Davide Pisani, Gert Wörheide: The Role of Homology and Orthology in the Phylogenomic Analysis of Metazoan Gene Content. In: Molecular Biology and Evolution. Band 36, 2019, S. 643–649.
- Peter Ax: Multicellular Animals · Volume 1. Springer-Verlag, Berlin/Heidelberg 1996, S. 82, 104.
- Casey W. Dunn, Joseph F. Ryan: The evolution of animal genomes. In: Current Opinion in Genetics & Development. Band 35, 2015, S. 26.
- Gert Wörheide, Martin Dohrmann, Dirk Erpenbeck, Claire Larroux, Manuel Maldonado, Oliver Voigt, Carole Borchiellini, Dennis V. Lavrov: Deep Phylogeny and Evolution of Sponges (Phylum Porifera). In: Advances in Marine Biology. Band 61, 2012, S. 1.
- Johanna Taylor Cannon, Bruno Cossermelli Vellutini, Julian Smith, Fredrik Ronquist, Ulf Jondelius, Andreas Hejnol: Xenacoelomorpha is the sister group to Nephrozoa. In: Nature. Band 530, 2016, S. 89.
- John Buckland-Nicks, Kennet Lundin, Andreas Wallberg: The sperm of Xenacoelomorpha revisited: implications for the evolution of early bilaterians. In: Zoomorphology. Band 138, 2019, S. 13.
- Andreas C. Fröbius, Peter Funch: Rotiferan Hox genes give new insights into the evolution of metazoan bodyplans. In: Nature Communications. Band 8, 2017, S. 3.
- Alfred Goldschmid: Deuterostomia. In: Wilfried Westheide, Gunde Rieger (Hg.): Spezielle Zoologie • Teil 1. Springer-Verlag, Berlin/Heidelberg 2013, S. 716.
- Edward M. de Robertis, Yoshiki Sasai: A common plan for dorsoventral patterning in Bilateria. In: Nature. Band 380, 1996, S. 37.
- François Bailly, Emmanuelle Pouydebat, Bruno Watier, Vincent Bels, Philippe Souères: Should Mobile Robots Have a Head?. In: Vasiliki Vouloutsi, José Halloy, Anna Mura, Michael Mangan, Nathan Lepora, Tony J. Prescott, Paul F.M.J. Verschure (Hg.): Biomimetic and Biohybrid Systems. Springer International Publishing, Cham, 2018, S. 30.
- Leonid L. Moroz: Phylogenomics meets neuroscience: how many times might complex brains have evolved?. In: Acta Biologica Hungarica. Band 63, 2011, S. 11.
- Linda Z. Holland, João E. Carvalho, Hector Escriva, Vincent Laudet, Michael Schubert, Sebastian M. Shimeld, Jr-Kai Yu: Evolution of bilaterian central nervous systems: a single origin?. In: EvoDevo. Band 04, 2013, Artikel 27, S. 15.
- José M. Martín-Durán, Kevin Pang, Aina Børve, Henrike Semmler Lê, Anlaug Furu, Johanna Taylor Cannon, Ulf Jondelius, Andreas Hejnol: Convergent evolution of bilaterian nerve cords. In: Nature. Band 553, 2018, S. 45.
- Linda Z. Holland, João E. Carvalho, Hector Escriva, Vincent Laudet, Michael Schubert, Sebastian M. Shimeld, Jr-Kai Yu: Evolution of bilaterian central nervous systems: a single origin?. In: EvoDevo. Band 04, 2013, Artikel 27, S. 3.
- Herve Philippe, Albert J. Poustka, Marta Chiodin, Katharina J. Hoff, Christophe Dessimoz, Bartlomiej Tomiczek, Philipp H. Schiffer, Steven Müller, Daryl Domman, Matthias Horn, Heiner Kuhl, Bernd Timmermann, Noriyuki Satoh, Tomoe Hikosaka-Katayama, Hiroaki Nakano, Matthew L. Rowe, Maurice R. Elphick, Morgane Thomas-Chollier, Thomas Hankeln, Florian Mertes, Andreas Wallberg, Jonathan P. Rast, Richard R. Copley, Pedro Martinez, and Maximilian J. Telford: Mitigating Anticipated Effects of Systematic Errors Supports Sister-Group Relationship between Xenacoelomorpha and Ambulacraria. In: Current Biology. Band 29, 2019, S. 1818.
- Gonzalo Giribet, Gregory D. Edgecombe: “Perspectives in Animal Phylogeny and Evolution”: A decade later. In: Giuseppe Fusco (Hg.): Perspectives on Evolutionary and Developmental Biology. Padova University Press, 2019, S. 170.
- Johanna Taylor Cannon, Bruno Cossermelli Vellutini, Julian Smith, Fredrik Ronquist, Ulf Jondelius, Andreas Hejnol: Xenacoelomorpha is the sister group to Nephrozoa. In: Nature. Band 530, 2016, S. 89.
- Ulf Jondelius, Olga I. Raikova, Pedro Martinez: Xenacoelomorpha, a Key Group to Understand Bilaterian Evolution: Morphological and Molecular Perspectives. In: Pierre Pontarotti (Hg.): Evolution, Origin of Life, Concepts and Methods. Springer International Publishing, Cham, 2019, S. 287.
- Herve Philippe, Albert J. Poustka, Marta Chiodin, Katharina J. Hoff, Christophe Dessimoz, Bartlomiej Tomiczek, Philipp H. Schiffer, Steven Müller, Daryl Domman, Matthias Horn, Heiner Kuhl, Bernd Timmermann, Noriyuki Satoh, Tomoe Hikosaka-Katayama, Hiroaki Nakano, Matthew L. Rowe, Maurice R. Elphick, Morgane Thomas-Chollier, Thomas Hankeln, Florian Mertes, Andreas Wallberg, Jonathan P. Rast, Richard R. Copley, Pedro Martinez, and Maximilian J. Telford: Mitigating Anticipated Effects of Systematic Errors Supports Sister-Group Relationship between Xenacoelomorpha and Ambulacraria. In: Current Biology. Band 29, 2019, S. 1824.
- Kim Mikkel Cohen, David A.T. Harper, Philip Leonard Gibbard, Jun-Xuan Fan: International Chronostratigraphic Chart v 2018/08. International Commission on Stratigraphy, 2018, (Link).
- Martin Dohrmann, Gert Wörheide: Dating early animal evolution using phylogenomic data. In: Scientific Reports. Band 07, 2017, S. 4.
- John A. Cunningham, Alexander G. Liu, Stefan Bengtson, Philip C. J. Donoghue: The origin of animals: Can molecular clocks and the fossil record be reconciled?. In: BioEssays. Band 39, 2017, S. 1.
- Ilya Bobrovskiy, Janet M. Hope, Andrey Ivantsov, Benjamin J. Nettersheim, Christian Hallmann, Jochen J. Brocks: Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals. In: Science. Band 361, Nr. 6408, 21. September 2018, S. 1246–1249, doi:10.1126/science.aat7228 (englisch).
- Martin F. Glaessner, Brian Daily: The geology and Late Precambrian fauna of the Ediacara fossil reserve. In: Records of the South Australian Museum. Band 13, 1959, S. 391.
- Martin F. Glaessner, Mary Wade: The late Precambrian fossils from Ediacara, South Australia. In: Palaeontology. Band 09, 1966, S. 611–612.
- Mikhail A. Fedonkin, Benjamin M Waggoner: The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism. In: Nature. Band 388, 1997, S. 869.
- Scott D. Evans, Ian V. Hughes, James G. Gehling, Mary L. Droser: Discovery of the oldest bilaterian from the Ediacaran of South Australia. In: Proc. Natl. Acad. Sci. USA. doi:10.1073/pnas.2001045117 (englisch).