Asselspinnen
Die Asselspinnen (Pycnogonida, auch Pantopoda) bilden eine Klasse innerhalb der Kieferklauenträger (Chelicerata). Trotz ihres Namens werden sie nicht zu den Spinnentieren gerechnet. Sie sind eine rein marine Tiergruppe mit einer Verbreitung in allen Weltmeeren und einem Verbreitungsschwerpunkt im Südlichen Ozean. Ihre Zahl wird auf über 1300 Arten geschätzt.
| Asselspinnen | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Asselspinne | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Pycnogonida | ||||||||||||
| Latreille, 1810 | ||||||||||||
| Familien | ||||||||||||
|
Merkmale
Allgemeines
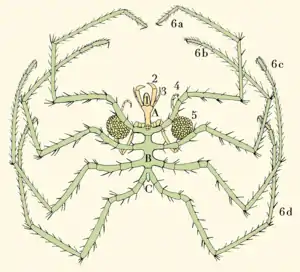
Die Pantopoda fallen vor allem durch einen, im Verhältnis zu den Beinen winzigen, stabförmigen Körper auf, der oft nur ein schmales Verbindungsstück zwischen den Beinbasen darstellt. Der Vorderkörper teilt sich in das ungegliederte Prosoma, das die ersten vier Extremitätenpaare trägt (darunter das erste Laufbeinpaar), und einen, durch Querfurchen in mehrere Segmente unterteilten, hinteren Abschnitt, dem die weiteren Laufbein-Paare anhängen. In der Regel sind es vier, bei einigen Arten bis zu sechs Paare. Der Hinterkörper (Abdomen, Opisthosoma) ist extrem reduziert und meist nur eine kleine Ausbuchtung ohne Anhänge, der am Ende den After trägt. Nur bei den fossilen Palaeopantopoden ist er noch sackförmig und lässt drei bis fünf Segmente erkennen. Neben Arten mit sehr langen Gliedmaßen kommen auch kompaktere Formen vor. Die kleinsten Asselspinnen haben eine Größe von 1 bis 10 mm, die größten unter den in der Tiefsee lebenden werden bis zu 900 mm groß. Die Länge des Körpers liegt zwischen 0,8 und 100 mm.
Gliedmaßen
Die Extremitäten ähneln denen anderer Chelicerata, doch sucht man bei ihnen die Laden (Enditen) vergeblich. Unterschieden werden vier verschiedene Arten von Gliedmaßen. Das erste Paar, die scherenbesetzten Cheliceren (bei den Pycnogonida meist Cheliforen genannt), bestehen meist aus drei, seltener aus vier Gliedern. Ihnen kommt eine Bedeutung bei der Ernährung zu. Es folgen tasterartige Palpen von wechselnder Länge (bis zehngliedrig), die mit ihrer dichten Behaarung der Reizaufnahme dienen.
Das dritte Extremitätenpaar ist das sogenannte Brutbeinpaar. Es entspringt ventral, ist gleichfalls tasterartig und dient beim Männchen als Eiträger (Oviger). Die Eipakete werden vom Männchen gebildet, indem es mit seinen Brutbeinen in der vom Weibchen abgelegten Eimasse rührt und diese mit von den Beinen abgegebenem Kitt zu Klumpen verklebt. Je nach Größe der Eier kann ein Paket 50 bis 1000 Eier beinhalten. Mit Hilfe eines aus den letzten vier Gliedern des Beines bestehenden Ringes, der mit vielen Borsten besetzt ist, gewährleistet das Männchen den sicheren Transport. Die Borsten dienen dem Männchen z. B. zur Reinigung der Eier. Bei den Weibchen hingegen ist dieses Beinpaar häufig zurückgebildet oder fehlt gänzlich.
Eine Besonderheit haben die nächsten Extremitätenpaare, die Laufbeine. Sie können, ähnlich denen der Weberknechte, bei Gefahr abgeworfen werden. Das Abwerfen hat zwei Vorteile: zum einen greifen Feinde die ihnen überlassenen langen Beine an, während die Asselspinne die Flucht ergreift; zum anderen ist es von Vorteil, ein verletztes Bein abzuwerfen, anstatt es mit sich herumzutragen und einen Flüssigkeitsverlust zu riskieren. Die Bruchstelle schließt sich sehr rasch, und das Bein wächst nach der nächsten Häutung nach. Die aus neun Gliedern bestehenden langen Laufbeine sind meist 4-, vereinzelt 5- (7 Arten, darunter Pentanymphon) oder selten sogar 6-paarig (2 Arten, Gattung Dodecalopoda). Diese mehrbeinigen Arten sind jeweils nahe mit achtbeinigen verwandt und gelten als sekundäre Abweichungen des Grundbauplans. Das Endglied, der Praetarsus, ist meist klauenförmig ausgebildet, dazu noch oft mit einer Nebenklaue besetzt und dient unter anderem dem Festhalten der Nahrung. Die drei vorderen Extremitäten-Paare können sehr variabel ausgebildet sein oder aber auch, genau wie Kiemen und Fühler, ganz fehlen.
Außenskelett
Auch diese Vertreter der Kieferklauenträger besitzen, genau wie andere Gliederfüßer, ein Exoskelett mit Chitin-Einlagerungen. Wie bei den Spinnen werden in die darunterliegende Haut Exkrete eingelagert, sodass die Tiere oft bunt gezeichnet sind. Aber auch Vorratsstoffe werden dort eingelagert. Das Exoskelett ist sehr undurchlässig und manchmal außerordentlich dick. Dagegen fehlt jedwede Einlagerung von Kalk, was zur Folge hat, dass die Haut der Pantopoda leder- oder pergamentartig ist.
Sinnesorgane und Nervensystem
In der Haut liegen zahlreiche Drüsen, wie z. B. Kittdrüsen an den Femora der Beine der Männchen und Spinndrüsen an den Cheliceren der Larven. Die Sinnesorgane sind gering entwickelt. Im Vorderkörper liegen auf einem Augenhügel vier kleine Linsenaugen (Medianaugen). An Hautsinnesorganen sind nur Sinnesborsten bekannt. Spaltsinnesorgane wurden bei dieser Klasse noch nicht gefunden.
Das Nervensystem ist primitiver als das anderer Chelicerata, da die Bauchganglien weitgehend getrennt bleiben. Das Unterschlundganglion innerviert Palpen- und Brutbeinsegment. Ein oder zwei Abdominalganglien treten während der Entwicklung noch auf, verschmelzen jedoch mit dem letzten Rumpfganglion. Die Ganglien im Rumpf sind meist deutlich sichtbar. Von diesen kann man oft starke Nervenstränge in die Beine ziehen sehen.
Verdauungstrakt
Der Mund liegt auf einem umfangreichen Rüssel (Proboscis), der ventralwärts oder nach vorn ragt. Der Rüssel besteht innen aus drei Längsteilen (Antimeren), einem dorsalen und zwei ventrolateralen. Der dreieckige Mund an der Rüsselspitze selbst ist mit drei borstenbesetzten Platten (Lippen) und drei beweglichen Chitinhaken besetzt. Der im Rüssel liegende Teil des Darms wird als Pharynx bezeichnet. Sein dreikantiges Lumen wird durch radiäre, zur Rüsselwand ziehende Muskeln erweitert, und der hintere Bereich wird durch in das Lumen ragende Chitinhaken zu einem Reusenapparat. Ein Oesophagus führt in den Mitteldarm. Da der Rauminhalt des Rumpfes bedeutend kleiner ist als der der Beine, gibt es zusätzlich lange Ausläufer des Mitteldarms (Blindsäcke), die bis in die Beine, bei manchen Arten aber auch bis in die Cheliceren und Rüssel ziehen. Das hat zur Folge, dass die aufnehmende und verdauende Oberfläche beträchtlich vergrößert wird. Die Speiseröhre, die von einem Apparat aus starren und beweglichen Borsten besetzt ist, der den groben aufgesogenen Nahrungsbrei fein zerkleinert, bis nur noch Zellbruchstücke zurückbleiben, ist lang und eng. Von hier aus gelangen diese in den Darm und werden hier von den Darmzellen, in denen die eigentliche Verdauung erfolgt, resorbiert. Der gerade Endteil des Darmes mündet mit endständigem After.
Die Exkretion läuft über sogenannte Nephrocysten, den Ausscheidungszellen, ab. Nephridien und Malpighische Gefäße fehlen diesen Vertretern der Chelicerata völlig.
Blutgefäßsystem und Atmung
Das Blutgefäßsystem besteht nur aus einem Rückengefäß. Es weist zwei Paar Einströmöffnungen (Ostien) für das farblose Blut auf und durchzieht den Rumpf vom Hinterende bis zur Region der Augenhügel und ist dorsal mit breiter Fläche an der Rückenwand, ventral am Pericardialseptum angewachsen. Oft kommt noch ein unpaares Ostium am Hinterende dazu. Das Pericardialseptum durchzieht den Rumpf horizontal dicht über dem Darm und erstreckt sich auch in die Beine.
Da den Asselspinnen Kiemen fehlen, wird die Atmung von einem anderen Organ übernommen, wahrscheinlich dem Darm[1] oder feinen Blutkapillaren, in die der Sauerstoff diffundiert.
Bau der Geschlechtsteile
Die Gonaden entstehen ventral am Pericardialseptum, erstrecken sich aber bis in die Beine, die auch den größten Anteil der Geschlechtsorgane enthalten. An den Coxen der Beine liegen auch die Genitalöffnungen, meist im zweiten Glied, daher findet man legereife Eier nie im Rumpf, sondern nur in den Beinen. Interessanterweise sind oft mehrere Paare von Öffnungen vorhanden, vielfach an allen Beinpaaren. Bei manchen Gattungen sind sie auf bestimmte Beinpaare beschränkt, am häufigsten jedoch auf die letzten. Zahl und Lage der Genitalöffnungen können in Abhängigkeit vom Geschlecht variieren.
Larvalentwicklung
Die abgelegten Eier, die je nach Art eine Größe von 0,02 bis 0,7 mm erreichen, werden wie schon beschrieben vom Männchen getragen. Die Entwicklung zeigt manche Eigenarten und ist auch innerhalb der Pantopoden nicht gleichartig. Die Furchung ist zunächst total und kann je nach Dottergehalt äqual oder inäqual ausfallen. Früher oder später verschmelzen aber Zellen zu syncytialen Massen. Die Keimblätterbildung ist schwer verständlich. Dorsal werden große Zellen ins Innere verlagert, welche Entoderm (zum Teil von einer Urentodermzelle ausgehend) und Mesoderm bilden. Später tritt ventral die Längsrinne auf, die dem Blastoporusgebiet anderer Arthropoden entspricht. Das Mesoderm scheint sich stets über ein einfaches Streifenstadium in Muskeln und Bindegewebe umzuwandeln.
Die Embryonalentwicklung führt zu einer typischen Larve, der Protonymphon-Larve. Bei den drei Extremitätenpaaren, über die die Larve anfangs verfügt, handelt es sich um Cheliceren, Palpen und Brutbeine. Die Cheliceren der Protonymphon-Larve tragen eine seitlich in eine Röhre mündende Spinndrüse und zum Teil Scherendrüsen. Die beiden anderen Extremitäten sind jedoch nur dreigliedrige Haken, die später mehr oder weniger zurückgebildet werden, während die definitiven Palpen und Brutbeine durch Neubildung entstehen. Herz und After fehlen der Larve. Die Weiterentwicklung erfolgt durch schrittweise Bildung der Beine am Hinterkörper, die Stadien sind durch Häutungen getrennt. Nur selten bleiben die Larvalstadien an den Brutbeinen der Männchen (zum Beispiel Chaetonymphon), meist verlassen sie als Protonymphon die Brutbeine und leben in der nächsten Phase als Ekto- oder Endoparasiten (Phoxichilidium, Anoplodactylus) an anderen Tieren, vor allem Polypen.
Lebensweise und Verbreitung
Die Vertreter der Pycnogonida sind ausschließlich marin zuhause und leben zwischen Bodenbewuchs aller Art. Dabei sind sie nicht an eine bestimmte Tiefe gebunden, sondern sind sowohl an der Oberfläche als auch in der Tiefsee in Tiefen von mehr als 4000 m heimisch. Einige besonders kleine Arten leben im Sandlückensystem (Interstitial). Abhängig sind sie einzig von einem bestimmten Salzgehalt, der bei ungefähr 3,5 % liegt. Des Weiteren bevorzugen sie kaltes Wasser. Daher findet man sie auffallend häufig in der Antarktis (etwa 250 der 1000 bekannten Arten, davon 100 Arten endemisch in der Antarktis und rund 60 in den subantarktischen Gewässern), dort auch in den großen Formen. In den warmen Meeren, zum Beispiel an den Küsten des Mittelmeeres, fand man bisher nur kleine Exemplare mit höchstens 30 mm Durchmesser. Alle Pantopoden sind durchweg träge Tiere, wobei sich die kurzbeinigen, plumpen Arten durch ganz besondere Schwerfälligkeit auszeichnen. Sie lassen sich, gibt man sie in eine Schale, zu Boden sinken und bleiben regungslos liegen. Die schlankeren Formen können jedoch mehr oder weniger grazil schwimmen und sich auf diese Weise längere Zeit im freien Wasser aufhalten. Alle Asselspinnen sind Kletterer, die sich langsam und bedächtig bewegen und sich an jedem geeigneten Gegenstand festklammern können. Die Fortbewegungsgeschwindigkeit ist recht langsam (ca. 1 bis 3 mm/s), kann jedoch in Gefahrensituation enorm gesteigert werden.
Nahrung
Alle Vertreter der Asselspinnen ernähren sich räuberisch. Zu ihrer Nahrung gehören ausschließlich weichhäutige Tiere, so beispielsweise Schnecken, Moostierchen und Schwämme, aber vor allem Hydroidpolypen. Die Nesselzellen der Polypen scheinen auf Asselspinnen keinerlei Wirkung zu haben. Die Nahrung, zum Beispiel ein Polypenköpfchen, wird mit einer Schere gefasst und mit dem Rüssel ausgesogen. Dieser Vorgang kann bis zu 10 Minuten dauern. Des Weiteren wurde auch die Aufnahme von Ruderfußkrebsen und Vielborstern beobachtet. Die Ruderfußkrebse werden mit Hilfe der Greifklauen der Laufbeine gepackt, zum Mund geführt und ausgesaugt. Außerdem wurde in Experimenten das Fressen von Muschelfleisch untersucht. Auch hier werden die Greifklauen der Beine eingesetzt, um das Fleisch festzuhalten und die Nahrung anschließend über den Proboscis aufzunehmen. Dieser Vorgang kann sich über mehrere Stunden erstrecken.[2][3]
Eine Reihe von Arten lebt ektoparasitisch (auf Hohltieren, Schwämmen, Mollusken und Stachelhäutern).
Abwehr von Fressfeinden
Bei der Knotigen Asselspinne (Pycnogonum litorale) wurde erstmals in einer marinen Räuber-Beute-Beziehung eine Methode der chemischen Abwehr gefunden.[4] Es konnte nachgewiesen werden, dass die Gemeine Strandkrabbe (Carcinus maenas), die sonst nahezu alles frisst, Asselspinnen meidet, weil diese in allen Stadien einen sehr hohen Gehalt an 20-Hydroxy-Ecdyson haben. Diese Substanz ist ein Hormon, das bei Insekten und Krebstieren die Häutung (Ecdysis) auslöst. Für die Strandkrabben sind häufige Häutungen nachteilig, nicht zuletzt weil frisch gehäutete Tiere noch sehr weiche Mundwerkzeuge haben, die eine Nahrungsaufnahme für eine gewisse Zeit unmöglich machen. Die Asselspinnen steuern ihre Häutungen offenbar anders. Ein Häutungshormon für diese Tiere ist bisher noch unbekannt.
Namensgebung
Auch für die Asselspinnen haben sich, wie bei allen Tier- und Pflanzengruppen, mehrere Namen erhalten. So wurden sie um 1815 vom Engländer William Elford Leach als Podosomata bezeichnet, was so viel wie „Körper (nur) aus Beinen“ bedeutet. 1863 wiederum beschrieb sie der deutsche Zoologe Carl Eduard Adolph Gerstäcker als Pantopoda, was man frei mit „die Allesbeinigen“ übersetzen kann. Ein gewisses Maß an Unsicherheit drückt der deutsche Name Asselspinnen aus. Lange Zeit wurden sie zu den Krebsen gestellt, da sie aber auch spinnenförmig aussehen und auch einige Gemeinsamkeiten aufwiesen, reihte man sie in die Klasse der Spinnentiere ein. Da man aber ihre Eigentümlichkeit betonen wollte, nannte man sie letztendlich Asselspinnen.[3]
Systematik
Zwischenzeitlich aufgekommene Vermutungen, die Asselspinnen bildeten einen eigenständigen Stamm basal zu allen anderen Arthropoda gelten heute als widerlegt. Die meisten Taxonomen setzen den Asselspinnen als basalster Gruppe der Chelicerata alle anderen Vertreter als Schwestergruppe gegenüber. Damit sind die Pfeilschwanzkrebse und die Spinnentiere miteinander näher verwandt als jede dieser Gruppen mit den Asselspinnen.
| Gliederfüßer (Arthropoda) |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Die so dargestellte traditionelle Systematik (basierend vor allem auf morphologischen Reihen mit fortschreitender Reduktion einzelner Gliedmaßen) wird von moderneren molekularen und kladistisch-morphologischen Studien nur teilweise gestützt. Die meisten Ordnungen und einige Familien erwiesen sich als paraphyletisch.
Fossile Überlieferung
Fossilien von Asselspinnen werden nur sehr selten gefunden. Eine Reihe problematischer Fossilien mit stark abweichendem Körperbau gelten heute nicht mehr als Stammgruppenvertreter, sondern wurden anderen Verwandtschaftskreisen zugeordnet.[6] Die verbleibenden Arten können aufgrund ihres Bauplans modernen Ordnungen zugewiesen werden, ihre früher übliche Zusammenfassung als „Palaeopantopoda“ ist demnach eine künstliche Einteilung. Besonders reich an fossilen Arten ist der unterdevonische Dachschiefer des Hunsrücks. Die ältesten Formen sind in Körpererhaltung (d. h. nicht nur als Abdruck) erhaltene Larven aus dem Oberkambrium von Schweden (dabei wurde die Körperwand durch Calciumphosphat ersetzt und das Tier anschließend in Kalkstein eingebettet, sog „Orsten“-Fossilien). Die adulten Formen dazu sind unbekannt.[7]
Literatur
- Claudia P. Arango, Ward C. Wheeler: Phylogeny of the sea spiders (Arthropoda, Pycnogonida) based on direct optimization of six loci and morphology. In: Cladistics. Band 23, 2007, S. 255–293, doi:10.1111/j.1096-0031.2007.00143.x.
- F. Arnaud, R. N. Bamber: The biology of Pycnogonida. In: J. H. S. Blaxter, Alan J. Southward (Hrsg.): Advances in Marine Biology. Band 24, Academic Press, 1988, ISBN 0-12-026124-3, S. 1–95.
- Volker Storch, Ulrich Welsch: Systematische Zoologie. 6. Auflage. Spektrum-Verlag. München 2004, ISBN 3-8274-1112-2.
- Hans Eckard Gruner, Hans-Joachim Hannemann, Gerhard Hartwich: Wirbellose Tiere. (= Urania Tierreich. Band 2). Urania, Freiburg/B. 1994, ISBN 3-332-00502-2.
- Joel W. Hedgpeth: Pycnogonid. In: Encyclopedia Britannica. 1977.
- Philip Ernst King: Pycnogonids. Hutchinson, London 1973, ISBN 0-09-116460-5.
- Jan C. Loman: Biologische Beobachtungen an einem Pantopoden. In: Tijdschrift van de Nederlandse Dierkundige Vereniging. Band 2, 1907, S. 255–284, Tafel V.
- T. Munilla, A. Membrives: Check-List of the pycnogonids from Antarctic and sub-Antarctic waters: zoogeographic implications. In: Antarctic Science. Band 21, 2009, S. 99–111, doi:10.1017/S095410200800151X.
Weblinks
Einzelnachweise
- Pantopōden. In: Meyers Konversations-Lexikon. 4. Auflage. Band 12, Verlag des Bibliographischen Instituts, Leipzig/Wien 1885–1892, S. 659–660.
- G. Lotz: Nahrungsaufnahme und Beutefang bei einem Pantopoden, Anoplodactylus petiolatus Krøyer. In: Oecologia. Band 1, 1968, S. 171–175. doi:10.1007/BF00383137
- H. Clémençon: Asselspinnen – Bewohner der Meeresküsten. Teil 1: Bau und Leben, Beobachtung und Präparation. Teil 2: Bestimmungsschlüssel. In: Mikrokosmos. Band 50, 1961, S. 262–270.
- K.-H. Tomaschko: Ecdysteroids from Pycnogonum littorale (Arthropoda, Pantopoda) act as chemical defense against Carcinus maenas (Crustacea, Decapoda). In: Journal of Chemical Ecology. Band 20, 1994, S. 1445–1455. doi:10.1007/BF02059872
- Arthur D. Chapman: Numbers of Living Species in Australia and the World. 2. Auflage. Report for the Australian Biological Resources Study, Canberra 2009, ISBN 978-0-642-56861-8. (environment.gov.au)
- J. W. Hedgpeth: A reappraisal of the Palaeopantopoda with description of a species from the Jurassic. In: Zoological Journal of the Linnean Society. Band 63, 1978, S. 23–34. doi:10.1111/j.1096-3642.1978.tb02088.x
- D. Waloszek, J. A. Dunlop: A larval sea spider (Arthropoda: Pycnogonida) from the upper Cambrian Orsten of Sweden, and the phylogenetic position of pycnogonids. In: Palaeontology. Band 45, 2002, S. 421–446. open access