Homologie (Biologie)
Als Homologie (altgriechisch ὁμολογεῖν homologein „übereinstimmen“) bezeichnet man in der biologischen Systematik und der vergleichenden Anatomie die grundsätzliche Übereinstimmung von Organen, Organsystemen, Körperstrukturen, physiologischen Prozessen oder Verhaltensweisen zweier Taxa aufgrund ihres gemeinsamen evolutionären Ursprungs. In diesem Artikel wird also auf die Homologie im Sinne des Phänotyps eingegangen.
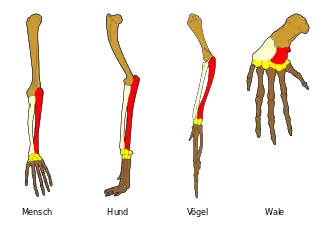
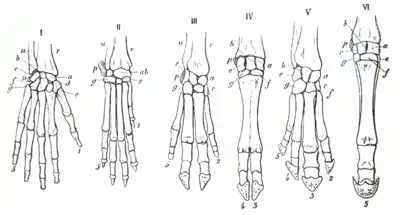
I Mensch II Hund III Schwein IV Kuh V Tapir VI Pferd
Homologe Merkmale gehen auf Merkmale des gemeinsamen Vorfahren zurück, sie sind also gleichwertig bezüglich ihrer stammesgeschichtlichen Herkunft. Die ursprünglichen Merkmale können sich danach in verschiedene Richtungen entwickelt haben und in unterschiedlichen Funktionen gebraucht werden.
„Ein Merkmal zweier oder mehrerer Taxa ist homolog, wenn es sich von demselben (oder einem entsprechenden) Merkmal ihres nächsten gemeinsamen Vorfahren ableitet.“
Homologe Merkmale spielen bei der Erstellung von Stammbäumen in der klassischen biologischen Systematik eine wichtige Rolle.
Begriffsabgrenzung: Homologie in der Genetik und Zytogenetik
Zytogenetik
Bei der Betrachtung der Chromosomen einer Zelle wird der Begriff Homologie in einem anderen Zusammenhang verwendet. Hier bezeichnet man jene zwei Chromosomen als homolog, welche die gleichen Gene enthalten. Wenn es sich um einen Organismus mit geschlechtlicher Fortpflanzung handelt, wurde je eines der beiden homologen Chromosomen vom Vater und eines von der Mutter ererbt.
Auch bei Vergleichen zwischen zwei nahe verwandten Arten können Chromosomen homologisiert werden, die auf einen gemeinsamen Vorläufer zurückzuführen sind. In diesem erweiterten Sinn sind zwei Chromosomen verschiedener Arten homolog, wenn sie einen sehr ähnlichen Inhalt aufweisen.
Genetik
In der Genetik und Evolutionsbiologie wird der Begriff der Homologie auch auf der Ebene der Gene angewandt, die in ihrer Sequenz auf einen gemeinsamen Vorläufer zurückzuführen sind.
Begriffsgeschichte
In vordarwinscher Zeit verband man mit dem Begriff der Homologie die Ähnlichkeit von Merkmalen bei verschiedenen Spezies unabhängig von ihrer Form und Funktion, aber in Abhängigkeit von ihrer Lage oder ihrer Entwicklung im Organismus.[3] Arbeiten dazu finden sich beim französischen Arzt Félix Vicq d’Azyr (um 1770) und etwas später bei Goethe (1790).[4] Der Begründer der Paläontologie Georges Cuvier ordnete (etwa ab 1803) Fossilien aufgrund des anatomischen Vergleichs homologer Strukturen mit rezenten Organismen erfolgreich in den Stammbaum ein. 1843 führte Richard Owen (1804–1892) den Begriff „Homology“ in die Literatur ein. Mit der Erkenntnis der Veränderlichkeit der Arten, von Darwin 1859 veröffentlicht, wurde es notwendig, Owens ursprüngliche „Erklärung“ der Homologien zu korrigieren. Anstelle der Strukturierung nach Lage oder Entwicklung wurden Homologien mehr und mehr in Abhängigkeit von der Abstammung definiert. Wobei die Vielzahl der gefundenen Homologien die evolutionäre Theorie Darwins schon zu Beginn förderte. In Unkenntnis der Vererbungsregeln wurde zunächst ein Konzept von abstrakten „Bauplänen“ postuliert, bis das Auftreten von Homologien schließlich durch die Erkenntnisse der Genetiker tatsächlich aufgeklärt wurde.
Homologiekriterien
Um eine Homologie zwischen anatomischen Merkmalen festzustellen, wurden 1952 von dem Zoologen Adolf Remane (1898–1976) die drei Hauptkriterien „Lage“, „Struktur“ und „Kontinuität“ vorgeschlagen. Von diesen dreien muss mindestens eines zutreffen, damit man von einer "Homologie" sprechen kann. Anhand der Ähnlichkeiten lasse sich dann auf die Verwandtschaft der Organismen, und damit eine Homologie rückschließen.
1. Kriterium der Lage
Strukturen sind dann homolog, wenn sie trotz unterschiedlicher Ausprägung in Gestalt und Anzahl in einem vergleichbaren Gefügesystem stets die gleiche Lagebeziehung aufweisen.
Beispiele:
- Innerhalb der Klasse der Insekten erfüllen die verschieden gestalteten Insektenbeine das Kriterium der Lage, denn sie sind alle am Thorax des Tieres angesetzt.
- Innerhalb der Klasse der Wirbeltiere gilt ähnliches für die Verdauungsorgane, die sich in Mund–Speiseröhre–Magen–Darm–After gliedern lassen.
- Ebenso der Aufbau des Herzens bei Säugetieren (linke Kammer, rechte Kammer, linker und rechter Vorhof sowie ihre Anbindung an die Blutgefäße).
- Aufbau der Vordergliedmaßen von Landwirbeltieren. Der Grundbauplan besteht aus Knochen, die in der Abfolge von einem Oberarmknochen, zwei Unterarmknochen, mehreren Handwurzelknochen, fünf Mittelhandknochen und fünf Fingern, wobei der Daumen zwei Fingerknochen hat und die restlichen vier jeweils aus drei Fingerknochen angelegt werden.
- Trotz identischer innerer Struktur kann das Aussehen und die Funktion der Vordergliedmaßen bei verschiedenen Wirbeltiereklassen sehr verschieden sein, denn sie dienen dem Laufen, Graben, Fliegen, Greifen oder Schwimmen. Im Verlauf der stammesgeschichtlichen Entwicklung haben sich Spezialisierungen der Vorderextremität herausgebildet. Erst der gleiche Grundbauplan weist auf gemeinsame Vorfahren hin, von denen aus sich die Entwicklung zu den heutigen Formen vollzogen hat.
2. Kriterium der spezifischen Qualität und Struktur
Ähnliche Strukturen können auch ohne Rücksicht auf die gleiche Lage homologisiert werden, wenn sie in zahlreichen Merkmalen übereinstimmen. Die Sicherheit wächst mit dem Grad der Komplexität der verglichenen Struktur. Maßgeblich für das Homologiekriterium der spezifischen Qualität ist also der „innere Aufbau“ eines Organs oder einer Struktur.
Beispiele:
- Nach dem Homologiekriterium der spezifischen Qualität sind der menschliche Zahn und die Hautschuppe eines Hais homolog, weil die äußerste Schicht aus Zahnschmelz und die darunterliegende Schicht aus Dentin besteht. Da sich Zähne wie Placoidschuppen als Spezifikationen eines urtümlich knöchernen Außenskeletts verstehen lassen, sind beide Strukturen tatsächlich homolog.
3. Kriterium der Kontinuität (Stetigkeit)
Organe sind dann homolog, wenn sie trotz unterschiedlicher Lage durch Zwischenformen, die sich homologisieren lassen, in Verbindung gebracht werden können.
Beispiele:
- Bestimmte Teile des Blutgefäßsystems (Halsschlagader) von Säugetieren lassen sich aufgrund von Embryonalstadien („Kiemenspalte“) mit den Kiemenbogenarterien der Fische homologisieren.
- die Schwimmblase von Knochenfischen und die Lunge von Landwirbeltieren sind homolog, da sich die Entwicklung von der Schwimmblase zur Lunge mit Hilfe der Amphibien und Sauropsiden nachvollziehen lässt.
Rezente Zwischenformen
Beispiel: Die Homologie des Articulare und Quadratum der Amphibien, Reptilien und Vögel mit Amboss und Hammer im Mittelohr der Säugetiere, sowie der hyomandibularen Columella der Vögel, Reptilien und Amphibien mit dem Steigbügel der Säugetiere wurde bereits 1837 von Carl Bogislaus Reichert (1811–1883) entdeckt.
Rudimente und Atavismen
Ein weiterer anatomischer Beleg für die Evolution sind Rudimente. Darunter versteht man rückgebildete Reste von Organen, die ihre Funktion verloren haben und dadurch überflüssig geworden sind. Zum Beispiel ist der Beckengürtel beim Bartenwal ein Rudiment. Erlangen diese Organe im Laufe der Zeit keine neuen Funktionen, können sie ganz verloren gehen: So ist die Blindschleiche keine Schlange, sondern eine Eidechse, deren Extremitäten vollständig zurückgebildet sind. In seltenen Fällen können rückgebildete Organe wieder auftreten, etwa beim Menschen der Fortsatz des Steißbeins als Schwanz. Man spricht in diesen Fällen von Atavismen.
Embryonale Zwischenformen
Während des embryonalen Entwicklungsprozesses werden Homologien zu spezifischen Arten sichtbar, die allerdings nur von ephemerer (vorübergehender) Dauer und beim endgültig entwickelten Lebewesen nicht mehr feststellbar sind. Diese Zwischenformen führten Ernst Haeckel zu der Annahme, dass jedes Säugetier während seiner embryonalen Entwicklung in kurzer Zeit eine vollständige Phylogenese (vollständige Stammesentwicklung seiner Art) durchläuft. Er formulierte dies in der Biogenetischen Grundregel, deren moderne Fassung so formuliert ist, dass in jedem Stadium der Ontogenese tiefgreifend umgestaltende Veränderungen von evolutionärer Bedeutung auftreten können.
Beispiele:
- Der menschliche Embryo weist in einem bestimmten Entwicklungsstadium eine Kiemenspalte (Kiemenanlage) und damit eine unverkennbare Ähnlichkeit mit den Fischen auf, die später wieder verschwindet.
- Der Vogelflügel ist mit der Hand des Menschen homologisierbar, da der Vogelembryo noch eine Hand mit fünf Fingern aufweist, die im Laufe seiner Entwicklung durch Verschmelzung und Reduktion umgebaut werden.
Fossile Zwischenformen (auch fossile Brückentiere)
Anhand von gefundenen und restaurierten Fossilien lässt sich die schrittweise Entwicklung der Evolution belegen. Dies geschieht entweder durch Progressionsreihen oder Regressionsreihen, also eine nachvollziehbare Entwicklung von weniger komplexen Strukturen zu sehr komplexen Strukturen bzw. umgekehrt.
Beispiel: Archaeopteryx. Der sogenannte Urvogel zeigt den Übergang zwischen Landbewohnern und dem Lebensraum Luft, da er sowohl Merkmale von Reptilien als auch von Vögeln besitzt.
Ethologische Übergänge
Auch ähnliche Verhaltensweisen können homolog sein, wie speziell die klassische ethologische Forschung herausgearbeitet hat. Beispiele: das (angeborene) Balzverhalten innerhalb verwandter Vogelgruppen, etwa von verschiedenen Entenarten oder Hühnerartigen.
Analogie
Die Ähnlichkeit eines Merkmals ist nicht immer auf einen gemeinsamen Vorfahren zurückzuführen. Waren verschiedene Arten einem ähnlichen Selektionsdruck ausgesetzt, können sie ähnliche Merkmale auch unabhängig voneinander entwickeln. Sind solche analogen Merkmale vorhanden, spricht man von konvergenter Entwicklung oder kurz von Konvergenz oder Analogie. Analogien erfüllen ähnliche Aufgaben, sind aber nicht auf einen gemeinsamen Vorfahren zurückzuführen. Sie beruhen daher auch nicht auf gemeinsamen Erbanlagen und sind nicht geeignet, eine enge evolutive Verwandtschaft zu begründen. Vereinfacht ausgedrückt beruhen homologe Merkmale auf gemeinsamen Vorfahren und analoge Merkmale auf gemeinsamer Umwelt.
Ein Beispiel für Analogie sind die Flügel der Vögel und die Flügel der Fledermäuse bezüglich der Tragfläche. Federn und Flughaut sind nicht aus einem gemeinsamen fliegenden Vorfahren hervorgegangen, sondern unabhängig voneinander zweimal entstanden. Weitere Beispiele für analoge Merkmale sind:
- Beinlosigkeit bei Schlangen und bei Eidechsen
- die zum Graben geeigneten Vorderextremitäten von Maulwurf (Säugetier) und Maulwurfsgrille (Insekt)
- die stechend-saugenden Rüssel von Wanzen (Hemiptera) und Stechmücken (Diptera)
- Sukkulenz bei Kakteen der Neuen Welt und bei einigen Euphorbiaceen der Alten Welt
- Weiße oder nicht gefärbte Eier bei Vögeln, die in Höhlen brüten
Die Unterscheidung zwischen analogen und homologen Merkmalen ist nicht in jedem Fall zweifelsfrei möglich, sie ist skalenabhängig, d. h. bei unterschiedlichen Zeit- und Betrachtungsebenen muss dasselbe Merkmal unterschiedlich bewertet werden. Beispielsweise sind die zum Fliegen verwendeten Vorderextremitäten von Vögeln und Fledermäusen anatomisch homolog. Betrachtet man aber anstelle z. B. des Knochenbaus das Merkmal „Flugfähigkeit“, handelt es sich um eine Analogie (da der letzte gemeinsame Vorfahre zwar eine anatomisch entsprechende Vorderextremität mit denselben Knochen besaß, aber nicht flugfähig war). Fachleute verwenden bei der Verwandtschaftsanalyse möglichst keine Merkmale, die „bekanntermaßen zu Konvergenz neigen“.[5]
Umgekehrt beobachtet man bei nahe verwandten Arten homologe Merkmale, die durch divergente Entwicklungen, Gendrift oder Flaschenhalseffekt ganz unterschiedlich erscheinen, manchmal sind Homologien auf den ersten Blick gar nicht mehr erkennbar, beispielsweise wenn ein Merkmal bei der einen Art vollständig reduziert wird. Ohne Fossilfunde, Embryonalstadien, genetische Untersuchungen oder die Kenntnis von Zwischenstadien bei rezenten Arten ist dann schwer entscheidbar, wie die Abwesenheit dieses Merkmals zu deuten ist. Zum Beispiel sind Nacktschnecken trotz des Fehlens eines Gehäuses mit den gehäusetragenden Schnecken eng verwandt, denn die gemeinsamen Vorfahren trugen ein Gehäuse, welches die Nacktschnecken dann verloren haben.
Anstelle des Begriffs Analogie wird in der neueren Fachliteratur der umfassendere Ausdruck Homoplasie als Gegensatz zur Homologie bevorzugt.[6] Homoplasie umfasst neben analoger Entwicklung außerdem die parallele Evolution (z. B. parallele Entwicklung eines Streifenmusters mit unterschiedlicher molekularer Grundlage aus einem ungestreiften Vorfahren bei zwei Fischarten) und die evolutionäre Rückentwicklung, bei der ein verlorenes Merkmal in einer Entwicklungslinie durch Mutation erneut auftritt. Während anatomische Rückentwicklungen sehr selten sind (ein Beispiel wäre die Wiederentwicklung funktionsfähiger Flügel innerhalb der flugunfähigen Gespenstschrecken[7]) spielt sie beim Vergleich homologer DNA-Sequenzen eine sehr große Rolle, da aufgrund der vier Basenpaare hier nur vier Permutationen für jede Position möglich sind.
Verständnis der phylogenetischen Systematik
Merkmale, die auf einen gemeinsamen Ursprung in der Phylogenese zurückzuführen sind, sind homolog. Daher gehörten Homologien zu den ersten Kriterien, die genutzt wurden, um Verwandtschaften zwischen den Taxa zu begründen.
1950 wird von Willi Hennig der Begriff der Homologie durch die Begriffe der phylogenetischen Systematik (Kladismus) ergänzt bzw. präzisiert:
- Apomorphie liegt bei einem vergleichsweise abgeleiteten (in der Stammesgeschichte später erworbenen) Merkmal vor.
- Plesiomorphie liegt bei einem vergleichsweise ursprünglichen (also stammesgeschichtlich früher erworbenen) Merkmal vor.
- Autapomorphie ist der Neuerwerb von Merkmalen in einer Stammart (Beispiel: die Feder bei der Stammart aller Vögel).
- Synapomorphie ist das Auftreten von abgeleiteten Merkmalen einer Stammart bei ihren beiden Tochterarten und deren Nachfahren (Beispiel: Echoortung bei Zahnwalen und Bartenwalen).
- Symplesiomorphie ist das Auftreten von gemeinsamen Merkmalen bei verschiedenen Arten, das nicht auf deren letzte gemeinsame Stammart zurückzuführen ist, sondern auf eine noch ältere Stammart, die auch der Vorfahre anderer Arten ist (Beispiel: Haare bei Schimpanse und Mensch).
Die Bewertung der Merkmale ist also abhängig von der Leserichtung (ursprünglich vs. abgeleitet) und Homologie (Rückführung auf einen gemeinsamen Grundplan): Das Merkmal „Neurocranium“ ist eine Autapomorphie der Schädeltiere, eine Synapomorphie aller zu den Schädeltieren gehörenden Gruppen und eine Plesiomorphie für einzelne zu den Schädeltieren gehörende Gruppen. Der apomorphe Merkmalszustand ist an den Grundplan geknüpft, in dem er zuerst auftritt, in Grundplänen, die aus der weiteren Aufspaltung der resultierenden Gruppe hervorgehen ist dasselbe Merkmal dann im plesiomorphen Zustand.
Der Begriff des Bauplans wird durch den Begriff Grundplan oder Grundmuster ersetzt. Letzterer ist definiert als die Summe aller Plesiomorphien und Autapomorphien der letzten gemeinsamen Stammart einer monophyletischen Gruppe.
Zur Anwendung dieses Konzeptes siehe Kladistik und phylogenetische Systematik, zu seiner Kritik siehe Kritische Evolutionstheorie.
Literatur
- Walter M. Fitch: Homology: a personal view on some of the problems. (PDF) In: Trends in Genetics. 16, Nr. 5, Mai 2000, S. 227–31. doi:10.1016/S0168-9525(00)02005-9. PMID 10782117.
Anmerkungen
- Abbildung aus: Gegenbaur 1870
- Ernst Mayr: Das ist Leben – die Wissenschaft vom Leben. Spektrum Akademischer Verlag, Heidelberg, 1997. S. 189
- Manfred Laubichler: Das Forschungsprogramm der evolutionären Entwicklungsbiologie, in: Philosophie der Biologie. Hrsg. Ulrich Krohs und Georg Toepfer, Suhrkamp, Frankfurt am Main 2005, S. 328
- Versuch die Metamorphose der Pflanzen zu erklären, gedruckt bei Carl Wilhelm Ettinger, Gotha 1790
- Douglas F. Futuyma: Evolutionsbiologie, Birkhäuser, Basel 1990, S. 334 f und 346f.
- vgl. z. B. Anastasia Thanukos: Bringing homologies into focus. Evolution: Education and outreach, 1, 2008, S. 498–504. doi:10.1007/s12052-008-0080-5.
- Michael F. Whiting, Sven Bradler, Taylor Maxwell: Loss and recovery of wings in stick insects. In: Nature, Band 421, 2003, S. 264–267. doi:10.1038/nature01313.