Schlauchpilze
Die Schlauchpilze (Ascomycota) bilden eine der großen Abteilungen im Reich der Pilze (Fungi). Sie tragen den Namen nach ihren charakteristischen Fortpflanzungsstrukturen, den schlauchförmigen Asci. Viele Hefe- und Schimmelpilze, aber auch begehrte Speisepilze wie Morcheln und Trüffeln gehören zu dieser Gruppe. Stammesgeschichtlich sind sie sehr wahrscheinlich die Schwestergruppe der Ständerpilze (Basidiomycota) und bilden mit diesen das Taxon Dikarya. Eine Mehrzahl der früher in eine eigenständige Gruppe gestellten Pilze ohne sexuelle Stadien im Lebenszyklus (Deuteromycota) können durch molekulargenetische Daten heute in die Schlauchpilze eingeordnet werden.
| Schlauchpilze | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Ascokarp der Frühjahrs-Giftlorchel | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Ascomycota | ||||||||||||
| Caval.-Sm.[1] |
Schlauchpilze haben eine große Bedeutung für den Menschen, da sie einerseits für zahlreiche Krankheiten von Pflanzen, Haustieren und Menschen verantwortlich sind, andererseits aber auch eine wichtige Rolle bei der Herstellung von Lebensmitteln wie Käse und Brot, Bier und Wein spielen und auch in der Medizin, wo das von Penicillium chrysogenum produzierte Antibiotikum Penicillin die Bekämpfung von bakteriellen Infektionskrankheiten revolutioniert hat, von nicht zu unterschätzender Wichtigkeit sind. Schlauchpilze übernehmen als Pilzpartner in Flechten und als so genannte Mykorrhiza-Pilze zudem eine wichtige Stellung in vielen Ökosystemen.
Aufbau
Viele Schlauchpilze setzen sich aus langen, aber nur etwa fünf Mikrometer dünnen Zellfäden, den Hyphen, zusammen, die sich oft wiederholt verzweigen und so ein wollig verknäultes Geflecht, das Myzel, bilden, das meist einige Zentimeter groß wird. Die darin enthaltenen Hyphen selbst würden dagegen (hypothetisch aneinandergelegt) meist eine Länge von einem oder sogar mehreren Kilometern erreichen. Das andere Extrem bilden die einzelligen Hefen, die oft nur mit dem Mikroskop zu sehen sind. Eine Reihe von Arten wie beispielsweise die klassische Backhefe (Saccharomyces cerevisiae) ist allerdings dimorph, das bedeutet, dass sie sowohl in ein- als auch in mehrzelliger Form auftreten können.
Die Zellwand besteht bei Schlauchpilzen fast immer aus Chitin und β-Glukanen; einzelne Zellen sind in charakteristischer Weise durch Querwände, die Septen, abgeteilt. Diese geben der Hyphe Stabilität und verhindern einen großflächigen Verlust von Zellplasma, falls die Zellmembran lokal einmal aufbrechen sollte. Als Resultat können sich Schlauchpilze im Gegensatz zu den feuchtigkeitsliebenden Jochpilzen (Zygomycota) auch in trockenen Lebensräumen halten. Die meisten Querwände sind allerdings zentral perforiert, weisen also eine kleine Öffnung in der Mitte auf, durch die sich Zellplasma und auch Zellkerne mehr oder weniger frei in der ganzen Hyphe bewegen können. Die meisten Hyphen enthalten nur einen Zellkern pro Zelle, man bezeichnet sie daher als uninukleat.
Das Ascokarp


Bei vielen, bei weitem aber nicht allen Schlauchpilzen kommt es im Zuge der geschlechtlichen Vermehrung zur Bildung eines makroskopischen, mit dem bloßen Auge sichtbaren Fruchtkörpers aus sehr eng ineinander verflochtenen Zellfäden, der Ascokarp oder auch Ascoma genannt wird und beispielsweise bei Trüffeln als „Speisepilz“ verzehrt wird. Er besteht aus sterilen und fruchtbaren Hyphen. Letztere bilden die Sporen genannten Vermehrungszellen und sind oft in einer eigenen Fruchtschicht, dem Hymenium, zusammengefasst. Sie entwickeln sich meist auf der inneren Oberfläche des Ascokarps.
Ohne Bedeutung für die Systematik der Pilze klassifiziert man Ascokarpe unter anderem nach ihrer Lage: Liegt der Fruchtkörper wie beispielsweise bei den Morcheln (Morchella) oberhalb der Erde nennt man ihn epigäisch, befindet er sich wie etwa bei den Trüffeln (Tuber) unterirdisch, heißt er hypogäisch. Daneben unterscheidet man als Formtypen
- das Apothecium: Hier ist das Ascokarp becherartig nach oben geöffnet. Die Fruchtschicht (Hymenium) liegt frei, so dass zahlreiche Sporen gleichzeitig verbreitet werden können. Apothecien besitzen beispielsweise die Morcheln und Lorcheln.
- das Kleistothecium: In diesem Fall ist das Ascokarp kugelförmig und geschlossen, die Sporen können somit nicht selbständig freigesetzt werden. Dies hat zur Folge, dass Pilze mit Kleistothecien neue Strategien zur Verbreitung ihrer Sporen entwickeln mussten. Die Trüffel haben dieses Problem beispielsweise dadurch gelöst, dass sie Tiere wie beispielsweise Wildschweine anlocken, welche die schmackhaften Ascokarpe aufbrechen und die ihnen daraufhin anheftenden Sporen über größere Strecken transportieren können. Kleistothecien finden sich meist bei Pilzen, die nur sehr wenig Raum für ihr Ascokarp zur Verfügung haben, beispielsweise bei solchen, die unterhalb der Borkenschicht von Bäumen oder wie die Trüffel im Boden leben. Auch die Hautpilze der Gattung Arthroderma bilden Kleistothecien aus.
- das Perithecium: Perithecien haben die Form eines Kegels oder einer Kugel. Ihre Eigenart ist, dass sich an der Oberseite eine kleine Pore, die Ostiole, befindet, durch welche die Sporen bei Reife – im Gegensatz zu Apothecien einzeln hintereinander – entlassen werden. Perithecien finden sich beispielsweise bei den Holzkeulen (Xylaria) aber auch bei den Pustelpilzen (Nectria).
- das Pseudothecium: Wie bei den Perithecien werden die Sporen in Pseudothecien in einer nach außen geöffneten Höhlung gebildet. Hierbei werden die Fruchtkörper ascolocular gebildet, indem Fruchtkörperanlagen aus haploidem Mycel schon vor der Befruchtung entstehen und die dikaryotischen Hyphen in eine später entstehende Höhlung, den Loculus mit den Asci, hineinwachsen.[2] Der Unterschied besteht in der unten näher beschriebenen Art der Sporenfreisetzung: In Pseudothecien treten dazu so genannte bitunicate Sporenschläuche auf, die sich bei Wasseraufnahme ausdehnen und so die in ihnen befindlichen Sporen regelrecht aus der Öffnung herausschieben. Beispielarten sind der Apfelschorf (Venturia inaequalis) oder die Rosskastanien-Blattbräune (Guignardia aesculi).
Stoffwechsel
Wie die meisten Pilze zersetzen auch Schlauchpilze in erster Linie lebende oder tote Biomasse. Dazu geben sie an ihre Umgebung kräftige Verdauungsenzyme ab, die organische Substanzen in kleine Bruchstücke aufspalten, welche daraufhin durch die Zellwand aufgenommen werden. Viele Arten leben von abgestorbenem pflanzlichem Material wie abgefallenen Blättern, Zweigen oder auch ganzen Baumstämmen. Andere befallen als Parasiten Pflanzen, Tiere oder andere Pilze und gewinnen ihre Stoffwechselenergie sowie alle notwendigen Nährstoffe aus dem Zellgewebe ihres Wirts. Besonders in dieser Gruppe finden sich teilweise extreme Spezialisierungen; so befallen viele Arten nur ein bestimmtes Bein einer spezifischen Insektenart. Daneben finden sich Schlauchpilze allerdings auch häufig in symbiotischen Beziehungen. So bilden sie beispielsweise mit diversen Algenzellen oder Cyanobakterien, von denen sie energiereiche Photosynthese-Produkte erhalten, die Flechten – andere kooperieren als Mykorrhiza-Pilze mit Waldbäumen. Schließlich gibt es sogar fleischfressende Pilze, die mit ihren Hyphen Fallen entwickelt haben, in denen sich kleinere Protisten wie Amöben, aber auch Tiere wie Fadenwürmer (Nematoda), Rädertierchen (Rotifera), Bärtierchen (Tardigrada) oder sogar kleine Gliederfüßer wie die Springschwänze (Collembola) verfangen können.
Schlauchpilze haben im Verlauf ihrer langen Stammesgeschichte die Fähigkeit erlangt, nahezu jede organische Substanz abzubauen. So sind sie im Gegensatz zu fast allen anderen Organismen dazu in der Lage, pflanzliche Zellulose oder das im Holz enthaltene Lignin durch geeignete Enzyme zu zersetzen. Auch Kollagen, ein wichtiges tierisches Strukturprotein oder Keratin, aus dem zum Beispiel Haare bestehen, dienen Pilzen als „Nahrungsquelle“. Exotischere Beispiele liefern Aureobasidium pullulans, der Wandfarbe verstoffwechselt oder der Kerosinpilz (Amorphotheca resinae), der sich von Kerosin ernähren kann und infolgedessen oft die Treibstoffleitungen von Flugzeugen verstopft.
Verbreitung und Lebensraum
Schlauchpilze finden sich weltweit in allen landbasierten Ökosystemen, als Flechten treten sie sogar in der Antarktis auf. Die Verbreitung einzelner Arten ist dagegen sehr variabel: Manche finden sich auf allen Kontinenten, andere wie beispielsweise die kulinarisch hoch begehrten weißen Trüffel (Tuber magnatum) nur regional in isolierten Gegenden Italiens und Frankreichs.
Wie bereits erwähnt erlauben die zwischen den Zellen befindlichen Querwände oder Septen Schlauchpilzen die Besiedelung weitaus trockenerer Lebensräume als beispielsweise den Jochpilzen. Diese Austrocknungsresistenz ist bei einzelnen Arten, die auf gesalzenem Fisch wachsen, wegen des dort auftretenden gewaltigen osmotischen Drucks extrem ausgeprägt. Auf der anderen Seite ist eine (allerdings in der Minderheit befindliche) Gruppe ins Wasser zurückgekehrt.
Fortpflanzung
Die Fortpflanzung von Schlauchpilzen ist sehr vielfältig, sie kann sowohl in ungeschlechtlicher als auch in geschlechtlicher Weise geschehen. Letztere ist durch die dabei ausgebildeten Fortpflanzungsstrukturen, die schlauchförmigen Asci besonders charakteristisch. Auf der anderen Seite spielt die ungeschlechtliche Fortpflanzung bei weitem die größere Rolle; viele Arten haben die geschlechtliche Vermehrung auch ganz aufgegeben.
Grundsätzlich unterscheidet man bei Schlauchpilzen zwei fundamental verschiedene Zustände im Lebenszyklus, den Anamorph, das ungeschlechtliche Stadium, und den Teleomorph, das geschlechtliche Stadium, das allerdings nicht immer vorhanden ist. Die Gesamtheit aus Anamorph und Teleomorph wird manchmal auch als Holomorph bezeichnet.
Da Teleomorph und Anamorph sich oft äußerlich in keiner Weise ähneln, war bis ins späte 20. Jahrhundert eine Zuordnung der Stadien zueinander oft nicht möglich. Dies hat zu der kuriosen, aber bis heute taxonomisch akzeptierten Situation geführt, das zwei verschiedene Lebensstadien ein und desselben Pilzes verschiedenen „Arten“ zugeordnet werden und damit unterschiedliche Namen tragen. So ist die geschlechtliche Erscheinungsform des Kerosinpilzes zum Beispiel unter dem Namen Amorphotheca resinae bekannt, während Hormoconis resinae das ungeschlechtliche Stadium bezeichnet. Durch molekulargenetische Methoden werden heute allerdings immer mehr Verbindungen zwischen diesen „Arten“ aufgedeckt, so dass die frühere Einteilung der ungeschlechtlichen Stadien in eine eigene Abteilung, die Fungi imperfecti (Deuteromycota) heute obsolet geworden ist.
Ungeschlechtliche Fortpflanzung
Die ungeschlechtliche Fortpflanzung ist bei Schlauchpilzen die dominante Form der Vermehrung, die für die schnelle Ausbreitung der Pilze in einem noch nicht erschlossenen Gebiet verantwortlich ist. Sie erfolgt durch mit dem Mutterpilz genetisch identische, meist einkernige Verbreitungsstrukturen, die Konidiosporen, kurz Konidien oder nach der Art ihrer Entstehung durch den zellulären Prozess der Mitose auch Mitosporen genannt werden. Gebildet werden sie in der Regel durch spezielle, als konidiogen bezeichnete Zellen, die an der Spitze spezialisierter Hyphen, der Konidiophoren, sitzen. Je nach Art können sie durch den Wind, durch Wasser, aber auch durch Tiere verbreitet werden.
Koelomyceten und Hyphomyceten
Die Typenvielfalt der ungeschlechtlichen Stadien, der Anamorphe, ist enorm. Sie lassen sich (ohne Bedeutung für die Systematik) je nachdem, ob die Sporen in einer abgeschlossenen Struktur, einem Konidioma, gebildet werden oder nicht, grob in zwei Gruppen unterteilen, die Koelomyceten und die Hyphomyceten.
- Koelomyceten bilden ihre Sporen in nach außen hin abgeschlossenen Konidiomata, die sich oft knapp unterhalb der Oberfläche eines Wirtsorganismus bilden. Man unterscheidet
- acervulare Konidiomata oder Acervuli: Sie entwickeln sich im Wirtsorganismus und können dabei unterhalb der Cuticula genannten Außenschicht der Pflanze (subcuticular), innerhalb der äußersten Zellschicht, der Epidermis (intraepidermal), direkt darunter (subepidermal) oder auch unter zahlreichen Wirtszellschichten liegen. Meist entwickelt sich dort eine flache Schicht mit relativ kurzen Konidiophoren, die daraufhin massenhaft Sporen produzieren. Der dadurch immer stärker anwachsende Druck führt schließlich zum Aufbrechen der Epidermis und Cuticula und ermöglicht so ein Entweichen der Konidien.
- pyknidiale Konidiomata oder Pycnidia: Im Gegensatz zu den Acervuli bilden sich pycnidiale Konidiomata im Pilzgewebe selbst. Sie haben meist die Form einer bauchigen Vase; durch die an der Spitze gelegen kleine Öffnung, die Ostiole, werden die Sporen in ihre Umgebung entlassen.
- Bei den Hyphomyceten liegen die Konidiophoren, also die Hyphen, die an der Spitze konidienbildende Zellen tragen, immer frei. Sie treten meist isoliert, manchmal allerdings auch in Form parallel ausgerichteter Bündel oder kissenförmiger Massen auf; bündelförmige Konidiomata werden als synnematal, kissenförmige als sporodochial bezeichnet.
Sporen
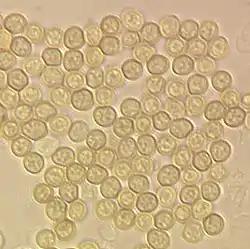 Konidiosporen von Trichoderma aggressivum, Durchmesser etwa 3 Mikrometer |
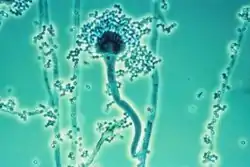 Konidiophoren von Schimmelpilzen der Gattung Aspergillus, Konidiogenese ist blastisch-phialidisch |
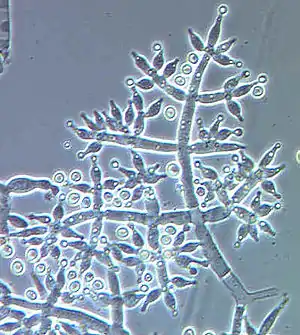 Konidiophoren von Trichoderma harzianum, Konidiogenese ist blastisch-phialidisch |
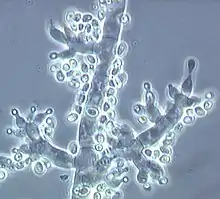 Konidiophoren von Trichoderma fertile mit vasenförmigen Phialiden und neu gebildeten Konidien an deren Spitzen (helle Punkte) |
Für die weitere Klassifikation der ungeschlechtlichen Lebenszustände der Schlauchpilze sind die Sporen wichtig, die nach Farbe, Form und Septierung unterschieden werden:
Den wohl häufigsten Sporentyp machen die als Amerosporen bezeichneten Einzelzellen aus. Ist dagegen die Spore durch eine Querwand (Septe) zweigeteilt, spricht man von Didymosporen. Bei zwei oder mehr Trennwänden kommt es auch auf die Form an. Sind die Septen innerhalb der Spore transversal, also wie die Sprossen einer Leiter angeordnet, nennt man sie Phragmosporen, bilden sie dagegen eher eine netzähnliche Struktur, spricht man von Dictyosporen. Bei manchen Sporen gehen von einem Zentralkörper strahlenförmige „Arme“ aus, sie heißen Staurosporen; bei anderen ist die ganze Spore wie eine Feder spiralig aufgewunden und wird dann Helicospore genannt. Schließlich bezeichnet man sehr lange, wurmförmige Sporen, die ein Verhältnis von Länge zu Durchmesser von mehr als 15:1 aufweisen, als Scolecosporen.
Konidiogenese und Dehiszenz
Zwei weitere wichtige Eigenschaften der Schlauchpilz-Anamorphen sind die Art, wie die Sporen gebildet werden, die Konidiogenese, und die Weise, wie sie sich abtrennen, die Dehiszens. Erstere entspricht der Embryologie bei Tieren und Pflanzen und lässt sich in zwei fundamentale Entwicklungsformen einteilen, die blastische Konidiogenese, bei der die Spore bereits wahrnehmbar ist, bevor sie durch eine Querwand von der sie bildenden konidiogenen Hyphe abgetrennt wird, und die thallische Konidiogenese, bei der sich erst die Querwand bildet und die so abgetrennte Zelle sich dann in eine Spore umwandelt.
Diese beiden Grundtypen lassen sich noch feiner untergliedern:
- Bei blastisch-acropetaler Sporenbildung entstehen die Konidien durch fortgesetzte Knospung an der Spitze der konidiogenen Hyphe, so dass sich eine lange Kette von Sporen bildet, deren jüngste immer an deren Spitze (apikal) liegt. Bilden sich zwei Knospen an einer Spore, die dann Ramoconidium genannt wird, so verzweigt sich die Kette. Blastisch-acropetale Konidiogenese ist somit eine Variante normalen Hyphenwachstums.
- Die blastisch-synchrone Konidiogenese zeichnet sich dadurch aus, dass sich zahlreiche Sporen gleichzeitig auf einer zentral gelegenen, meist aufgeschwollenen Zelle, bilden. Manchmal entstehen aus diesen Sporen sekundär wieder acropetale Ketten.
- Die blastisch-sympodiale Entwicklung ist dadurch charakterisiert, dass sich nach der Sporenbildung an der Spitze der Konidiophore hinter der Spore eine neue Spitze bildet, die dann seitlich (sympodial beziehungsweise zymös) hervorwächst. Diese bildet ihrerseits schließlich eine Spore, unterhalb derer wiederum eine neue Spitze entsteht usw. Unweigerlich werden bei diesem Prozess die konidiogenen Zellen immer länger.
- Besonders auffällig ist die blastisch-annellidische Form der Konidiogenese: Nachdem sich eine Spore gebildet und abgetrennt hat, bleibt eine ringförmige Narbe an der Spitze der konidiogenen Zelle zurück. Diese wächst durch diesen Ring hindurch und bildet dann die nächste Spore, die bei der Abtrennung ihrerseits eine Narbe hinterlässt, so dass man an der Zahl der Ringe einer konidiogenen Zelle die Zahl der von ihr gebildeten Sporen ablesen kann.
- Bei der blastisch-phialidischen Bildung der Sporen entstehen am offenen Ende der in diesem Fall Phialiden genannten konidiogenen Zellen immer neue Sporen. Da die Phialide selbst ihre Form und Länge nicht ändert, wächst die Sporenkette ähnlich wie bei der blastisch-acropetalen Entwicklung nach oben; im Gegensatz zu dieser ist hier aber die am so genannten konidiogenen Lokus gebildete unterste Spore immer die jüngste. Der Anamorph des Schimmelpilzes Penicillium ist ein bekanntes Beispiel für diese Konidiogeneseform.
- Als Variante der blastisch-phialidischen Entwicklungsform kann die basauxische Bildung gelten. Bei ihr befinden sich die Sporen der entstehenden Kette in unterschiedlichen Entwicklungsstadien: Während die am oberen Ende befindlichen ältesten Sporen ausgereift sind, haben sich die unten gelegenen jüngsten Sporen noch kaum von der darunter gelegenen Hyphe differenziert.
- Ganz anders verläuft die blastisch-retrogressive Sporenbildung. Hierbei bildet sich an der Spitze der konidiogenen Hyphe eine Spore und teilt sich durch eine Querwand von dieser ab. Daraufhin entwickelt sich direkt darunter eine weitere Spore, die sich nun ihrerseits durch eine Septe vom Rest der Hyphe trennt. Durch mehrfache Wiederholung dieses Prozesses bilden sich aus einer Hyphe von der Spitze nach unten immer weitere Sporen, während die Hyphenzelle selbst sich immer weiter verkürzt.
- Anders als bei den blastischen Formen werden bei thallisch-arthrischer Konidiogenese vor der Sporenbildung erst die Septen gebildet. Dazu hört eine Hyphe erst auf zu wachsen und bildet dann in irregulären Abständen doppelte Querwände aus. Die so entstandenen Einzelzellen trennen sich auf eine schizolytisch genannte Weise voneinander ab und beginnen sich zu differenzieren, so dass eine Kette kurzer zylinderförmiger Sporen, der so genannten Arthrokonidien, entsteht, die gelenkig miteinander verbunden zu sein scheinen. Bei einer Variante dieser Entwicklungsform bilden sich nur alternierende Zellen zu Sporen aus; die dazwischenliegenden Zellen degenerieren, sterben ab und setzen so die zwischen ihnen gelegenen Arthrokonidien frei.
- Schließlich existiert noch die thallisch-solitäre Entwicklungsform. Hier trennt sich am Ende einer konidiogenen Hyphe eine große bauchige Zelle ab, in der sich dann zahlreiche transversal angeordnete interne Querwände bilden. Die Zelle differenziert sich nun zu einer Phragmospore, die als ganzes (auf eine rhexolytisch genannte Weise) abgetrennt wird.
Die Dehiszens kann im Wesentlichen auf zwei unterschiedlichen Wegen geschehen. Bei der schizolytischen Variante bildet sich zwischen den Sporen eine Doppeltrennwand mit zentral gelegener Mittellamelle aus, die sich dann zur Abtrennung der Sporen auflöst. Bei rhexolytischer Dehiszens degeneriert dagegen einfach die Zellwand, welche die Sporen außen verbindet und setzt die Konidien auf diese Weise frei.
Heterokaryose und Parasexualität
Bei einer ganzen Reihe von Schlauchpilzen existieren entweder keine sexuellen Stadien oder solche sind nicht bekannt. Zwei Wege, trotzdem die genetische Vielfalt zu erhalten, sind Heterokaryose und Parasexualität. Erstere wird einfach durch Verschmelzung zweier Hyphen verschiedener Organismen ausgelöst, ein Prozess, der als Anastomose bezeichnet wird. Als Resultat befinden sich mehr Zellkerne im Myzel als gewöhnlich, die zudem von genetisch unterschiedlichen „Elternorganismen“ stammen. Parasexualität bezeichnet dagegen ein Phänomen, bei der es ohne eigentliche sexuelle Vorgänge zu einer Verschmelzung zweier Zellkerne und dadurch zu einer Verdoppelung der Chromosomenzahl kommt. Daran schließt sich eine komplexe Form der Mitose genannten Kernteilung an, bei der es zum „Crossing over“, einem Austausch von Genmaterial zwischen je zwei einander entsprechenden Chromosomen kommt. (Bei sexuellen Formen der Vermehrung tritt „Crossing over“ dagegen nur bei der Meiose genannten Reifeteilung auf.) Endlich entstehen durch Haploidisation wieder zwei Zellkerne mit je einem Chromosomensatz, die sich aber genetisch nun von den beiden Ausgangskernen unterscheiden.
Geschlechtliche Fortpflanzung

Die geschlechtliche Fortpflanzung der Schlauchpilze ist durch eine charakteristische Struktur geprägt, den Ascus, der sie von allen anderen Pilzen unterscheidet. Ein Ascus ist ein schlauchförmiges Meiosporangium, das bedeutet, dass in ihm durch die Meiose genannte Reifeteilung die geschlechtlichen Sporen gebildet werden, die dann zur Abgrenzung von den ungeschlechtlich gebildeten Konidiosporen auch Ascosporen heißen.
Von Ausnahmen wie der Bäckerhefe (Saccharomyces cerevisiae) abgesehen sind fast alle Schlauchpilze im Normalzustand haploid, ihre Kerne besitzen also jeweils nur einen Chromosomensatz, was sie besonders anfällig für Mutationen macht. Bei der geschlechtlichen Fortpflanzung kommt es zu einer in der Regel sehr kurzen diploiden Phase (mit dann zwei Chromosomensätzen), an die sich aber meist sehr schnell eine Meiose anschließt, so dass der haploide Zustand wiederhergestellt ist.
Ablauf der sexuellen Sporenbildung
Der sexuelle Teil des Lebenszyklus wird eingeleitet, sobald zwei passende Hyphen aufeinandertreffen. Diese stammen aus demselben Hyphengeflecht, das auch die ungeschlechtlichen Sporen ausbildet. Entscheidend dafür, ob eine Konjugation, also eine sexuelle Verschmelzung stattfindet, ist zunächst ob die Hyphen demselben Organismus angehören oder ob sie von einem anderen Pilz stammen. Während zahlreiche Pilze durchaus zur Selbstbefruchtung in der Lage sind, eine Eigenschaft, die als homothallisch bezeichnet wird, benötigen andere einen genetisch nicht-identischen Partner und sind damit heterothallisch. Daneben müssen die zwei betroffenen Hyphen auch noch passenden Paarungstypen angehören. Diese sind eine Besonderheit der Pilze und entsprechen in etwa den Geschlechtern bei Pflanzen und Tieren; es kann allerdings mehr als zwei Paarungstypen geben.
Bei Verträglichkeit bilden sich auf den Hyphen nun die Gametangien aus, die Bildungszellen für die Gameten, in denen sich zahlreiche Zellkerne sammeln. Durch eine sehr feine Hyphe, die Trichogyne, die aus einem der Gametangien, nun Ascogonium genannt, herauswächst, bildet sich eine Verbindung aus, durch die Zellkerne aus dem anderen Gametangium, nun Antheridium genannt, in das Ascogonium übertreten können. Anders als bei Tieren oder Pflanzen kommt es nach der Vereinigung der Zellplasmen der beiden beteiligten Gametangien (Plasmogamie) in der Regel nicht zu einer sofortigen Verschmelzung der Kerne (Karyogamie). Stattdessen ordnen sich die eingewanderten Kerne aus dem Antheridium paarweise mit Kernen des Ascogoniums an, bleiben aber unabhängig neben diesen bestehen. Damit hat die Dikaryophase im Lebenszyklus des Pilzes begonnen, während deren sich die Kernpaare wiederholt synchron teilen, so dass sich ihre Zahl vervielfacht. Aller Wahrscheinlichkeit nach ist die Dikaryophase eine evolutionäre Anpassung und dient dazu, bei einem seltenen Aufeinandertreffen zweier Individuen das Potenzial der sexuellen Fortpflanzung voll auszuschöpfen. Durch die wiederholte synchrone Teilung der Kernpaare vermehrt sich das genetische Ausgangsmaterial und wird bei der Rekombination während der Meiose in vielfältigster Weise miteinander kombiniert, so dass eine möglichst große Zahl genetisch unterschiedlicher Sporen entstehen kann. Eine ähnliche Lösung eines gleichgelagerten Problems hat sich unabhängig davon auch bei den Rotalgen (Rhodophyta) ausgebildet.
Aus dem befruchteten Ascogonium wachsen nun millionenfach neue, dinukleate Hyphen aus, in die jeweils pro Zelle zwei Zellkerne einwandern. Sie werden auch ascogen oder fertil genannt. Ernährt werden sie durch gewöhnliche, uni- oder mononukleate Hyphen mit nur einem Zellkern, die als steril bezeichnet werden. Das Geflecht aus sterilen und fertilen Hyphen bildet sich nun in vielen Fällen zum makroskopisch sichtbaren Fruchtkörper des Pilzes, dem Ascokarp aus, das mehrere Millionen fertiler Hyphen enthalten kann.
In der eigentlichen Fruchtschicht, dem Hymenium, entstehen nun die Asci. Dazu bildet eine ascogene Hyphe an der Spitze einen U-förmigen Haken aus, der entgegen der Wachstumsrichtung der Hyphe zeigt. Die in dieser endständigen, das heißt an der Spitze der Hyphe gelegenen Zelle enthaltenen zwei Kerne teilen sich nun derart, dass die Spindelfasern ihrer Mitosespindeln parallel verlaufen und dadurch jeweils zwei (genetisch verschiedene) Tochterkerne an der Spitze des Hakens zu liegen kommen, ein Tochterkern an der jetzt gegen die Wachstumsrichtung ausgerichteten Spitze der Hyphe und ein weiterer an der Basis der Hyphenzelle positioniert ist. Jetzt werden zwei parallele Querwände eingezogen, welche den Haken in drei Teile teilen: Die ursprüngliche, jetzt gegen die Wachstumsrichtung zeigende Spitze der Hyphe mit einem Kern, die daneben gelegene Basis der Hyphe, ebenfalls mit einem Zellkern und ein an der Spitze gelegener Mittelbereich mit zwei Zellkernen. In letzterem findet bei richtiger Positionierung in der Fruchtschicht endlich die eigentliche Kernverschmelzung statt, durch welche die diploide Zygote entsteht. Er verlängert sich jetzt zu einer langgezogenen schlauch- oder zylinderförmigen Kapsel, dem eigentlichen Ascus. Hier findet nun auch die Reifeteilung oder Meiose statt, durch die sich die Zahl der Kerne auf vier erhöht und gleichzeitig der haploide Zustand wiederhergestellt wird. Fast immer schließt sich eine weitere Kernteilung (Mitose) an, so dass am Ende acht Tochterkerne im Ascus zu liegen kommen. Diese werden nun mit etwas Zellplasma von eigenen Membranen und meist auch einer festen Zellwand umhüllt und bilden so schließlich die eigentlichen Verbreitungszellen, die Ascosporen. Diese liegen im Ascus zunächst wie Erbsen in einer Hülse, werden aber bei passender Gelegenheit freigesetzt.
Ascosporen sind grundsätzlich nicht begeißelt, so dass sie auf andere Verbreitungsmechanismen angewiesen sind. Manche Sporen werden durch den Wind ausgebreitet, bei anderen kommt es beim Kontakt mit Wasser zum Aufbrechen des reifen Ascus und zur Freisetzung der Sporen. Einzelne Arten haben regelrechte „Sporenkanonen“ entwickelt, mit denen die Sporen bis zu 30 cm weit geschleudert werden können. Vermutlich um ihre Sporen in die turbulente Luftzone oberhalb des Bodens zu verbringen, wird bei Pilzen der Gattung Ascobolus die Ausrichtung der Sporenkanone sogar durch den Lichteinfall bestimmt; sie haben hierzu ein regelrechtes Linsensystem entwickelt.
Sobald die Sporen auf einem geeigneten Substrat auftreffen, keimen sie aus, bilden neue Hyphen und beginnen so den Lebenszyklus von vorn.
Ascus-Klassifikation
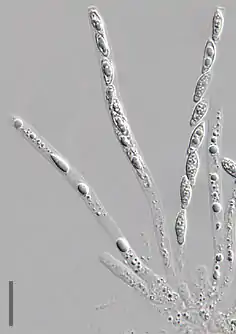
Bei der Klassifikation der Schlauchpilze spielt auch die Form des Ascus, also des schlauchförmigen Behälters, in dem die geschlechtlichen Sporen gebildet werden, eine Rolle. Es lassen sich hier grundsätzlich vier Typen unterscheiden:
- Ein unitunikat-operculater Ascus hat einen eingebauten „Deckel“, das Operculum, der über eine eingebaute Schwachstelle verfügt, an der er nach der Ausreifung der Sporen aufbricht und jene auf diese Weise freisetzt. Unitunikat-operculate Asci kommen nur in den Apothecien genannten Ascokarpen vor, also beispielsweise bei den Morcheln.
- Ein unitunikat-inoperculater Ascus besitzt dagegen kein Operculum. An dessen Stelle tritt ein elastischer Ring, der wie ein Druckventil funktioniert. Bei Reife dehnt er sich kurzzeitig aus und lässt so die Sporen hinausschießen. Dieser Typ findet sich sowohl in Apothecien als auch in Perithecien. Eine Beispielart ist der abgebildete Goldschimmel (Hypomyces chrysospermus).
- Ein bitunikater Ascus ist dadurch charakterisiert, dass er von einer Doppelwand umgeben ist. Diese setzt sich aus einer dünnen spröden Außenhülle und einer dicken elastischen Innenwand zusammen. Sobald die Sporen reif sind, spleißt die Hülle offen, so dass die Innenwand Wasser aufnehmen kann. Infolgedessen beginnt diese sich mitsamt der in ihr enthaltenen Sporen auszudehnen und zwar so lange, bis sie sich über den Rest des Ascokarps erhebt, so dass die Sporen unbehindert vom Fruchtkörper des Pilzes in den freien Luftstrom gelangen können. Bitunicate Asci treten ausschließlich in Pseudothekien auf und finden sich beispielsweise bei Apfelschorf (Venturia inaequalis) und Rosskastanien-Blattbräune (Guignardia aesculi).
- Gar keinen aktiven Freisetzungsmechanismus haben die meist kugelförmig gebauten prototunikaten Asci. Bei ihnen löst sich die reife Ascuswand entweder einfach auf, so dass die Sporen herausquellen können, oder sie wird durch äußere Einflüsse wie beispielsweise Tiere aufgebrochen. Prototunikate Asci finden sich sowohl in Perithekien als auch in Kleistothekien, beispielsweise bei den Bläuepilzen (Ophiostoma). Da es sich eher um eine Sammelbezeichnung für nicht zu den anderen drei Typen gehörige Asci handelt, gilt es als wahrscheinlich, dass sie sich mehrfach unabhängig aus unitunikaten Asci entwickelt haben.
Ökologie
Schlauchpilze übernehmen zentrale Rollen in den meisten landbasierten Ökosystemen. Sie sind wichtige Zersetzer organischer Materialien wie toter Blätter, Zweige, abgestorbener Bäume etc. und erleichtern zudem detrivoren, also von organischen Abfallstoffen lebenden Organismen die Aufnahme ihrer Nahrung. Durch das Aufbrechen sonst nur schwer abbaubarer Substanzen wie Zellulose oder Lignin nehmen sie einen wichtigen Platz im Stickstoff- und Kohlenstoffkreislauf der Natur ein.
Umgekehrt bilden die Fruchtkörper der Schlauchpilze selbst die Nahrung der verschiedensten Tiere, von Insekten (Insecta) und Schnecken (Gastropoda) hin zu Nagetieren (Rodentia) und größeren Säugetieren wie Rehen oder Wildschweinen.
Daneben sind Schlauchpilze für ihre zahlreichen symbiotischen Beziehungen zu anderen Lebewesen bekannt.
Flechten
Wahrscheinlich schon früh in ihrer Stammesgeschichte haben Schlauchpilze Grün- (Chlorophyta) sowie in Einzelfällen andere Algen und Cyanobakterien (Cyanobakteria) „domestiziert“ und bilden mit diesen als Flechten bekannte Lebensgemeinschaften, die in den unwirtlichsten Gegenden der Erde, in der Arktis und Antarktis, in Wüsten oder Hochgebirgen leben und Temperaturextreme von −40 Grad Celsius bis +80 Grad Celsius überstehen können. Während der photoautotrophe Algenpartner durch Photosynthese die Stoffwechselenergie bereitstellt, bietet der Pilz ein stabiles Stützskelett und schützt vor Strahlungseinwirkung und Austrocknung. Etwa 42 Prozent oder umgerechnet 18.000 aller Schlauchpilzarten sind flechtenbildend; umgekehrt sind die Pilzpartner fast aller Flechten Schlauchpilze – der Anteil der Ständerpilze liegt wahrscheinlich bei nicht mehr als zwei bis drei Prozent.
Mykorrhizapilze und Endophyten
Mit Pflanzen bilden Schlauchpilze zwei besonders wichtige Lebensgemeinschaften, als Mykorrhizapilze und als Endophyten. Erstere sind symbiotische Assoziationen der Pilze mit dem Wurzelsystem der Pflanzen, die für Bäume, insbesondere Nadelbäume, lebensnotwendig sind und ihnen erst den Aufschluss von Mineralsalzen aus dem Boden ermöglichen. Während der Pilzpartner durch sein fein verzweigtes Myzel viel eher als die Pflanze zur Aufnahme von Mineralen in der Lage ist, wird er von der Pflanze mit Stoffwechselenergie in Form von Photosyntheseprodukten versorgt. Es ist sogar bekannt, dass Mykorrhizapilze in der Lage sind, durch ihr weit gespanntes Hyphengeflecht Nährstoffe von einer Pflanze zu einer anderen zu transportieren, um diese zu stabilisieren. Als sehr wahrscheinlich gilt, dass erst die Assoziation zwischen Mykorrhizapilzen und den Wurzeln der Pflanzen letzteren die Eroberung des Landes ermöglicht hat. Jedenfalls sind schon die ersten erhaltenen Fossilien von Landpflanzen mit Mykorrhizapilzen assoziiert.
Endophyten leben dagegen in der Pflanze, insbesondere im Stamm und in den Blättern, schädigen diese dabei allerdings in der Regel nicht. Die genaue Natur der Beziehung zwischen Pilz und Wirtspflanze ist noch nicht gut verstanden, es scheint allerdings, dass Endophytenbesiedelung einer Pflanze höhere Widerstandskraft gegen Schadinsekten, Fadenwürmer und Bakterien verleiht sowie die Produktion spezieller Alkaloide, giftiger Pflanzenstoffe, die auf pflanzenfressende Säugetiere gesundheitsschädigend wirken, heraufsetzt oder erst ermöglicht. Man unterscheidet zwischen nur punktuell auftretenden und systemischen Endophyten; letztere kommen überall in der Pflanze vor.
Symbiotische Beziehungen zu Tieren
Eine Reihe von Arten aus der Schlauchpilz-Gattung Xylaria findet sich in den Nestern von Blattschneiderameisen und verwandten Arten aus dem Tribus Attini und in den Pilzgärten der Termiten (Isoptera). Da sie erst Fruchtkörper ausbilden, wenn die Ameisen die Nester verlassen haben oder im Absterben begriffen sind, wurde zwischenzeitlich in Betracht gezogen, dass sie wie eine Reihe von Ständerpilzen (Basidiomycota) von den Insekten kultiviert werden. Dies konnte bisher nicht bestätigt werden.
Wichtige Symbiosepartner sind hingegen die Borkenkäfer (Scolytidae). Die Käferweibchen transportieren die Pilzsporen in charakteristischen Einstülpungen ihrer Außenhaut, den Mycetangien, zu neuen Pflanzen. Dort fressen sie Gänge in das Holz, die sich nach innen zu größeren Kammern weiten, in denen sie ihre Eier ablegen. Gleichzeitig werden hier auch die Sporen freigesetzt. Von den entstehenden Pilzhyphen, die anders als die Käfer das Holz enzymatisch abbauen können, ernähren sich wiederum die geschlüpften Larven, die sich nach ihrer Verpuppung und Umwandlung zum erwachsenen Insekt mit Pilzsporen kontaminiert auf die Suche nach neuen Pflanzen machen.
Bedeutung für den Menschen

Schlauchpilze tragen erheblich sowohl zum Nutzen als auch zum Schaden des Menschen bei.
Schadwirkungen
Eine der bedeutendsten Schadwirkungen ist ihre Funktion als Erreger von zahlreichen Pflanzenkrankheiten. Ein großflächiges Ulmensterben wurde zum Beispiel in Nordamerika und Europa durch die eng verwandten Arten Ophiostoma ulmi und Ophiostoma novo-ulmi ausgelöst, für den Befall von Esskastanien (Castanea sativa) kann die aus Asien stammende Art Cryphonectria parasitica verantwortlich gemacht werden, eine besonders in Nordamerika grassierende Krankheit der Maispflanzen (Zea mays) wird durch Cochliobolus heterostrophus hervorgerufen und Taphrina deformans greift Pfirsichblätter an. Uncinula necator ist für den Befall von Weinreben mit echtem Mehltau verantwortlich, während Schlauchpilze der Gattung Monilia insbesondere Stein- und Kernobst befallen und dort die so genannte Polsterfäule auslösen; daneben werden aber auch die Blütenstände von Pfirsich (Prunus persica) und Sauerkirsche (Prunus ceranus) infiziert.
Schlauchpilze wie Stachybotrys chartarum sind für die Ausbleichung von Baumwolltextilien verantwortlich und stellen insbesondere in den Tropen ein großes Problem dar. Blaugrüner, roter und brauner Schimmel befällt und verdirbt hingegen Lebensmittel, Penicillium italicum zum Beispiel Orangen.
Mit Fusarium graminearum infiziertes Getreide enthält Mykotoxine wie Deoxynivalenol, die bei Verzehr durch Schweine bei diesen zu Haut- oder Schleimhautläsionen führen. Direkt für den Menschen bedeutsam ist der Mutterkornpilz (Claviceps purpurea), der Roggen oder Weizen befällt und hochgiftige, eventuell sogar karzinogene Alkaloide bildet, die bei Einnahme unter anderem zu Darmkrämpfen und schweren Halluzinationen führen können. Hochgradig krebserzeugend und leberschädigend sind dagegen die von Aspergillus flavus, einem unter anderem auf Erdnüssen wachsenden Schlauchpilz, gebildeten Aflatoxine. Im Vergleich dazu eher harmlos ist Candida albicans, ein Hefepilz, der menschliche Schleimhäute befallen kann und dort Kandidosen auslöst. Auch die für Pilzinfektionen der Haut verantwortlichen Hautpilze der Gattung Epidermophyton sind für Menschen mit gesundem Immunsystem eher ungefährlich. Ist dieses jedoch gestört, so kann es zu lebensgefährlichen Erkrankungen kommen; Pneumocystis jirovecii ist beispielsweise für schwere Lungenentzündungen verantwortlich, wie sie beispielsweise bei AIDS-Patienten auftreten.
Nutzwirkungen
Andererseits haben Schlauchpilze dem Menschen auch bedeutenden Nutzen gebracht. Am berühmtesten ist wohl die Art Penicillium chrysogenum, deren vermutlich zur Bekämpfung konkurrierender Bakterien entwickeltes Antibiotikum als Penizillin im 20. Jahrhundert eine Revolution in der Behandlung bakterieller Infektionskrankheiten ausgelöst hat. Doch auch die Bedeutung von Tolypocladium niveum, ein Schlauchpilz der mit Cyclosporin eines der effektivsten und schonendsten Mittel zur Immunsuppression produziert, lässt sich kaum überschätzen. Neben dem Einsatz bei Organtransplantationen zur Verhinderung der Abstoßung körperfremden Gewebes wird Cyclosporin auch zur Behandlung von Autoimmunerkrankungen wie zum Beispiel der Multiplen Sklerose in Betracht gezogen, allerdings bestehen hier Zweifel über die langfristige Verträglichkeit des Wirkstoffs.
Daneben können manche Schlauchpilze verhältnismäßig leicht durch gentechnologische Verfahren genetisch verändert werden. Sie produzieren dann wichtige Proteine wie Insulin, menschliche Wachstumsfaktoren oder tPA, einen zur Auflösung von Blutgerinnseln eingesetzten Wirkstoff. Der Brotschimmel Neurospora crassa ist ein wichtiger Modellorganismus der Biologie; sein Genom wurde mittlerweile vollständig sequenziert.
Auf profanerer Ebene werden Schlauchpilze zur Produktion von Lebensmitteln eingesetzt: Die Backhefe (Saccharomyces cerevisiae) kommt beim Backen mit Hefeteig und bei der Herstellung von Bier und Wein zum Einsatz, indem sie Zucker wie Glukose oder Saccharose zu Alkohol vergärt und Kohlendioxid freisetzt, das dazu dient, den Teig zu lockern. Enzyme von Penicillium camembertii spielen bei der Produktion der Käsesorten Camembert und Brie, solche von Penicillium roqueforti bei der Herstellung von Gorgonzola, Roquefort und Blue Stilton eine Rolle. Aspergillus oryzae wird insbesondere in Asien einem Brei aus eingeweichten Sojabohnen zugesetzt; durch die Fermentation entsteht die in vielen fernöstlichen Gerichten eingesetzte Sojasauce.
Schließlich sind einige Schlauchpilze auch begehrte Speisepilze: Mögen die Ständerpilze (Basidiomycetes) in dieser Hinsicht zahlenmäßig auch bedeutender sein; die Schlauchpilze stellen mit Morcheln (Morchella) und Trüffeln (Tuber) die beiden kulinarisch herausragendsten Pilz-Delikatessen.
Gefährdung

Informationen über die globale Gefährdungssituation der Schlauchpilze fehlen weitgehend. Anders als etwa bei Säugetieren oder Vögeln sind die Bestände in der Regel viel zu wenig bekannt, um genaue Aussagen über die globale Bedrohung von Schlauchpilzen treffen zu können. Dies gilt insbesondere für die zahlreichen, oft nur mikroskopisch sichtbaren Mykorrhiza-Pilze, die dennoch wichtige ökologische Aufgaben übernehmen, doch auch bei Großpilzen sind die vorliegenden Informationen noch sehr spärlich. Erst seit 2003 sind auf der Roten Liste auch Pilze aufgeführt, im Jahr 2004 werden dort allerdings nur zwei nordamerikanische Flechtenarten, Cladonia perforata und Erioderma pedicellatum gelistet, die als einzige bisher evaluiert wurden – die Aussagekraft der Liste ist daher bisher vernachlässigbar.
Auf nationaler Ebene wurde in Deutschland erstmals 1992 eine Rote Liste der gefährdeten Großpilze erstellt. Auf der 1996 herausgegebenen Nachfolgerliste werden etwa 55 % der bekannten Flechtenarten (in denen Schlauchpilze die weitaus überwiegende Zahl der Pilzpartner stellen) als gefährdet eingeschätzt, daneben werden auch eine ganze Reihe von Großpilzen wie der Leuchtende Prachtbecher (Caloscypha fulgens) oder die Riesenlorchel (Gyromitra gigas) gelistet. In der Schweiz gibt es seit 1995 eine „Provisorische Rote Liste der gefährdeten Höheren Pilze der Schweiz“, auf der ebenfalls einige Schlauchpilzarten aufgeführt sind.
Stammesgeschichte
Schlauchpilze sind sehr wahrscheinlich im Meer entstanden. Wann sie sich von ihrer evolutionären Schwestergruppe, den Ständerpilzen, getrennt haben, lässt sich zu Beginn des 21. Jahrhunderts nicht genau beantworten. Molekularbiologische Befunde verweisen diesen Zeitpunkt in das Erdzeitalter des späten Proterozoikums vor etwa 600 Millionen Jahren. Obwohl bereits aus dem Silur Fossilien bekannt sind, die als Schlauchpilze interpretiert werden können, ist ihre Zuordnung zu dieser Gruppe heute umstritten.

Zweifelsfrei nachgewiesen sind Schlauchpilze aus dem unteren Devon vor etwa 400 Millionen Jahren. In einer berühmten Fundstätte, dem Rhynie Chert im schottischen Aberdeenshire, wurden in Dünnschnitten unterhalb der Epidermis von Blättern, Sprossen und Rhizomen der frühen Landpflanze Asteroxylon Asci, Ascosporen und perithekienförmige Ascokarpe identifiziert, die sich eindeutig mit Schlauchpilzen in Verbindung bringen lassen. Die Wechselwirkungen zwischen Pilz und Pflanze sind damit schon sehr früh entstanden; es wird sogar spekuliert, dass die enge symbiotische Beziehung zwischen Pilzen und Pflanzen eine Voraussetzung für die Besiedelung des Landes gewesen sein könnte. Interessanterweise sind auch die ältesten Flechten aus den Fundstätten bei Rhynie bekannt.
Systematik

Schlauchpilze bilden mit sehr hoher Wahrscheinlichkeit ein monophyletisches Taxon, umfassen also alle Nachkommen eines gemeinsamen Vorfahren. Dies erscheint vor allem aufgrund der Synapomorphie des Ascus, der nur bei Schlauchpilzen auftritt, als nahezu sicher. Bei den Pilzen ohne geschlechtliches Stadium ist oft aufgrund eindeutiger molekularbiologischer oder ultrastruktureller Daten eine Einordnung in die Schlauchpilze möglich.
Die Zusammenfassung mit den Ständerpilzen (Basidiomycota) zu einer Dikarya genannten Gruppe wird durch die bei beiden Gruppen vorhandene regelmäßige Septierung durch Querwände mit zentraler Perforation, die ihnen gemeinsame zweikernige (dikaryotische) Phase im Lebenszyklus nach der Verschmelzung der Zellplasmen und die bei beiden auftretende, im Abschnitt zur Heterokaryose angesprochene Möglichkeit der Fusion steriler Hyphen (Anastomose) begründet. Die an der Spitze ascogener Zellen kurz vor der Ascusbildung auftretenden Haken und die bei Ständerpilzen vorzufindenden Schnallen (clamp connections) werden als Homologien gesehen, also als Strukturen, die auf eine gemeinsame Vorläuferstruktur zurückgehen.
Die Schlauchpilze bilden mit einem Anteil an der Gesamtzahl aller echten Pilze von 75 % eine sehr artenreiche Gruppe; mehr als 32.000 Arten wurden bisher beschrieben; die tatsächliche Anzahl dürfte allerdings insbesondere in Anbetracht der bisher kaum untersuchten Pilzflora tropischer Gebiete um ein Mehrfaches darüber liegen. Es lassen sich innerhalb des Taxons drei große Gruppen unterscheiden, die als Unterabteilungen klassifiziert werden[3]:
- Die Pezizomycotina (auch Euascomycetes) bilden die größte Klasse. Bei ihnen entwickeln sich die Asci aus ascogenen Hyphen, die fast immer in Ascokarpen vereinigt sind. Die Ascosporenmembran bildet sich in dieser Klasse nicht aus der Membran des Zellkerns, sondern aus der außenliegenden Plasmamembran. Zu den Euascomycetes gehören die meisten Schimmelpilze wie auch die medizinisch so bedeutsame Art Penicillium chrysogenum, zahllose flechtenbildende Pilze, aber auch die Trüffeln und Morcheln.
- Die Saccharomycotina (Hemiascomycetes) sind sehr einfach gebaute Schlauchpilze, die nur kurze oder gar keine Hyphengeflechte ausbilden. In dieser Gruppe finden sich keine Ascokarpe; die Asci entstehen vielmehr direkt, ohne den Umweg über spezialisierte ascogene Zellen: Nach der Konjugation passender haploider Zellen und der Verschmelzung zur Zygote wandelt sich diese selbst in einen Ascus um, in dem sich dann vier tetraederförmig angeordnete Ascosporen bilden, die durch Sprossung auskeimen. Im Gegensatz zu den Pezizomycotina stammt die Sporenmembram von der Membran des Sporenkerns ab. Zu den Saccharomycotina zählen zahlreiche Hefepilze, darunter die Backhefe (Saccharomyces cerevisiae), aber auch die für Schleimhautinfektionen verantwortliche Art Candida albicans. Es gilt als weitgehend unumstritten, dass die Saccharomycotina eine natürliche Gruppe bilden.
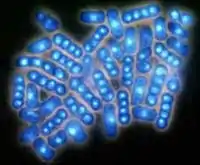 Hefezellen bei der Teilung
Hefezellen bei der Teilung - Die Taphrinomycotina (Archaeoascomycetes) schließlich sind eine erst zu Ende des 20. Jahrhunderts etablierte relativ kleine Gruppe von Arten, die klassisch zu den Hemiascomyceten gezählt wurden. Sie haben sich wahrscheinlich schon früh von allen anderen Schlauchpilzen getrennt. Die Gruppe gilt heute als monophyletisch[3], auch wenn sie recht vielfältig ist. Auch Taphrinomycotina bilden keine Ascomata aus.
Gliederung
Wijayawardene und Autoren (2020) gliedern die Abteilung Ascomycota wie folgt:[4] Es wird bis zur Unterklasse bzw. bis zur Ordnung angezeigt, siehe auch Systematik der Pilze:
- Unterabteilung Taphrinomycotina
- Klasse Taphrinomycetes
- Ordnung Taphrinales
- Klasse Neolectomycetes
- Ordnung Schlauchkeulenartige (Neolectales)
- Klasse Pneumocystidomycetes
- Ordnung Pneumocystidales
- Klasse Schizosaccharomycetes
- Ordnung Schizosaccharomycetales
- Klasse Archaeorhizomycetes
- Klasse Taphrinomycetes
- Unterabteilung Saccharomycotina
- Klasse Saccharomycetes
- Ordnung Saccharomycetales
- Klasse Saccharomycetes
- Unterabteilung Echte Schlauchpilze – Pezizomycotina
- Klasse Arthoniomycetes
- Ordnung Arthoniales
- Ordnung Lichenostigmatales
- Klasse Coniocybomycetes
- Ordnung Coniocybales
- Klasse Dothideomycetes
- Unterklasse Dothideomycetidae
- Unterklasse Pleosporomycetidae
- keiner Unterklasse zugeordnet – incertae sedis
- Ordnung Abrothallales
- Ordnung Acrospermales
- Ordnung Aulographales
- Ordnung Botryosphaeriales
- Ordnung Catinellales
- Ordnung Collemopsidiales
- Ordnung Dyfrolomycetales
- Ordnung Eremithallales
- Ordnung Jahnulales
- Ordnung Kirschsteiniotheliales
- Ordnung Lembosinales
- Ordnung Lichenotheliales
- Ordnung Minutisphaerales
- Ordnung Monoblastiales
- Ordnung Murramarangomycetales
- Ordnung Muyocopronales
- Ordnung Natipusillales
- Ordnung Parmulariales
- Ordnung Stigmatodiscales
- Ordnung Strigulales
- Ordnung Superstratomycetales
- Ordnung Tubeufiales
- Ordnung Valsariales
- Ordnung Zeloasperisporiales
- Klasse Eurotiomycetes
- Unterklasse Chaetothyriomycetidae
- Unterklasse Eurotiomycetidae
- Unterklasse Coryneliomycetidae
- Unterklasse Mycocaliciomycetidae
- Klasse Geoglossomycetes
- Ordnung Erdzungenartige – Geoglossales
- Klasse Laboulbeniomycetes
- Ordnung Laboulbeniales
- Ordnung Pyxidiophorales
- Klasse Lecanoromycetes
- Unterklasse Acarosporomycetidae
- Unterklasse Candelariomycetidae
- Unterklasse Lecanoromycetidae
- Unterklasse Ostropomycetidae
- Unterklasse Umbilicariomycetidae
- keiner Unterklasse zugeordnet – incertae sedis
- Ordnung Micropeltidales
- Ordnung Turquoiseomycetales
- Klasse Leotiomycetes
- Ordnung Golfkugelpilzartige – Cyttariales
- Ordnung Mehltaupilzartige – Erysiphales
- Ordnung Helotiales
- Ordnung Lahmiales
- Ordnung Leotiales
- Ordnung Medeolariales
- Ordnung Phacidiales
- Ordnung Runzelschorfartige – Rhytismatales
- Ordnung Thelebolales
- Ordnung Triblidiales
- Klasse Lichinomycetes
- Ordnung Lichinales
- Klasse Orbiliomycetes
- Ordnung Knopfbecherchenartige – Orbiliales
- Klasse Pezizomycetes
- Ordnung Becherlingsartige – Pezizales
- Klasse Sordariomycetes
- Unterklasse Diaporthomycetidae
- Unterklasse Lulworthiomycetidae
- Unterklasse Savoryellomycetidae<
- Unterklasse Hypocreomycetidae
- Unterklasse Sordariomycetidae
- Unterklasse Xylariomycetidae
- keiner Unterklasse zugeordnet – incertae sedis
- Ordnung Amplistromatales
- Ordnung Spathulosporales
- keiner Unterklasse zugeordnet – incertae sedis
- Klasse Xylonomycetes
- Ordnung Xylonomycetales
- Klasse Arthoniomycetes
Forschung
Schlauchpilze sind bis heute Gegenstand aktiver Forschungsarbeiten. Ein wichtiges Forschungsgebiet ist immer noch die Systematik; die Stellung der Schlauchpilze als Schwestergruppe der Ständerpilze gilt zwar als relativ unumstritten, Fragen nach der weiteren Einteilung sind aber nach wie vor aktuell. Insbesondere das Verhältnis der Archaeoascomycetes zu den restlichen Schlauchpilzen ist noch nicht aufgeklärt; auch die detailliertere Klassifikation der Schlauchpilz-Ordnungen ist noch im Fluss.
Ein anderes fruchtbares Forschungsgebiet ist die Untersuchung der Stammesgeschichte der Schlauchpilze anhand fossiler Sporen, Asci oder ganzer Ascomata. Insbesondere die Assoziation von Schlauchpilzen mit den Wurzeln der ersten Landpflanzen ist bei weitem noch nicht aufgeklärt; weitere Fossilienfunde gelten als dringend notwendig, um die Vorgänge bei der Landbesiedelung von Pflanzen und Pilzen besser zu verstehen.
Die Rolle der Schlauchpilze in natürlichen Ökosystemen ist ein weiteres aktives Gebiet mykologischer Studien. Während die Flechtenkunde (Lichenologie) sich traditionell als selbständige Wissenschaft versteht, fällt die Untersuchung von Mykorrhiza-Symbiosen und endophytischen Pilzen in das Arbeitsgebiet des Pilzforschers oder Mykologen. Gerade die Endophyten sind in ihrer Auswirkung auf die jeweiligen Wirtspflanzen erst sehr wenig erforscht.
Einige Pilze dienen auch als Modellorganismen in Genetik und Molekularbiologie. Unter anderem an Neurospora crassa und der Backhefe Saccharomyces cerevisiae werden zahlreiche zellbiologische Phänomene erforscht, die von grundsätzlicher Bedeutung für das Verständnis der allen Organismen zugrundeliegenden Lebensprozesse sind.
Immer mehr in den Vordergrund rückt die Frage des Artenschutzes: Die Gefährdung einer Art lässt sich dann zuverlässig beurteilen, wenn seriöse Daten zur Bestandsentwicklung vorliegen – diese existieren jedoch selbst in den gemäßigten Breiten nur in sehr wenigen Ausnahmefällen. Wahrscheinlich ist zu Beginn des 21. Jahrhunderts nur ein sehr geringer Teil der Vielfalt der Schlauchpilzearten erfasst.
Quellen
Literatur
- C. J. Alexopoulos, M. Blackwell, C. W. Mims: Introductory Mycology. 4. Auflage. Wiley, New York NY 1996, ISBN 0-471-52229-5.
- B. Kendrick: The Fifth Kingdom. Kapitel 4. Focus Publ., Newburyport MA 2001, ISBN 1-58510-022-6.
- G. J. Krieglsteiner: Verbreitungsatlas der Großpilze Deutschlands (West). Bd. 2. Schlauchpilze. Ulmer, Stuttgart 1993, ISBN 3-8001-3318-0.
- Josef Breitenbach, Fred Kränzlin (Hrsg.): Pilze der Schweiz. Beitrag zur Kenntnis der Pilzflora der Schweiz. Band 1: Ascomyceten (Schlauchpilze). Mykologia, Luzern 1981, ISBN 3-85604-010-2.
Einzelnachweise
- Index Fungorum (Memento vom 13. März 2012 im Internet Archive)
- Definition von ascolocular im Lexikon der Biologie bei Spektrum.de, abgerufen am 12. Mai 2018.
- David S. Hibbett, Manfred Binder, Joseph F. Bischoff, Meredith Blackwell, Paul F. Cannon, Ove E. Eriksson, Sabine Huhndorf, Timothy James, Paul M. Kirk, Robert Lücking, H. Thorsten Lumbsch, François Lutzoni, P. Brandon Matheny, David J. McLaughlin, Martha J. Powell, Scott Redhead, Conrad L. Schoch, Joseph W. Spatafora, Joost A. Stalpers, Rytas Vilgalys, M. Catherine Aime, André Aptroot, Robert Bauer, Dominik Begerow, Gerald L. Benny, Lisa A. Castlebury, Pedro W. Crous, Yu-Cheng Dai, Walter Gams, David M. Geiser, Gareth W. Griffith, Cécile Gueidan, David L. Hawksworth, Geir Hestmark, Kentaro Hosaka, Richard A. Humber, Kevin D. Hyde, Joseph E. Ironside, Urmas Kõljalg, Cletus P. Kurtzman, Karl-Henrik Larsson, Robert Lichtwardt, Joyce Longcore, Jolanta Miądlikowsk, Andrew Miller, Jean-Marc Moncalvo, Sharon Mozley-Standridge, Franz Oberwinkler, Erast Parmasto, Valérie Reeb, Jack D. Rogers, Claude Roux, Leif Ryvarden, José Paulo Sampaio, Arthur Schüßler, Junta Sugiyama, R. Greg Thorn, Leif Tibell, Wendy A. Untereiner, Christopher Walker, Zheng Wang, Alex Weir, Michael Weiß, Merlin M. White, Katarina Wink, Yi-Jian Yao, Ning Zhang: A higher-level phylogenetic classification of the Fungi. In: Mycological Research. Band 111(5). British Mycological Society, 2007, S. 509–547, doi:10.1016/j.mycres.2007.03.004 (PDF; 1,01 MB (Memento vom 22. November 2010 im Internet Archive)). A higher-level phylogenetic classification of the Fungi (Memento vom 22. November 2010 im Internet Archive)
- Wijayawardene, N. N., K. D. Hyde, L. K. T. Al-Ani, L. Tedersoo, Danny Haelewaters, K. C. Rajeshkumar, R. L. Zhao, A. Aptroot, V Leontyev, D., R. K. Saxena, Y. S. Tokarev, D. Q. Dai, P. M. Letcher, S. L. Stephenson, D. Ertz, H. T. Lumbsch, M. Kukwa, V Issi, I., H. Madrid, A. J. L. Phillips, L. Selbmann, W. P. Pfliegler, E. Horvath, K. Bensch, P. M. Kirk, K. Kolarikova, H. A. Raja, R. Radek, V Papp, B. Dima, J. Ma, E. Malosso, S. Takamatsu, G. Rambold, P. B. Gannibal, D. Triebel, A. K. Gautam, S. Avasthi, S. Suetrong, E. Timdal, S. C. Fryar, G. Delgado, M. Reblova, M. Doilom, S. Dolatabadi, J. Z. Pawlowska, R. A. Humber, R. Kodsueb, I Sanchez-Castro, B. T. Goto, D. K. A. Silva, F. A. de Souza, F. R. Oehl, G. A. da Silva, I. R. Silva, J. Blaszkowski, K. Jobim, L. C. Maia, F. R. Barbosa, P. O. Fiuza, P. K. Divakar, B. D. Shenoy, R. F. Castaneda-Ruiz, S. Somrithipol, A. A. Lateef, S. C. Karunarathna, S. Tibpromma, P. E. Mortimer, D. N. Wanasinghe, R. Phookamsak, J. Xu, Y. Wang, F. Tian, P. Alvarado, D. W. Li, I Kusan, N. Matocec, A. Masic, Z. Tkalcec, S. S. N. Maharachchikumbura, M. Papizadeh, G. Heredia, F. Wartchow, M. Bakhshi, E. Boehm, N. Youssef, V. P. Hustad, J. D. Lawrey, A. L. C. M. A. Santiago, J. D. P. Bezerra, C. M. Souza-Motta, A. L. Firmino, Q. Tian, J. Houbraken, S. Hongsanan, K. Tanaka, A. J. Dissanayake, J. S. Monteiro, H. P. Grossart, A. Suija, G. Weerakoon, J. Etayo, A. Tsurykau, V. Vazquez, P. Mungai, U. Damm, Q. R. Li, H. Zhang, S. Boonmee, Y. Z. Lu, A. G. Becerra, B. Kendrick, F. Q. Brearley, J. Motiejunaite, B. Sharma, R. Khare, S. Gaikwad, D. S. A. Wijesundara, L. Z. Tang, M. Q. He, A. Flakus, P. Rodriguez-Flakus, M. P. Zhurbenko, E. H. C. McKenzie, M. Stadler, D. J. Bhat, J. K. Liu, M. Raza, R. Jeewon, E. S. Nassonova, M. Prieto, R. G. U. Jayalal, M. Erdogdu, A. Yurkov, M. Schnittler, O. N. Shchepin, Y. K. Novozhilov, A. G. S. Silva-Filho, E. Gentekaki, P. Liu, J. C. Cavender, Y. Kang, S. Mohammad, L. F. Zhang, R. F. Xu, Y. M. Li, M. C. Dayarathne, A. H. Ekanayaka, T. C. Wen, C. Y. Deng, O. L. Pereira, S. Navathe, D. L. Hawksworth, X. L. Fan, L. S. Dissanayake, E. Kuhnert, and M. Thines.: Outline of Fungi and Fungus-like Taxa. In: Mycosphere. Band 11, Nr. 1, 2020, S. 1060–1456., doi:10.5943/mycosphere/11/1/8.
Weblinks
- Anamorph-Teleomorph-Datenbank
- Fotografien einiger Fruchtkörper (Ascomata)
- Funghiparadise, Seite mit zahlreichen Abbildungen von Schlauchpilzen
- Video: Fruchtkörpertypen und Sporenverbreitung bei Discomyceten (Ascomycetes). Institut für den Wissenschaftlichen Film (IWF) 1985, zur Verfügung gestellt von der Technischen Informationsbibliothek (TIB), doi:10.3203/IWF/C-1593.