Ringelwürmer
Die Ringelwürmer (Anneliden[1][2], Annelida)[an 1] oder auch Gliederwürmer bilden aufgrund ihres eigenständigen Bauplans einen Stamm innerhalb der Stammgruppe der Lophotrochozoen (Lophotrochozoa), die zu den Urmündern (Protostomia) gehören. Ringelwürmer werden in zwei Klassen eingeteilt: Vielborster (Polychaeta) und Gürtelwürmer (Clitellata), die sich in Wenigborster (Oligochaeta) und Egel (Hirudinea) aufteilen. Insgesamt gibt es etwa 18.000 verschiedene Arten.
| Ringelwürmer | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Gemeiner Regenwurm (Lumbricus terrestris) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Annelida | ||||||||||||
| Lamarck, 1809 |
Die größten bekannten Arten sind der in Australien beheimatete Riesenregenwurm Megascolides australis, der eine Länge von bis zu 3 Metern erreichen kann, sowie der Meereswurm Eunice aphroditois mit einer ähnlichen Länge und bis zu 1000 Einzelsegmenten. Die kleinsten Arten leben im Grundwasser und in der Sandlückenfauna und erreichen Längen von 300 Mikrometern (Gattung Diurodrilus), die Zwergmännchen der Art Dinophilus gyrociliatus werden sogar nur 50 Mikrometer lang.
Aufbau
Die wichtigsten evolutionären Änderungen gegenüber den Bauplänen einfacher gebauter bilateralsymmetrischer Tiere wie den Fadenwürmern oder den Plattwürmern sind bei den Anneliden der Erwerb eines echten Coeloms und einer Segmentierung. Das Coelom bietet Platz für komplexe Organe und erlaubt neuartige differenzierte Bewegungsformen. Hautmuskelschlauch und Eingeweidemuskulatur können unabhängige Bewegungen vollführen. Durch die Segmentierung ist es möglich, einzelne Segmentabschnitte für bestimmte Aufgaben zu spezialisieren (heteronome Segmentierung). Insgesamt lässt sich der Körper eines Ringelwurms in drei Regionen gliedern:
- Kopf (Prostomium und Peristomium)
- Rumpf (gleichartige Segmente)
- Hinterende (Pygidium)
Diese Einteilung kann bei einigen Untergruppen der Anneliden nicht mehr klar erkennbar sein.
Segmente und Parapodien
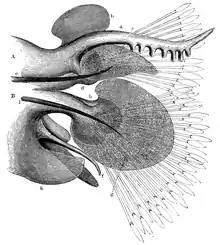

Die deutsche Bezeichnung „Ringelwurm“ geht darauf zurück, dass der Körper dieser Tiere aus zahlreichen sogenannten Segmenten aufgebaut ist, was von außen als Ringelmuster erscheint. Tatsächlich wurden diese Segmente historisch als „Ringe“ oder „Ringel“ bezeichnet.vgl. z. B. [3][4] An den einzelnen Rumpfsegmenten befinden sich, vor allem bei den Polychaeten gut ausgeprägt, paarförmige laterale Stummelfüße (Parapodien), in denen Borsten, die so genannten Chaetae, verankert sind. Die Parapodien sind Ausstülpungen der Körperwand, in die hinein sich die Leibeshöhle (Coelom) ausdehnt und in die sich Muskeln ziehen. Die Segmente werden nacheinander an der Sprossungszone vor dem Körperhinterende gebildet (Teloblastie). Bei manchen Arten ist die Anzahl der Segmente bei allen Individuen konstant, bei anderen Arten schwankt die Anzahl um wenige Segmente. Die meisten Ringelwürmer leben im Wasser, einige Arten (vor allem größere) besitzen auch Kiemen, diese sitzen meistens an den Parapodien.
Leibeshöhle und Fortbewegung
Die sekundäre Leibeshöhle (Coelom) des segmentierten Körpers, die bei den Ringelwürmern das erste Mal in der Evolution auftaucht, ist vollständig mit Flüssigkeit gefüllt und wird von einem geraden, rohrartigen Darm durchzogen. Das hydrostatische Skelett ermöglicht im Zusammenspiel mit den Ringmuskeln, den Längsmuskeln und den Borsten ein effektives Fortbewegungssystem. „Errante“, also nicht in Röhren oder Bauten lebende Ringelwürmer, erhalten durch dieses System ihr typisches Fortbewegungsmuster. Je nach Lebensraum und zum Teil auch nach unterschiedlicher Lebensphase können Anneliden kriechen, schwimmen, sich schlängeln oder auch undulieren (rhythmische Bewegung, um für Wasserdurchströmung in Wohnröhren und damit für Durchlüftung zu sorgen).
Blutkreislauf
Das Blutgefäßsystem ist geschlossen und besteht aus einem Rücken- und einem Bauchgefäß. Das Bauchgefäß führt das Blut von vorne nach hinten, das Rückensystem liegt oberhalb des Darms und führt das Blut von hinten nach vorne. Meist ist das Rückengefäß kontraktil und manchmal auch doppelt vorhanden. Die Längsgefäße werden häufig durch Seitengefäße verbunden. Es besteht kein Endothel, das Blut fließt in den Spalträumen zwischen aneinandergrenzenden Geweben. Als Atmungspigmente wurden Hämoglobine, Chlorocruorin (grün) und Hämerythrin (violett) nachgewiesen. Der Gasaustausch und damit auch die Atmung erfolgt über die Haut.
Nervensystem
Vor allem bei ursprünglich gebauten Ringelwürmern findet eine Cephalisation statt. Die neuronalen Zentren werden ursprünglich im Kopfbereich (Prostomium) zusammen mit den wichtigsten Sinneszellen als Oberschlundganglion konzentriert. Das Prostomium ist mit Licht- und Chemorezeptororganen und beweglichen Anhängen ausgerüstet. Auf diesen Anhängen (Palpen und Antennen) befinden sich ebenfalls Rezeptoren. Zur Steuerung der einzelnen Segmente finden sich aber nach wie vor Anhäufungen von Nervenzellen, sogenannte Ganglien, paarig in den einzelnen Segmenten. Auch hier kann es zur Konzentration von Ganglien kommen.
Das für die Anneliden typische Strickleiternervensystem findet sich als Bauchmark auf der ventralen Seite dieser Tiere. Die in Lehrbüchern oft schematisch dargestellte Nervenstrickleiter ist im präparierten Tier nicht ohne weiteres zu erkennen. Mit bloßem Auge sieht man auch bei großen Ringelwürmern nur eine Ansammlung von Nervenfasern, die auf den ersten Blick keiner Strickleiter ähnelt. Dies liegt daran, dass bei diesen Taxa die Nervenstränge zu einem einheitlichen Strang verschmolzen sind, oder die Nervenzellkörper verteilen sich über die Konnektive.
An das Prostomium schließt sich das Peristomium mit der Mundöffnung und weiteren Anhängen an. Diese Anhänge können sekundär fehlen. Auch das Hinterende ist ursprünglich mit Anhängen versehen, auf denen sich Rezeptoren befinden. Augen kommen in vielen Ausprägungen vor – von den zweizelligen Ocellen bis hin zum komplexen Linsenauge (Tomopteris). Am häufigsten liegen sie am Prostomium, können jedoch auch an anderen Stellen (z. B. den Segmenten) vorkommen. Nuchalorgane kommen innerhalb der Anneliden nur bei den Polychaeten vor. Es sind paarige, stark bewimperte Strukturen am hinteren Ende des Prostomiums, die vor allem Chemorezeptoren enthalten.
Epidermis und Hautmuskelschlauch
Die weiche Epidermis der Anneliden besitzt viele Drüsen. Diese Drüsen produzieren beispielsweise Schleime, Sekrete zum Bau von Röhren oder sogar Leuchtsekrete. Die Epidermis wird außen von einer Cuticula umgeben welche meistens ein Gitter aus Kollagenfasern enthält. Die für die meisten Anneliden charakteristischen Borsten, welche beweglich sind, sind ebenfalls Abscheidungen der Epidermis. Das heißt, diese Borsten stehen segmental angeordnet in tiefen Epidermistaschen. Die Anzahl und Form der Borsten ist bei jeder Art sehr konstant und bilden ein wichtiges Unterscheidungsmerkmal.
Unter der Epidermis verlaufen zwei Muskelschichten: eine Ringmuskelschicht und eine Längsmuskelschicht. Zwischen diesen Schichten können diagonale Fasern liegen. Zu diesen Muskeln kommen diverse weitere Muskeln wie jene, die die Borsten bewegen, Parapodialmuskeln oder Dorsoventralmuskeln. Eine Kräfteübertragung findet durch das wassergefüllte Coelom statt, welches mit den Kollagenfasern der Cuticula das hydrostatische Skelett bildet. Die Muskelzellen sind einkernig und gehören zum Typ schräggestreifter Muskulatur.
Darmtrakt und Exkretionsorgane
Der Darmtrakt der Ringelwürmer ist primär ein mit einem einschichtigen Epithel umhülltes Rohr, welches gerade und in drei Abschnitte gegliedert ist. Er zieht sich von der ventralen Mundöffnung am Peristomium bis zum After am Pygidium. Davon ausgehend gibt es je nach Ernährungsweise und Lebensweise viele Variationen.
Die Exkretion erfolgt durch die Nephridien. Ursprünglich sind diese in jedem Segment paarweise vorhanden. Die Mehrzahl der Anneliden besitzt Metanephridien, es kommen jedoch auch Protonephridien vor.
Lebensräume, Ernährung
Die Vielborster sind meist Meerestiere. Die festsitzenden Arten („Sedentaria“) ernähren sich von Plankton, die freilebenden Arten („Errantia“) ernähren sich meist räuberisch oder als Weidegänger. Einige Vielborster sind zwar grundsätzlich freibeweglich, werden jedoch zu den Sedentaria gezählt, weil sie in Röhren leben und an eine ortsgebundene Lebensweise angepasst sind, wie der Pier- oder Wattwurm (Arenicola marina). Sie und auch die zu den Wenigborstern gehörenden Regenwürmer wie Lumbricus terrestris ernähren sich von Bakterien und organischen Stoffen, indem sie das umgebende Substrat fressen und die nährstoffreichen Anteile verdauen. Regenwürmer und Wattwürmer sind damit erheblich an der Umwälzung und Auflockerung der bewohnten Böden beteiligt. Auch die vorwiegend im Süßwasser vorkommenden Egel gehören zu den Anneliden. Sie sind entweder blutsaugende Ektoparasiten (beispielsweise erwachsene Tiere des Medizinischen Blutegels, Fischegel und Schneckenegel) oder leben räuberisch (beispielsweise Pferdeegel, Rollegel).
Fortpflanzung
Die Fortpflanzung der Ringelwürmer ist äußerst vielgestaltig. Es gibt zwittrige und geschlechtliche Fortpflanzung, lebend gebärende Arten, Larvalentwicklung, Sprossung, Generationswechsel. Je nach Lebensweise haben sich passende Formen der Fortpflanzung herausgebildet.
Reparative Regeneration
Die Möglichkeit, Amputationen durch neue Sprossungszonen zu reparieren, ist bei den Anneliden weit entwickelt und wird von ihnen auch zur Fortpflanzung genutzt. Bei einigen Arten reicht ein einzelnes Segment, um die restlichen fehlenden zu ergänzen. Regenerierende Regenwürmer verfallen in eine Körperstarre.
Geschlechtliche Fortpflanzung
Die geschlechtliche Fortpflanzung der Anneliden findet ursprünglich frei im Wasser statt. Das heißt, die Gameten werden ins Wasser abgegeben und verschmelzen dort. Die direkte Übertragung des Spermas auf den Geschlechtspartner hat sich hiervon mehrfach unabhängig voneinander entwickelt, einige Arten besitzen auch äußere Geschlechtsorgane.
Larvalentwicklung
Ursprünglich geht die Larvalentwicklung über eine Trochophora-Larve. Eine Reihe von Polychaeten und alle Clitellaten entwickeln sich jedoch direkt.
Systematik
Lange Zeit wurden die Ringelwürmer in Vielborster – meist Meeresbewohner mit Parapodien, an denen zahlreiche Borsten sitzen – und Gürtelwürmer – meist Süßwasser- oder Landbewohner ohne Parapodien sowie mit einem Clitellum (Gürtel) und wenigen oder überhaupt keinen Borsten – unterteilt. Die Vielborster zerfallen wiederum in sessile oder eingeschränkt bewegliche Sedentaria und mobile Errantia, die Gürtelwürmer dagegen in Wenigborster mit wenigen Borsten und räuberische oder blutsaugende Egel ohne Borsten, mit Saugnäpfen, einem von kräftigen Muskeln durchzogenen Körper und reduziertem Coelom.
Der Großteil der Arten der Ringelwürmer verteilt sich in der traditionellen Systematik auf die Vielborster (Polychaeta) mit etwa 10000 Arten und die Wenigborster (Oligochaeta) mit etwa 7700 Arten, während die Egel nur rund 300 Arten zählen – die Gürtelwürmer (Clitellata) zusammen also etwa 8000 Arten. Hinzu kommt noch eine systematisch umstrittene Gruppe an Stachelhäutern parasitierender Formen, die Myzostomida mit etwa 140 Arten.
Phylogenetische Untersuchungen auf Grundlage der Anatomie der Vielborster führten zur Systematik nach Rouse & Fauchald (1998). Während die Errantia hiernach monophyletisch sind und in der Gruppe Aciculata aufgehen, also Vielborstern mit kräftigen, muskulösen, durch ein Innenskelett von Aciculae gestützten Parapodien, werden die Sedentaria aufgelöst in die primitiven, meist eingeschränkt mobilen Scolecida und die sessilen Canalipalpata – bereits unter Einschluss der ehemaligen Bartwürmer – mit teilweise zu ausgedehnten Tentakelsystemen entwickelten Palpen. Die Canalipalpata sind nach Rouse & Fauchald (1998) näher mit den Aciculata als mit den Scolecida verwandt und bilden mit den Aciculata das gemeinsame Taxon Palpata. Bei Betrachtung sämtlicher Ringelwürmer wie auch der später mit einbezogenen ehemaligen eigenen Stämme Sipuncula und Echiura kann folgendes Kladogramm gezeichnet werden:
| |||||||||||||||||||||||||||||||||||||
Die Spritzwürmer (Sipuncula), die Igelwürmer (Echiura) und die Bartwürmer (ehemals Pogonophora, heute nur noch Familie Siboglinidae innerhalb der Sabellida) galten bis vor kurzem als eigenständige Stämme, bzw. ihre systematische Einordnung war lange umstritten. Auf Grund jüngerer phylogenetischer Untersuchungen auf molekulargenetischer Grundlage wie von Andrade und anderen (2015), Parry und anderen (2016), Torsten Hugo Struck und anderen (2015) sowie Anne Weigert und Christoph Bleidorn (2016) müssen diese Taxa aus kladistischer Sicht den Ringelwürmern zugeordnet werden. Die Bartwürmer besitzen wie alle Anneliden bewimperte Trochophoralarven. Daneben verweisen auch die Segmentierung des Endstücks und das Sauerstofftransportmolekül Hämoglobin auf eine Einordnung in die Ringelwürmer. Die bisherigen Taxa Oligochaeta (Wenigborster) und Polychaeta (Vielborster) gibt es hiernach nicht mehr, denn sie sind paraphyletisch. Wiederbelebt werden hier die alten Taxa „Errantia“ und „Sedentaria“, allerdings in ganz anderem Umfang als früher.
Kladogramm nach Struck et al. 2015 sowie Weigert & Bleidorn 2016:
| Annelida s. lato |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Anmerkung
- Aus der Sicht des klassischen Latein müsste der Name Anellida bzw. Anelliden geschrieben werden, entsprechend ānellus „kleiner Ring“ im klassischen Latein, vgl. Charlton T. Lewis und Charles Short, A Latin Dictionary, Stichwort ānellus. Im nachklassischen Latein bildete sich die Variante ānnellus heraus, auf die sich die Schreibweise Annelida bzw. Anneliden zurückführen lässt; vgl. annellus im englischen Wiktionary.
Einzelnachweise
- Duden online: Anneliden
- Meyers Großes Konversations-Lexikon (1908), Stichwort Ringelwürmer zeno.org
- Oscar Schmidt: Brehms Thierleben. Allgemeine Kunde des Thierreichs. Große Ausgabe. Zweite umgearbeitete und vermehrte Auflage. Vierte Abtheilung: Wirbellose Thiere. Zweiter Band: Die Niederen Thiere. Verlag des Bibliographischen Instituts, Leipzig 1878, doi:10.5962/bhl.title.1067, S. 68 ff.
- Hermann Burmeister: Handbuch der Naturgeschichte. Zweite Abtheilung: Zoologie. Verlag von Theodor Christian Friedrich Enslin, Berlin 1837 (HathiTrust), S. 531 ff.
Literatur
- Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Band 1. Spektrum Akademischer Verlag, 2003, ISBN 3-8274-1482-2.
- Gregory W. Rouse, Kristian Fauchald (1998): Recent views on the status, delineation, and classification of the Annelida. (PDF; 959 kB). American Zoologist. 38 (6), S. 953–964. doi:10.1093/icb/38.6.953
- Luke A. Parry, Gregory D. Edgecombe, Danny Eibye-Jacobsen, Jakob Vinther (2016): The impact of fossil data on annelid phylogeny inferred from discrete morphological characters. The Royal Society Publishing 2016. DOI: 10.1098/rspb.2016.1378
- Torsten Hugo Struck, Anja Golombek, Anne Weigert, Franziska Anni Franke, Wilfried Westheide, Günter Purschke, Christoph Bleidorn, Kenneth Michael Halanych (2015): The Evolution of Annelids Reveals Two Adaptive Routes to the Interstitial Realm Current Biology. Current Biology 25 (15), S. 1993–1999. DOI: 10.1016/j.cub.2015.06.007
- Anne Weigert, Christoph Bleidorn (2016), Current status of annelid phylogeny. Organisms Diversity and Evolution 16 (2), S. 345–362. DOI: 10.1007/s13127-016-0265-7
- Sónia C. S. Andrade, Marta Novo, Gisele Yukimi Kawauchi, Katrine Worsaae, Fredrik Pleijel, Gonzalo Giribet, Gregory W. Rouse (2015): Articulating “Archiannelids”: Phylogenomics and Annelid Relationships, with Emphasis on Meiofaunal Taxa. Molecular Biology and Evolution 32 (11), S. 2860–2875.
Weblinks
- Annelida im Tree-of-Life-Projekt
- Introduction to the Annelida
- Center for Annelida Ressources
- Annelida-Quellen zusammengestellt von Geoff Read mit Wissenschaftlerkontakten