Großer Igeltenrek
Der Große Igeltenrek oder Große Igeltanrek (Setifer setosus), teilweise auch Gewöhnlicher Igeltanrek, ist eine Säugetierart aus der Familie der Tenreks (Tenrecidae). Er ist endemisch auf Madagaskar verbreitet und kommt auf der gesamten Insel vor. Die Tiere bevorzugen zwar die tropisch-feuchten Landschaften des östlichen Inselteils, sind aber auch in den trockenen westlichen Gebieten recht häufig. Zudem dringen sie in städtische Regionen vor und können sich an durch Menschen veränderte Landschaften gut anpassen. Äußerlich ähnelt der Große Igeltenrek den europäischen Igeln, er ist aber deutlich kleiner und hat einen kürzeren Schwanz, ebenfalls charakteristisch sind die langen Vibrissen. Die Fellfärbung kann regional variieren. Von anderen Tenreks und vor allem von seinem nächsten Verwandten, dem Kleinen Igeltenrek, unterscheidet er sich durch den Gebissaufbau.
| Großer Igeltenrek | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
.jpg.webp)
Großer Igeltenrek (Setifer setosus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Setifer | ||||||||||||
| Froriep, 1806 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Setifer setosus | ||||||||||||
| (Schreber, 1777) |
In der Regel lebt der Große Igeltenrek einzelgängerisch und ist nachtaktiv. Er bewegt sich am Boden vorwärts, kann aber auch in den Bäumen klettern. Die Befähigung ist jedoch nicht so deutlich ausgeprägt wie beim Kleinen Igeltenrek. Die Tiere unterhalten Eigenreviere, die sich an den Rändern mit denen anderer Individuen überlappen. Die Territorien sind im Vergleich zu Tierarten mit ähnlichen Körperausmaßen sehr groß. Zur Ruhe zieht sich der Große Igeltenrek in Baumlöcher und unterirdische Baue zurück. Die Nahrung umfasst Wirbellose und kleinere Wirbeltiere, insgesamt herrscht eine eher allesfresserische Ernährungsweise vor. Der Nachwuchs kommt zumeist in der feuchteren und wärmeren Jahreszeit zur Welt, ein Wurf umfasst bis zu sieben Jungtiere, die relativ schnell heranwachsen. In den trockenen Gebieten des westlichen Madagaskar können Weibchen in einer Fortpflanzungsperiode bis zu drei Würfe gebären. Charakteristisch für den Großen Igeltenrek ist die variierende Körpertemperatur, die von den Außentemperaturen und der Aktivität abhängt und über den Tag und das Jahr einen zyklischen Ablauf aufweist. Gleiches gilt für die Stoffwechselrate, die allgemein sehr gering ist. Die Trockenzeit verbringt der Große Igeltenrek in einem mehrmonatigen Torpor.
In Europa wurde erstmals Mitte des 17. Jahrhunderts über den Großen Igeltenrek berichtet. Die wissenschaftliche Erstbeschreibung der Art erfolgte dann rund einhundert Jahre später, im Jahr 1777. Allgemein wurden die Tiere anfangs mit den europäischen Igeln in Verbindung gebracht. Erst zu Beginn des 19. Jahrhunderts kam es zu einer stärkeren Differenzierung. Seinen heute gültigen Gattungsnamen erhielt der Große Igeltenrek im Jahr 1806. Doch wurde er im weiteren Verlauf des 19. und teilweise auch des 20. Jahrhunderts häufig in andere Gattungen verwiesen. Der Bestand des großen Igeltenreks gilt als nicht gefährdet. In einigen Regionen Madagaskars wird er zu Nahrungszwecken gejagt.
Merkmale
Habitus
_(9614561811).jpg.webp)
Der Große Igeltenrek ist ein mittelgroßer Vertreter der Tenreks. Mit einer Gesamtlänge von 16 bis 22,5 cm wird er nur wenig größer als der Kleine Igeltenrek (Echinops telfairi), wie bei diesem besteht der Schwanz aus einem nur 15 bis 16 mm langen Stummel. Das Körpergewicht variiert von 150 bis 220 g, kann mitunter aber auch 300 g und mehr erreichen,[1][2] allgemein ist es abhängig von der Kondition des einzelnen Individuums und der Jahreszeit. Ein auffälliger Geschlechtsdimorphismus liegt nicht vor. Im äußeren Erscheinungsbild ähneln die Tiere den Igeln, was durch den robusten, rundlichen Körper, die spitze Schnauze und durch das Stachelkleid hervorgerufen wird. Unterschiede bestehen in dem sehr kurzen, kegelförmigen Schwanz, den sehr langen Vibrissen und der geringeren Körpergröße, die nur die die Hälfte eines europäischen Igels erreicht. Das Rückenfell wird durch sehr dicht stehende, durchschnittlich 15 mm lange Stacheln gebildet. Generell haben Tiere aus dem feuchteren östlichen Madagaskar dunklere Stachelspitzen als jene aus dem trockeneren westlichen. Die Körperunterseite, das Gesicht und die Gliedmaßen zeigen eine nur spärliche, Fellbedeckung aus hellgrauem, weichem Haar. Die Augen sind relativ klein, ebenso die Ohren, die etwa die Länge der Stacheln besitzen. Die vergleichsweise langen Gliedmaßen enden vorn und hinten in jeweils fünf Strahlen. Diese tragen Krallen, an den Händen sind sie 6, an den Füßen 7 mm lang.[3] Der gesamte Hinterfuß misst etwa 31 mm in der Länge. Weibliche Tiere haben fünf oder sechs Paare an Zitzen.[4][5][6][7][8][9]
Schädel- und Gebissmerkmale
Die durchschnittliche Schädellänge liegt bei 46,2 mm, die durchschnittliche Breite am Jochbogen gemessen 23,1 mm. Die schmalste Stelle wird an den Augen mit einem Durchschnittswert von 10,9 mm erreicht.[10][2] Allgemein ist der Schädel moderat robust, das Rostrum wirkt breit und hoch, der Hirnschädel kurz. Das Verhältnis des vorderen zum hinteren Schädelabschnitts beträgt 0,61:0,39.[11] Im Unterschied zum Kleinen Igeltenrek wölbt sich die Stirnlinie stärker auf, da das Stirnbein im hinteren Bereich aufgebläht ist. Der Jochbogen ist wie bei allen Tenreks nicht geschlossen. Im hinteren Schädelbereich sind kräftige Knochenwülste als Muskelansatzstellen ausgebildet. Das Gebiss setzt sich aus 36 Zähnen zusammen, die Zahnformel lautet folgendermaßen: . Es ist somit nicht ganz so stark reduziert wie beim Kleinen Igeltenrek. Der obere innere Schneidezahn erreicht fast die Größe des Eckzahns und ist gut entwickelt. Im unteren Gebiss bestehen kurze Diastemata beidseitig des Eckzahns und des vordersten Prämolaren. Die beiden hinteren Prämolaren sind molarisiert und ähneln so den Mahlzähnen. Diese besitzen ähnlich den anderen Tenreks ein zalambdodontes Kauflächenmuster bestehend aus drei Haupthöckern. In der oberen Gebissreihe zeigen die Molaren nicht ganz so starke Größenreduktionen wie beim Kleinen Igeltenrek. Demzufolge ist beim Großen Igeltenrek der vorderste Molar der größte Zahn in der Reihe und nicht der letzte Prämolar wie beim Kleinen Igeltenrek.[11][10]

Skelettmerkmale
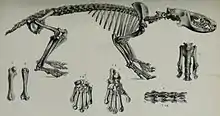
Die Wirbelsäule setzt sich aus 7 Hals-, 16 bis 17 Brust-, 6 bis 7 Lenden-, zwei Kreuzbein- und 9 Schwanzwirbel zusammen.[11] Im Bau des Bewegungsapparates finden sich einzelne Merkmale, die sowohl auf eine bodenlaufende als auch eine baumbewohnende Lebensweise schließen lassen, letztere sind in der Regel nicht so deutlich ausgeprägt wie beim Kleinen Igeltenrek. Das Schulterblatt ist eher breit und nicht so langschmal wie bei den grabenden Tenrekformen. Am Oberarmknochen ist der Kopf eher rundlich ausgebildet, was mit dem Kleinen Igeltenrek übereinstimmt, abweichend von diesem aber analog zum Großen Tenrek (Tenrec ecaudatus) verbreitert sich die untere Gelenkrolle (Ellenbogengelenk) vorn und hinten. Wie bei den beiden anderen Tenrekvertretern auch zeigt der obere Gelenkfortsatz der Elle, das Olecranon, mit einer Länge von nur 16 % des Gesamtknochens kaum Tendenzen zur Streckung, die hier ansetzende Unterarmmuskulatur ist demnach nicht übermäßig kräftig. Im Unterschied zu den grabenden Vertretern der Tenreks treten an den Handwurzelknochen keine Verwachsungen auf, was eine höhere Rotationsfähigkeit der Hand befürwortet. Die Mittelhandknochen sind wiederum verlängert, aber nicht in dem Maße wie beim Kleinen Igeltenrek. Ebenso ist der innere Strahl insgesamt kürzer und die Greiffähigkeit damit eher eingeschränkt. Unterschiede finden sich auch an den Krallen, die beim Großen Igeltenrek länger und nicht ganz so stark gekrümmt sind wie bei seinem Verwandten. Sie entsprechen aber mit ihrer insgesamt hochschmalen Form den Krallen anderer kletternder Säugetiere.[12][13] Auch die Füße besitzen einzelne Modifikationen wie die flachen Knöchel, die wie beim Kleinen Igeltenrek die Mobilität weniger einschränken. Dass der Fuß des Großen Igeltenreks nicht so extrem beweglich ist wie bei Kleinen Igeltenrek zeigen wiederum die stärkeren Leisten auf der Sprungbeinrolle des Sprungbeins und die dadurch bedingte tiefere Einwölbung der Gelenkfläche, wodurch das Schienbein besser fixiert wird. Ebenso ist der Übergang zum Hals des Sprungsbeins stärker strukturiert.[14][9]
Verbreitung
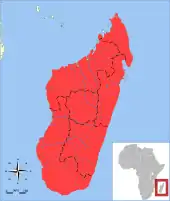
Der Große Igeltenrek kommt endemisch auf Madagaskar vor und ist dort über die gesamte Insel verbreitet. Er kann in nahezu allen Landschaftsräumen angetroffen werden mit Ausnahme von Überschwemmungs- und Marschgebieten. Zusätzlich tritt er auch in einzelnen städtischen Regionen wie beispielsweise Antananarivo auf und ist selbst in extrem stark vom Menschen überprägten Bereichen beobachtet worden. Die Tiere favorisieren zwar die feuchteren östlichen Wälder, sie sind aber auch in den trockeneren Landschaften des Westens heimisch. Dort wurden seit Untersuchungen in den 1990er Jahren fast 30 Fundplätze mit Nachweisen der Art registriert, im Südwesten tritt sie sympatrisch mit dem Kleinen Igeltenrek auf.[15] In der Regel besiedeln die Tiere die Tieflandsregionen unter 900 m, lokal reicht die Höhenverteilung aber bis etwa 2250 m. In einigen Gebieten, wo der Große Igeltenrek vorkommt, gehört er mit zu den am häufigsten nachgewiesenen Vertretern der Tenreks.[16]
Lebensweise
Territorial- und Sozialverhalten
Die Lebensweise des Großen Igeltenreks ist durch Beobachtungen in Gefangenschaft und durch Felduntersuchungen relativ gut erforscht. Die Tiere sind nachtaktiv mit einer bimodalen Verteilung der Aktivität. Die erste Phase beginnt bereits gegen 14:00 Uhr und erreicht gegen 19:00 und 20:00 Uhr ihren Höhepunkt, wonach eine Ruhepause in den zwei Stunden vor Mitternacht eintritt. Ein zweiter Höhepunkt wird gegen 02:00 Uhr erreicht, die Aktivität ebbt dann in den folgenden vier Stunden ab. Daraus ergibt sich, dass die die Tiere etwa 16 Stunden eines Tages aktiv verbringen. Allerdings verzögert sich häufig der Beginn bei niedrigeren Außentemperaturen. Die Tiere sind weitgehend bodenbewohnend, wo sie sich im Kreuzgang und mit dem ganzen Fuß aufsetzend vorwärts bewegen. Im schnellen Lauf berühren aber meist nur die Zehen den Boden. Ebenso ist der Große Igeltenrek ein vergleichsweise guter Kletterer, sein Fuß kann jedoch nicht so stark gedreht werden wie beim Kleinen Igeltenrek. Prinzipiell sieht man ihn seltener in den Bäumen als seinen kleineren Verwandten. Wie am Boden so ist er auch im Geäst eher langsam und klettert mit wechselseitigen Bewegungen der Gliedmaßen. Hände und Füße umgreifen die Zweige, teilweise wird der Schwanz zum Abstemmen eingesetzt. In unbekanntem Gelände pausiert der Kleine Igeltenrek häufig und schnüffelt in der Luft, mitunter erhebt er sich dabei auf die Hinterbeine.[5][17][6][7][9]

In der Regel tritt der Große Igeltenrek mit Ausnahme von Mutter-Jungtier-Gruppen einzeln auf. Die Tiere unterhalten Eigenreviere, die nach Untersuchungen im Nationalpark Ankarafantsika im Nordwesten von Madagaskar relativ groß sind für verhältnismäßig kleine Säugetiere. Demnach beanspruchen Männchen ein Territorium von 4,9 bis 21,1 ha Größe (durchschnittlich 13,7 ha), bei Weibchen sind es 0,9 bis 9,2 ha (durchschnittlich 6,7 ha). Die Reviere überschneiden sich an den Rändern mit anderen, dabei sind die Überlappungen mehr oder weniger unabhängig vom Geschlecht des Nachbarn. Innerhalb der Territorien ruhen die Tier in Baumhöhlen und hohlen, abgestorbenen Bäumen. In Ankarafantsika befanden sich die Ruheplätze in 0,5 bis über 2 m Höhe in Bäumen mit überwiegend 10 bis 20 cm Stammdurchmesser. Säugende Weibchen suchen sich dabei durchschnittlich höhere Lagen, wohl als Schutz vor Beutegreifern. Zudem sollten die Unterschlüpfe wenig Einblick bieten. Mutter- und Jungtiere bleiben auch über längere Zeit in den Baumhöhlen, etwa 20 bis 25 Tage. Ansonsten wechselt der Kleine Igeltenrek seinen Schlafplatz täglich und benutzt einen älteren nur selten erneut. Mitunter kann aber ein Nachbartier einen bereits gebrauchten Unterschlupf aufsuchen und besetzen.[1] Die Rückzugsplätze enthalten Nester aus Pflanzenmaterial, das von den Tieren im Maul herangetragen wird. In Regionen mit geeignetem, nicht sandigem Untergrund zieht sich der Kleine Igeltenrek auch in unterirdische Baue zurück, die er teilweise selbst in Hangbereichen anlegt. Zwei beobachtete Baue bei Manandroy nördlich von Fianarantsoa im zentral-östlichen Madagaskar bestanden aus einem 60 cm langen, rund 6 cm breiten Gang, der in eine Nestkammer von 7,5 cm Weite mündete. Derartige Baue werden möglicherweise regelmäßiger aufgesucht.[7] In Ruheposition rollt sich ein Tier auf den Hinterbeinen hockend zusammen, selten nimmt es eine Seitenlage ein.[17][7][9]
Begegnungen zwischen gleichgeschlechtlichen Tieren zeichnen sich durch Vermeidungsverhalten aus. In der Regel bestehen sie aus dem gegenseitigen Beschnuppern an der Nase und am Körper, wonach sich die Tiere wieder trennen. Bei aggressiven Verhalten werden die Stacheln aufgestellt und das Maul geöffnet. Stärkere Bedrängung resultiert in typischen auf- und abwärtsführenden oder seitlichen Kopfbewegungen und in Beißattacken. Bedrohte Tiere fliehen zumeist, wobei sie bis zu 3,2 km/h erreichen, oder rollen sich zu einer Kugel zusammen, bei der die Stacheln maximal aufgestellt sind. Für die innerartliche Kommunikation ist der Geruchssinn bedeutend. Der Große Igeltenrek defäziert häufig nahe den Eingängen seiner Baue. Auch reibt er seinen Körper an Ästen und Zweigen und setzt so Duftmarken. Die Lautkommunikation erfolgt über verschiedene Töne. Am häufigsten sind Tschilp-Laute mit einer Frequenz von 1000 bis 3000 Hz und einer Dauer von 0,03 bis 0,05 s. Unter Bedrängnis stoßen die Tiere einen „putt-putt“-Ruf aus, etwa beim Einrollen, oder ein Zischen. Das Komfortverhalten umfasst neben dem Kratzen und Lecken auch das für andere Tenreks typische „Gesichtwaschen“ in sitzender Position mit beiden Vorderbeinen gleichzeitig.[5][17][6][7][9]
Ernährung und Energiehaushalt
In seiner Ernährungsweise ist der Große Igeltenrek wenig spezialisiert. Er nimmt sowohl Wirbellose wie Heuschrecken, sonstige Insekten und Regenwürmer sowie kleinere Wirbeltiere, etwa Frösche und Mäuse zu sich. In menschlicher Gefangenschaft vertilgte er auch Innereien oder Fleisch größerer Säugetiere.[5][18] Das Gebiss des Großen Igeltenreks ist aber nicht geeignet, Beute größer als Insekten oder Jungmäuse zu töten. In der Umgebung von Siedlungen wurden Tiere beim Durchwühlen von Abfall beobachtet, teilweise fressen sie auch Aas.[19][20] Die prinzipiell allesfresserische Ernährung bestätigten auch Isotopenuntersuchungen an Individuen aus Tsinjoarivo im zentral-östlichen Madagaskar.[21] Die Beute wird mit den Zähnen aufgegriffen, die Vorderfüße spielen bei der Manipulation der Nahrung kaum eine Rolle. Die täglich aufgenommene Nahrungsmenge variiert und ist abhängig von der Kondition der Tiere und der Jahreszeit. Unter menschlicher Obhut fraßen die Tiere durchschnittlich 15 bis 20 g Fleisch und 5 bis 6 Mehlwürmer oder 2 bis 3 Wüstenheuschrecken. Der Große Igeltenrek trinkt regelmäßig Wasser.[5][17][7][9]
Entsprechend zahlreichen anderen Tenreks variiert die Körpertemperatur beim Großen Igeltenrek beträchtlich und ist abhängig von den Umgebungsbedingungen und der Aktivität. Laboruntersuchungen unter kontrollierten Bedingungen zufolge wurden die tiefsten Werte für die Körpertemperatur während der Ruhezeit von den frühen Morgenstunden bis zur Mittagszeit erreicht. Sie lagen dann nur wenige Grad über der Außentemperatur, im Mittel bei 22 °C. Die höchsten Werte stellten sich in der aktiven Phase in den frühen Abend- und Nachtstunden ein, der mittlere Wert betrug 31,5 °C. Die Tiere schränkten bei sinkenden Außentemperaturen ihre Aktivitäten ein und wurden lethargisch, bei weniger als 15 °C fielen sie in einen Torpor.[22][5][17][23] Freilandstudien im Nationalpark Ankarafantsika bestätigten die Ergebnisse teilweise. Diesen zufolge ist während des Südsommers (November bis Mai) die Körpertemperatur in der inaktiven Phase tatsächlich durchschnittlich geringer als in der aktiven, die entsprechenden Werte betragen 30,8 °C und 32,2 °C. Die absoluten Grenzwerte schwanken von 26,5 bis 35,1 °C, was eine Spanne von mehr als 8 °C ausmacht. Während der aktiven Phase ist die Körpertemperatur deutlich stabiler als in der inaktiven Phase. Bei letzterer sind die Schwankungen teilweise so stark, dass sie im oberen Temperaturbereich mitunter die in der aktiven Phase erreichten Maximalwerte überschreiten. Demnach findet während der Aktivitätszeit eine Temperaturregulierung statt. Die jeweiligen Außentemperaturen waren im Untersuchungszeitraum wenig veränderlich und lagen bei 20 bis 30 °C.[24][25][26] Ähnliche Werte ergaben Studien in der Umgebung von Fianarantsoa. Die hier gemessenen durchschnittlichen Körpertemperaturen sind aber etwas niedriger, ebenso wie die Außentemperaturen geringer ausfielen.[7] Die Feldbeobachtungen im Nationalpark Ankarafantsika zeigten zudem, dass der Torpor im Südsommer zumindest im trockenen westlichen Madagaskar bei jeder Außentemperatur eintreten kann und dann bis zu drei Stunden andauert, vorzugsweise geschieht dies am Tage während der inaktiven Phase. Dagegen vermelden Berichte aus dem feuchten östlichen Madagaskar, dass der Große Igeltenrek im Südsommer nur selten in einen Starrezustand wechselt.[19][7] Während des Torpors sinkt die Körpertemperatur ab und nähert sich der Außentemperatur, es kommt aber auch hier zu Schwankungen, die teils sehr hohe Körpertemperaturen beinhalten. Die Fluktuationen im hohen Bereich der Körpertemperatur sind wahrscheinlich der Energieersparnis geschuldet. Sie ermöglichen, dass in dem eher tropisch warmen Klima die Temperaturregulierung passiv einsetzt und nicht wie in kühleren Gegenden aktiv gestartet werden muss. Im Südwinter von Mai bis September verstärkt sich der Torpor, die Phase kann, abhängig von der geographischen Lage, fünf bis sieben Monate anhalten. Auffallend im Nationalpark Ankarafantsika sind die geschlechtsabhängig abweichenden Eintrittszeiten in die Winterruhe, bei weiblichen Tieren beginnt sie zwischen April und Mai, während sie bei männlichen im Februar bis März und damit deutlich früher einsetzt. Der Unterschied wird durch die Aufzucht des Nachwuchses durch die Weibchen bedingt.[2][24][25][26] Ähnlichen rhythmischen Schwankungen im Tagesverlauf wie die Körpertemperatur unterliegen auch die Atem- und Herzfrequenz.[5][17][9]
Eine Ausnahme bilden weibliche Tiere in der Fortpflanzungsphase oder während der Milchproduktion. Diese zeigen eine deutlich stabilere Körpertemperatur mit durchschnittlich höheren Werten, die etwa 33,1 °C betragen, die Schwankungsbreite liegt bei rund 3,5 °C. Außerdem neigen sie dann seltener dazu, in einen Torpor zu verfallen. Die stabilere Körpertemperatur ist mit einer höheren Stoffwechselrate verbunden. Normalerweise zeichnet sich der Große Igeltenrek durch einen sehr geringen Metabolismus aus,[22][23] dessen Werte etwa denen des Kleinen Igeltenreks entsprechen, er steigt aber während der Reproduktion um bis zum Zweifachen an.[24][25] Ebenfalls zyklisch verlaufen deutliche Gewichtsveränderungen. Tiere direkt nach der Winterruhe wiegen zwischen 120 und 160 g. Das während des Südsommers stark ansteigende Körpergewicht erreicht sein Maximum kurz vor der nächsten Trockenzeit, Männchen können dann bis zu 327 g wiegen. Weibchen nehmen ebenfalls an Gewicht zu, zusätzlich tragen sie noch den Nachwuchs aus, so dass einzelne Tiere dann auch über 350 g wiegen können.[1][2][17][7][9]
Fortpflanzung
Die hauptsächliche Fortpflanzungsphase fällt in den Südsommer und beginnt zumeist direkt nach dem Ende der Torporphase. Es wird aber angenommen, dass der Große Igeltenrek möglicherweise je nach geographischer Lage auch im Südwinter paarungswillig ist. Wahrscheinlich besitzt der Große Igeltenrek ein eher promiskes Paarungsverhalten, was unter anderem durch die Verteilung der einzelnen Territorien Unterstützung findet.[1] Das Männchen bewirbt das Weibchen durch Markieren des Baus und der näheren Umgebung. Die Kontaktaufnahme erfolgt durch gegenseitiges Beschnüffeln an Nase, Körper und Hinterteil. Teilweise defäkiert auch das Weibchen und die Reste werden vom Männchen abgeleckt. Beim anschließenden Aufsitzen versucht das Männchen das Weibchen mit charakteristisch scharrenden Bewegungen seiner Hinterfüße gegen das Geschlechtsteil zu stimulieren. Ein einzelner Geschlechtsakt kann bis zu 28 Minuten dauern, der gesamte Paarungsvorgang wärt teilweise über zwei Stunden, da die Trennung der Partner durch das verhaken des Penis im weiblichen Geschlechtsteil verzögert wird.[7][27][9]
Die Tragzeit beträgt zwischen 51 und 69 Tagen,[4] sie ist witterungsabhängig, bei wärmerem Wetter dauert sie kürzer als bei kälterem.[8] Etwa eine Woche vor der Geburt steigert das Weibchen seine Nestbauaktivitäten. Die Wurfgröße liegt bei bis zu sieben Neugeborenen,[4] zumeist kommen eins bis fünf Tiere zur Welt, durchschnittlich sind es drei.[28] Ein gemessenes Junges war 78 mm lang und wog 24,7 g. Die Jungen kommen als Nesthocker zur Welt, ihre Augen und Ohren sind geschlossen, der Körper ist nur spärlich behaart, allerdings sind am Rücken schon Stacheln von 0,5 mm Länge zu erkennen. Die Hautfarbe ist generell weißlich und die Haare und Stacheln haben eine gräulich-helle Tönung. Die Jungen können sich bereits in eine Kugel einrollen und mit allen vieren krabbeln. Die Mutter zeigt in den ersten Wochen starke Fürsorge und verteidigt das Nest. Nach rund einer Woche sind die Stacheln der Jungen bereits 4 mm lang. Die Augen öffnen sich zwischen dem 9. und dem 14. Tag. Zu diesem Zeitpunkt folgen die Jungen dem Muttertier aus dem Bau hinaus und beginnen, feste Nahrung zu sich zu nehmen. Eine zweite Gruppe von Stacheln von meist dunkler Färbung entwickelt sich in der dritten Woche, diese sind anderes orientiert als die vorhandenen Stacheln, so dass beide Gruppen einen effektiven Schutz geben. Zudem wächst zwischen den Stacheln ein weicher Flaum an Haaren, der später wieder verloren geht und bei Alttieren nicht vorkommt. Die Entwöhnung fängt ebenfalls in diesem Zeitraum an und endet rund zwei Wochen später. Die sexuelle Reife wird nach rund 74 Tagen erreicht.[6][7][2][9]
Innerhalb einer Fortpflanzungsperiode haben Weibchen gemäß Untersuchungen in Ankarafantsika bis zu drei Würfe. Sie können noch während der Aufzucht der Jungen wieder trächtig werden und den nächsten Nachwuchs austragen. Möglich ist dies höchstwahrscheinlich durch einen Post-partum-Östrus, der es den Weibchen ermöglicht, schon kurz nach der Geburt wieder empfangsbereit zu sein, ein Effekt, der bisher nur vom Erdtenrek (Geogale aurita) bekannt war.[2] Ob eine derartige schnelle Geburtenfolge auch bei Tieren aus dem feuchteren Osten Madagaskars vorkommt, ist unklar, bisherige Berichte sprechen meist von einem Wurf, ohne dass das Weibchen danach wieder trächtig wurde.[6][7] Es ist zu vermuten, dass die schnellere Fortpflanzung des Großen Igeltenreks im westlichen Inselteil durch die dortigen trockeneren Verhältnisse bestimmt wird, hier kommt es während der Südwinters zu einem rapiden Rückgang an verfügbaren Nahrungsquellen. Die Überlebensrate des Nachwuchses ist gering. Von sechs im Nationalpark Ankarafantsika beobachteten Würfen im Zeitraum 2010 und 2011 gingen vier verloren.[2] Das Lebenserwartung in freier Wildbahn ist nicht bekannt, das höchste Alter eines Tieres in menschlicher Obhut betrug etwa 14 Jahre.[29][9]
Fressfeinde und Parasiten
Ähnlich wie der Kleine Igeltenrek ist möglicherweise auch der Große Igeltenrek durch sein Stachelkleid und durch seine Befähigung sich in eine Kugel zusammenzurollen vor zahlreichen kleineren Beutegreifern geschützt. Gelegentlich fällt ein Individuum aber der Fossa zum Opfer.[30][31] Wesentlich häufiger ist die Erbeutung durch Schlangen dokumentiert, zu den bedeutenden Fressfeinden gehören hier die Madagaskar-Hundskopfboa und die Nördliche Madagaskarboa.[2] Äußere Parasiten umfassen vor allem Flöhe der Gattungen Paractenopsyllus und Synopsyllus,[32][33][34] außerdem liegt ein Nachweis des Rattenflohs vor, dessen Hauptwirt die Hausratte darstellt.[35] Daneben finden sich auch Zecken der Gattungen Ixodes und Haemaphysalis.[36] Als innerer Parasit wurden der Kleine Leberegel[27] und der Einzeller Eimeria identifiziert.[37] Zusätzlich sind Neoplasien dokumentiert.[38]
Systematik
Innere Systematik der Tenreks nach Everson et al. 2016[39]
|
Der Große Igeltenrek ist eine Art aus der heute als monotypisch eingestuften Gattung Setifer. Art und Gattung gehören zur Familie der Tenreks (Tenrecidae), innerhalb dieser bilden sie zusammen mit dem Großen Tenrek (Tenrec ecaudatus) sowie dem Kleinen Igeltenrek (Ehinops telfairi) und den Streifentenreks (Hemicentetes) die Unterfamilie der Igeltenreks (Tenrecinae). Die Igeltenreks wiederum stellen eine der Hauptlinien der Tenreks dar. Als charakteristisch ist ihr stacheliges Haarkleid anzusehen, wodurch sie sich von den übrigen weichhaarigen Tenreks wie den sehr diversen Reiswühlern (Oryzorictinae) und den monotypischen Erdtenreks (Geogalinae) unterscheiden. Innerhalb der Unterfamilie der Igeltenreks stellen der Große und der Kleine Igeltenrek laut molekulargenetischen Analysen Schwesterformen dar. Beide Arten ähneln sich äußerlich sehr, sie sind als einzige Tenreks befähigt, sich zu einer schützenden Kugel zusammenzurollen. Ein bedeutendes Unterscheidungsmerkmal stellt die unterschiedliche Zahnanzahl dar, zusätzlich gibt es einzelne abweichende Zahn- und Schädelmerkmale. Ihnen gegenüber wiederum steht eine Klade, die aus dem Großen Tenrek und den Streifentenreks gebildet wird.[40][41][39] Die Ursprungslinie des Großen und des Kleinen Igeltenreks trennte sich bereits im Mittleren Miozän vor etwa 10,2 Millionen Jahren auf.[39]
Die Eigenständigkeit des Großen Igeltenreks war seit seiner Erstbeschreibung durch Johann Christian von Schreber im Jahr 1777 nie angezweifelt worden. Allerdings ordnete ihm Fredericus Anna Jentink im Jahr 1879 den Kleinen Igeltenrek bei, welchen William Charles Linnaeus Martin 1838 eingeführt hatte.[42] Jentink verwies dabei auf den Holotyp des Kleinen Igeltenreks, der seiner Meinung nach aufgrund der geringeren Körpergröße und der nur zwei ausgebildeten Mahlzähne ein nicht ganz ausgewachsenes Exemplar des Großen Igeltenreks darstellen würde.[43] In der Folgezeit übernahmen einige Wissenschaftler diese Auffassung, so unter anderem George Edward Dobson in seiner klassischen Abhandlung über Insektenfresser.[44] Jedoch widersprach Oldfield Thomas dem 1892 und hob den Kleinen Igeltenrek erneut in den Artstatus, was bis heute anerkannt bleiben sollte. Für Thomas war dabei nicht nur der abweichende Gebissaufbau ausschlaggebend, er verwies zusätzlich auf die unterschiedlich geformten Krallen, die beim Großen Igeltenrek länger und breiter sind als beim Kleinen.[3]
Forschungsgeschichte
Entdeckung und Erstbeschreibung


Die ersten Berichte über den Großen Igeltenrek reichen bis in das 17. Jahrhundert zurück. In seinem im Jahr 1658 erschienenen und mehrfach aufgelegten Werk Histoire de la grande isle Madagascar erwähnte Étienne de Flacourt, ehemals Gouverneur von Madagaskar, ein igelartiges Tier, das er als „Tendrac“ bezeichnete. Nach Aussage Flacourts würde das Tier das halbe Jahr schlafen, wozu es sich in den Boden eingräbt und zudem während dieser Zeit keine Nahrung zu sich nimmt. Neben einer Zeichnung des „Tendrac“ gibt er auch den lokalen Namen „Sora“ an.[45] Mehr als einhundert Jahre später beschrieb Georges-Louis Leclerc de Buffon im Jahr 1764 im zwölften Band seines Werkes Histoire naturelle, générale et particulière den „Tendrac“ genauer und bildete ihn ebenfalls ab. Demnach hatte der „Tendrac“ ein charakteristisch igelartiges Stachelkleid, allerdings war er deutlich kleiner als die damals bekannten europäischen Igel, so dass seine Größe in etwa der eines Maulwurfs glich. Buffon stellte dem „Tendrac“ den „Tanrec“ zur Seite, letzterer unterscheidet sich von ersterem durch seine allgemein größeren Ausmaße und wird heute mit dem Großen Tenrek (Tenrec ecaudatus) in Verbindung gebracht. Seine Ausführungen zum „Tendrac“ und zum „Tanrec“ stützte Buffon auf zwei Tiere aus der Sammlung des damaligen Cabinet du roi, aus dem das Muséum national d’histoire naturelle in Paris hervorging, und welche über einen Händler aus Lyon in diese gelangten.[46] Das Individuum, das Buffon dem „Tendrac“ zuwies, besaß späteren Angaben zufolge eine Körperlänge von etwa 16 cm.[43] Johann Christian von Schreber legte im Jahr 1777 in seinem umfassenden Werk Die Säugthiere in Abbildungen nach der Natur mit Beschreibungen eine deutschsprachige Beschreibung des „Tendrac“ vor und berief sich dabei auf Buffon. In einem Zusatzband fügte er auch eine Abbildung des Tieres bei, die stark der von Buffon ähnelt. Zusätzlich ist dort aber noch die wissenschaftliche Bezeichnung Erinaceus setosus vermerkt, weswegen Schreber als wissenschaftlicher Erstbeschreiber des Großen Igeltenreks gilt. Schreber stellte die Art dabei in den Verwandtschaftskreis der europäischen Igel, das Artepitheton leitet sich von dem lateinischen Wort saetōsus für „borstig“ oder „haarig“ her und verweist auf das Stachelkleid. Als Herkunftsgebiet gab Schreber Madagaskar allgemein an, er nahm aber auch ein Vorkommen in Ostindien an.[47]
Unterarten und Synonyme

Neben der von Schreber benannten Nominatform Erinaceus setosus waren im Laufe der Forschungsgeschichte teilweise verschiedene Unterarten anerkannt. Étienne Geoffroy Saint-Hilaire führte im Jahr 1839 die Form Ericulus nigrescens ein, die er als extrem dunkelhaarig beschrieb. Dabei verwies er auf einen sechs Jahre zuvor veröffentlichten Brief von William Telfair, in dem dieser der Zoological Society of London einen Tenrek vorstellte, den er für ein Jungtier des Großen Tenrek hielt.[48] Allerdings hatte bereits 1838 William Charles Linnaeus Martin das Individuum mit dem von ihm benannten Kleinen Igeltenrek (Echinops telfairi) in Verbindung gebracht.[42] Im Jahr 1926 erkannte Oldfield Thomas mit Ericulus setosus setosus und Ericulus setosus nigrescens zwei Unterarten an und führte mit Ericulus setosus melantho eine dritte ein, die wie letztere dunkelhaarig aber deutlich größer als die beiden anderen war.[49] Die drei Unterarten wurden später teilweise auch angeführt,[5] sie sind aber heute nicht allgemein anerkannt. Eine vor allem im 19. Jahrhundert gebrauchte Form ist Centenes spinosus, welche von Anselme Gaëtan Desmarest im Jahr 1820 unter Berufung auf Buffons „Tendrac“ etabliert worden war.[50] Auch diese gilt heute als synonym zu Setifer setosus.[51]
Setiger, Ericulus und Dasogale – Zur Gattungsproblematik

Der Verweis des Großen Igeltenreks zu den Igeln war im 18. Jahrhundert durchaus üblich, erst im 19. Jahrhundert erfolgte eine stärkere Differenzierung der verschiedenen Formen. Der heute gültige Gattungsname Setifer stammt von Ludwig Friedrich von Froriep aus dem Jahr 1806. Froriep hatte das im gleichen Jahr von André Marie Constant Duméril erstellte Werk Zoologie analytique aus dem Französischen ins Deutsche übersetzt, darin fügte er die Bezeichnung Setifer hinzu und setzte sie mit Erinaceus setosus in eine eindeutige Beziehung.[52] Beides ist im Originalwerk nicht zu finden.[53] Dem Wort Setifer liegt (wie setosus) der lateinische Begriff saeta für „Borste“ zugrunde. Bereits 1803 war von Geoffroy Saint-Hilaire das prinzipiell ähnliche Wort Setiger genutzt worden um die heutigen Igeltenreks von den europäischen Igeln abzugrenzen.[54] Setiger fand daraufhin im Verlauf des 19. Jahrhunderts vielfach Verwendung in der wissenschaftlichen Literatur, meist stand der Gattungsname in Verbindung mit dem Großen Igeltenrek.[43] Nach Geoffroy Saint-Hilaire schloss Setiger aber nicht nur den Großen Igeltenrek, sondern auch den Großen Tenrek und den Eigentlichen Streifentenrek (Hemicentetes semispinosus) ein. In dieser erweiterten Sichtweise und bezogen auf alle damals bekannten Tenreks hatte bereits Georges Cuvier die Bezeichnung drei Jahre zuvor im ersten Teil seines Werkes Leçons d'anatomie comparée genutzt.[55] Aufgrund der Ungenauigkeiten bei der Namensgebung im Bezug auf den Großen Igeltenrek, dem Großen Tenrek und dem Eigentlichen Streifentenrek wurde im Jahr 1930 vorgeschlagen, Setiger als ein direktes Synonym für Tenrec einzustufen.[56]
Eine weitere, im 19. und teilweise bis weit in das 20. Jahrhundert häufig gebrauchte Bezeichnung ist Ericulus. Diese wurde im Jahr 1837 wiederum von Geoffroy Saint-Hilaire eingeführt. Er verband sie aber zunächst nicht mit einer bestimmten Art, schloss in die Gattung aber Tenreks mit kleinem Wuchs, kräftigen Krallen, kurzem Schwanz und langgestrecktem Schädel sowie mit insgesamt vier Schneidezähnen und sechs Mahlzähnen im Oberkiefer ein.[57] In einem zwei Jahre später erschienenen umfangreicheren Aufsatz benannte Geoffroy Saint-Hilaire ausdrücklich Ericulus spinosus und Ericulus nigrescens, also heutige Synonymformen des Großen Igeltenreks.[48] Teilweise stand neben dem Großen auch der Kleine Igeltenrek innerhalb der Gattung Ericulus, wobei letzterer gelegentlich auf Ebene der Untergattung (Echinops) abgetrennt wurde.[58][59] Beide Formen gelten aber heute als getrennt, ebenso setzte sich in der zweiten Hälfte des 20. Jahrhunderts die Gattungsbezeichnung Setifer gegenüber Ericulus durch.[5][60][51]
Im Jahr 1928 benannte Guillaume Grandidier die Gattung Dasogale mit der einzigen Art Dasogale fontoynonti zu Ehren des damaligen Vorsitzenden der madagassischen Akademie der Wissenschaften Antoine Maurice Fontoynont. Gattung und Art basierten auf zwei Individuen, von denen der Holotyp aus einem beschädigten Schädel samt Unterkiefer, einigen Extremitätenknochen und Hautstücken mit Fell besteht. Als Herkunft gab Grandidier nur vage grande forêt orientale („großer östlicher Wald“) an. Er beschrieb das Gebiss mit insgesamt 32 Zähnen als ähnlich zu dem des Kleinen Igeltenreks, da je Kieferhälfte nur zwei Molaren ausgebildet sind. Jedoch bemerkte er Unterschiede im Gebissaufbau, etwa in der speziellen Gestaltung der Mahlzähne. Daher befürwortete er im Bezug auf die abweichende Morphologie und die unterschiedliche Verbreitung eine eigenständige Stellung, sah aber Dasogale in der Nähe des Kleinen Igeltenreks.[61] Weitere Exemplare von Dasogale wurden in der Folgezeit nicht entdeckt, allerdings zweifelte kaum ein Forscher an der Existenz des Taxons.[5][7][60] Anfang der 1980er Jahre erkannte Walter Poduschka bei der Untersuchung des Originalmaterials, dass es sich bei den beiden Exemplaren um Jungtiere des Großen Igeltenreks handelt. Hierfür spricht unter anderem das vordere Gebiss, das aus Milchzähnen besteht und etwa gleich große Schneidezähne besitzt. Zudem war beim Holotyp der dritte Molar noch nicht durchgebrochen, was etwa an der Stellung des letzten Zahns der Zahnreihe erkennbar ist: bei allen Tenreks steht der letzte Mahlzahn deutlich zu den anderen Zähnen eingedreht, bei Dasogale ist dies jedoch nicht der Fall. Schlussendlich indizieren auch andere Merkmale wie etwa die Beschaffenheit der Stacheln, dass die beiden Individuen mit dem Großen Igeltenrek identisch sind.[10] Unabhängig von Poduschkas Untersuchung kam Ross D. E. MacPhee nur wenige Jahre später zu einem übereinstimmenden Ergebnis bei nahezu identischer Beweisführung.[62]
Stammesgeschichte
Allgemein sind Fossilfunde von Tenreks eher selten. Vom Großen Igeltenrek liegen einige vereinzelte subfossile Reste vor. Zu den bedeutendsten gehören jene der Ankilitelo-Höhle, die sich am Südrand des Mikoboka-Plateau nördlich von Toliara im südwestlichen Madagaskar befindet. Hier wurde seit dem Untersuchungsbeginn 1994 eine sehr reichhaltige Fauna ausgegraben. Sie umfasst vor allem Knochen und Zähne von verschiedenen Primaten, Fleder- und Nagetieren, es sind aber auch rund ein halbes Dutzend Arten an Tenreks überliefert. Der Große Igeltenrek stellt dabei den am häufigsten nachgewiesenen Vertreter der Familie dar, er ist mit zahlreichen Kiefer, Zahn- und Schädelresten sowohl ausgewachsener als auch juveniler Tiere belegt. Die Funde werden auf ein Alter von etwa 510 bis 630 Jahren datiert.[63] Die Art ist zudem aus der Andrahomana-Höhle im äußersten Südosten Madagaskars dokumentiert. Die Höhle liegt eingebettet in den Eolianit-Aufschlüssen der Steilküste und enthält ebenfalls ein ungemein reiches Fossilmaterial. Sie wird bereits seit dem Ende des 19. Jahrhunderts wissenschaftlich untersucht. Die Schichtenfolge und die daraus geborgenen Funde decken den Zeitraum der letzten rund 8.700 Jahre ab.[64] Beide Fundregionen zeichnen sich durch trockene Klimate aus, im Bereich der Andrahomana-Höhle ist aber auch noch ein gewisser Einfluss der feuchten Bedingungen des östlichen Madagaskars bemerkbar.[63][64][65]
Bedrohung und Schutz
Größere Bedrohungen für den Bestand des Großen Igeltenreks sind nicht bekannt. Sein Fleisch wird als Nahrungsressource genutzt und auf Märkten und in Restaurants verkauft. Dadurch kann es durch Überjagung lokal zu stärkeren Bestandsrückgängen kommen. In einigen Regionen gibt es aber ein Tabu, das die Jagd auf die Tiere verbietet.[66] Die IUCN listet die Art als „nicht gefährdet“ (least concern). Der Große Igeltenrek ist in zahlreichen Naturschutzgebieten anwesend, unter anderem im Nationalpark Montagne d’Ambre, im Nationalpark Ankarafantsika im Nationalpark Ranomafana, im Nationalpark Zombitse-Vohibasia, im Nationalpark Andringitra, im Nationalpark Andohahela und im Nationalpark Isalo.[16]
Literatur
- J. F. Eisenberg und Edwin Gould: The Tenrecs: A Study in Mammalian Behavior and Evolution. Smithsonian Institution Press, 1970, S. 1–138
- Kathryn M. Everson, Voahangy Soarimalala, Steven M. Goodman und Link E. Olson: Multiple loci and complete taxonomic sampling resolve the phylogeny and biogeographic history of tenrecs (Mammalia: Tenrecidae) and reveal higher speciation rates in Madagascar’s humid forests. Systematic Biology 65 (5), 2016, S. 890–909 doi: 10.1093/sysbio/syw034
- Nick Garbutt: Mammals of Madagascar. A complete guide. Yale University Press, 2007, S. 1–304 (S. 32–56)
- Paulina D. Jenkins: Tenrecidae (Tenrecs and Shrew tenrecs). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 134–172 (S. 163–164) ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. The Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9
Einzelnachweise
- Danielle L. Levesque, D. Rakotondravony und Barry G. Lovegrove: Home range and shelter site selection in the greater hedgehog tenrec in the dry deciduous forest of Western Madagascar. Journal of Zoology 287, 2012, S. 161–168
- Danielle L. Levesque, O. M.A. Lovasoa, S. N. Rakotoharimalala und Barry G. Lovegrove: High mortality and annual fecundity in a free-ranging basal placental mammal, Setifer setosus (Tenrecidae: Afrosoricida). Journal of Zoology 291, 2013, S. 205–212
- Oldfield Thomas: On the insectivorous genus Echinops, Martin, with notes oh the dentition of allied species. Proceedings of the.Zoological Society of London, 1892, S. 500–505 ()
- Walter Poduschka: Hyperthelie, Wurfgröße und Trächtigkeitsdauer bei der Unterfamilie Tenrecinae Cabrera, 1925 (Mammalia: Insectivora: Tenrecidae), nebst Bemerkungen zur Längsstreifung der Gattung Hemicentetes. Contributions to Zoology 66 (2), 1996, S. 119–128
- Konrad Herter: Über die Borstenigel von Madagaskar (Tenrecinae). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin N. F. 2, 1962, S. 5–37
- Edwin Gould und John F. Eisenberg: Notes on the biology of the Tenrecidae. Journal of Mammalogy 47 (4), 1966, S. 660–686
- J. F. Eisenberg und Edwin Gould: The Tenrecs: A Study in Mammalian Behavior and Evolution. Smithsonian Institution Press, 1970, S. 1–138
- Nick Garbutt: Mammals of Madagascar. A complete guide. Yale University Press, 2007, S. 1–304 (S. 32–56)
- Paulina D. Jenkins: Tenrecidae (Tenrecs and Shrew tenrecs). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 134–172 (S. 163–164) ISBN 978-84-16728-08-4
- Walter Poduschka und Christl Poduschka: Die taxonomische Zugehörigkeit von Dasogale fontoynonti G. Grandidier, 1928. Sitzungsberichte der Österreichischen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse, Abteilung 1, 191, 1982, S. 253–264
- Wilhelm Leche: Zur Entwicklungsgeschichte des Zahnsystems der Säugetiere, zugleich ein Beitrag zur Stammesgeschichte dieser Tiergruppe. Zweiter Teil: Phylogenie. Zweites Heft: Familien der Centetidae, Solenodontidae und Chrysochloridae. Zoologica 20, 1906/1908, S. 1–157 ()
- Justine A. Salton und Eric J. Sargis: Evolutionary morphology of the Tenrecoidea (Mammalia) carpal complex. Biological Journal of the Linnean Society, 93, 2008, S. 267–288
- Justine A. Salton und Eric J. Sargis: Evolutionary Morphology of the Tenrecoidea (Mammalia) Forelimb Skeleton. In: E.J. Sargis und M. Dagosto (Hrsg.): Mammalian Evolutionary Morphology: A Tribute to Frederick S. Szalay, Springer Science, 2008, S. 51–71
- Justine A. Salton und Frederick S. Szalay: The Tarsal Complex of Afro-Malagasy Tenrecoidea: A Search for Phylogenetically Meaningful Characters. Journal of Mammalian Evolution 11 (2), 2004, S. 73–104
- Voahangy Soarimalala: Les Afrosoricides de la forêt sèche malgache. Afrotherian Conservation 8, 2011, S. 4–9
- P. J. Stephenson, Voahangy Soarimalala und Steven M. Goodman: Setifer setosus. The IUCN Red List of Threatened Species 2016. e.T40594A97203842 (); zuletzt abgerufen am 3. April 2017
- Konrad Herter: Untersuchungen an lebenden Borstenigeln (Tenrecinae), 4: Über das Verhalten und die Aktivitätsrhythmik eines Setifer setosus (Schreber) in Gefangenschaft. Zoologische Beiträge 10, 1964, S. 161–187
- Peter J. Stephenson, Paul A. Racey und Félix Rakotondraparany: Maintenance and reproduction of tenrecs (Tenrecidae) at Parc Tsimbazaza, Madagascar. International Zoo Yearbook 33, 1994, S. 194–201
- A. L. Rand: On the Habits of Some Madagascar Mammals. Journal of Mammalogy 16 (2), 1935, S. 89–104
- J. J. Petter und A. Petter-Rousseaux: Notes biologiques sur les Centetinae. La Terre et la Vie 17 (1), 1963, S. 66–80
- Melanie Dammhahn, Voahangy Soarimalala und Steven M. Goodman: Trophic Niche Differentiation and Microhabitat Utilization in a Species-rich Montane Forest Small Mammal Community of Eastern Madagascar. Biotropica 45 (1), 2013, S. 111–118
- C. Kayser: Consommation d’oxygène et temperature central au cours de l’hiver austral de deux insectivores de Madagascar, Centetes ecaudatus et Setifer setosus. Comptes rendus des séances de la Société de Biologie 154, 1960, S. 1873–1876 ()
- G. Hildwein: Evolution saisonnière de la termorégulation chez l'Ericulus (Setifer setosus) (Mammifère Insectivore). Comptes Rendus des Séances de la Société de Biologie et de ses Filiales 158, 1964, S. 1580–1583 ()
- Danielle L. Levesque und Barry G. Lovegrove: Increased homeothermy during reproduction in a basal placental mammal. The Journal of Experimental Biology 217, 2014, S. 1535–1542
- Danielle L. Levesque, Kerileigh D. Lobban und·Barry G. Lovegrove: Effects of reproductive status and high ambient temperatures on the body temperature of a free‑ranging basoendotherm. Journal of Comparative Physiology B, 2014 doi:10.1007/s00360-014-0858-4
- Barry G. Lovegrove, Cindy Canale, Danielle Levesque, Gerhard Fluch, Milada Řeháková-Petrů und Thomas Ruf: Are Tropical Small Mammals Physiologically Vulnerable to Arrhenius Effects and Climate Change? Physiological and Biochemical Zoology 87 (1), 2014, S. 30–45
- Walter Poduschka: Das Paarungsverhalten des Großen Igel-Tenrek (Setifer setosus, Froriep 1806) und die Frage des phylogenetischen Alters einiger Paarungseinzelheiten. Zeitschrift für Tierpsychologie 34, 1974, S. 345–358
- John F. Eisenberg: Tenrecs and solenodons in captivity. International Zoo Yearbook 15, 1975, S. 6–12
- Richard Weigl: Longevity of mammals in captivity; from the Living Collections of the world A list of mammalian longevity in captivity. Kleine Senckenberg-Reihe, Band 48, 2005, S. 1–214 (S. 34)
- Steven M. Goodman, O. Langrand und B. P. N. Rasolonandrasana: The food habits of Cryptoprocta ferox in the high mountain zone of the Andringitra Massif, Madagascar (Carnivore, Viverridae). Mammalia 61 (2), 1997, S. 185–192
- Luke Dollar, Jörg U. Ganzhorn und Steven M. Goodman: Primates and other prey in the seasonally variable diet of Cryptoprocta ferox in the deciduous forest of Western Madagascar. In: S. Gursky und K. A. Nekaris (Hrsg.): Predation on primates. New York, 2007, S. 63–76
- Jean-Claude Beaucournu, H. Rico Randrenjarison Andriniaina und Steven M. Goodman: Puces (Insecta: Siphonaptera) d’Ambohitantely, Madagascar: Spécificité et phénologie. Malagasy Nature 9, 2015, S. 39–48
- J.-B. Duchemin: Leptopsyllines from Madagascar (Insecta: Siphonaptera: Ceratophyllidae): Description of five new species of Paractenopsyllus Wagner, (1938). Parasite 11, 2004, S. 59–70
- Steven M. Goodman, H. Rico Randrenjarison Andriniaina, Voahangy Soarimalala und Jean-Claude Beaucournu: The Fleas of Endemic and Introduced Small Mammals in Central Highland Forests of Madagascar: Faunistics, Species Diversity, and Absence of Host Specificity. Journal of Medical Entomology 52 (5), 2015, S. 1135–1143
- Mireille Harimalala, Soanandrasana Rahelinirina und Romain Girod: Presence of the Oriental Rat Flea (Siphonaptera: Pulicidae) Infesting an Endemic Mammal and Confirmed Plague Circulation in a Forest Area of Madagasca. Journal of Medical Entomology, 2020, S. tjaa026 doi:10.1093/jme/tjaa026
- Malala N. Rakotomanga, Steven M. Goodman, Voahangy Soarimalala, Sebastien Boyer und Dmitry Apanaskevich: Les tiques dures (Acari : Ixodidae) ectoparasites de micromammifères non-volants dans la forêt d’Ambohitantely, Madagascar. Malagasy Nature 12, 2017, S. 59–67
- Lee Couch, Juha Laakkonen, Steven Goodman und Donald W. Duszynski: Two New Eimerians (Apicomplexa) from Insectivorous Mammals in Madagascar. Journal of Parasitology 97 (2), 2011, S. 293–296
- Mina K. Khoii, Elizabeth W. Howerth, Roy B. Burns, K. Paige Carmichael und Zoltan S. Gyimesi: Spontaneous Neoplasia in Four Captive Greater Hedgehog Tenrecs (Setifer setosus). Journal of Zoo and Wildlife Medicine 39 (3), 2008, S. 392–397
- Kathryn M. Everson, Voahangy Soarimalala, Steven M. Goodman und Link E. Olson: Multiple loci and complete taxonomic sampling resolve the phylogeny and biogeographic history of tenrecs (Mammalia: Tenrecidae) and reveal higher speciation rates in Madagascar’s humid forests. Systematic Biology 65 (5), 2016, S. 890–909 doi: 10.1093/sysbio/syw034
- Link E. Olson und Steven M. Goodman: Phylogeny and biogeography of tenrecs. In: Steven M. Goodman und Jonathan P. Benstead (Hrsg.): The natural history of Madagascar. University of Chicago Press, 2003, S. 1235–1242
- Matjaž Kuntner, Laura J. May-Collado und Ingi Agnarsson: Phylogeny and conservation priorities of afrotherian mammals (Afrotheria, Mammalia). Zoologica Scripta 40 (1), 2011, S. 1–15
- William Charles Linnaeus Martin: On a new genus of insectivorous Mammalia. Proceedings of the Zoological Society of London 6, 1838, S. 17–19 ()
- Fredericus Anna Jentink: On the hedgehogs from Madagascar. Notes from the Royal Zoological Museum at Leyden 1879, S. 137–151 ()
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part I London, 1883, S. 71 ()
- Étienne de Flacourt: Histoire de la grande isle Madagascar. Paris, 1661, S. 1–471 (S. 152) ()
- Georges-Louis Leclerc de Buffon: Histoire naturelle, générale et particulière., Tome Douzième. Paris, 1764, S. 440–441, 450 und Tafel 57 ()
- Johann Christian Daniel Schreber: Die Säugthiere in Abbildungen nach der Natur mit Beschreibungen. Dritter Teil. Erlangen, 1777, S. 583–584 () und Tafelband 1 Tafel 164 ()
- Étienne Geoffroy Saint-Hilaire: Tanrec. Cuv. Centetes. Illig. et Éricule. Ericulus. Is. Geoff. Magasin de Zoologie Serie 2 1, 1839, S. 1–37 ()
- Oldfield Thomas: On some new mammals from Madagascar. Journal of Natural History 17, 1926, S. 250–252
- Anselme Gaëtan Desmarest: Mammalogie ou description des espèces des Mammifères. Premiere Partie. Paris, 1820, S. 1–276 (S. 162) ()
- Don E. Wilson und DeeAnn M. Reeder: Mammal Species of the World. A taxonomic and geographic Reference. Johns Hopkins University Press, 2005 ()
- Ludwig Friedrich von Froriep: Dumeril, Analytische Zoologie. Aus dem Französischen mit Zusätzen. Weimar, 1806, S. 1–345 (S. 15) ()
- Constant Dumeril: Zoologie analytique: ou, Méthode naturelle de classification des animaux; rendue plus facile a l'aide de tableaux synoptiques. Paris, 1906, S. 1–344 (S. 14) ()
- Étienne Geoffroy Saint-Hilaire: Catalogue des mammifères du Muséum National d'Histoire Naturelle. Paris, 1803, S. 1–272 (S. 71) ()
- Georges Cuvier: Leçons d'anatomie Comparée. Tome 1. Paris, 1800, S. 1–522 (S. ??) ()
- C. W. Stiles und Samuel F. Stanley: Erinaceus Ecaudatus Designated Type of Setiger 1800. Journal of Mammalogy 11 (2), 1930, S. 226, doi:10.1093/jmammal/11.2.226
- Étienne Geoffroy Saint-Hilaire: Notice sur les mammiferes épineux de Madagascar. Comptes rendus hebdomadaires des séances de l'Académie des sciences 5, 1837, S. 372–374 ()
- P. Mathias und P. Rode: Contribution à l'etude des insectivores.1. Les insectivores à piquants. Bulletin de la Societété zoologique de France 55, 1930, S. 429–437 ()
- Gabriel Petit: Insectivora. In: A. Gruvel (Hrsg.): Faune des colonies Françaises. Tome Quatriène Paris, 1931, S. 570–575 ()
- Henri Heim de Balsac: Insectivores. In: R. Battistini und G. Richard-Vindard (Hrsg.): Biogeography and ecology in Madagascar. Den Haag, 1972, S. 629–660
- Guillaume Grandidier: Un nouveau type de Mammifere insectivore de Madagascar, le Dasogale fontoynonti G. Grand. Bulletin de l'Academie Malgache NS 11, 1928, S. 84–90
- Ross D. E. MacPhee: Systematic status of Dasogale fontoynonti (Tenrecidae, Insectivora). Journal of Mammalogy 68 (1), 1987, S. 133–135
- Kathleen M. Muldoon, Donald D. de Blieux, Elwyn L. Simons und Prithijit S. Chatrath: The Subfossil Occurrence and Paleoecological Significance of Small Mammals at Ankilitelo Cave, Southwestern Madagascar. Journal of Mammalogy 90 (5), 2009, S. 1111–1131
- D. A. Burney, N. Vasey, L. R. Godfrey, Ramilisonina, W. L. Jungers, M. Ramarolahy und L. Raharivony: New Findings at Andrahomana Cave, Southeastern Madagascar. Journal of Cave and Karst Studies 70 (1), 2008, S. 13–24
- Steven M. Goodman und William L. Jungers: Extinct Madagaskar. Picturing the island's past. University of Chicago Press, 2014, S 1–206 (S. 65–73 und 94–101)
- Julia P. G. Jones, Mijasoa M. Andriamarovololona und Neal Hockley: The Importance of Taboos and Social Norms to Conservation in Madagascar. Conservation Biology 22 (4), 2008, S. 976–986
Weblinks
- Setifer setosus in der Roten Liste gefährdeter Arten der IUCN 2017. Eingestellt von: P. J. Stephenson, Voahangy Soarimalala und Steven M. Goodman, 2015. Abgerufen am 03.04.2017.