Streifentenreks
Die Streifentenreks (Hemicentetes) sind eine Säugetiergattung aus der Unterfamilie der Igeltenreks (Tenrecinae) innerhalb der Familie der Tenreks (Tenrecidae). Ihr werden zwei Arten zugerechnet, der Eigentliche Streifentenrek und der Schwarzkopftenrek. Sie kommen ausschließlich auf Madagaskar vor und bewohnen dort tropische Regenwälder des Tief- und Gebirgslandes. Darüber hinaus treten sie auch in Offenlandschaften auf und dringen bis in menschliche Siedlungen vor. Neben ihrem igelartigen Aussehen zählen vor allem das gestreifte Fellmuster und die lange, ausgezogene Schnauze zu den charakteristischen Merkmalen der Tiere. Im Nacken ist eine Halskrause aus Stacheln mit Widerhaken ausgebildet, die aufgestellt werden kann und zur Verteidigung dient. Eine weitere Besonderheit findet sich in dem Stridulationsorgan am hinteren Rücken, das aus besonderen Stacheln ohne Widerhaken besteht. Mit diesem können verschiedene Töne zur innerartlichen Kommunikation erzeugt werden. Aufgrund dieser und einiger weiterer Eigenschaften zählen die Streifentenreks zu den am stärksten spezialisierten Vertretern der Tenreks.
| Streifentenreks | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Eigentlicher Streifentenrek (Hemicentetes semispinosus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Hemicentetes | ||||||||||||
| Mivart, 1871 |
Die Streifentenreks leben bodenbewohnend und graben unterirdische Baue als Unterschlupf. Sie orientieren sich mittels des Geruchssinns, sind aber auch zu einer einfachen Form der Echoortung befähigt. Die Hauptaktivitäten erfolgen im Südsommer oder während der Regenzeit. Im Südwinter beziehungsweise in der Trockenzeit fallen die Tiere aufgrund ihrer schwankenden Körpertemperatur in einen Torpor. Vom Eigentlichen Streifentenrek ist bekannt, dass er während der Sommermonate in Familiengruppen oder Kolonien lebt, was innerhalb der Tenreks eher selten auftritt. Der Zeitraum bildet gleichzeitig die Fortpflanzungsphase. Die Würfe können bis zu elf Jungtiere umfassen, deren Individualentwicklung sehr schnell verläuft. Die Ernährung der Streifentenreks basiert überwiegend auf Regenwürmer, von denen sie im Verlauf eines Tages die Menge ihres eigenen Körpergewichts vertilgen können.
Aus forschungsgeschichtlicher Sicht wurden die ersten Streifentenreks bereits in der zweiten Hälfte des 18. Jahrhunderts erwähnt und wenig später als eigenständige Art beschrieben. In der Regel wurden sie im 19. Jahrhundert aber in die gleiche Gattung wie der Große Tenrek verwiesen. Erst rund einhundert Jahre nach der Ersterwähnung, im Jahr 1871, etablierte St. George Mivart eine eigene Gattung für die Streifentenreks. Im Verlauf des 20. Jahrhunderts war teilweise mit dem Eigentlichen Streifentenrek nur eine Art anerkannt, während der Schwarzkopftenrek als Unterart angesehen wurde.
Merkmale
Habitus

Die Streifentenreks gehören zu den mittelgroßen Tenreks. Sie erreichen eine Kopf-Rumpf-Länge von 12 bis 19 cm und ein Körpergewicht von 70 bis 220 g. Die beiden bekannten Arten, der Eigentliche Streifentenrek (Hemicentetes semispinosus) und der Schwarzkopftenrek (Hemicentetes nigriceps), sind dabei annähernd gleich groß mit etwas größeren Durchschnittsmaßen bei ersterem. Typisch für die Streifentenreks ist ihr igelartiges Aussehen mit einem gedrungenen Körper und kurzem Stummelschwanz. Die Gliedmaßen sind vergleichsweise lang und mit kräftigen Zehen ausgestattet. Die Tiere haben kein durchgehendes Stachelkleid, sondern ein eher borstiges Fell, aus dem die Stacheln herausragen. Diese besitzen kleine Widerhaken und regenerieren bei Verlust. Die Grundfärbung am Rücken ist schwarz, sie hebt sich deutlich von der helleren Unterseite ab. Der dunkle Farbton am Rücken wird von drei helleren Streifen durchbrochen, welche beim Eigentlichen Streifentenrek gelblich und beim Schwarzkopftenrek weißlich getönt sind. Ein zusätzlicher heller Bereich befindet sich am Nacken. Die Stacheln in den helleren Farbstreifen weisen Längen von 13 bis 16 mm auf und sind somit durchschnittlich größer als die 8 bis 14 mm langen Stacheln der dunklen Bereiche. Hier kommen auch Haare und Borsten vor, die beim Schwarzkopftenrek eine dichte Unterwolle bilden, so dass er insgesamt weicher wirkt als der Eigentliche Streifentenrek. Die mit 22 bis 30 mm längsten Stacheln treten am Nacken auf. Dort formen sie eine markante Halskrause, die aufgestellt werden kann. Zusätzlich vermischen sich die langen Stacheln im Nacken mit einzelnen helleren und dunkleren Stacheln von 11 bis 16 mm Länge sowie mit Halbstacheln und Haaren, deren Länge zwischen 10 und 13 mm variiert.[1] Eine besondere Bildung findet sich am Hinterteil, wo einige kräftige Stacheln ein Stridulationsorgan bilden. Der Kopf der Streifentenreks ist ausgesprochen grazil. Er weist ebenfalls eine schwarze Färbung auf. Diese wird beim Eigentlichen Streifentenrek durch einen gelblichen Mittelstreifen halbiert, beim Schwarzkopftenrek fehlt eine entsprechende Musterung. Die Schnauze ist sehr lang ausgezogenen und nach vorn spitz zulaufend, sie endet in einer nackten Nase. Hervorzuheben sind weiter markante Vibrissen und die sehr kleinen Augen.[2][3][4][5][6] Weibchen besitzen insgesamt 18 bis 20 Zitzen.[7] Insgesamt wird das markante Streifenmuster teilweise als Warnhinweis gegenüber Beutegreifern interpretiert[3] oder als Mimikry angesehen, bei der die Streifentenreks die Jungtiere des sympatrisch vorkommenden Großen Tenreks (Tenrec ecaudatus) imitieren, die von ihrer Mutter stark beschützt werden.[7]
Schädel- und Gebissmerkmale
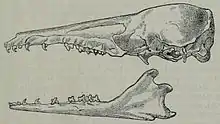
Der Schädel der Streifentenreks ist sehr grazil gebaut. Besondere Kennzeichen sind das ausgesprochen lange Rostrum und der kurze Hirnschädel. Das Verhältnis des vorderen zum hinteren Schädelabschnitt beträgt 0,61:0,39.[8] Das Rostrum ist in der Aufsicht schmal, es zieht in etwa auf der Höhe der hinteren Prämolar ein, wodurch der Schädel insgesamt nicht so zylindrisch wirkt wie beim Großen Tenrek. Des Weiteren fehlt eine deutliche Einschnürung hinter den Augen. In Seitenansicht besitzt der Schädel dagegen eine konische Gestalt. Das Nasenbein ist langgestreckt und verwächst zumeist nur in seinem hinteren Abschnitt. Ebenso ist das Gaumenbein deutlich ausgedehnt und reicht bis hinter den letzten Molaren. Der Mittelkieferknochen hat eine kleine Gestalt. Wie bei allen Tenreks bildet sich der Jochbogen nicht vollständig aus, der vordere Bogenansatz kragt aber nicht so weit nach außen wie beim Großen Tenrek. Am hinteren Schädelteil sind nur wenige markante Knochenrippeln als Muskelansatzstelle ausgebildet. Das Hinterhauptsbein ist steil aufgestellt. Das Gebiss besteht aus 40 Zähnen, die Zahnformel lautet folgendermaßen: . Auffallend sind die allgemein geringe Größe der Zähne und die großen Diastemata zwischen allen vorderen Zähnen einschließlich der Prämolaren, beide Merkmale bilden typische Eigenschaften von Nahrungsspezialisten, die sich hauptsächlich von Ameisen oder Würmern ernähren.[9] Die oberen Schneidezähne sowie der Eckzahn sind hakenartig nach hinten gekrümmt und weisen zusätzliche Höckerchen auf. Dagegen haben die unteren Schneidezähne eine schaufelartige Gestalt, während der Eckzahn hier wiederum seinem oberen Gegenstück gleicht. Im Gegensatz zum oberen überragt der untere Eckzahn aber die Schneidezähne des Unterkiefers. Die Mahlzähne verfügen in Übereinstimmung mit den anderen Tenreks über ein zalambdodontes Kauflächenmuster bestehend aus drei Haupthöckern.[10][8][11]

Skelettmerkmale
Die Wirbelsäule setzt sich aus 7 Hals-, 16 bis 17 Brust-, 4 bis 5 Lenden-, 2 Kreuzbein- und 9 bis 10 Schwanzwirbel zusammen.[8] Der Skelettbau zeigt einige Anpassungen an die grabende Lebensweise, die besonders deutlich an den Vorderbeinen hervortreten. Unter anderem finden sich diese in dem vergleichsweise langen und schmalen Schulterblatt, dass aber nicht so extrem gestreckt wirkt wie bei den Reiswühlern (Oryzorictes). Auch ist der Oberarmknochen kurz und robust, die innere Gelenkrolle am Ellenbogengelenk zieht seitlich sehr stark aus. Darüber hinaus besitzt die Elle gegenüber anderen Igeltenreks ein deutlich verlängertes oberes Gelenkende, das Olecranon. Es macht etwa 22 bis 23 % der Gesamtlänge des Knochens aus, innerhalb der Tenreks wird dieser Wert nur von den Reiswühler übertroffen. Beide Gelenkenden fungieren als Ansatzstelle für eine kräftige Unterarmmuskulatur. Die Handwurzelknochen sind kurz und breit gestaltet. Das Kahnbein bildet eine verwachsene Einheit mit dem Os centrale. Dadurch wird das Handgelenk stabiler und weniger beweglich. Ähnliches liegt von den Reiswühlern vor, doch ist hier zusätzlich noch das Mondbein mit einbezogen. Der zweite und der dritte Mittelhandknochen sind deutlich robuster gebaut, dagegen weist der erste gegenüber den kletternden Tenrekvertretern Kürzungen auf. Ebenso sind die Krallen des zweiten und dritten Fingerstrahls sehr robust und lang, weitaus länger als bei anderen Igeltenreks.[12][13]
Stridulationsorgan
Eine Besonderheit stellt das Stridulationsorgan dar, sich im hinteren Abschnitt der Rückenmittellinie befindet. Es besteht aus 7 bis 18, in drei Reihen angeordneten speziellen Stacheln. Die drei Reihen erstrecken sich laut einem vermessenen Exemplar über eine Länge von 1,68 cm entlang der Rückenmittellinie und über eine Breite von 0,86 cm, sie nehmen so insgesamt eine Fläche von rund 1 cm² ein. Im nackten Zustand ist die Fläche des Stridulationsorgans eindeutig sichtbar, sie enthält neben den Stacheln zusätzlich noch Schweißdrüsen. Die hier ausgebildeten Stacheln sind deutlich schwerer und etwas anders geformt als ihre Gegenstücke am Nacken. Sie besitzen zudem keine Widerhaken und regenerieren bei ausgewachsenen Tieren nicht.[14] Ihre Länge beträgt 8 bis 9 mm, ihre maximale Breite 0,8 bis 0,9 mm, die Stacheln der Außenreihen sind durchschnittlich dicker als die der Mittelreihe. Der größte Durchmesser wird im unteren Teil erreicht, nach oben laufen die Stacheln in einer Spitze aus, so dass ihr Umriss in etwa einer Flasche gleicht. In der unteren Hälfte werden die Stacheln von zehn dunkleren Linien umlaufen, ansonsten sind sie überwiegend hellbraun gefärbt. Die Stacheln durchdringen die Haut und reichen bis in den Bereich der Muskeln hinein. An den hier ansetzenden Basen werden die Stacheln von Kapseln aus Bindegewebe und von einem speziellen Fettgewebe umschlossen. Wie die Stacheln des Nackens können die Stacheln des Stridulationsorgans bewegt werden, wofür unter anderem der Musculus arrectores pilorum verantwortlich ist. Gleichzeitig stecken die Bindegewebskapseln im Musculus cutaneus trunci, einem der Hautmuskel, so dass hier ein stark miteinander verflochtenes Muskelgewebe entsteht.[15][16][1][17]
Durch Kontraktion der verschiedenen Muskelbereiche können die Stacheln im Uhrzeiger- oder entgegen dem Uhrzeigersinn rotiert werden. Die Drehrichtung ist veränderbar, sie verhält sich aber für beide Körperseiten gegenläufig zueinander. Durch die Bewegungen berühren sich die Stacheln und erzeugen rasselnde Klänge. Die so produzierten Töne liegen im höheren Frequenzbereich zwischen 2 und 80 kHz, mitunter erreichen sie bis zu 200 kHz. Innerhalb dieser Spanne bewegen sich verschiedene Maxima zwischen 20 und 45 sowie 50 und 150 kHz, die Lautstärke beträgt bis zu 63 db. Die Töne halten zwischen 7 und 100 ms an, während einer etwa zwei minütigen Sequenz werden so bis zu 70 Töne erzeugt. Die beiden Arten der Streifentenreks unterscheiden sich etwas in der Anzahl und Dauer der Tonimpulse, da der Schwarzkopftenrek durchschnittlich längere Töne erzeugt als der Eigentliche Streifentenrek. Die Tiere selbst können nur einen eingeengten Bereich ihrer Stridulationsklänge wahrnehmen, ihre optimale Hörfrequenz reicht von 10 bis 15 beziehungsweise bis 20 kHz. Dies ist aber ausreichend für eine Übertragungsreichweite von 4 bis maximal etwa 10 m.[18][14][16][3] Die Befähigung der Streifentenreks zu stridulieren wurde erstmals 1935 von Austin Loomer Rand beschrieben.[19]
Verbreitung und Lebensraum
Die Streifentenreks sind endemisch auf Madagaskar verbreitet. Beide Arten kommen im östlichen Inselteil vor. Der Schwarzkopftenrek ist dabei eher auf die zentral- bis südöstlichen Bereiche beschränkt, während der Eigentliche Streifentenrek ein in Nord-Süd-Richtung ausgedehnteres Verbreitungsgebiet aufweist. Die Tiere bewohnen vorwiegend tropische Regenwälder, sie sind aber auch in offeneren Landschaften und in anthropogen beeinflussten Gebieten wie Landwirtschaftsflächen oder Gärten anzutreffen und dringen teilweise bis in menschliche Siedlungen vor. In einigen Regionen wie dem Andringitra-Gebirge oder den Bergländern um Ivohibe in der Provinz Fianarantsoa kommen beide Arten allopatrisch vor. Unter diesen Umständen bevorzugt der Schwarzkopftenrek höher Lagen bis oberhalb der Baumgrenze, der Eigentliche Streifentenrek die Tiefländer und mittleren Gebirgslagen. Lediglich im 1550 m hoch gelegenen Waldgebiet von Mahatsìnjo südöstlich von Tsinjoarivo in der Provinz Antananarivo konnte bisher eine sympatrische Verbreitung der beiden Arten beobachtet werden.[20][21] Allgemein gelten sowohl der Schwarzkopftenrek als auch der Eigentliche Streifentenrek als relativ häufig.[5][6][22][23]
Lebensweise
Territorial- und Sozialverhalten
Die Lebensweise der Streifentenreks ist gut untersucht. Sie sind bodenlebend und Sohlengänger, die sich im Kreuzgang fortbewegen und auch mit alternierenden Bewegungen der Gliedmaßen klettern und schwimmen können, es aber selten tun.[24] In den Aktivitätszeiten unterscheiden sich die beiden Arten, da der Schwarzkopftenrek strikt nachtaktiv ist, der Eigentliche Streifentenrek aber auch verstärkt am Tage auftritt. In den Ruhephasen ziehen sich die Tiere in selbst gegrabene unterirdische Baue zurück, sie schlafen dann zusammengerollt mit dem Kopf am Bauch, so dass die Hauptlast des Gewichts auf den Füßen zu tragen kommt, teilweise liegen sie auch seitlich. Die Baue sind bei Einzeltieren etwa 45 cm lang und reichen rund 7,5 cm unter die Erde. Die Tiere heben ihre Baue mit den breiten Vorderpfoten aus, wobei hier ebenfalls wechselnde Armbewegungen vorherrschen, der dabei anfallende Aushub wird mit den Hinterbeinen weggedrückt. Ab einer Länge von etwa 30 cm beginnen die Tiere Pflanzenmaterial für ein Nest mit dem Maul heranzutransportieren. Der einzelne Eingang wird ebenfalls mit Pflanzenteilen abgedeckt. Vom Eigentlichen Streifentenrek ist bekannt, dass er in der Fortpflanzungsphase größere Familiengruppen und Kolonien bestehend aus erwachsenen Tieren beiderlei Geschlechts und mehreren Generationen an Jungtieren bildet. Größere Gruppen können bis zu zwei Dutzend Individuen umfassen. Diese Gruppen nutzen dann komplexere Tunnelsysteme mit Längen von bis zu 3 m. In der Winterzeit zerfallen die Gruppen aber dann wieder.[14][3][4]

Sozialkontakte zwischen Artgenossen bestehen in der Regel aus gegenseitigem Beschnüffeln. Sie verlaufen beim Eigentlichen Streifentenrek deutlich friedlicher ab. Antagonistisches Verhalten drückt sich allgemein in dem Aufrichten der Stacheln am Nacken und am Körper aus. Vor allem die Nackenstacheln können als Indikator für den Grad der Erregung angesehen werden. Leicht aufgestellte Stacheln zeigen meist aufmerksame Tiere an während senkrecht stehende oder nach vorn übergekippte Stacheln stark aggressives Verhalten signalisieren. Dieses Stadium ist zudem mit typischen Auf- und Abwärtsbewegungen des Kopfes verbunden, wodurch die Stacheln einen potentiellen Kontrahenten aufgrund ihrer leichten Ablösbarkeit vom Körper und der kleinen Widerhaken verletzen können. Tiere, die in ihrem Bau gestört werden, richten meist ihre Stacheln direkt vollständig auf. Begegnungen mit potentiellen Fressfeinden führen überwiegend zur Flucht, im Gegensatz zum Großen (Setifer setosus) und zum Kleinen Igeltenrek (Echinops telfairi) können sich die Streifentenreks nicht zu einer Kugel zusammenrollen. Ebenso fehlen das für andere Tenreks typische Aufreißen des Maules als Drohgebärde oder Bisse zur Verteidigung, was beides bei den eher kleinen Zähnen wenig effektiv ist.[3][4]
Die Streifentenreks besitzen einen guten Geruchssinn, der unter anderem der Orientierung und der Nahrungssuche dient. In unbekannten Gebieten schnüffeln die Tiere in der Luft und am Boden, was teilweise vier- bis fünfmal innerhalb einer Sekunde geschieht.[15] Er wird auch für die innerartliche Kommunikation eingesetzt, etwa beim Markieren der Eingänge der Baue. Darüber hinaus verfügen die Streifentenreks über eine Reihe von Lauten, darunter Grunz-, Quietsch- und Zwitschertöne. Andere Laute werden aber offensichtlich ebenfalls bei der Orientierung in unübersichtlichem Gelände oder bei Nacht eingesetzt. Hierzu gehören hochfrequente Zungenklicks im Bereich von 11 bis 16 kHz und einer Dauer von 0,2 bis 0,7 ms, die als Signale für eine Echoortung fungieren.[18] Teilweise wurde angenommen, dass auch die Stridulation der Echoortung dient, dies ließ sich in Experimenten aber nicht bestätigen. Die Funktion der Stridulation ist nicht eindeutig, sie erfolgt vor allem bei Aufmerksamkeit und Erregung, verläuft aber unabhängig von dem Aufrichten der Stacheln im Nacken. Die Tiere stridulieren etwa bei allgemeiner Aktivität, bei der Nahrungssuche, während Sozialkontakten oder beim Geschlechtsakt und bei Flucht. Die Rückenstacheln rasseln jedoch nicht während der Nahrungsaufnahme oder beim Putzen. Möglicherweise übertragen die Rasselgeräusche wichtige Informationen über den Absender an andere Artgenossen. Experimente mit Mutter- und Jungtieren zeigten, dass stridulierende Tiere sich häufig weiter entfernt voneinander aufhielten als solche, die keine Rasselgeräusche von sich gaben. Außerdem werden Jungtiere stimuliert, sich in die Richtung des Senders der Stridulationen zu bewegen, sie unterscheiden aber den Experimenten zufolge nicht zwischen der Stridulation der Mutter und anderen Artgenossen. Ein weiterer Grund für das Rasseln besteht eventuell in einem Warnsignal an Fressfeinde.[3][4]
Ernährung und Energiehaushalt
Die Nahrung der Streifentenreks besteht aus Wirbellosen, der Großteil wird über Regenwürmer abgedeckt, seltener fressen sie auch Larven von Insekten. Allerdings besteht eine trophische Differenz zwischen dem Schwarzkopftenrek und dem Eigentlichen Streifentenrek im Gebiet ihres gemeinsamen Auftretens bei Tsinjoarivo wie Isotopenuntersuchungen an Tieren aus der Region ergaben. Demnach ernährt sich ersterer gemäß den Untersuchungen häufiger von Sekundärkonsumenten, während letzterer ein für unterirdisch lebende Tiere typisches Isotopenspektrum aufweist.[25] Die Suche nach Nahrung erfolgt meist in Waldlichtungen mit lockeren und feuchten Böden Die Beute erschnuppern die Tiere mit ihrer langen Schnauze im Erdreich oder unter Blättern. Aufgespürte Regenwürmer werden mit dem Maul gepackt und durch ruckartige Rückwärtsbewegungen des Kopfes aus dem Boden gezogen. Anschließend wird er mit den Vorderpfoten seitlich gerieben und so von Erde befreit, was gleichzeitig der Ermittlung der Größe der Beute dient. Das typische Gebiss der Streifentenreks mit den kleinen Zähnen und den großen Zahnlücken stellt eine Anpassung an die wurmhaltige Nahrung dar.[9] Da das Gebiss kaum zum Kauen geeignet ist, wird die Nahrung meist vollständig verschlungen, zu große Würmer zerstückeln die Streifentenreks durch Trampeln mit den Füßen und Krallen. Die hakenartig zurückgebogenen Schneidezähne fixieren später die Nahrung im Maul und verhindern ein Herausgleiten. Die täglich aufgenommenen Menge an Nahrung ist relativ hoch und entspricht nahezu dem Körpergewicht eines Tieres.[24][14][3][4]
Wie bei anderen Tenreks auch ist die Körpertemperatur der Streifentenreks stark schwankend und zeigt sowohl im Tages- als auch im Jahresverlauf zyklische Veränderungen, sie ist abhängig von der Aktivität und passt sich den äußeren Bedingungen an. Vor allem im Südwinter von Juni bis Oktober liegt die Körpertemperatur nur wenige Grad über der Außentemperatur. In diesem Zeitraum tritt auch ein Torpor auf, der unter anderem mit der Verfügbarkeit an Regenwürmer als Hauptnahrung verbunden ist, deren Anzahl im trockeneren Winter stark zurückgeht. Beim Schwarzkopftenrek fällt der Torpor aufgrund seiner Verbreitung in durchschnittlich höheren Gebirgslagen intensiver aus als beim Eigentlichen Streifentenrek, bei dem die Starrephasen stärker von der individuellen Konstitution abhängig sind. Ersterer kann daher als obligatorischer Winterschläfer, letzterer als fakultativer angesehen werden. Die Stoffwechselrate ist im Vergleich zu ähnlich großen Säugetieren sehr gering und erreicht nur 49 bis 69 % des zu erwartenden Wertes, sie variiert ebenfalls im Jahresverlauf mit niedrigeren Werten im Winter als im Sommer. In Phasen erhöhten Stresses wie der Fortpflanzung steigt der Metabolismus zusätzlich an, was häufig mit einer Stabilisierung der Körpertemperatur während dieser Zeit in Zusammenhang gebracht wird. Auffallend sind ebenfalls jährlich starke Körpergewichtsschwankungen mit niedrigeren Gewichten im Winter und höheren im Sommer.[26][3][4]
Fortpflanzung
Die Fortpflanzung beschränkt sich bei beiden Vertretern der Streifentenreks auf den Südsommer von November bis Mai. Die Partnerwerbung besteht aus dem gegenseitigen Beschnüffeln verschiedener Körperteile in einer festen Reihenfolge, angefangen von der Nase über das Hinterteil bis zum Nacken, worauf der Geschlechtsakt erfolgt. Nach einer 55- bis 63-tägigen Tragzeit kommen zwei bis elf Jungtiere zur Welt, wobei die durchschnittliche Wurfgröße beim Eigentlichen Streifentenrek mit 6 bis 8 Jungtieren deutlich größer ist als beim Schwarzkopftenrek mit 3 bis 5. Der Nachwuchs kommt als Nesthocker zur Welt und verbleibt anfangs in einem Nest aus Pflanzenmaterial. Erst mit dem Öffnen der Augen nach rund 9 Tagen verlassen die Jungen erstmals den Wohnbau. Die Individualentwicklung der Jungen erfolgt sehr schnell, so dass Weibchen bereits nach viereinhalb bis fünf Wochen die Geschlechtsreife erlangen, was eine der kürzesten Generationenfolgen innerhalb der Säugetiere darstellt.[27] Beim Schwarzkopftenrek ist das Vatertier nicht in die Aufzucht des Nachwuchses involviert, weibliche Tiere mit Jungen verteidigen ihr Nest strikt. Im Gegensatz dazu zieht sich beim Eigentlichen Streifentenrek ein Muttertier in eine entferntere Ecke des Tunnelsystems zurück, verbleibt aber ansonsten in der Familiengruppe. Außerdem toleriert sie ab einem gewissen Alter der Jungen die Gegenwart von fremden Männchen. Möglicherweise finden sich die Ursachen für die Koloniebildung des Eigentlichen Streifentenreks in der durchschnittlich größeren Wurfgröße verbunden mit der geringeren innerartlichen Aggressivität.[14][3][4] Die Lebenserwartung frei lebender Streifentenreks ist unbekannt, das höchste bekannte Alter eines Tieres in Gefangenschaft betrug knapp drei Jahre.[28]
Systematik
Innere Systematik der Tenreks nach Everson et al. 2016[29]
|
Die Streifentenreks sind eine Gattung innerhalb der Familie der Tenreks (Tenrecidae). Sie bilden gemeinsam mit dem Großen Tenrek (Tenrec ecaudatus) sowie dem Großen (Setifer setosus) und dem Kleinen Igeltenrek (Echinops telfairi) die Unterfamilie der Igeltenreks (Tenrecinae), eine der drei Hauptlinien der Tenreks. Die anderen werden durch die sehr diversen Reiswühler (Oryzorictinae) und die monotypischen Erdtenreks (Geogalinae) repräsentiert, welche abweichend von den stacheligen Igeltenreks ein weichhaariges Fell aufweisen. Die Streifentenreks gehören zu den am stärksten spezialisierten Tenreks, was sich neben ihrer grabenden Lebensweise und der kurzen Generationenfolge, den aufgrund ihrer besonderen Ernährungsweise extrem kleinen Zähnen oder den Stacheln mit Widerhaken auch durch das Stridulationsorgan ausdrückt. Während widerhakenbesetzte Stacheln bei anderen Igeltenreks fehlen, kommt das Stridulationsorgan etwa in abgeschwächter Form auch beim Großen Tenrek vor, bei dem aber nur die Jungtiere über eine entsprechende Befähigung verfügen. Der Große Tenrek ist laut molekulargenetischen Analysen auch die Schwesterform der Streifentenreks.[30][31][29] Die beiden Linien trennten sich bereits im Unteren Miozän vor etwa 16,1 Millionen Jahren, die Aufspaltung von Hemicentetes vollzog sich dagegen erst im Pliozän vor 4,75 Millionen Jahren.[29]
Innerhalb der Streifentenreks werden heute zwei Arten unterschieden:[29]
- Eigentlicher Streifentenrek (Hemicentetes semispinosus (G. Couvier, 1798))
- Schwarzkopftenrek (Hemicentetes. nigriceps Günther, 1875)
Andere überwiegend im 19. Jahrhundert eingeführte Arten wie H. madagascariensis, H. variegatus oder H. buffoni erwiesen sich als synonym zum Eigentlichen Streifentenrek oder zum Schwarzkopftenrek.[4][32] Vor allem in den 1970er bis 1990er Jahren wurde der Schwarzkopftenrek teilweise als Unterart des Eigentlichen Streifentenreks angesehen.[4] Dagegen sprechen aber neben den auffallenden Abweichungen im äußeren Erscheinungsbild auch evidente Schädel- und Zahnmerkmale,[33][11] weswegen zahlreiche Forscher in dieser Zeit die Zusammenführung beider Arten nicht unterstützten.[34][3] Letztendlich führte die Entdeckung des sympatrischen Auftretens der beiden Formen bei Tsinjoarivo in der Provinz Antananarivo Ende der 1990er Jahre wieder zur Auftrennung der Streifentenreks in zwei Arten.[20]
Forschungsgeschichte

Die ersten Berichte über gestreifte Tenreks gehen bis in das 18. Jahrhundert zurück. Zuvor hatte Étienne de Flacourt 1661 in seiner Monographie Histoire de la grande isle Madagascar ein Tier namens „tendrac“ erwähnt sowie abgebildet, er hob auch das prinzipiell igelähnliche Erscheinungsbild des „tendrac“ hervor und bezog sich dadurch wohl auf den Großen Igeltenrek.[35] Rund 100 Jahre später, 1764, unterschied Georges-Louis Leclerc de Buffon im zwölften Band seines umfassenden Werkes Histoire naturelle, générale et particulière zwischen dem „tendrac“ und dem „tanrec“. Seinen Aussagen zufolge war der „tanrec“ deutlich größer als der „tendrac“ und hatte eine lang ausgezogene spitze Schnauze sowie eine markante Halskrause im Nacken. Auf zwei beigehefteten Tafeln bildete Buffon die Tiere ab, die sich durch eine einheitliche Farbgebung des Fells auszeichneten. In zwei Zusatzbänden der Histoire naturelle, die 1776 und 1789 veröffentlicht wurden, waren dagegen jeweils Abbildungen gestreifter Tenreks enthalten. Die Abbildung des Jahres 1776 interpretierte Buffon als Jungtier des „tanrec“ (le jeune tanrec), die des Jahres 1789 als ebensolches des „tendrac“.[36] Zwei Dekaden später, 1798, nutzte Georges Cuvier die Abbildung des Jahres 1776 zur Aufstellung einer eigenständigen weiteren Art der Tenreks und bezeichnete sie mit Erinaceus semispinosus.[37] Die gleiche Abbildung diente kurz darauf sowohl George Shaw zur Etablierung des wissenschaftlichen Artnamens Erinaceus madagascariensis[38] als auch Étienne Geoffroy Saint-Hilaire für die wissenschaftliche Bezeichnung Setiger variegatus.[39] Alle drei Namen beziehen sich auf den Eigentlichen Streifentenrek, wobei den Regeln der zoologischen Nomenklatur der ICZN folgend lediglich der erste Artname (Erinaceus semispinosus) Gültigkeit besitzt.[32]
In der Folgezeit ordneten zahlreiche Autoren die neue Form in die Nähe des Großen Tenreks, der im Verlauf des 19. und dem Beginn des 20. Jahrhunderts zumeist unter dem alten Synonymnamen Centetes geführt wurde.[40][41] Die Gattung Hemicentetes wurde dann schließlich im Jahr 1871 von St. George Mivart wissenschaftlich erstbeschrieben. Mivart verwies lediglich auf das markante äußere Erscheinungsbild der Streifentenreks und stellte darauf folgend die Skelettanatomie genauer vor. Er sah in seiner Veröffentlichung H. madagascariensis als Nominatform an. Darüber hinaus verglich er Hemicentetes mit dem Großen Tenrek (damals Centetes, heute Tenrec) und setzte gleichzeitig beide voneinander ab. Darauf bezieht sich auch der Gattungsname Hemicentetes (von altgriechisch ἥμι hemi für „halb“ und κεντητήϛ kentetes, hergeleitet von κεντητος kentetos für „gestickt“ oder κεντεω kenteo für „stechen“).[42][10]
Bedrohung und Schutz
Die beiden Arten der Streifentenreks sind weit verbreitet und gelten als relativ häufig. Zudem kommen sie auch mit anthropogen veränderten Landschaften wie Reisfeldern und Plantagen zurecht und dringen in menschliche Siedlungen vor. Größere Gefährdungen bestehen für die Bestände nicht, die IUCN stuft beide Arten als „nicht gefährdet“ (least concern) ein. Sie sind in mehreren Naturschutzgebieten vertreten. Dadurch sind sie von der Umwandlung der Regenwälder in Felder weniger betroffen als andere Tierarten.[22][23]
Literatur
- J. F. Eisenberg und Edwin Gould: The Tenrecs: A Study in Mammalian Behavior and Evolution. Smithsonian Institution Press, 1970, S. 1–138
- Kathryn M. Everson, Voahangy Soarimalala, Steven M. Goodman und Link E. Olson: Multiple loci and complete taxonomic sampling resolve the phylogeny and biogeographic history of tenrecs (Mammalia: Tenrecidae) and reveal higher speciation rates in Madagascar’s humid forests. Systematic Biology 65 (5), 2016, S. 890–909 doi: 10.1093/sysbio/syw034
- Nick Garbutt: Mammals of Madagascar. A complete guide. Yale University Press, 2007, S. 1–304 (S. 32–56)
- Christopher D. Marshall und John F. Eisenberg: Hemicentetes semispinosus. Mammalian Species 541, 1996, S. 1–4
- Ronald M. Nowak: Walker's Mammals of the World. The Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9
- Peter J. Stephenson: Streaked tenrecs, Hemicentetes. Afrotherian Conservation 5, 2007, S. 1–3
Einzelnachweise
- Olga V. Zherebtsova: Morphofunctional interpretation of the quills stridulating in tenrecs (Lipotyphla, Tenrecidae). Russian Journal of Theriology 5 (1), 2006, S. 1–11
- Konrad Herter: Über die Borstenigel von Madagaskar (Tenrecinae). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin N. F. 2, 1962, S. 5–37
- J. F. Eisenberg und Edwin Gould: The Tenrecs: A Study in Mammalian Behavior and Evolution. Smithsonian Institution Press, 1970, S. 1–138
- Christopher D. Marshall und John F. Eisenberg: Hemicentetes semispinosus. Mammalian Species 541, 1996, S. 1–4
- Nick Garbutt: Mammals of Madagascar. A complete guide. Yale University Press, 2007, S. 1–304 (S. 32–56)
- Peter J. Stephenson: Streaked tenrecs, Hemicentetes. Afrotherian Conservation 5, 2007, S. 1–3
- Walter Poduschka: Hyperthelie, Wurfgröße und Trächtigkeitsdauer bei der Unterfamilie Tenrecinae Cabrera, 1925 (Mammalia: Insectivora: Tenrecidae), nebst Bemerkungen zur Längsstreifung der Gattung Hemicentetes. Contributions to Zoology 66 (2), 1996, S. 119–128
- Wilhelm Leche: Zur Entwicklungsgeschichte des Zahnsystems der Säugetiere, zugleich ein Beitrag zur Stammesgeschichte dieser Tiergruppe. Zweiter Teil: Phylogenie. Zweites Heft: Familien der Centetidae, Solenodontidae und Chrysochloridae. Zoologica 20, 1906/1908, S. 1–157 ()
- Cyril Charles, Floréal Solé, Helder Gomes Rodrigues und Laurent Viriot: Under pressure? Dental adaptions to termitophagy and vermivory among mammals. Evolution 67 (6), 2013, S. 1792–1804
- St. George Mivart: On Hemicentetes, a new Genus of Insectivora, with some additional remarks on the osteology of that order. Proceedings of the Zoological Society of London, 1871, S. 58–79 ()
- Percy M. Butler: A Comparison of the Skulls and Teeth of the Two Species of Hemicentetes. Journal of Mammalogy 22 (1), 1941, S. 65–81
- Justine A. Salton und Eric J. Sargis: Evolutionary morphology of the Tenrecoidea (Mammalia) carpal complex. Biological Journal of the Linnean Society, 93, 2008, S. 267–288
- Justine A. Salton und Eric J. Sargis: Evolutionary Morphology of the Tenrecoidea (Mammalia) Forelimb Skeleton. In: E.J. Sargis und M. Dagosto (Hrsg.): Mammalian Evolutionary Morphology: A Tribute to Frederick S. Szalay, Springer Science, 2008, S. 51–71
- Edwin Gould und John F. Eisenberg: Notes on the biology of the Tenrecidae. Journal of Mammalogy 47 (4), 1966, S. 660–686
- J. J. Petter und A. Petter-Rousseaux: Notes biologiques sur les Centetinae. La Terre et la Vie 17 (1), 1963, S. 66–80
- Ernest Glen Wever und Paul N. Herman: Stridulation and Hearing in the Tenrec. Journal of Auditory Research 8, 1968, S. 39–42
- Hideki Endo, Daisuke Koyabu, Junpei Kimura, Felix Rakotondraparany, Atsushi Matsui, Takahiro Yonezawa, Akio Shinohara und Masami Hasegawa: A Quill Vibrating Mechanism for a Sounding Apparatus in the Streaked Tenrec (Hemicentetes semispinosus). Zoological Science 27, 2010, S. 427–432
- Edwin Gould: Evidence for echolocation in the Tenrecidae of Madagaskar. Proceedings of the American Philosophical Society 109 (6), 1965, S. 352–360
- A. L. Rand: On the Habits of Some Madagascar Mammals. Journal of Mammalogy 16 (2), 1935, S. 89–104
- Steven M. Goodman, Daniel Rakotondravony, Voahangy Soarimalala, J. B. Duchemin und J.-M. Duplantier: Syntopic occurrence of Hemicentetes semispinosus and H. nigriceps (Lipotyphla: Tenrecidae) on the central highlands of Madagascar. Mammalia 64, 2000, S. 113–116
- Steven M. Goodman, Daniel Rakotondravony, Marie Jeanne Raherilalao, Domoina Rakotomalala, Achille P. Raselimanana, Voahangy Soarimalala, Jean-Marc Duplantier, Jean-Bernard Duchemin und J. Rafanomezantsoa: Inventaire biologique de la Foret de Tsinjoarivo, Ambatolampy. Akon'ny Ala 27, 2000, S. 18–27
- P. J. Stephenson, Voahangy Soarimalala und Steven M. Goodman: Hemicentetes semispinosus. The IUCN Red List of Threatened Species 2016. e.T40593A97189434 (); zuletzt abgerufen am 2. März 2017
- P. J. Stephenson, Voahangy Soarimalala und Steven M. Goodman: Hemicentetes nigriceps. The IUCN Red List of Threatened Species 2016. e.T62011A97189172 (); zuletzt abgerufen am 2. März 2017
- Konrad Herter: Untersuchungen an lebenden Borstenigeln (Tenrecinae), 3: Über das Verhalten und die Lebensweise des Streifentanreks (Hemicentetes semispinosus G. Cuvier) in Gefangenschaft. Zoologische Beiträge 9 (2), 1963, S. 237–274
- Melanie Dammhahn, Voahangy Soarimalala und Steven M. Goodman: Trophic Niche Differentiation and Microhabitat Utilization in a Species-rich Montane Forest Small Mammal Community of Eastern Madagascar. Biotropica 45 (1), 2013, S. 111–118
- Peter J. Stephenson und Paul A. Racey: Seasonal variation in resting metabolic rate and body temperature of streaked tenrecs, Hemicentetes nigriceps and H. semispinosus (Insectivora: Tenrecidae). Journal of Zoology 232, 1994, S. 285–29
- Link E. Olson: Tenrecs. Current Biology 23 (1), 2013, S. R5–R8
- Peter J. Stephenson, Paul A. Racey und Félix Rakotondraparany: Maintenance and reproduction of tenrecs (Tenrecidae) at Parc Tsimbazaza, Madagascar. International Zoo Yearbook 33, 1994, S. 194–201
- Kathryn M. Everson, Voahangy Soarimalala, Steven M. Goodman und Link E. Olson: Multiple loci and complete taxonomic sampling resolve the phylogeny and biogeographic history of tenrecs (Mammalia: Tenrecidae) and reveal higher speciation rates in Madagascar’s humid forests. Systematic Biology 65 (5), 2016, S. 890–909 doi: 10.1093/sysbio/syw034
- Link E. Olson und Steven M. Goodman: Phylogeny and biogeography of tenrecs. In: Steven M. Goodman und Jonathan P. Benstead (Hrsg.): The natural history of Madagascar. University of Chicago Press, 2003, S. 1235–1242
- Matjaž Kuntner, Laura J. May-Collado und Ingi Agnarsson: Phylogeny and conservation priorities of afrotherian mammals (Afrotheria, Mammalia). Zoologica Scripta 40 (1), 2011, S. 1–15
- Don E. Wilson und DeeAnn M. Reeder: Mammal Species of the World. A taxonomic and geographic Reference. Johns Hopkins University Press, 2005 ()
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part I. London, 1883, S. 70–71 und 72–77 ()
- Henri Heim de Balsac: Insectivores. In: R. Battistini und G. Richard-Vindard (Hrsg.): Biogeography and ecology in Madagascar. Den Haag, 1972, S. 629–660
- Étienne de Flacourt: Histoire de la grande isle Madagascar. Paris, 1661, S. 1–471 (S. 152) ()
- Georges-Louis Leclerc de Buffon: Histoire naturelle, générale et particulière., Tome Douzième. Paris, 1764, S. 440–441 und Tafeln 56 und 57 (), Supplément, Tome troisième. Paris, 1776, Tafel 37 () und Supplément, Tome Septième. Paris, 1789, Tafel 76 ()
- Georges Cuvier: Tableau elementaire de l’histoire naturelle des animaux. Paris, 1798, S. 1–710 (S. 108) ()
- George Shaw: General zoology. Vol. 1 London, 1800, S. 1–552 (S. 548) ()
- Étienne Geoffroy Saint-Hilaire: Catalogue des mammifères du Muséum National d'Histoire Naturelle. Paris, 1803, S. 1–272 (S. 72) ()
- Anselme Gaëtan Desmarest: Mammalogie, ou, Description des espèces de mammifères. Paris, 1820, S. 1–276 (S. 161) ()
- Fredericus Anna Jentink: On the hedgehogs from Madagascar. Notes from the Royal Zoological Museum at Leyden 1879, S. 137–151 ()
- Johann Karl Wilhelm Illiger: Prodromus systematis mammalium et avium additis terminis zoographicis utriudque classis. Berlin, 1811, S. 1–301 (S. 124) ()
Weblinks
- Hemicentetes semispinosus in der Roten Liste gefährdeter Arten der IUCN 2017. Eingestellt von: P. J. Stephenson, Voahangy Soarimalala und Steven M. Goodman, 2014. Abgerufen am 02.03.2017.
- Hemicentetes nigriceps in der Roten Liste gefährdeter Arten der IUCN 2017. Eingestellt von: P. J. Stephenson, Voahangy Soarimalala und Steven M. Goodman, 2014. Abgerufen am 02.03.2017.