Eigentlicher Streifentenrek
Der Eigentliche Streifentenrek (Hemicentetes semispinosus), auch Gemeiner Streifentenrek, Streifentanrek oder Gelbstreifentanrek, ist eine Säugetierart aus der Gattung der Streifentenreks innerhalb der Familie der Tenreks (Tenrecidae). Er kommt endemisch in Madagaskar vor, wo er über weite Bereiche des östlichen Teils des Inselstaates verbreitet ist. Die bevorzugten Habitate umfassen tropische Regenwälder des Tief- und mittleren Berglands. Die Tiere sind aber auch in offenen Landschaften und in vom Menschen überprägten Gebieten anzutreffen. Namensgebend ist das schwarz-gelbe Streifenmuster des Rückens, zudem besitzt der Eigentliche Streifentenrek eine charakteristisch langgestreckte Schnauze. Das Fell besteht aus Borsten und Stacheln, die vor allem im Nackenbereich als Abwehrreaktion aufgestellt werden können. Am Hinterteil wächst eine Gruppe besonders kräftiger Stacheln, die ein Stridulationsorgan bilden, mit denen die Tiere rasselnde Geräusche zur Kommunikation erzeugen können.
| Eigentlicher Streifentenrek | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Eigentlicher Streifentenrek (Hemicentetes semispinosus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Hemicentetes semispinosus | ||||||||||||
| (G. Cuvier, 1798) |
Die Lebensweise des Eigentlichen Streifentenreks ist gut erforscht. Die Tiere sind tag- und nachtaktiv und weitgehend bodenlebend, wo sie unterirdische Baue graben. Das Sozialgefüge ist ausgesprochen entwickelt, während der Fortpflanzungsphase bilden sich große Familienverbände oder Kolonien, die komplexe Tunnelsysteme bewohnen. Zum Ende der Reproduktionszeit zerfallen diese Gruppenbildungen aber wieder. Die Nahrung besteht aus Wirbellosen, hauptsächlich Regenwürmer, von denen ein Individuum täglich nahezu die Menge seines Körpergewichtes verzehren kann. Wie bei vielen Tenreks ist die Körpertemperatur ebenfalls nicht stabil. Während der wenig ertragreichen Trockenzeit fallen die Tiere in einen Torpor. Weibchen bringen pro Wurf bis zu elf Jungtiere zur Welt. Die Individualentwicklung des Nachwuchses erfolgt sehr schnell, so dass die Jungen nach etwas mehr als einem Monat bereits geschlechtsreif sind.
Die wissenschaftliche Erstbeschreibung des Eigentlichen Streifentenreks datiert in das Jahr 1798. Dafür stand eine ältere Zeichnung eines gestreiften Tenreks Pate. Die gleiche Zeichnung diente in den nächsten fünf Jahren als Grundlage für die Etablierung von zwei weiteren wissenschaftlichen Artnamen, diese sind heute aber nicht mehr gültig. Aufgrund der weiten Verbreitung und relativen Anpassungsfähigkeit gilt der Bestand des Eigentlichen Streifentenreks als nicht gefährdet.
Merkmale
Habitus
Der Eigentliche Streifentenrek ist ein mittelgroßer Vertreter der Tenreks, er wird etwas größer als der nahe verwandte Schwarzkopftenrek (Hemicentetes nigriceps). Seine Kopf-Rumpf-Länge beträgt 13,0 bis 19,0 cm, der Schwanz ist nur als kurzer Stummel ausgebildet. Das Körpergewicht variiert von 90 bis 220 g mit einem Durchschnittswert von 130 g. Unterschiede zwischen männlichen und weiblichen Tieren liegen nicht vor. Im äußerlichen Erscheinungsbild ähneln die Tiere den Igeln, die Schnauze ist lang ausgezogen und vorn spitz zulaufend. Ein charakteristisches Merkmal stellt die Fellzeichnung dar. Diese besteht am Rücken aus drei hellen, gelblich gefärbten Streifen auf schwärzlich braunem Grund. Der Nacken zeigt ebenfalls einen gelblichen Farbton, der Kopf ist schwarz, diese Grundfärbung wird aber mittig auf der Oberseite von der Stirn bis zur Nase von einem weiteren gelblichen Streifen unterbrochen. Eine ähnliche Streifenbildung auf dem Kopf fehlt beim Schwarzkopftenrek. Das Haarkleid besteht aus Borsten und Stacheln, eine Unterwolle ist ebenfalls ausgebildet, kommt aber weitgehend nur bei Jungtieren vor und verliert sich mit zunehmendem Alter der Tiere. Dadurch wirkt der Eigentliche Streifentenrek deutlich stacheliger als der Schwarzkopftenrek. Die Stacheln in den hellen Streifen sind markant größer als die in den dunklen Farbabschnitten. Die längsten kommen am Nacken vor, wo sie zwischen 22 und 30 mm messen und austauschbar sind. Echtes Haar besteht nur auf der Unterseite und ist dort kastanienbraun getönt, zusätzlich treten noch zwei dunkle Streifen auf. Eine Besonderheit besteht am Hinterteil. Hier sind 14 bis 18, durchschnittlich meist aber 14 besonders breite Stacheln ausgebildet, die in drei Reihen auf einer Fläche von circa 1 cm² stehen.[1][2] Sie regenerieren nicht bei Verlust und können bei Anspannung der subdermalen Muskulatur gegeneinander gerieben werden, wodurch sie ein rasselndes Geräusch erzeugen. Die Stacheln wirken als Stridulationsorgan, es kommt auch beim Schwarzkopftenrek vor, besteht hier aber aus im Mittel weniger Stacheln.[3][4][5][6][7]
Schädel- und Gebissmerkmale
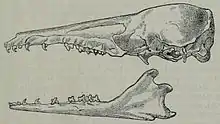
Der Schädel ist äußerst grazil gebaut, ein gemessenes Exemplar besitzt eine Länge von 41,6 mm.[8] Das Rostrum ist lang ausgezogen und schmal, wobei die Verschmälerung etwa auf Höhe des vorletzten und letzten Prämolaren beginnt und damit im Bezug auf die Zahnstellung weiter hinten als beim Schwarzkopftenrek. Das Nasen- und Gaumenbein sind langgestreckt, der Mittelkieferknochen ist klein. Der Scheitelkamm hat nur eine schwache Ausprägung, ebenso ragt der Wulst am Hinterhauptsbein weniger prominent heraus, der gesamte hintere Schädel wirkt dadurch noch steiler als beim Schwarzkopftenrek. Der Jochbogen ist wie bei allen Tenreks nicht geschlossen. Der vordere Bogenansatz liegt im Gegensatz zum Schwarzkopftenrek enger am Schädel an und ist länger. Der Unterkiefer zeigt einen leichteren Bau als beim Schwarzkopftenrek, im Unterschied zu diesen besitzt der Kronenfortsatz eine hakenförmige Gestalt,[9] sein vorderer Rand führt nicht ganz so steil aufwärts. Das Gebiss setzt sich aus 40 Zähnen zusammen, die folgende Zahnformel bilden: . Die gesamte untere Zahnreihe ist im Vergleich zum Schwarzkopftenrek weiter nach vorn versetzt, so dass mehr Platz zwischen dem aufsteigenden Gelenkast des Unterkiefers und dem letzten Molaren entsteht. Dadurch verschiebt sich auch die Position des Foramen mentale, das beim Eigentlichen Streifentenrek unter dem ersten Molaren, beim Schwarzkopftenrek aber unter dem letzten Prämolaren liegt. Insgesamt sind die Zähne in ihrer Größe stark reduziert und noch einmal 30 % kleiner als beim Schwarzkopftenrek. Zwischen den vorderen Zähnen und den Prämolaren liegen breite Diastemata, ihre Ausdehnung nimmt nach hinten deutlich zu. Sowohl die geringe Zahngröße als auch die weiten Zahnlücken gelten als Anpassung an die spezifische Ernährung des Eigentlichen Streifentenreks. Im oberen Gebiss sind die beiden vorderen Schneidezähne etwa gleich groß und weisen hakenförmig nach hinten. Der dritte Schneidezahn ist eher klein, der Eckzahn entspricht in Größe und Aussehen weitgehend den vorderen Incisiven. Die Schneidezähne des Unterkiefers besitzen alle etwa die gleiche Größe und haben eine schaufelartige Form. Der Eckzahn hier überragt die Schneidezähne und gleicht seinem oberen Pendant. Die Zahnkronen der vorderen Zähne sind zumeist mit zusätzlichen, manchmal aber schwach entwickelten Höckerchen ausgestattet. Entsprechend den anderen Tenreks haben die Mahlzähne ein zalambdodontes Kauflächenmuster bestehend aus drei Haupthöckern.[10][11][12]

Verbreitung
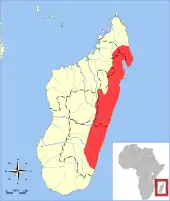
Der Eigentliche Streifentenrek kommt endemisch in Madagaskar vor. Sein Verbreitungsgebiet befindet sich im Ostteil des Inselstaates, wo es sich in einem mehr oder weniger breiten Streifen von Nord nach Süd erstreckt. Die Tiere bewohnen Tiefländer und mittlere Gebirgslagen. In der Regel reicht die Höhenverbreitung bis etwa 1550 m über dem Meeresspiegel. Die bevorzugten Habitate bestehen aus primären und sekundären tropischen Regenwäldern, darüber hinaus ist der Eigentliche Streifentenrek auch auf landwirtschaftlichen Nutzflächen, in Gärten oder auf von Menschen geschaffenen Wiesen- oder Offenlandflächen (“Pseudosteppe”) anzutreffen, einzelne Tiere dringen auch in Siedlungen vor. Aufgrund dessen gilt die Art als relativ häufig. Bedeutende Fundregionen im Norden finden sich in den Bergmassiven von Marojejy[13] und Anjanaharibe,[14][15][16] im südlich daran anschließenden Waldgebiet von Makira[17] sowie auf der östlich davon gelegenen Halbinsel Masoala,[18][19] alle zur Provinz Antsiranana gehörig. Abseits davon wurde auch eine Population am Tsaratanana-Massiv in der Provinz Mahajanga nachgewiesen, wobei die höchsten Fundpunkte hier bei 2050 m liegen.[20] In den zentralen Landesteilen sind die Tiere aus dem Waldgebiet von Ambatovy-Analamay-Tsorotorofotsy[21] in der Provinz Toamasina und vom Waldkorridor von Anjozorobe-Angavo[22] im Grenzgebiet der Provinzen Toamasina und Antananarivo belegt, zusätzlich auch vom Waldgebiet von Mahatsìnjo südöstlich von Tsinjoarivo[23][24] und südlich der Hauptstadt Antananarivo in der gleichnamigen Provinz. Weiter südlich gelegene Fundregionen bilden der Nationalpark Ranomafana, das Andringitra-Gebirge und die Bergländer um Ivohibe in der Provinz Fianarantsoa.[25] An den letzten beiden genannten Stellen tritt der Eigentliche Streifentenrek zusammen mit dem Schwarzkopftenrek auf, der aber generell höhere Gebirgslagen bis oberhalb der Baumgrenze bevorzugt. Lediglich bei Tsinjoarivo in 1550 m Höhe konnten beide Arten bisher in direkter Sympatrie nachgewiesen werden. Der Eigentliche Streifentenrek bewohnt hier Waldränder und Talgründe in Agrarflächen. Im nur 6 km entfernten Waldgebiet von Ankilahila, dessen Höhenniveau rund 150 m niedriger liegt, ist dagegen bisher nur der Eigentliche Streifentenrek belegt.[23][24][26][7]
Lebensweise
Territorialverhalten
.jpg.webp)
Die Lebensweise des Eigentlichen Streifentenreks ist relativ gut erforscht, dies betrifft sowohl Freilandstudien als auch Beobachtungen in Gefangenschaft. Im Gegensatz zum Schwarzkopftenrek ist der Eigentliche Streifentenrek weniger strikt nachtaktiv, sondern kann auch tagsüber in Erscheinung treten. Die Aktivitäten beginnen meist gegen 17.00 Uhr und erreichen zwischen 18.00 und 20.00 Uhr ihren Höhepunkt, danach nehmen sie kontinuierlich ab, bilden aber im Verlauf der Nacht verschiedene kleinere Maxima. Sporadische Aktivitäten herrschen von 09.00 bis 13.00 Uhr vor, während diese dann gegen 14.00 Uhr zum Erliegen kommen.[27][4] Am Boden bewegen sich die Tiere im Kreuzgang vorwärts und erreichen im Lauf eine Geschwindigkeit von 1,7 bis 2,9 km/h. Sie sind ebenfalls befähigt zu klettern und zu schwimmen,[27] allerdings können sie nicht springen. In unbekannten Gebieten bewegen sich die Tiere langsam vorwärts, pausieren häufig und schnüffeln beständig in der Luft und am Boden. Bei unübersichtlichem Gelände stoßen sie Klicklaute mit der Zunge aus, die eine Frequenz von 11 bis 16 kHz und einer Dauer von 0,2 bis 0,7 Millisekunden aus, die einer einfachen Echoortung dienen.[28] Zum Komfortverhalten gehören Kratzen mit den Hinterfüßen und das Bekauen der Nägel. Das typische „Gesichtwaschen“ anderer Tenreks mit beiden Vorderpfoten ist weniger stark ritualisiert und wird zumeist nur mit einer Pfote ausgeführt.[29][4][8][7]
Der Eigentliche Streifentenrek lebt während der Fortpflanzungszeit in Familiengruppen oder Kolonien, die mehrere Generationen einschließen, aber zum Ende der Phase wieder zerfallen. Die Art besitzt dadurch eines der komplexesten Sozialgefüge innerhalb der Gruppe der Afrotheria.[6] Große Gruppen können bis zu zwei Dutzend Individuen mit ausgewachsenen Tieren beiderlei Geschlechts sowie Jungtieren und Neugeborenen umfassen. Sie bewohnen dann komplexe, selbst gegrabene Tunnelsysteme, von denen eines in den 1960er Jahren im Gebiet von Ranomafana untersucht werden konnte. Es beherbergte eine Gruppe von 18 Tieren bestehend aus je zwei Männchen und zwei Weibchen sowie 14 Jungtieren. Das Tunnelsystem war in einen steilen, nach Süden orientierten Hang eingegraben und bestand aus drei Einheiten: einem etwa 300 cm langen Gang, bei dem eine Nestkammer in 225 cm Abstand zum Eingang bestand und auf der ein weiterer Tunnel folgte. In einer Entfernung von einem halben und rund 19 m gab es jeweils einen weiteren Gang mit einer Länge von 71 und 180 cm und einer Nestkammer am Ende. Die drei Einheiten besaßen keine direkte Verbindung zueinander, aber die einzelnen Individuen der Kolonie wechselten zwischen den Gängen hin und her. Kleinere Gruppen oder einzelne Tiere nutzen dagegen kleinere Baue, die häufig 30 bis 60 cm lang sind und 5 bis 7,5 cm unter der Erdoberfläche liegen. In der Regel sind sie in Hanglage eingegraben und folgen dem Hangverlauf oder winden sich um Baumwurzeln. Sie enthalten prinzipiell ein mit Pflanzen ausgelegtes Nest, das Material wird von den Tieren während des Grabens mit dem Maul transportiert und abgelegt. Zumeist haben die Tunnel nur einen Eingang, der ebenfalls mit Pflanzen abgedeckt ist. Die Eingänge werden häufig mit Kot markiert. Die Tiere graben dafür kleine Kuhlen und setzen den Kot ab. Ein ritualisiertes Bedecken durch Scharren mit den Hinterbeinen ist beim Eigentlichen Streifentenrek weniger stark ausgebildet als beim Schwarzkopftenrek. Die Tiere wechseln ihrer Baue in einer Periode von 10 bis 15 Tagen.[3][4][8][7]

Die Kommunikation innerhalb der Familiengruppe ist komplex. Dabei spielt möglicherweise die Stridulation mit den Stacheln des Hinterleibs eine bedeutende Rolle, bei der Töne im Frequenzbereich von 2 bis 200 kHz erzeugt werden. Die dabei produzierten Rasseltöne setzen sich je zur Hälfte aus kurzen, nur 7 bis 40 Millisekunden andauernden Tönen und aus längeren, bis zu 100 Millisekunden andauernden Sequenzen zusammen. Das Gehör des Eigentlichen Streifentenreks vermag aber am besten Töne von 10 bis 15 kHz wahrzunehmen, weswegen die effektive Entfernung der Wahrnehmung wahrscheinlich rund 4 m beträgt.[2] Daneben sind auch mehrere Lautäußerungen bekannt, die aus verschiedenen Grunz-, Quietsch- und Zwitscherlauten während der Sozialkontakte bestehen. Ein Knirschen wird bei Bedrohung ausgestoßen, während ein „putt-putt“-ähnlicher Laut ertönt, wenn ein Tier in seinem Nest gestört wird. Begegnungen zwischen Artgenossen laufen wenig aggressiv ab. Zumeist bestehen sie im gegenseitigen Beschnüffeln bestimmter Körperteile wie der Nase, dem Gesäß oder den Ohren. Bei antagonistischem Verhalten stellen die Tiere die Stacheln des Nackens auf, der Grad des Aufrichtens zeigt hierbei die Intensität der Erregung an. Etwaige Kämpfe werden mit den Vorderbeinen ausgetragen. Daneben versuchen die Tiere mit typischen auf- und abwärts gerichteten Bewegungen des Kopfes den Gegner mit den Stacheln zu treffen. Gelegentlich kommen auch Bisse zum Einsatz, ein geöffnetes Maul als Drohgebärde, häufig bei Tenreks belegt, ist vom Eigentlichen Streifentenrek nicht bekannt. Sowohl die Bisse als auch das geöffnete Maul sind wohl aufgrund der geringen Zahngrößen wenig effektiv.[3][4][8][7]
Ernährung und Energiehaushalt
Die Hauptnahrung des Eigentlichen Streifentenreks besteht aus Wirbellosen. Isotopenuntersuchungen an Tieren aus Tsinjoarivo ergaben Werte, wie sie typisch für andere grabende Tenreks etwa einigen Vertretern der Kleintenreks (Microgale) oder dem Eigentlichen Reiswühler (Oryzorictes hova) sind.[30] Die wichtigste Nahrungsressource stellen Regenwürmer dar, in Gefangenschaft verzehrten die Tiere auch Larven von Käfern und Fleisch. Beobachtungen an einem Individuum in menschlicher Obhut zufolge fraß dieses innerhalb eines Tages insgesamt 99,4 g an Nahrung, die jeweils zur Hälfte aus Fleisch und aus Regenwürmern (130 Stück) bestand, was nahezu dem Körpergewicht des Tieres entsprach.[29] Dies ist vergleichbar mit wildlebenden Tieren, die laut Untersuchungen in der Umgebung von Perinet bei Andasibe in der Provinz Fianarantsoa in den 1960er Jahren zwischen 85,5 und 104,4 g an Regenwürmern konsumierten. Gleichzeitig vorgenommene Schätzungen der Menge an Regenwürmern im Beobachtungsgebiet ergab etwa 83 kg auf einer Fläche von rund 4050 m² (1 Acre), was ausreichend ist, um zehn Tiere für 83 Tage zu ernähren (ohne Berücksichtigung der Reproduktion der Regenwürmer).[4] Die Tiere suchen ihre Beute mit der Nase am Boden unter Blätterabfall und ergreifen sie mit dem Maul. Durch Rückwärtsbewegungen oder Schütteln des Kopfes beziehungsweise mit Rückwärtsschritten ziehen sie den Wurm aus dem Boden. Mit den Vorderfüßen reinigen sie den Regenwurm von der Erde, wodurch das Tier gleichzeitig die Größe der Beute feststellt. In der Regel wird der Wurm vollständig verschluckt, die kleinen Zähne sind eher wenig zum Kauen geeignet. Nur wenn ein Exemplar zu groß ist, zerreißt der Eigentliche Streifentenrek ihn durch Betrampeln mit den Füßen und den Krallen in mehrere Teile und wiederholt die Prozedur. In der Fortpflanzungsperiode fressen die Tiere häufig in ihren Familiengruppen, die einzelnen Individuen drehen sich bei der Nahrungsaufnahme oder schwingen ihren Körper seitlich, was andere möglicherweise davon abhält, die Beute zu erhaschen. Sofern zwei Tiere den gleichen Wurm von unterschiedlichen Enden fressen, beansprucht ihn zumeist das größere Individuum.[3][4] Bei gefangenen Tiere wurde beobachtet, dass diese ihre Nahrung teilweise erbrechen und erneut verschlucken. Der Sinn ist nicht ganz eindeutig, eventuell wird die Beute dadurch von übermäßigem Schleim befreit. Ebenso trinken Tiere in menschlicher Obhut und halten ihre lange Nase dabei leicht nach oben gebogen. Ob der Eigentliche Streifentenrek in freier Wildbahn auch trinkt oder seinen Wasserbedarf über die Regenwürmer deckt, ist unklar.[31][29][3][4][8][7]
Wie zahlreiche andere Tenreks auch weist der Eigentliche Streifentenrek eine variierende Körpertemperatur auf, die sich den Bedingungen der Umgebung anpasst. Im Tagesrhythmus fluktuiert die Körpertemperatur mit zwei Hochwerten um 30 °C in der Mittagszeit und um Mitternacht, wonach sie jeweils auf rund 26,8 bis 28,2 °C abfällt.[27][29] Der Zyklus stimmt in etwa mit dem Aktivitätsverhalten überein, ist aber nicht in jedem Fall synchron.[4] Die Aktivität der Tiere steigert sich bei Umgebungstemperaturen um 18 bis 19 °C, was wahrscheinlich die Regulierung der Körpertemperatur fördert. Bei tieferen Außentemperaturen tritt häufig ein Torpor ein, der vor allem für die Monate des Südwinters charakteristisch ist. Er verläuft individuell verschieden und ist abhängig von der Körperkondition und den Fettreserven eines Tieres und den allgemeinen äußeren Bedingungen. Dadurch ist er nicht so strikt und lang andauernd wie beim Schwarzkopftenrek, der in deutlich höheren Gebirgslagen lebt. Die Variation der Körpertemperatur ist auch über den Jahresverlauf auffallend, sie liegt im Südsommer (November bis April) durchschnittlich 2,5 °C über der Umgebungstemperatur, im Südwinter (Mai bis Oktober) beträgt die Differenz nur 1,7 °C. Die Stoffwechselrate ist sehr gering und erreicht etwa 49 % des Wertes, der für ein ähnlich großes Säugetier zu erwarten wäre. Sie verringert sich im Winter noch einmal deutlich, in Phasen körperlichen Stresses wie während der Fortpflanzung steigt sie markant an. Die Fluktuation der Körpertemperatur und des Metabolismus ist verbunden mit Schwankungen des Körpergewichtes, allerdings liegen die Maximal- und Minimalwerte leicht versetzt dazu. So werden im März mit durchschnittlich 147,5 g die höchsten Werte erreicht, im Dezember sind die Tiere mit 98,5 g am leichtesten.[32][4][8][7]
Fortpflanzung
Die Fortpflanzung des Eigentlichen Streifentenreks ist durch Feld- und Laboruntersuchungen in den 1960er bis 1980er Jahren gut untersucht. Sie findet hauptsächlich während des Südsommers oder der Regenzeit von November bis etwa Mai statt, je nach Bedingung vor Ort kann sie lokal um etwa einen Monat schwanken. Während der Fortpflanzungsphase finden sich die Tiere in ihren Familiengruppen zusammen. Männchen werben um Weibchen in einem bestimmten Ritual, dass vornehmlich aus dem Beschnüffeln verschiedener Körperregionen besteht, etwa angefangen vom Nacken über die Nase zum Hinterteil, zu den Ohren – hier teilweise mit Bissen verbunden – und zur Körperseite. Dem folgt anschließend das Aufsitzen des Männchens auf dem Weibchen mit dem Geschlechtsakt, der bis zu 20 Minuten andauern kann.[33][34][3][4][8][7]
Die Tragzeit beläuft sich auf 58 bis 63 Tage, sie liegt damit in der Variation anderer Tenreks, ist aber im Vergleich zu ähnlich großen Säugetieren ausgesprochen lang.[33][34][35] Es kommen in einem Wurf zwischen einem und elf Jungtiere zur Welt, die Durchschnittswerte variieren je nach Region zwischen 6,2 und 8,8 Jungtieren je Wurf. Die Neugeborenen wiegen etwa 8,3 g bis 12,9 g und weisen eine Gesamtlänge von 6 bis 7 cm auf.[4] (Anderen Angaben zufolge beträgt die Länge der Neugeborenen rund 2,5 cm.[31]) Als Nesthocker sind die Augen und Ohren geschlossen und der Körper unbehaart mit Ausnahme der Vibrissen, die bereits rund 6 mm messen. Allerdings zeigt der Rumpf auffällige Pigmentierungen mit drei hellen Streifen auf dunklerem Grund, die die spätere Behaarung widerspiegeln. Am Nacken verbreitert sich der mittlere helle Streifen und zeigt die Stelle an, wo später die kräftigen Nackenborsten wachsen. Ebenso dehnt er sich leicht am Gesäß aus, wo das Stridulationsorgan entsteht. Die Stacheln und Borsten des Mittelstreifens wachsen zuerst und werden schon am Tag der Geburt sichtbar. Schon unmittelbar nach der Geburt können die Jungen kriechen und geben niederfrequente Laute von sich. Die Jungen verbleiben zunächst in einem Nest aus Pflanzenmaterial, das sowohl vom Mutter- als auch vom Vatertier herbeigeschafft wird. Sofern sie das Nest verlassen, werden sie mit der Nasenspitze von beiden Elterntieren zurückgeschoben, manchmal trägt sie die Mutter auch im Maul. Die Individualentwicklung schreitet sehr schnell voran. Mit vier Tagen stehen Jungtiere schon stabil auf den Beinen und beginnen zu laufen. Einen Tag später sind alle Stacheln sichtbar und am Nacken rund 4 mm lang, sie können als Abwehrhaltung bereits aufgerichtet werden. Die Stacheln des Stridulationsorgan lassen sich schon gegeneinanderreiben, doch sind sie noch zu kurz, um Töne zu erzeugen. Die Augen und Ohren öffnen sich nach acht bis zehn Tagen, was deutlich früher ist als bei anderen Tenreks. Zu diesem Zeitpunkt verlassen die Jungen erstmals das Nest und halten sich bevorzugt in der Nähe des Eingangs auf oder folgen der Mutter. Allerdings lässt damit auch die Fürsorge des Muttertiers wie das Lecken und Säubern der Jungen mit der Zunge langsam nach. Ab dem 16. Lebenstag ist das Stridulationsorgan voll entwickelt und erzeugt Töne. Die erste feste Nahrung nehmen Jungtiere etwa mit rund 18 Tagen zu sich, nach spätestens 25 Tagen sind sie entwöhnt. Bei Weibchen setzt der erste Östrus bereits nach 35 bis 40 Tagen ein. Dann verlangsamt sich auch das Wachstum, die Tiere sind dann durchschnittlich 14 bis 15 cm lang. Die Lebenserwartung in freier Wildbahn ist unbekannt, in menschlicher Obhut überlebten Tiere bis zu zweieinhalb Jahre.[36][3][4][35][8][7]
Fressfeinde und Parasiten
Als wichtigste Fressfeinde sind einige Madagassische Raubtiere wie der Ringelschwanzmungo, die Fossa und die Fanaloka zu nennen. Letztere hat aber nach Untersuchungen von Kotresten im Nationalpark Ranomafana nur einen geringen Einfluss aus die Population.[37] Vor allem auf den Geruch des Ringelschwanzmungos reagiert der Eigentliche Streifentenrek sehr stark. Abwehrverhalten besteht zumeist in der Flucht, bei Bedrängung im Aufrichten der Nackenstacheln. Mit den typischen Auf- und Abwärtsbewegungen des Kopfes versucht es dann den Beutegreifer mit den Stacheln zu treffen, wobei es teilweise auch Drehungen vollführt, die an kleine Sprünge erinnern.[4] Äußere Parasiten bilden Flöhe, Milben aus der Familie der Gamasidae und Schildzecken. Als innere Parasiten wurden der Einzeller Eimeria[38] und Fadenwürmer aus der Gruppe der Ascaroidea nachgewiesen.[4][7]
Systematik
Innere Systematik der Tenreks nach Everson et al. 2016[39]
|
Der Eigentliche Streifentenrek ist eine Art aus der Gattung der Streifentenreks (Hemicentetes), in der außerdem noch der Schwarzkopftenrek (Hemicentetes nigriceps) steht. Die Streifentenreks wiederum bilden einen Teil der Familie der Tenreks (Tenrecidae). Innerhalb dieser werden sie zu den Igeltenreks (Tenrecinae) geordnet, einer der drei Hauptlinien der Tenreks. Die Igeltenreks zeichnen sich durch ein stacheliges Haarkleid aus, was ihnen ein igelartiges Erscheinungsbild verleiht, allerdings haben sie einen deutlich kürzeren Schwanz. Charakteristisch für die Streifentenreks ist ihr auffallendes Fellmuster sowie das Stridulationsorgan am Hinterleib. Laut molekulargenetischen Untersuchungen stellt der Große Tenrek (Tenrec) den nächsten Verwandten dar. Während sich die Tenrecinae bereits im Oberen Eozän vor rund 35 Millionen Jahren von den übrigen Entwicklungslinien der Tenreks abgespalten hatten, trennten sich die Gattungen Tenrec und Hemicentetes im Mittleren Miozän vor rund 16 Millionen Jahren. Die Gattung Hemicentetes splittete sich im Pliozän vor etwa 4,75 Millionen Jahren in die beiden heutigen Linien.[39]


Die wissenschaftliche Erstbeschreibung stammt von Georges Cuvier aus dem Jahr 1798, Cuvier benannte den Eigentlichen Streifentenrek darin mit Erinaceus semispinosus und verwies ihn damit zu den Igeln. Außerdem berief er sich auf Georges-Louis Leclerc de Buffon, der 1776 im dritten Zusatzband seines Werkes Histoire naturelle, générale et particulière einen gestreiften Tenrek abgebildet und als Jungtier (le jeune tanrec) angesehen hatte.[40][41] Späteren Angaben von Fredericus Anna Jentink zufolge basierte die detailgenaue Abbildung auf einem Individuum von circa 11 cm Länge,[42] als Terra typica wird allgemein Madagaskar angegeben.[43] Nur zwei Jahre nach Cuvier benannte George Shaw, ebenfalls unter Berufung auf Buffon, den Eigentlichen Streifentenrek mit Erinaceus madagascariensis. Zusätzlich zitierte Shaw noch den französischen Forschungsreisenden Pierre Sonnerat, der bei seinem Besuch auf Madagaskar während seiner Expeditionen von 1774 bis 1781 die Tiere offensichtlich gesehen hatte und sie in seinem 1782 veröffentlichten Reisebericht beschrieb.[44][45] Wiederum nur kurze Zeit darauf, im Jahr 1803, kreierte Étienne Geoffroy Saint-Hilaire die wissenschaftliche Bezeichnung Setiger variegatus, bei der er sich ebenfalls von Buffons Abbildung leiten ließ.[46] Sowohl Erinaceus madagascariensis als auch Setiger variegatus gelten heute als synonym zu Hemicentetes semispinosus,[43] der erstere Artname wurde aber vor allem im 19. Jahrhundert sehr häufig für den Eigentlichen Streifentenrek benutzt. Die Gattungsbezeichnung Hemicentetes führte St. George Mivart erst im Jahr 1871 ein, er erhob dabei auch Hemicentetes madagascariensis zur Nominatform.[10] Der Schwarzkopftentrek wurde 1875 von Albert Günther als zweiter eindeutiger Vertreter der Streifentenreks etabliert, er galt im Verlauf des 20. Jahrhunderts teilweise als Unterart des Eigentlichen Streifentenreks.[8] Jedoch hatten bereits George Edward Dobson in den 1880er Jahren und Percy M. Butler in den 1940er Jahren auffallende Skelett- und zahnanatomische Unterschiede zwischen den beiden Formen herausgearbeitet,[9][12] so dass diese Ansicht nicht allgemein anerkannt war.[47] Aufgrund dessen werden beide heute als eigenständige Arten geführt.[43]
Bedrohung und Schutz
Der Eigentliche Streifentenrek ist weit verbreitet und tritt relativ häufig auf, darüber hinaus toleriert er anthropogene Landschaftsveränderungen. Größere Bedrohungen für den Bestand sind nicht bekannt, lokal haben der Holzeinschlag und die Landwirtschaft einen gewissen Einfluss. Außerdem werden die Tiere lokal als Nahrungsressource gejagt. Die IUCN stuft die Art in die Kategorie „nicht gefährdet“ (least concern) ein. Sie kommt in mehreren Naturschutzgebieten vor, so im Nationalpark Marojejy, im Nationalpark Masoala, Nationalpark Mantadia Andasibe, im Nationalpark Ranomafana und im Nationalpark Andringitra.[26]
Einziger Halter in Deutschland ist der Tierpark Donnersberg in Rockenhausen, ansonsten wird die Art in Europa nur noch von einem niederländischen Zoo allerdings nicht öffentlich zugänglich betreut.[48]
Literatur
- Georges Cuvier: Tableau elementaire de l’histoire naturelle des animaux. Paris, 1798, S. 1–710 (S. 108) ()
- J. F. Eisenberg und Edwin Gould: The Tenrecs: A Study in Mammalian Behavior and Evolution. Smithsonian Institution Press, 1970, S. 1–138
- Kathryn M. Everson, Voahangy Soarimalala, Steven M. Goodman und Link E. Olson: Multiple loci and complete taxonomic sampling resolve the phylogeny and biogeographic history of tenrecs (Mammalia: Tenrecidae) and reveal higher speciation rates in Madagascar’s humid forests. Systematic Biology 65 (5), 2016, S. 890–909 doi: 10.1093/sysbio/syw034
- Nick Garbutt: Mammals of Madagascar. A complete guide. Yale University Press, 2007, S. 1–304 (S. 32–56)
- Paulina D. Jenkins: Tenrecidae (Tenrecs and Shrew tenrecs). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 134–172 (S. 164–165) ISBN 978-84-16728-08-4
- Christopher D. Marshall und John F. Eisenberg: Hemicentetes semispinosus. Mammalian Species 541, 1996, S. 1–4
- Peter J. Stephenson: Streaked tenrecs, Hemicentetes. Afrotherian Conservation 5, 2007, S. 1–3
Einzelnachweise
- Olga V. Zherebtsova: Morphofunctional interpretation of the quills stridulating in tenrecs (Lipotyphla, Tenrecidae). Russian Journal of Theriology 5 (1), 2006, S. 1–11
- Ernest Glen Wever und Paul N. Herman: Stridulation and Hearing in the Tenrec. Journal of Auditory Research 8, 1968, S. 39–42
- Edwin Gould und John F. Eisenberg: Notes on the biology of the Tenrecidae. Journal of Mammalogy 47 (4), 1966, S. 660–686
- J. F. Eisenberg und Edwin Gould: The Tenrecs: A Study in Mammalian Behavior and Evolution. Smithsonian Institution Press, 1970, S. 1–138
- Nick Garbutt: Mammals of Madagascar. A complete guide. Yale University Press, 2007, S. 1–304 (S. 32–56)
- Peter J. Stephenson: Streaked tenrecs, Hemicentetes. Afrotherian Conservation 5, 2007, S. 1–3
- Paulina D. Jenkins: Tenrecidae (Tenrecs and Shrew tenrecs). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 134–172 (S. 164–165) ISBN 978-84-16728-08-4
- Christopher D. Marshall und John F. Eisenberg: Hemicentetes semispinosus. Mammalian Species 541, 1996, S. 1–4
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part I. London, 1883, S. S. 70–71 und 72–77 ()
- St. George Mivart: On Hemicentetes, a new Genus of Insectivora, with some additional remarks on the osteology of that order. Proceedings of the Zoological Society of London, 1871, S. 58–79 ()
- Wilhelm Leche: Zur Entwicklungsgeschichte des Zahnsystems der Säugetiere, zugleich ein Beitrag zur Stammesgeschichte dieser Tiergruppe. Zweiter Teil: Phylogenie. Zweites Heft: Familien der Centetidae, Solenodontidae und Chrysochloridae. Zoologica 20, 1906/1908, S. 1–157 ()
- Percy M. Butler: A Comparison of the Skulls and Teeth of the Two Species of Hemicentetes. Journal of Mammalogy 22 (1), 1941, S. 65–81
- Steven M. Goodman und Paulina D. Jenkins: Tenrecs (Lipotyphla; Tenrecidae) of the Parc National de Marojejy, Madagascar. Fieldana Zoology 97, 2000, S. 201–229
- Steven M. Goodman und Paulina D. Jenkins: The Insectivores of the Réserve Spéciale d’Anjanaharibe-Sud, Madagascar. Fieldiana Zoology 90, 1998, S. 139–161
- Franco Andreone, Jasmin E. Randrianirina, Paula D. Jenkins und Gennaro Aprea: Species diversity of Amphibia, Reptilia and Lipotyphla (Mammalia) at Ambolokopatrika, a rainforest between the Anjanaharibe-Sud and Marojejy massifs, NE Madagascar. Biodiversity and Conservation 9, 2000, S. 1587–1622
- Voahangy Soarimalala und Steven M. Goodman: Diversité biologique des micromammifères non volants (Lipotyphla et Rodentia) dans le complexe Marojejy-Anjanaharibe-sud. In: Steven M. Goodman und Lucienne Wilmé (Hrsg.): Nouveaux résultats faisant référence à l’altitude dans la région des massifs montagneux de Marojejy et d’Anjanaharibe-sud. Recherche pour le développement, Série Sciences biologiques, Centre d’Information et de Documentation Scientifique et Technique 19, 2003, S. 231–276
- Zafimahery Rakotomalala, Vonjy Andrianjakarivelo, Volatiana Rasataharilala und Steven M. Goodman: Les petits mammifères non volant de la forêt de Makira, Madagascar. Bulletin de la Société zoologique de France 132, 2007, S. 205–221
- Vonjy Andrianjiakarivelo, Emilienne Razafimahatratra, Yvette Razafindrakoto und Steven M. Goodman: The terrestrial small mammals of the Parc National de Masoala, northeastern Madagascar. Acta Theriologica 50 (4), 2005, S. 537–549
- Voahangy Soarimalala: Exploitation des bois précieux à Masoala, Madagascar: quel impact sur les tenrecs? Afrotherian Conservation 14, 2018, S. 20–32
- Claudette Patricia Maminirina, Steven M. Goodman und Christopher J. Raxworthy: Les microammifères (Mammalia, Rodentia, Afrosoricida et Soricomorpha) du massif du Tsaratanana et biogéographie des forêts de montagne de Madagascar. Zoosystema 30 (3), 2008, S. 695–721
- Voahangy Soarimalala und Martin Raheriarisena: The non-volant and non-primate mammals of the Ambatovy-Analamay forest. In: Steven. M. Goodman und V. Mass (Hrsg.): Biodiversity, exploration, and conservation of the natural habitats associated with the Ambatovy project. Malagasy Nature 3, 2010, S. 153–177
- Voahangy Soarimalala, Landryh T. Ramanana, José M. Ralison und Steven M. Goodman: Les petits mammifères non-volants du „Couloir forestier d’Anjozorobe – Angavo“. In: Steven M. Goodman, Achille P. Raselimanana und Lucienne Wilmé (Hrsg.): Inventaires de la faune et de la flore du couloir forestier d’Anjozorobe – Angavo. Recherche pour le développement, Série Sciences biologiques, Centre d’Information et de Documentation Scientifique et Technique 24, 2007, S. 141–182
- Steven M. Goodman, Daniel Rakotondravony, Voahangy Soarimalala, J. B. Duchemin und J.-M. Duplantier: Syntopic occurrence of Hemicentetes semispinosus and H. nigriceps (Lipotyphla: Tenrecidae) on the central highlands of Madagascar. Mammalia 64, 2000, S. 113–116
- Steven M. Goodman, Daniel Rakotondravony, Marie Jeanne Raherilalao, Domoina Rakotomalala, Achille P. Raselimanana, Voahangy Soarimalala, Jean-Marc Duplantier, Jean-Bernard Duchemin und J. Rafanomezantsoa: Inventaire biologique de la Foret de Tsinjoarivo, Ambatolampy. Akon'ny Ala 27, 2000, S. 18–27
- Voahangy Soarimalala, Stephen M. Goodman, H. Ramiaranjanahary, L. L. Fenohery und W. Rakotonirina: Les micromammifères non-volants du Parc National de Ranomafana et du couloir forestier qui le relie au Parc National d’Andringitra. Dans Inventaire biologique du Parc National de Ranomafana et du couloir forestier qui la relie au Parc National d’Andringitra. In: Stephen M. Goodman und V. R. Razafindratsita (Hrsg.): Recherches pour le Développement. Série Sciences Biologiques 17, 2001, S. 199–229
- P. J. Stephenson, Voahangy Soarimalala und Steven M. Goodman: Hemicentetes semispinosus. The IUCN Red List of Threatened Species 2016. e.T40593A97189434 (); zuletzt abgerufen am 19. Februar 2017
- Konrad Herter: Über die Borstenigel von Madagaskar (Tenrecinae). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin N. F. 2, 1962, S. 5–37
- Edwin Gould: Evidence for echolocation in the Tenrecidae of Madagaskar. Proceedings of the American Philosophical Society 109 (6), 1965, S. 352–360
- Konrad Herter: Untersuchungen an lebenden Borstenigeln (Tenrecinae), 3: Über das Verhalten und die Lebensweise des Streifentanreks (Hemicentetes semispinosus G. Cuvier) in Gefangenschaft. Zoologische Beiträge 9 (2), 1963, S. 237–274
- Melanie Dammhahn, Voahangy Soarimalala und Steven M. Goodman: Trophic Niche Differentiation and Microhabitat Utilization in a Species-rich Montane Forest Small Mammal Community of Eastern Madagascar. Biotropica 45 (1), 2013, S. 111–118
- Heinrich Francke: Gefangenschaftsbeobachtungen an Hemicentetes semispinosus. Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin N. F. 1, 1961, S. 118–123
- Peter J. Stephenson und Paul A. Racey: Seasonal variation in resting metabolic rate and body temperature of streaked tenrecs, Hemicentetes nigriceps and H. semispinosus (Insectivora: Tenrecidae). Journal of Zoology 232, 1994, S. 285–29
- John F. Eisenberg: Tenrecs and solenodons in captivity. International Zoo Yearbook 15, 1975, S. 6–12
- Walter Poduschka: Hyperthelie, Wurfgröße und Trächtigkeitsdauer bei der Unterfamilie Tenrecinae Cabrera, 1925 (Mammalia: Insectivora: Tenrecidae), nebst Bemerkungen zur Längsstreifung der Gattung Hemicentetes. Contributions to Zoology 66 (2), 1996, S. 119–128
- Peter J. Stephenson, Paul A. Racey und Félix Rakotondraparany: Maintenance and reproduction of tenrecs (Tenrecidae) at Parc Tsimbazaza, Madagascar. International Zoo Yearbook 33, 1994, S. 194–201
- Richard Weigl: Longevity of mammals in captivity; from the Living Collections of the world A list of mammalian longevity in captivity. Kleine Senckenberg-Reihe, Band 48, 2005, S. 1–214 (S. 34)
- Steven M. Goodman, F. J. Kerridge und R. C. Ralisomalala: A note on the diet of Fossa fossana (Carnivora) in the central eastern humid forests of Madagascar. Mammalia 67 (4), 2003, S. 595–598
- Lee Couch, Juha Laakkonen, Steven Goodman und Donald W. Duszynski: Two New Eimerians (Apicomplexa) from Insectivorous Mammals in Madagascar. Journal of Parasitology 97 (2), 2011, S. 293–296
- Kathryn M. Everson, Voahangy Soarimalala, Steven M. Goodman und Link E. Olson: Multiple loci and complete taxonomic sampling resolve the phylogeny and biogeographic history of tenrecs (Mammalia: Tenrecidae) and reveal higher speciation rates in Madagascar’s humid forests. Systematic Biology 65 (5), 2016, S. 890–909 doi: 10.1093/sysbio/syw034
- Georges-Louis Leclerc de Buffon: Histoire naturelle, générale et particulière., Supplément, Tome troisième. Paris, 1776, Tafel 37 ()
- Georges Cuvier: Tableau elementaire de l’histoire naturelle des animaux. Paris, 1798, S. 1–710 (S. 108)
- Fredericus Anna Jentink: On the hedgehogs from Madagascar. Notes from the Royal Zoological Museum at Leyden 1879, S. 137–151 ()
- Don E. Wilson und DeeAnn M. Reeder: Mammal Species of the World. A taxonomic and geographic Reference. Johns Hopkins University Press, 2005 ()
- Pierre Sonnerat: Voyage aux Indes Orientales et à la Chine. hier Tome second. Paris, 1782, S. 146–147 und Tafel 93 ()
- George Shaw: General zoology. Vol. 1 London, 1800, S. 1–552 (S. 548) ()
- Étienne Geoffroy Saint-Hilaire: Catalogue des mammifères du Muséum National d'Histoire Naturelle. Paris, 1803, S. 1–272 (S. 72) ()
- Henri Heim de Balsac: Insectivores. In: R. Battistini und G. Richard-Vindard (Hrsg.): Biogeography and ecology in Madagascar. Den Haag, 1972, S. 629–660
- Zootierliste (), zuletzt abgerufen am 10. März 2017
Weblinks
- Hemicentetes semispinosus in der Roten Liste gefährdeter Arten der IUCN 2017. Eingestellt von: P. J. Stephenson, Voahangy Soarimalala und Steven M. Goodman, 2014. Abgerufen am 19.2.2017.