Großer Tenrek
Der Große Tenrek (Tenrec ecaudatus), auch Großer Tanrek oder einfach nur Tenrek beziehungsweise Tanrek, ist eine Säugetierart aus der Gruppe der Igeltenreks, die zur Familie der Tenreks (Tenrecidae) gehört. Er ist der größte, weitestverbreitete und wohl bekannteste Vertreter seiner Familie. Äußerlich zeichnet er sich durch einen kräftigen Körperbau und einen großen breiten Kopf mit langer Schnauze aus. Das Fell ist borstig, am Nacken und am Kopf sind Stacheln ausgebildet. Charakteristische Merkmale stellen auch der Schwanzstummel und die langen Eckzähne dar. Das natürliche Ursprungsgebiet der Tiere ist Madagaskar. Dort kommen sie in nahezu allen Lebensräumen vor: sie bewohnen sowohl die feuchten tropischen Regenwälder im östlichen Teil als auch die laubabwerfenden trockenen Wälder des westlichen sowie die sehr trockenen Dornenbuschgebiete des südwestlichen Teils der Insel. Sie können sich an von Menschen veränderte Landschaften anpassen – so treten sie auch in städtischen Gebieten auf. Es gibt Populationen auf Mauritius, Réunion, den Seychellen und den Komoren.
| Großer Tenrek | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Großer Tenrek (Tenrec ecaudatus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Tenrec | ||||||||||||
| Lacépède, 1799 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Tenrec ecaudatus | ||||||||||||
| (Schreber, 1777) |
Die Lebensweise des Großen Tenrek ist relativ gut erforscht. Er ist nachtaktiv und bodenbewohnend. Als Unterschlupf nutzt er selbst gegrabene Tunnel, die mit einem Nest aus Pflanzen ausgestattet sind. Dort verbringt er auch den trockenen und nahrungsarmen Südwinter in einem Starrezustand (Torpor), der mehrere Monate anhalten kann. Der allgemeine Lebensrhythmus wird dabei von stark schwankenden Körpertemperaturen begleitet, die von den Außentemperaturen und der Aktivität abhängen. Die Tiere sind einzelgängerisch und kommen nur während der Fortpflanzungsperiode zusammen. Ein Wurf besteht aus durchschnittlich 15 Jungen; maximal wurden bisher 32 beobachtet, was die höchste Anzahl an Jungtieren unter den Säugetieren ist. Die Jungtiere haben ein gestreiftes Fell, das zudem deutlich mehr Stacheln aufweist. Am Hinterteil sind die Stacheln zu einem Stridulationsorgan umgestaltet, mit dem sie rasselnde Töne erzeugen können. Die Aufzucht der Jungen dauert etwa zwei Monate. Die Ernährung des Großen Tenreks basiert hauptsächlich auf wirbellosen Tieren wie Insekten und Würmern, zudem verzehrt er auch kleine Wirbeltiere und pflanzliches Material wie Früchte.
Der Große Tenrek wurde bereits in der Mitte des 18. Jahrhunderts in Europa erwähnt, seine wissenschaftliche Erstbeschreibung als Art datiert in das Jahr 1777. Obwohl die Bezeichnung der Gattung mit Tenrec etwa zwei Jahrzehnte später erfolgte, war der Große Tenrek im Verlauf des 19. Jahrhunderts unter der jüngeren Bezeichnung Centetes bekannt. Erst Mitte des 20. Jahrhunderts setzte sich der heute gültige Name durch. Der Bestand des Großen Tenreks gilt als ungefährdet, lokal kann es aber durch intensive Jagd zu Rückgängen kommen.
Merkmale
Habitus
Der Große Tenrek ist der größte Vertreter der Tenreks. Ausgewachsene Tiere haben eine Kopf-Rumpf-Länge von 26,5 bis 39 cm und einen 1,0 bis 1,5 cm langen Schwanz. Das Körpergewicht variiert von 600 bis 2400 g je nach Jahreszeit,[1][2] Tiere in Gefangenschaft können bis zu 3000 g wiegen.[3] Männchen sind in der Regel etwas schwerer als Weibchen. Der Körper ist ausgesprochen muskulös gebaut. Am Rücken besteht kein echtes Stachelkleid, sondern lediglich ein borstiges Fell aus 27 bis 60 mm langen Haaren, das mit kürzeren, 8 bis 15 mm langen Stacheln durchsetzt ist. Am Nacken und auf dem Kopf sind die Stacheln, die kurz und dick (10 bis 15 mm) oder lang und schlank (18 bis 24 mm) sein können, dichter ausgebildet.[4] Die Stacheldichte geht meist mit dem Lebensalter der Tiere zurück. Aus dem Fell ragen zusätzlich lange, schwarze Haare als Tastorgane. Die Färbung des Rückenfells variiert je nach geographischer Region und reicht von rötlich- über gelblich- bis agouti braun. Weibchen sind meist etwas dunkler gefärbt als Männchen. Die Unterseite zeigt sich spärlich behaart und cremefarben. Der Kopf ist hell bräunlich und relativ groß, er nimmt fast ein Drittel der Gesamtlänge eines Tieres ein.[5] Der Große Tenrek hat außerdem eine sehr breite Gestalt. Es zeigt sich ein deutlicher Geschlechtsdimorphismus, da bei Männchen die Augen deutlich weiter auseinander stehen. Das breite Gesicht wird bei den Männchen durch den mächtigen Musculus masseter bewirkt. Zusätzlich ist die Schnauze langgestreckt und mit markanten Vibrissen ausgestattet. Die Vorderbeine sind länger als die Hinterbeine, Hände und Füße weisen jeweils fünf Strahlen auf und verfügen über kurze, kräftige Krallen.[6][1][3][2] Die Weibchen besitzen 12 bis 14 Zitzenpaare.[7][8]
Jungtiere unterscheiden sich deutlich von ausgewachsenen Individuen. Sie haben bis zu einer Körperlänge von 16 cm eine dunkelbraune Rückenfärbung, die von fünf längsgerichteten Streifen aus weißlichen Stacheln unterbrochen wird. Auf der Mittellinie befindet sich ein Stridulationsorgan wie es vergleichbar bei den Streifentenreks vorkommt. Es erstreckt sich auf einer Länge von 40 mm und einer Breite von 2 mm und besteht aus 30 bis 80 Stacheln. Diese stehen in drei bis fünf Reihen und haben eine cremefarbene Tönung. Sie sind hier in eine spezielle Unterhautmuskulatur eingebettet und können durch Anspannung der Muskeln aneinander reiben, wodurch Töne erzeugt werden. Im Übergang zum ausgewachsenen Tier gehen das Streifenmuster und das Stridulationsorgan verloren und werden durch einfachere Haare und Borsten ersetzt.[9][3][4][8]
Schädel- und Gebissmerkmale
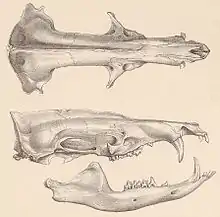
Der Schädel ist langgestreckt sowie zylindrisch und mit einem ausgezogenen Rostrum versehen. Der Hirnschädel zeigt dagegen einen kurzen, schmalen und winkligen Bau; hier sind außerdem kräftige Knochenwülste in der Sagittal- und Lambdaregion vorhanden. Das Verhältnis von vorderem zu hinterem Schädelabschnitt beträgt 0,64:0,36.[10] Die Zwischenaugenregion zeigt sich relativ schmal und gestreckt. Der Jochbogen ist nicht geschlossen, ein typisches Kennzeichen aller Tenreks. Das Gebiss besteht aus insgesamt 38 Zähnen mit folgender Zahnformel: . In der oberen Zahnreihe bestehen beidseits des Eckzahns und hinter dem ersten Prämolaren kurze Diastemata, in der unteren Zahnreihe fehlt die Zahnlücke vor dem Eckzahn. Die oberen beiden ersten Schneidezähne ähneln sich in ihrer Größe, der dritte ist nur im Milchgebiss vorhanden, deutlich kleiner als die vorderen und im Dauergebiss nicht mehr präsent. Im Unterkiefer sind alle Schneidezähne eher klein, nur der mittlere ist geringfügig höher. Die Invicisiven werden durch zusätzliche kleine Höckerchen an den Zahnkronen charakterisiert. Diese fehlen beim Eckzahn, der dadurch wie ein echter Eckzahn wirkt (caniniform). Außerdem ist er sowohl im oberen als auch im unteren Gebiss ausgesprochen groß, er kann bei Männchen bis zu 15 mm lang werden. Im Zwischenkieferknochen sind Einstülpungen ausgebildet, die den unteren Eckzahn bei Gebissschluss aufnehmen. Die vorderen beiden Prämolaren weisen eine kegelförmige Gestalt auf, der letzte ähnelt deutlich den Molaren. Die Mahlzähne kennzeichnet ein zalambdodontes Kauflächenmuster, bestehend aus drei Haupthöckern (Para-, Proto- und Metaconus; bezogen auf die Oberkiefermolare) in dreieckiger Anordnung.[5][11][10]

Skelettmerkmale

Die Wirbelsäule setzt sich aus 7 Hals-, 17 bis 19 Brust-, 4 bis 7 Lenden-, 2 bis 3 Kreuzbein- und 8 bis 10 Schwanzwirbeln zusammen.[10] Die weitgehend generalisierte, bodenbewohnende Lebensweise des Großen Tenrek lässt sich auch am Skelettbau erkennen. Das Schulterblatt hat eine kurze und hohe Form und weicht so von der langschmalen grabender Tenreks ab. Der Oberarmknochen ist beim Großen Tenrek, verglichen mit anderen Tenreks, deutlich länger als die Speiche. Er besitzt einen massiv ausgebildeten Großen Rollhügel, was für kraftvolle Längsbewegungen des Vorderbeins spricht. Dagegen ist der Kleine Rollhügel eher verkleinert, als Ansatzstelle des Musculus subscapularis verweist dies auf geringere Rotationsfähigkeiten des Arms. Ebenso zeigt die deltopectorale Leiste am Humerusschaft nur eine geringe Entwicklung. Das untere Gelenkende (Ellenbogengelenk) ist vergleichsweise breit. Da hier ein Teil der Unterarmmuskulatur entspringt, kann auf eine gewisse Kratz- und Grabbefähigung geschlossen werden. Dies unterstützt auch der verhältnismäßig lange obere Gelenkfortsatz (Olecranon) der Elle, der etwa 23 % der Länge des Gesamtknochens ausmacht. Das Gelenkende am unteren Schaftende zur Artikulation mit der Hand ist eher breit und flach gestaltet. An den Handwurzelknochen treten keine Verwachsungen auf, wie sie bei einigen grabenden Arten nachgewiesen sind. Sie haben zudem eine eher lange Form, während die Fingerglieder kurz erscheinen. Der Prepollex, eine fingerartige Verlängerung am Großen Vieleckbein zur Innenseite des Daumens hin, ist sehr lang. Die Krallen wiederum wirken sehr lang und flach, sie sind nicht so stark gebogen wie bei baumkletternden Tenreks.[12][13] Am Hinterfuß deuten die etwas aufgewölbteren Knöchel stärkere Einschränkungen in der Bewegungsfreiheit an, außerdem spricht der kräftige Nacken des Sprungbeins für eine terrestrische Fortbewegung. Dem gegenüber verweist der gerundete Kopf des Sprungbeins auf eine gewisse Befähigung zum Klettern.[14]
Verbreitung und Lebensraum
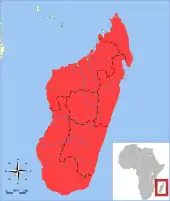
Der Große Tenrek lebt endemisch auf ganz Madagaskar. Er kommt in nahezu allen Waldlandschaften der Insel vor, von den feuchten tropischen Regenwäldern im Osten über die laubabwerfenden trockenen Wälder des Westens bis hin zu den sehr trockenen Dornenbuschgebieten des Südwestens. In den Wäldern des westlichen Madagaskars konnte die Art an nahezu allen untersuchten Plätzen nachgewiesen werden (35 von 37 Lokalitäten).[15] Zusätzlich ist er auch auf Plantagen, auf Farmland, in sekundär geöffneten Waldlandschaften und in Stadtgebieten anzutreffen. Die Tiere bevorzugen Habitate mit Wasser in der Nähe und dichtem Bodenbewuchs, der als Deckung dient. Die Höhenverbreitung reicht vom Meeresspiegelniveau bis auf etwa 2050 m Höhe. Die Art ist ebenfalls auf den Komoren häufig, ihr Ursprung dort gilt als ungeklärt.[16] Auf den Maskarenen (Mauritius und Réunion) wurde sie als Nahrungsressource eingeführt, ebenso auf den Seychellen, wo sie seit etwa 1880 nachgewiesen ist. Auf den ersten beiden Inselgruppen werden die Tiere noch gejagt, nicht jedoch auf letzterer. Dort kann die Populationsdichte in zuträglichen Habitaten bis zu 15 Individuen je Hektar betragen.[17][1][8]
Lebensweise
Territorialverhalten

Die Lebensweise des Großen Tenreks ist relativ gut erforscht, sie wurde an Freilandtieren auf Madagaskar und den Seychellen studiert. Die Tiere sind hauptsächlich nachtaktiv mit einer bimodalen Verteilung: die erste Aktivitätsphase findet am frühen Abend zwischen 19:30 und 21:00 Uhr statt mit einem Höhepunkt um 20.00 Uhr. Die zweite erstreckt sich von 00:30 bis 05:00 Uhr mit der Hauptaktivität von 01:00 bis 02:00 Uhr. Über das Jahr gesehen haben die Tiere ebenfalls eine zyklische Aktivität. Im Südwinter (Mai bis Oktober), wenn die Nahrungsressourcen zurückgehen, verfallen sie in einen Torpor, der mitunter bis zu sechs Monate anhalten kann. Der Große Tenrek ist bodenbewohnend und bewegt sich dabei im Kreuzgang vorwärts. Er kann zudem gut schwimmen und durchquert so Reisfelder. Auch beim Klettern in Felsen wurde er bereits beobachtet, auf Bäumen sieht man ihn hingegen fast nie. In unbekanntem Gelände bewegt sich der Große Tenrek langsam vorwärts, pausiert häufig und schnüffelt beständig in der Luft, manchmal hebt er dabei ein Vorderbein, bleibt aber ansonsten reglos stehen. Teilweise stößt er auch pfeifartige Laute aus, die wohl der Kommunikation mit Artgenossen dienen. Durch Zungenklicks können die Tiere hochfrequente Töne im Bereich von 10 bis 16 kHz und mit einer Dauer von 0,1 bis 3,6 ms erzeugen. Sie gehören wahrscheinlich zu einer einfachen Echoortung, mit der sich die Tiere ebenfalls orientieren. Im Gegensatz zu anderen Tenreks ist beim Großen Tenrek der Sehsinn besser entwickelt und wird auch aktiv eingesetzt.[9][3][8]
Der Große Tenrek nutzt individuelle Aktionsräume von 1 bis 3 ha Größe, die Ausmaße sind abhängig vom Nahrungsangebot. Sie überlappen sich im Grenzgebiet mit denen anderer Individuen. Die Ruhezeit verbringen die Tiere in natürlichen oder selbst gegrabenen unterirdischen Bauen. Die Baue variieren je nach Jahreszeit, Alter und Geschlecht. Einer von zwei untersuchten Bauen bei Ivohibe im südöstlichen Madagaskar, die während des Südwinters in Benutzung waren, hatte eine Länge von rund 2 m, wobei die ersten anderthalb Meter flach unter dem Erdboden verliefen und der Tunnel dann nach unten abknickte und 30 cm unter der Erde endete. Der Eingang war mit Blättern abgedeckt. Weibchen mit Nachwuchs legen kompliziertere und tiefere Tunnel an, die zwei Eingänge haben, so dass der Bau eine Y-Form erhält. Männchen nutzen dagegen wohl flachere Baue. Die Tunnel befinden sich häufig an Flusshängen unter Baumwurzeln oder umgefallenen Bäumen. Sie enden blind in einer Nestkammer aus Pflanzenmaterial. Einige vermessene Nestkammern waren 8 bis 13 cm lang, 13 bis 18 cm weit und 7 bis 11,5 cm hoch.[18][6][19] Die Pflanzenteile werden von den Tieren im Maul heran transportiert. In den Tunneln herrscht ein weitgehend ausgeglichenes Klima. Die Innentemperaturen in Tiefen von 6 bis 8 cm betragen 19,7 bis 25,5 °C bei Außentemperaturen von 18 bis 29 °C. Zum Schlaf rollen sich die Tiere auf den Hinterbeinen hockend ein oder liegen seltener seitlich, bei hohen Temperaturen nehmen sie eine gestreckte Position ein. Das Komfortverhalten besteht aus Kratzen mit dem Hinterfuß und Lecken. Das für andere Igeltenreks und auch die Kleintenreks (Microgale) teils typische „Gesichtwaschen“ mit beiden Vorderbeinen kommt beim Großen Tenrek nicht vor.[6][3][2][8]

Allgemein lebt der Große Tenrek einzelgängerisch, Ausnahmen stellen Mutter-Jungtiergruppen dar, während der Paarung bleiben Männchen und Weibchen teilweise bis zu drei Tage zusammen. Gleichgeschlechtliche Artgenossen vermeiden Begegnungen untereinander. Finden diese doch statt, inspizieren sich die Tiere eine kurze Zeit gegenseitig, teilweise begleitet von schnaufenden Tönen, und trennen sich dann wieder. Bei aggressivem Verhalten werden die Stacheln am Hals aufgestellt und es kommt zu Beißattacken. Als Drohgebärde dienen ein aufgerissenes Maul, dessen Spanne bis zu 13 cm betragen kann, und ein Fußstampfen. Die weitere Lautkommunikation umfasst Grunz- und Zischlaute, die häufig in Bedrängnis ausgestoßen werden. Außerdem erfolgt die Verständigung über chemische Reize. Der Große Tenrek defäziert häufig neben den Eingängen seiner Baue. Mitunter hebt er mit den Vorderbeinen kleine Löcher aus und vergräbt seinen Kot darin mit kickenden Bewegungen der Hinterbeine. Muttertiere markieren beim Verlassen ihre Baue, teilweise unter Beteiligung des Nachwuchses. Bei Begegnungen mit Artgenossen oder in fremdem Terrain reiben die Tiere ihr Hinterteil oder ihre Seiten an Objekte.[20][3][2][8]
Ernährung und Energiehaushalt
Der Große Tenrek ist ein Allesfresser, der hauptsächlich wirbellose Tiere wie Insekten und Würmer zu sich nimmt. Untersuchungen von 22 Mageninhalten aus dem Waldgebiet von Kirindy im westlichen Madagaskar ergaben einen großen Anteil an Ameisen, darunter die Gattungen Pachycondyla, Camponotus, Monomorium und Pheidole, und Termiten, etwa Coarctotermes. Ebenfalls häufig waren Hundertfüßer und verschiedene Entwicklungsstadien von Käfern, hier dominierten Blatthornkäfer und Pflanzenkäfer.[21] Weitere fünf Mageninhalte, die bereits in den 1930er Jahren analysiert wurden, enthielten vor allem Raupen und Käfer.[18] Darüber hinaus umfasst das Nahrungsspektrum auch Regenwürmer und Heuschrecken. Ebenso verzehrt der Große Tenrek kleinere Wirbeltiere, beispielsweise Frösche, Eidechsen oder Mäuse, zum Teil auch Vogeleier, Tiere in Gefangenschaft fraßen des Weiteren rohes Fleisch. Pflanzen spielen ebenfalls eine gewisse Rolle, sie werden in Form von Früchten überwiegend gegen Ende des Südsommers aufgenommen, wenn die Tiere Fett ansetzen.[1][6][3][2][8]
Insgesamt besteht die Hauptnahrung aus kleinen Beutetieren mit einer Spezialisierung auf Gliederfüßer. Größere Beute wie die Wirbeltiere wird wahrscheinlich eher zufällig bei Gelegenheit erlegt. Bei der Nahrungssuche schreitet der Große Tenrek langsam voran und sucht am Boden unter Blätterabfall und an abgestorbenen Bäumen. Dabei scharrt er häufig kleine Löcher von 5 cm Tiefe; seine wichtigsten Beutetiere leben bis maximal 10 cm unter der Erdoberfläche. Die Schnauze spielt beim Aufspüren der Beute eine wichtige Rolle. Sie ist mit Tasthaaren und warzigen Buckeln bedeckt, die eine taktile Funktion haben. Ebenso wird der gute Geruchssinn eingesetzt, da die Nase sich bei der Suche nur wenige Millimeter über dem Boden befindet, allerdings ist auch der Sehsinn besser entwickelt als bei den meisten anderen Tenreks. Kleinere Beutetiere wie Ameisen und Termiten werden mit der langen und beweglichen Zunge aufgenommen und unmittelbar zerkaut. Die spitzen Höckerchen der Mahlzähne sind speziell an Insektennahrung angepasst, sie vermögen aber auch kleinere Wirbeltiere zu zerlegen. Pflanzliches Material kann mit den Zähnen nur zerschnitten, nicht jedoch zermahlen werden. Daher findet die primäre Verdauung und Auflösung der Pflanzenzellen wohl eher im Magen-Darm-Trakt statt. Allgemein erfolgt die hauptsächliche Manipulation der Nahrung mit dem Maul, die Vorderpfoten spielen kaum eine Rolle.[21][6][3][2][8]
Vergleichbar zu anderen Vertretern der Tenreks besitzt auch der Große Tenrek eine variierende Körpertemperatur, die sich zyklisch über den Tag und das Jahr ändert. Laboruntersuchungen aus den 1950er Jahren ergaben eine Schwankungsbreite zwischen 24,1 und 34,8 °C mit einem Durchschnitt von 28,4 °C. Die Umgebungstemperaturen reichten von 12 bis 24,4 °C. Die Beobachtungen an einem ausgewachsenen Tier ließen damals aber kaum eine Beziehung zwischen der Umgebung und der Aktivität erkennen, lediglich ein Jungtier wurde bei Körpertemperaturen unter 25 °C lethargisch.[22] Andere Untersuchungen zeigten aber deutliche Bindungen zwischen der Außen- und der Körpertemperatur. So bewegte sich die Körpertemperatur innerhalb eines Tages mit 15,2 und 28,1 °C nur wenige Grad über den Außentemperaturen von 12 bis 27 °C.[23] Studien unter annähernd natürlichen Bedingungen bei Perinet im zentral-östlichen Madagaskar erbrachten ähnliche Ergebnisse.[3] Bei Tieren auf den Seychellen schwankt die Körpertemperatur bei aktiven Tieren von 23 bis 35,5 °C, die höchsten Werte stellen sich bei Individuen während der nächtlichen Nahrungsaufnahme ein. Im Ergebnis deuteten die verschiedenen Studien darauf hin, dass die Körpertemperatur beim Großen Tenrek abhängig ist von der Umgebungstemperatur und der Aktivität eines Tieres.[24][25][2][1][8] Abweichend dazu ergab eine Untersuchung aus dem Jahr 2019 stark veränderliche Körpertemperaturen beim Großen Tenrek über den Tag mit Maximalwerten zwischen 12 und 34 °C. Demnach nähert sich die Körper- der Außentemperatur bis zu Tiefstwerten um 12 °C an, selbst bei körperlich aktiven Tieren. Die Schwankungen sind um so heftiger, je niedriger die Umgebungstemperatur ist. Über gewisse kurze Zeiträume des Tages stabilisiert sich die Körpertemperatur, unabhängig davon zeigen trächtige Weibchen eine weniger stark variierende Körpertemperatur, vor allem kurz vor und während der Geburtsphase. Andererseits stellen sich bei einzelnen Individuen in ihrer Aktivitätsperiode lethargische Momente bei Körpertemperaturen um 28 °C ein, die für mehrere Stunden bis Tage anhalten können. Hier tritt aber bei äußeren Störungen relativ schnell wieder ein Wachzustand ein, der wiederum häufig mit einer leicht ansteigenden Körpertemperatur verbunden ist. Generell erweist sich der Große Tenrek als intolerant gegenüber Außentemperaturen von unter 8 beziehungsweise über 34 °C und bildet dann Stresssymptome aus. Schlussfolgernd lässt sich sagen, dass die Körpertemperatur zwar zum Aktivitätszyklus der Tiere beiträgt, diesen aber nicht vollständig steuert.[26]
Den Südwinter beziehungsweise die Trockenzeit auf Madagaskar verbringen die Tiere in einem Torpor. Der Zeitraum reicht etwa von Mai bis Oktober und ist durch einen starken Rückgang des Nahrungsangebotes gekennzeichnet. Im Nationalpark Ankarafantsika wurden Tiere beobachtet, die insgesamt acht bis neun Monate im Starrezustand verbrachten (von Februar/März bis November). Bemerkenswert ist dabei, dass die Unterbrechung des Torpors, die normalerweise alle 12 bis 24 Stunden eintritt und die Tiere vor Langzeitschäden wie Durchblutungs- oder Stoffwechselstörungen bewahrt, beim Großen Tenrek nicht einsetzt, solange die Körpertemperatur über 22 °C liegt.[27] Auf den Seychellen zeigt der Große Tenrek ebenfalls eine Phase des Starrezustands. Sie findet weitgehend von Januar/Februar bis Juli/August statt und dauert wenigstens sechs Monate. Auch hier ist die Phase mit einem starken Schwinden der Nahrungsressourcen verbunden, was mit dem Wechsel des Monsuns von Nordwest nach Südost einhergeht. Männliche Tiere beginnen ihren Torpor dabei durchschnittlich einen Monat früher als weibliche. Die leichte zeitliche Verschiebung zeigt aber zusätzlich, dass sich der Große Tenrek an verschiedene lokale Bedingungen anpassen kann. Auch während der Starrezeit ist ein zyklischer Verlauf der Körpertemperatur bemerkbar, in der Regel liegt sie nur wenige Grad über der Außentemperatur. Vereinzelte starke Anstiege der Körpertemperatur auf bis zu maximal 32 °C in der Nacht sind mit gelegentlichen Aktivitäten der Tiere in Verbindung zu bringen. Manchmal graben die Tiere dabei eine neue Kammer, die bis zu 1 m von der alten entfernt liegen kann.[24][25] In der Regel verbleiben die Tiere aber über den gesamten Zeitraum des Torpors im gleichen Unterschlupf, zu einem eventuellen Ortswechsel kommt es lediglich bei heftigen Regenfällen oder anderen äußeren Einflüssen.[1] Zum Ende der Torporphase kommt es zu einem graduellen Anstieg der Körpertemperatur, der über mehrere Wochen andauert. Die Aufwachphase ist abhängig von den äußeren Bedingungen und kann bei höheren Umgebungstemperaturen innerhalb weniger Sekunden vollzogen sein, bei niedrigen Außentemperaturen aber dementsprechend länger andauern.[26] Neben der Körpertemperatur senkt sich während des Starrezustands auch die Atemfrequenz auf bis zu 30 Atemzüge je Minute, ebenso kann die Herzrate um 34 % zurückgehen. Begleitet wird die jahreszeitlich bedingte Aktivität und Inaktivität des Großen Tenreks von einer zyklischen Veränderung des Körpergewichts. So erhöht sich das Gewicht der Tiere vor Beginn der Starrezeit drastisch, einige Individuen haben dann einen Fettanteil von bis zu 50 % des Körpergewichts. Dies wird teilweise mit einer verstärkten Aufnahme von Früchten erreicht. Das Körpergewicht sinkt dann wieder während der Torporphase um 1,1 bis 1,4 g täglich.[24][25][2][1][8][26]
Fortpflanzung

Die Fortpflanzungsphase beginnt unmittelbar nach dem Ende der Trockenruhe im Frühling (September bis Oktober), sie kann sich, abhängig von der geographischen Lage, um etwa einen Monat verschieben. Auf den eher tropischen Seychellen startet sie einen Monat früher. Es finden ausgeprägte Rivalenkämpfe zwischen den Männchen um das Vorrecht der Paarung statt. Die Kämpfe werden ritualisiert mit den langen Eckzähnen ausgetragen, wobei es dabei auch zu Verletzungen kommen kann. Der bei anderen Tenrekarten beobachtete Nackenbiss ist beim Großen Tenrek modifiziert und die Tiere werfen ihren Kopf bei den Kämpfen seitlich hin und her. Der Sieger wirbt um das Weibchen, indem er ihm folgt und es an verschiedenen Stellen beschnuppert, beginnend an der Nase über die Seiten. Begleitet wird dies von Grunzlauten. Mit dem Aufsteigen des Männchens auf das Weibchen beginnt der Geschlechtsakt. Er dauert etwa 4½ bis 12 Minuten,[28] kann aber auch 25 Minuten anhalten und wird mehrfach unterbrochen.[1][3][2][8]
Die Tragzeit beträgt 58 bis 64 Tage, die Dauer wird wahrscheinlich durch den Torpor des Muttertieres beeinflusst, was ebenfalls vom Erdtenrek (Geogale) belegt ist.[1] Etwa 25 % aller trächtigen Weibchen zeigen eine Polyovulation, bei der bis zu 5 Eizellen in den Follikel eingelagert sind. Eine weitere Besonderheit ist, dass die Follikel sich nicht mit Flüssigkeit füllen, um einen Graafschen Follikel zu bilden, vielmehr reißt der feste Follikel langsam auf und die Eizelle wandert zur Befruchtung zur Oberfläche.[29][1] Kurz vor der Geburt steigert das Weibchen die Nestbauaktivitäten und konstruiert ein kugeliges Pflanzennest im hintersten Teil des Baues. Im Durchschnitt kommen 15 Jungtiere zur Welt, die Wurfgröße kann aber bis zu 32 betragen, der Rekord unter den Säugetieren.[28][7] Generell haben Tenreks aus dem trockenen Süden und Südwesten Madagaskars größere Würfe als solche aus dem feuchteren Osten. Untersuchungen von den Seychellen ergaben ebenfalls geringere Wurfgrößen mit maximal 18 bis 20 Jungen. Außerdem reduziert sich die Wurfgröße mit dem zunehmenden Alter der Tiere: Weibchen in der dritten Fortpflanzungsperiode bringen, bedingt durch intrauterine Sterblichkeit oder Resorption einzelner Föten, durchschnittlich weniger Junge zur Welt als solche in der ersten.[1][8]
Jungtiere kommen als Nesthocker blind und taub zur Welt, sie sind aber weiter entwickelt als bei den meisten anderen Tenreks. Sie sind mit einem charakteristischen, hell-dunkel gestreiften Fell aus 5 mm langen Haaren bedeckt und besitzen gut entwickelte Krallen. Die Geburtsgröße liegt bei 84 bis 92 mm, das Gewicht beträgt 22,8 bis 27,4 g. Außerdem können sie zischende und quiekende Laute ausstoßen und sich auf allen vieren krabbelnd fortbewegen. Das Muttertier ist anfangs besonders wachsam und verteidigt das Nest mit aufgerichteter Halskrause, Bissen und Fußstampfen. Sie leckt ihren Nachwuchs häufig, zum Säugen steht sie schützend über den Jungen. Am siebten Tag sind in den hellen Fellstreifen deutliche Stacheln zu sehen, besonders prominent treten sie am Stridulationsorgan in der Rückenmitte hervor. Ab dem neunten Tag beginnen sich die Augen zu öffnen, der Prozess dauert aber bis zum 14. Tag an. Ein Kratzreflex ist ab 11 Tagen entwickelt, außerdem kann ein Jungtier zu diesem Zeitpunkt bereits seine Vorder- und Hinterbeine besser koordinieren. Mit drei Wochen unternehmen die Jungen erste Streifzüge zur Nahrungssuche mit der Mutter, hierbei hat das Streifenmuster der Jungtiere einen tarnenden Charakter. Sie laufen in einer Reihe hinter der Mutter und ballen sich um sie, wenn sie Nahrung aufnimmt oder stoppt, etwa bei einer möglichen Gefahr. Bei letzterem stridulieren die Jungen dann, was mit dem Aufstellen der Stacheln und Flucht verbunden ist. Die Funktion der Stridulation bei Jungtieren ist nicht ganz geklärt, möglicherweise übertragen sie damit auch Informationen zu ihrem Standort oder warnen andere Nestmitglieder. Die bei den Rasselgeräuschen der Stacheln erzeugten Frequenzen liegen im Bereich von 12 bis 15 kHz.[9] Ebenfalls in dieser Periode beginnt die Zeit der Entwöhnung, die Jungtiere haben dann das bis zu 40fache ihres Geburtsgewichtes erreicht.[26] Mit etwa 35 Tagen nimmt die Aufmerksamkeit der Mutter ab und die Jungen gehen häufiger alleine auf Nahrungssuche. Von da an bis zum 60. Tag wechselt das Fell und die Jungen bekommen das Haarkleid der Alttiere. Sie sind dann zu Beginn dieses Prozesses durchschnittlich 16 cm lang. Danach löst sich der Zusammenhalt der Familiengruppe auf.[20][3][2] Die maximale Lebenserwartung von Tieren in menschlicher Gefangenschaft war bisher knapp neun Jahre,[30] für Tiere in freier Wildbahn liegen kaum Daten vor. Auf den Seychellen erreichten nur wenige Weibchen das vierte Lebensjahr.[1][8]
Fressfeinde und Parasiten
Zu den wichtigsten Fressfeinden gehört die Fossa, nach Analysen von 114 Kotproben des Raubtiers aus dem Waldgebiet von Kirindy erreicht der Große Tenrek aber nur einen geringen Individuen- und Biomasseanteil im gesamten Beutespektrum (jeweils rund 2,5 %).[31] Ähnlich selten wurde die Art auch im Nationalpark Ankarafantsika durch die Fossa erbeutet, die meisten Tiere kamen zudem im Südsommer zu Tode.[32][33] Von Bedeutung sind außerdem der Madagaskar-Bussard und die Nördliche Madagaskarboa.[27][3] Der Große Tenrek kann sich im Gegensatz zum Großen oder zum Kleinen Igeltenrek nicht zu einer Kugel zusammenrollen. Im Falle eines Angriffs oder einer Berührung richtet er die borstigen Nackenhaare auf, gibt Zisch- oder Quietschlaute von sich und stampft mit den Füßen. Außerdem zeigt er ein geöffnetes Maul oder beißt. In der Regel flieht ein Tier aber und kann dabei Geschwindigkeiten von 3,6 bis 4,7 km/h erreichen.[3] Als äußere Parasiten sind Flöhe der Gattung Paractenopsyllus[34][35] und Milben der Gattung Andreacarus identifiziert,[36] innere Parasiten schließen neben Fadenwürmern[3] unter anderem den Einzeller Eimeria ein.[37] Außerdem tritt der Große Tenrek als Wirt von Leptospira-Bakterien auf.[38]
Systematik
Innere Systematik der Tenreks nach Everson et al. 2016[39]
|
Der Große Tenrek ist eine Art aus der Gattung Tenrec, die wiederum zur Familie der Tenreks (Tenrecidae) gehört. Innerhalb dieser bildet er zusammen mit dem Großen Igeltenrek (Setifer setosus), dem Kleinen Igeltenrek (Ehinops telfairi) sowie den Streifentenreks (Hemicentetes) die Unterfamilie der Igeltenreks (Tenrecinae). Die Igeltenreks stellen eine der drei Hauptlinien der Tenreks dar. Ihr stacheliges Haarkleid setzt sie von den übrigen weichhaarigen Tenreks wie den sehr diversen Reiswühlern (Oryzorictinae) und den monotypischen Erdtenreks (Geogalinae) ab. Laut molekulargenetischen Untersuchungen ist der Große Tenrek näher mit den Streifentenreks verwandt. Beide Gattungen vereint neben dem weniger stacheligen Haarkleid im Vergleich zu den Igeltenreks das Streifenmuster und die Befähigung zur Stridulation, beides ist beim Großen Tenrek allerdings nur bei Jungtieren ausgebildet. Ihnen gegenüber wiederum steht eine Klade, die sich aus dem Großen und dem Kleinen Igeltenrek zusammensetzt.[40][41][39] Eine nähere Verwandtschaft zwischen dem Großen Tenrek und den Streifentenreks wurde auch schon aus morphologischen Gründen erwogen.[42] Die Linien des Großen Tenreks und der Streifentenreks trennten sich bereits im ausgehenden Unteren Miozän vor etwa 16,1 Millionen Jahren auf.[39]
Die Gattung Tenrec enthält nur eine Art, ebenso sind keine Unterarten des Großen Tenreks bekannt. Sowohl Gattung als auch Art sind daher als monotypisch anzusehen.[43]
Tenrec und Centetes – Zur Forschungsgeschichte der Gattung


Erstmals wurde in Europa durch Étienne de Flacourt, ehemals Gouverneur von Madagaskar, über Tenreks berichtet. In seinem im Jahr 1658 veröffentlichten und mehrfach wieder aufgelegten Reisebericht erwähnte er ein igelartiges Tier und bezeichnete es als „Tendrac“.[44] Heute wird der „Tendrac“ allgemein mit dem Großen Igeltenrek in Verbindung gebracht. Gut einhundert Jahre später beschrieb Georges-Louis Leclerc de Buffon im Jahr 1764 im zwölften Band seines Werkes Histoire naturelle, générale et particulière den „Tendrac“ genauer und stellte ihm den „Tanrec“ zur Seite. Das Werk enthielt auch Abbildungen der beiden Formen. Nach Aussagen Buffons war der „Tanrec“ deutlich größer als der „Tendrac“, er hatte außerdem eine längere Schnauze und besaß Stacheln nur im Nacken, während den restlichen Körper Borsten bedeckten. Buffons Ausführungen zum „Tendrac“ und zum „Tanrec“ basierten auf zwei Tieren, die sich zur damaligen Zeit im Cabinet du roi, dem Vorgänger des Muséum national d’histoire naturelle in Paris, befanden und aus einer Schenkung eines Händlers aus Lyon stammten.[45] Späteren Angaben zufolge wies der „Tanrec“ eine Länge von 21 cm auf.[46] Im Jahr 1777 legte dann Johann Christian von Schreber in seinem umfassenden Werk Die Säugthiere in Abbildungen nach der Natur mit Beschreibungen eine deutschsprachige Beschreibung des „Tanrec“ vor. Er stützte sich dabei auf Buffon und versah seine Beschreibungen ebenfalls mit einer Abbildung, die an jene aus der Histoire naturelle angelehnt war. Die Abbildung enthielt außerdem die wissenschaftliche Bezeichnung Erinaceus ecaudatus, weswegen Schreber als der Erstbeschreiber des Großen Tenreks gilt. Der Artname ecaudatus ist lateinischen Ursprungs und bezieht sich auf den kaum sichtbaren Schwanz (von ecaudis für „schwanzlos“, hergeleitet von cauda für „Schwanz“).[47]

Schreber hatte mit dem Artnamen Erinaceus ecaudatus den „Tanrec“ zu den heutigen Kleinohrigeln gestellt. Die Zuweisung der damals bekannten Formen der Tenreks zu den Igeln war im ausgehenden 18. Jahrhundert durchaus üblich. Gut zwei Dekaden nach Schrebers Veröffentlichung, 1798, trennte Georges Cuvier die Tenreks (Tenrecs) erstmals formell von den Igeln (Hérissons) ab, benannte aber keine neue Gattung.[48] Dies blieb dann Bernard Germain Lacépède im folgenden Jahr vorbehalten, der den Gattungsnamen Tenrec für den Großen Tenrek prägte. Er charakterisierte die Gattung über drei gleich große Schneidezähne, einen dreimal so langen Eckzahn und über den stachelbesetzten Körper.[49] Johann Karl Wilhelm Illiger etablierte im Jahr 1811 die Gattung Centetes und setzte sie wie zuvor Lacépède mit Tenrec auch in direkten Bezug zu Erinaceus ecaudatus. Seine Merkmalsbeschreibung ähnelte der von Lacépède, zusätzlich hob Illiger noch die lange Schnauze und die fünfstrahligen Hände und Füße hervor. Der Name Centetes ist der griechischen Sprache entnommen (κεντητήϛ kentetes) und leitet sich von κεντητος (kentetos) für „gestickt“ oder κεντεω (kenteo) für „stechen“ her.[50] Auf Illigers Centetes basiert auch der von Georges Cuvier geschaffene Name Centenes,[51] der gelegentlich in Gebrauch war, aber als Falschschreibung anzusehen ist.[52] Von Anselme Gaëtan Desmarest stammt wiederum die Bezeichnung Tenrecus, welche er 1820 einführte und Lacépède zuschrieb.[53]
Der Gattungsname Tenrec wurde nach seiner Einführung durch Lacépède von den meisten Autoren weitgehend ignoriert.[52] Vielmehr fand Centetes weite Verbreitung im 19. Jahrhundert und galt darüber bis zur Mitte des 20. Jahrhunderts als gültige Gattungsbezeichnung. Auf Centetes beruht auch der Familienname Centetidae, den Andrew Murray im Jahr 1866 kreierte. Murray setzte damit die Tenreks von den Igeln ab, er schloss alle damals bekannten Tenrekgattungen in die Centetidae ein (Centetes, Ericulus und Echinogale; heute unter Tenrec, Setifer und Echinops geführt).[54] Zuvor hatte bereits 1838 Charles Lucien Jules Laurent Bonaparte die Centetina als eine Untergruppe in die Igel eingegliedert.[55] Im Jahr 1945 wies George Gaylord Simpson in seiner Taxonomie der Säugetiere darauf hin, dass Centetes als Gattungsbezeichnung nicht gültig ist, da Tenrec bereits zuvor benannt wurde.[56]
Die Eigenständigkeit des Großen Tenreks war nie Gegenstand des wissenschaftlichen Disputes und ist allgemein anerkannt. Lediglich George Shaw sah im Jahr 1800 die Form als Varietät der Streifentenreks an,[57] was aber kaum anerkannt wurde.[46] Im Jahr 1837 führte Saint Hilaire mit wenigen Worten Centetes armatus ein.[58] Nur zwei Jahre später beschrieb er die Form genauer, sie basierte auf einem Individuum, dass dem Muséum national d’histoire naturelle von einem französischen Marinekapitän übergeben worden war.[52] Allerdings ließen sich kaum Unterschiede zum Großen Tenrek ausmachen, so dass Centetes armatus bereits im 19. Jahrhundert als synonym zu Tenrec ecaudatus galt.[46][5] Ein weiteres, nur selten gebrauchtes Synonym ist das von Pieter Boddaert schon 1785 verwendete Erinaceus tanrec.[59][43]
Stammesgeschichte
Die Tenreks sind in der Regel nur selten fossil nachweisbar. Dies trifft auch auf den Großen Tenrek zu. Ein einzelner Unterkiefer eines nicht ausgewachsenen Tieres wurde in der Ankilitelo-Höhle entdeckt. Die Höhle befindet sich am Südrand des Mikoboka-Plateaus nördlich von Toliara im südwestlichen Madagaskar. Sie wird seit dem Jahr 1994 wissenschaftlich untersucht. Ihre sehr reichhaltigen Fauna-Funde setzen sich aus Knochen und Zähnen von verschiedenen Primaten, Fleder- und Nagetieren zusammen, darüber hinaus sind rund ein halbes Dutzend Arten an Tenreks überliefert. Der Große Tenrek ist aber im Vergleich zum Großen und zum Kleinen Igeltenrek, zum Erdtenrek (Geogale) oder zu den verschiedenen Vertretern der Kleintenreks (Microgale) ein eher seltenes Faunenelement. Das Alter der Funde beträgt etwa 510 bis 630 Jahre.[60] Ein weiterer Fundort ist die Andrahomana-Höhle im äußersten Südosten Madagaskars. Diese liegt eingebettet in den Eolianit-Aufschlüssen der Steilküste und wird bereits seit dem Ende des 19. Jahrhunderts erforscht. Ihr ebenfalls ungemein reiches Fossilmaterial, das die letzten rund 8.700 Jahre abdeckt, umfasst Reptilien, Vögel und Säugetiere, darunter auch mehrere Tenrek-Arten wie den Großen Tenrek.[61] An beiden Fundregionen herrschen hauptsächlich trockene Klimate vor, jedoch wirken im Bereich der Andrahomana-Höhle noch teilweise die feuchten Bedingungen des östlichen Madagaskars ein.[60][61][62]
Bedrohung

Der Große Tenrek ist zu einem gewissen Grad Kulturfolger und kommt sowohl auf Agrar- und Farmland als auch in menschlichen Siedlungen vor. Die Rodung von Wäldern hat keinen großen Einfluss auf den Bestand der Art,[63] in den Trockenwäldern des Südwestens können aber Waldbrände lokal eine Bedrohung sein. In weiten Bereichen Madagaskars wird der Große Tenrek seines Fleisches wegen gejagt. In einigen Gebieten, hauptsächlich um größere Städte herum, kann er durch Überjagung daher verschwunden sein. Regional gibt es bezüglich der Jagd auf den Großen Tenrek einzelne Tabus, etwa in zentral-östlichen Madagaskar, wo die Tiere nur zum Ende des Südsommers im April/Mai erbeutet werden dürfen.[64][65] Ebenso gelten die Tiere auf den Maskarenen als Nahrungsressource, während sie auf den traditionell islamischen Komoren lange Zeit einem Tabu unterlagen, dieses scheint aber in jüngerer Zeit gelockert zu sein.[1] Die IUCN stuft den Großen Tenrek aufgrund seiner weiten Verbreitung und seiner Anpassungsfähigkeit als „nicht gefährdet“ (least concern) ein. Er kommt in nahezu allen Naturschutzgebieten Madagaskars vor, zu den wichtigsten zählen etwa der Nationalpark Montagne d’Ambre, der Nationalpark Marojejy, der Nationalpark Masoala, der Nationalpark Ankarafantsika, der Nationalpark Isalo, der Nationalpark Zombitse, der Nationalpark Ranomafana, der Nationalpark Andringitra und der Nationalpark Andohahela.[17]
Seit der Haltungsaufgabe 2015 in Stuttgart wird die Art in Deutschland nicht mehr gepflegt. In Europa gibt es sie in Frankreich, Großbritannien, Polen und Tschechien zu sehen.[66]
Literatur
- J. F. Eisenberg und Edwin Gould: The Tenrecs: A Study in Mammalian Behavior and Evolution. Smithsonian Institution Press, 1970, S. 1–138
- Kathryn M. Everson, Voahangy Soarimalala, Steven M. Goodman und Link E. Olson: Multiple loci and complete taxonomic sampling resolve the phylogeny and biogeographic history of tenrecs (Mammalia: Tenrecidae) and reveal higher speciation rates in Madagascar’s humid forests. Systematic Biology 65 (5), 2016, S. 890–909 doi: 10.1093/sysbio/syw034
- Nick Garbutt: Mammals of Madagascar. A complete guide. Yale University Press, 2007, S. 1–304 (S. 32–56)
- Paulina D. Jenkins: Tenrecidae (Tenrecs and Shrew tenrecs). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 134–172 (S. 164) ISBN 978-84-16728-08-4
- Martin Nicoll: The common tenrec, Tenrec ecaudatus. Afrotherian Conservation 7, 2009, S. 2–3
- Ronald M. Nowak: Walker's Mammals of the World. The Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9
Einzelnachweise
- Martin Nicoll: The common tenrec, Tenrec ecaudatus. Afrotherian Conservation 7, 2009, S. 2–3
- Nick Garbutt: Mammals of Madagascar. A complete guide. Yale University Press, 2007, S. 1–304 (S. 32–56)
- J. F. Eisenberg und Edwin Gould: The Tenrecs: A Study in Mammalian Behavior and Evolution. Smithsonian Institution Press, 1970, S. 1–138
- Olga V. Zherebtsova: Morphofunctional interpretation of the quills stridulating in tenrecs (Lipotyphla, Tenrecidae). Russian Journal of Theriology 5 (1), 2006, S. 1–11
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part I. London, 1883, S. 68–69 ()
- Konrad Herter: Über die Borstenigel von Madagaskar (Tenrecinae). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin N. F. 2, 1962, S. 5–37
- Walter Poduschka: Hyperthelie, Wurfgröße und Trächtigkeitsdauer bei der Unterfamilie Tenrecinae Cabrera, 1925 (Mammalia: Insectivora: Tenrecidae), nebst Bemerkungen zur Längsstreifung der Gattung Hemicentetes. Contributions to Zoology 66 (2), 1996, S. 119–128
- Paulina D. Jenkins: Tenrecidae (Tenrecs and Shrew tenrecs). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 134–172 (S. 164) ISBN 978-84-16728-08-4
- Edwin Gould: Evidence for echolocation in the Tenrecidae of Madagaskar. Proceedings of the American Philosophical Society 109 (6), 1965, S. 352–360
- Wilhelm Leche: Zur Entwicklungsgeschichte des Zahnsystems der Säugetiere, zugleich ein Beitrag zur Stammesgeschichte dieser Tiergruppe. Zweiter Teil: Phylogenie. Zweites Heft: Familien der Centetidae, Solenodontidae und Chrysochloridae. Zoologica 20, 1906/1908, S. 1–157 ()
- Steven M. Goodman und Paulina D. Jenkins: Tenrecs (Lipotyphla; Tenrecidae) of the Parc National de Marojejy, Madagascar. Fieldiana Zoology 97, 2000, S. 201–229
- Justine A. Salton und Eric J. Sargis: Evolutionary morphology of the Tenrecoidea (Mammalia) carpal complex. Biological Journal of the Linnean Society, 93, 2008, S. 267–288
- Justine A. Salton und Eric J. Sargis: Evolutionary Morphology of the Tenrecoidea (Mammalia) Forelimb Skeleton. In: E.J. Sargis und M. Dagosto (Hrsg.): Mammalian Evolutionary Morphology: A Tribute to Frederick S. Szalay, Springer Science, 2008, S. 51–71
- Justine A. Salton und Frederick S. Szalay: The Tarsal Complex of Afro-Malagasy Tenrecoidea: A Search for Phylogenetically Meaningful Characters. Journal of Mammalian Evolution 11 (2), 2004, S. 73–104
- Voahangy Soarimalala: Les Afrosoricides de la forêt sèche malgache. Afrotherian Conservation 8, 2011, S. 4–9
- David Ross Stoddart: Biogeography and ecology of the Seychelles Islands. Springer, 1984. ISBN 906193107X, S. 616
- P. J. Stephenson, Voahangy Soarimalala und Steven M. Goodman: Tenrec ecaudatus. The IUCN Red List of Threatened Species 2016. e.T40595A97204107 (); zuletzt abgerufen am 18. April 2017
- A. L. Rand: On the Habits of Some Madagascar Mammals. Journal of Mammalogy 16 (2), 1935, S. 89–104
- J. J. Petter und A. Petter-Rousseaux: Notes biologiques sur les Centetinae. La Terre et la Vie 17 (1), 1963, S. 66–80
- Edwin Gould und John F. Eisenberg: Notes on the biology of the Tenrecidae. Journal of Mammalogy 47 (4), 1966, S. 660–686
- M. Ade: Examination of the digestive tract contents of Tenrec ecaudatus Schreber 1777 (Tenrecidae, Insectivora) from Western Madagascar. Primate Report 46-1, 1996, S. 233–249
- M. Eisentraut: Temperaturschwankungen bei niederen Säugetieren. Zeitschrift für Säugetierkunde 21, 1956, S. 49–52
- C. Kayser: Consommation d’oxygène et temperature central au cours de l’hiver austral de deux insectivores de Madagascar, Centetes ecaudatus et Setifer setosus. Comptes rendus des séances de la Société de Biologie 154, 1960, S. 1873–1876 ()
- Martin Nicoll: Responses to Seychelles tropical forest seasons by a litter-foraging mammalian insectivore, Tenrec ecaudatus, native to Madagascar. Journal of Animal Ecology 54, 1985, S. 71–88
- Martin Nicoll: Diel variation in body temperature in Tenrec ecaudatus during seasonal hypothermia. Journal of Mammalogy 67, 1986, S. 759–762
- Michael D. Treat, Lori Scholer, Brandon Barrett, Artur Khachatryan, Austin J. McKenna, Tabitha Reyes, Alhan Rezazadeh, Charles F. Ronkon, Dan Samora, Jeremy F. Santamaria, Claudia Silva Rubio, Evan Sutherland, Jeffrey Richardson, John R. B. Lighton und Frank van Breukelen: Extreme physiological plasticity in a hibernating basoendothermic mammal, Tenrec ecaudatus. Journal of Experimental Biology 221, 2018, S. jeb185900 doi:10.1242/jeb.185900
- Barry G. Lovegrove, Kerileigh D. Lobban und Danielle L. Levesque: Mammal survival at the Cretaceous-Palaeogene boundary: metabolic homeostasis in prolonged tropical hibernation in tenrecs. Proceedings of the Royal Society B 281, 2014, S. 20141304 doi:10.1098/rspb.2014.1304
- John F. Eisenberg: Tenrecs and solenodons in captivity. International Zoo Yearbook 15, 1975, S. 6–12
- Martin Nicoll und P. A. Racey: Follicular development, ovulation, fertilization and fetal development in tenrecs (Tenrec ecaudatus). Journal of Reproduction and Fertility 74 (1), 1985, S. 47–55
- Richard Weigl: Longevity of mammals in captivity; from the Living Collections of the world A list of mammalian longevity in captivity. Kleine Senckenberg-Reihe, Band 48, 2005, S. 1–214 (S. 35)
- Rodin M. Rasoloarison, Bernardin P. N. Rasolonandrasana, Jörg U. Ganzhorn und Steven M. Goodman: Predation on vertebrates in the Kirindy forest, Western Madagascar. Ecotropica 1, 1995, S. 59–65
- Luke Dollar, Jörg U. Ganzhorn und Steven M. Goodman: Primates and other prey in the seasonally variable diet of Cryptoprocta ferox in the deciduous forest of Western Madagascar. In: S. Gursky und K. A. Nekaris (Hrsg.): Predation on primates. New York, 2007, S. 63–76
- Clare E. Hawkins und Paul A. Racey: Food habits of an endangered carnivore, Cryptoprocta ferox in the dry deciduous forests of Western Madagascar. Journal of Mammalogy 89 (1), 2008, S. 64–74
- Jean-Claude Beaucournu, H. Rico Randrenjarison Andriniaina und Steven M. Goodman: Puces (Insecta: Siphonaptera) d’Ambohitantely, Madagascar: Spécificité et phénologie. Malagasy Nature 9, 2015, S. 39–48
- Michael W. Hastriter und Carl W. Dick: A description of Paractenopsyllus madagascarensis sp. n. and the female of Paractenopsyllus raxworthyi Duchemin & Ratovonjato, 2004 (Siphonaptera, Leptopsyllidae) from Madagascar with a key to the species of Paractenopsyllus. ZooKeys 13, 2009, S. 43–55
- Ashley P. G. Dowling, Andre V. Bochkov und Barry M. Oconnor: Revision of the genus Andreacarus (Acari: Laelapidae) with description of seven new species and a new genus for Australian species formerly placed in Andrreacarus. Journal of Medical Entomology 44 (3), 2007, S. 405–421
- Lee Couch, Juha Laakkonen, Steven Goodman und Donald W. Duszynski: Two New Eimerians (Apicomplexa) from Insectivorous Mammals in Madagascar. Journal of Parasitology 97 (2), 2011, S. 293–296
- Erwan Lagadec, Yann Gomard, Gildas Le Minter, Colette Cordonin, Eric Cardinale, Beza Ramasindrazana, Muriel Dietrich, Steven M Goodman, Pablo Tortosa und Koussay Dellagi: Identification of Tenrec ecaudatus, a Wild Mammal Introduced to Mayotte Island, as a Reservoir of the Newly Identified Human Pathogenic Leptospira mayottensis. PLoS Neglected Tropical Diseases 10 (8), 2016, S. e0004933. doi:10.1371/journal.pntd.0004933
- Kathryn M. Everson, Voahangy Soarimalala, Steven M. Goodman und Link E. Olson: Multiple loci and complete taxonomic sampling resolve the phylogeny and biogeographic history of tenrecs (Mammalia: Tenrecidae) and reveal higher speciation rates in Madagascar’s humid forests. Systematic Biology 65 (5), 2016, S. 890–909 doi: 10.1093/sysbio/syw034
- Link E. Olson und Steven M. Goodman: Phylogeny and biogeography of tenrecs. In: Steven M. Goodman und Jonathan P. Benstead (Hrsg.): The natural history of Madagascar. University of Chicago Press, 2003, S. 1235–1242
- Matjaž Kuntner, Laura J. May-Collado und Ingi Agnarsson: Phylogeny and conservation priorities of afrotherian mammals (Afrotheria, Mammalia). Zoologica Scripta 40 (1), 2011, S. 1–15
- Henri Heim de Balsac: Insectivores. In: R. Battistini und G. Richard-Vindard (Hrsg.): Biogeography and ecology in Madagascar. Den Haag, 1972, S. 629–660
- Don E. Wilson und DeeAnn M. Reeder: Mammal Species of the World. A taxonomic and geographic Reference. Johns Hopkins University Press, 2005 ()
- Étienne de Flacourt: Histoire de la grande isle Madagascar. Paris, 1661, S. 1–471 (S. 152) ()
- Georges-Louis Leclerc de Buffon: Histoire naturelle, générale et particulière., Tome Douzième. Paris, 1764, S. 440–441, S. 450 und Tafel 56 ()
- Fredericus Anna Jentink: On the hedgehogs from Madagascar. Notes from the Royal Zoological Museum at Leyden 1879, S. 137–151 ()
- Johann Christian Daniel Schreber: Die Säugthiere in Abbildungen nach der Natur mit Beschreibungen. Dritter Teil. Erlangen, 1777, S. 584 () und Tafelband 1 Tafel 165 ()
- Georges Cuvier: Tableau elementaire de l’histoire naturelle des animaux. Paris, 1798, S. 1–710 ( 108) ()
- Bernard Germain Lacépède: Tableau des divisions, sous-divisoins, ordres et genres des mammifères. Paris, 1799; Nachdruck in den Memoirs de l'Institut national des sciences et arts 3, 1801, S. 489–519 (S. 492) ()
- Johann Karl Wilhelm Illiger: Prodromus systematis mammalium et avium additis terminis zoographicis utriudque classis. Berlin, 1811, S. 1–301 (S. 124) ()
- Georges Cuvier: Le règne animal distribué d’après son organisation. Paris, 1817, S. 1–540 (S. 166) ()
- Étienne Geoffroy Saint-Hilaire: Tanrec. Cuv. Centetes. Illig. et Éricule. Ericulus. Is. Geoff. Magasin de Zoologie Serie 2 1, 1839, S. 1–37 ()
- Anselme Gaëtan Desmarest: Mammalogie, ou, Description des espèces de mammifères. Paris, 1820, S. 1–276 (S. 162) ()
- Andrew W. Murray: The geographic distribution of mammals. London, 1866, S. 1–420 (S. XIV) ()
- Charles Lucien Jules Laurent Bonaparte: Synopsis vertebratorum systematis. Nuovi Annali delle Scienze Naturali 2, 1838, S. 105–133 ()
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 50 und 175–179)
- George Shaw: General zoology. Vol. 1 London, 1800, S. 1–552 (S. 548) ()
- Étienne Geoffroy Saint-Hilaire: Notice sur les mammiferes épineux de Madagascar. Comptes rendus hebdomadaires des séances de l'Académie des sciences 5, 1837, S. 372–374 ()
- Pieter Boddaert: Elenchus animalium. Rotterdam, 1785, S. 1.174 (S. 129) ()
- Kathleen M. Muldoon, Donald D. de Blieux, Elwyn L. Simons und Prithijit S. Chatrath: The Subfossil Occurrence and Paleoecological Significance of Small Mammals at Ankilitelo Cave, Southwestern Madagascar. Journal of Mammalogy 90 (5), 2009, S. 1111–1131
- D. A. Burney, N. Vasey, L. R. Godfrey, Ramilisonina, W. L. Jungers, M. Ramarolahy und L. Raharivony: New Findings at Andrahomana Cave, Southeastern Madagascar. Journal of Cave and Karst Studies 70 (1), 2008, S. 13–24
- Steven M. Goodman und William L. Jungers: Extinct Madagaskar. Picturing the island's past. University of Chicago Press, 2014, S 1–206 (S. 65–73 und 94–101)
- J. U. Ganzhorn, A. W. Ganzhorn, J.-P. Abraham, L. Andriamanarivo und A. Ramananjatovo: The impact of selective logging on forest structure and tenrec populations in western Madagascar. Oecologia 84, 1990, S. 126–133
- Julia P. G. Jones, Mijasoa M. Andriamarovololona und Neal Hockley: The Importance of Taboos and Social Norms to Conservation in Madagascar. Conservation Biology 22 (4), 2008, S. 976–986
- Kim E. Reuter und Brent J. Sewall: Taboos and sustainability of tenrec hunting in Madagascar. Afrotherian Conservation 12, 2016, S. 11–15
- Zootierliste (); zuletzt abgerufen am 6. Mai 2017
Weblinks
- Tenrec ecaudatus in der Roten Liste gefährdeter Arten der IUCN 2017. Eingestellt von: P. J. Stephenson, Voahangy Soarimalala und Steven M. Goodman, 2014. Abgerufen am 18.4.2017.