Mimivirus
Mimivirus ist eine Gattung von Viren aus der Familie der Mimiviridae,[6] denen Amöben als natürliche Wirte dienen. Mit der Familie Mimiviridae gehört Mimivirus zu den Riesenviren im Phylum der Nucleocytoviricota (auch englisch Nucleocytoplasmic large DNA viruses, NCLDV; ursprünglicher Vorschlag „Nucleocytoplasmaviricota“; ein noch früherer Vorschlag ist der einer Ordnung „Megavirales“ s. l.[7] [1]).
| Acanthamoeba polyphaga mimivirus | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
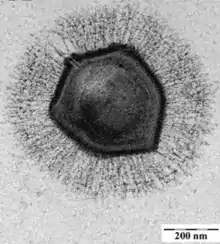
EM-Aufnahme eines Virions (Virusteilchens) des | ||||||||||||||||||||||
| Systematik | ||||||||||||||||||||||
| ||||||||||||||||||||||
| Taxonomische Merkmale | ||||||||||||||||||||||
| ||||||||||||||||||||||
| Wissenschaftlicher Name | ||||||||||||||||||||||
| Acanthamoeba polyphaga mimivirus | ||||||||||||||||||||||
| Kurzbezeichnung | ||||||||||||||||||||||
| APMV | ||||||||||||||||||||||
| Links | ||||||||||||||||||||||
|
In der Gattung Mimivirus gibt es mit Stand März 2019 eine einzige vom International Committee on Taxonomy of Viruses (ICTV) bestätigte Art Acanthamoeba polyphaga mimivirus (APMV), die daher auch die Typusart ist.[8] In der Umgangssprache und in älterer Literatur wird APMV üblicherweise nur als Mimivirus bezeichnet. Es gibt jedoch eine ganze Reihe phylogenetisch verwandter großer Viren, die als weitere Mitglieder dieser Gattung vorgeschlagen sind.
Entdeckung und Forschungsgeschichte
Das Mimivirus APMV wurde 1992 bei Forschungsarbeiten über die Legionärskrankheit (Legionellose) in einem Industriekühlturm in Bradford (England) entdeckt, wobei man feststellte, dass es sich in der Amöbe Acanthamoeba polyphaga vermehrt. Im Jahr 2003 wurde es an der Université de la Méditerranée in Marseille von einer Arbeitsgruppe um Didier Raoult identifiziert.[9] Mit einem Durchmesser von 400 nm haben die Viruspartikel (Virionen) von APMV die Größe von kleinen Bakterien.[10] Wegen dieser Größe und der äußeren Ähnlichkeit mit kugelförmigen Bakterien (Kokken) hielt man es zunächst für ein grampositives Bakterium und nannte es Bradfordcoccus. Als man den Irrtum erkannte, benannte man das neu entdeckte Virus in Anspielung auf seine Größe und Färbungseigenschaften Mimicking Virus, täuschendes Virus. Schließlich wurde daraus kurz Mimivirus, mit dem Namensteil mimi als eine Abkürzung für englisch mimicking microbe. Im Oktober 2004 wurde dann von Didier Raoult und Kollegen die Struktur seines Erbguts in der Fachzeitschrift Science veröffentlicht.[11] Mimivirus blieb übrigens nicht das einzige Virus, dem das passierte: Ein weiteres Beispiel ist das Misannotatedvirus (vorgeschlagen für Pithoviridae), zunächst Amina Cherif Louazani, Sarah Aherfi, Rania Francis, Rodrigo Rodrigues, Ludmila Santos Silva, Dehia Sahmi, Said Mougari, Nisrine Chelkha, Meriem Bekliz, Lorena Silva, Felipe Assis, Fábio Dornas, Jacques Yaacoub Bou Khalil, Isabelle Pagnier, Christelle Desnues, Anthony Levasseur, Philippe Colson, Jônatas Abrahão, Bernard La Scola.[12]
Das gleiche Team, das das APMV entdeckte, entdeckte später ein etwas größeres Virus, das Acanthamoeba castellanii mamavirus (ACMV, kurz Mamavirus) zusammen mit dem Sputnik-Virophagen, der es infiziert. ACMV und APMV sind so nahe verwandt, dass sie gewöhnlich in dieselbe Gattung Mimivirus gestellt werden.[13]
Bis 2013, als ein noch größeres Virus, das Pandoravirus, beschrieben wurde, wiesen die Viren der Gattung Mimivirus den größten Kapsiddurchmesser aller bekannten Viren auf; inzwischen wird es aber auch von nahen Verwandten Megavirus chilensis, Tupanvirus, Platanovirus, sowie anderen Riesenviren wie dem Pithovirus übertroffen.
Wirte
Der erste bekannte Mimivirus-Wirt ist die Amöbe Acanthamoeba polyphaga (Gattung Acanthamoeba, Amoebozoa). Bisher konnten labormäßig nur Vertreter der Gattung Acanthamoeba, außer A. polyphaga noch A. castellanii und A. mauritaniensis, als Wirte dieses Virus eingesetzt werden, keine Zellen anderer einzelliger oder mehrzelliger Organismen.[14] Die natürlichen Wirte sind unbekannt (Stand 2015).[15]
Aufbau
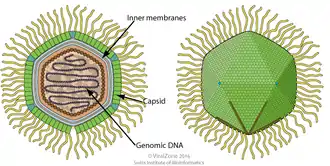

Die Kapside der Virionen (Viruspartikel) von APMV erscheinen unter einem Elektronenmikroskop sechseckig, daher ist die Kapsidgeometrie ikosaedrisch.[17] Es scheint keine äußere virale Hülle zu geben, was darauf hindeutet, dass das Virus die Wirtszelle nicht durch Exozytose verlässt.[14]
Das Hauptprotein des Mimivirus-Kapsids besteht aus zwei Domänen vom Biskuitrollen-Typ (englisch Jelly roll fold). Dieses Protein bildet homotrimere Kapsomere als Organisationseinheit der Kapside. Die Kapsomere sind hexagonal in Form von „Gänseblümchen“ gepackt: Sechs Kapsomere umgeben eine Vertiefung zwischen ihnen.[17][18]
Die Virionen von Mimivirus haben einen Kapsiddurchmesser von 400 nm.
Fibrillen
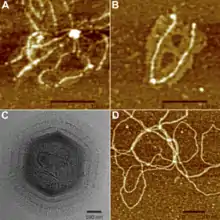
B: AFM-Aufnahme von zwei abgelösten Oberflächenfasern des Mimivirus.
C: Cryo-EM-Bild eines Mimivirus nach teilweisem Abbau der Fibrillen mittels Bromelain.
D: AFM-Bild von inneren Fasern des Mimivirus.
Das Kapsid ist mit einer kompakten Schicht von Fibrillen bedeckt (Tegument). Die aus der Oberfläche des Kapsids herausragenden Proteinfilamente (Fibrillen) haben eine Länge von etwa 100 nm (80–125 nm), und bringen damit die Gesamtlänge eines Virions auf 600 nm.[1][15] Abweichungen in der wissenschaftlichen Literatur lassen die Zahlen als sehr ungenau erscheinen, wenn zum Beispiel die ‚Größe’ des Virions gelegentlich als irgendwo zwischen 400 und 800 nm angegeben wird. Abgesehen von Unterschieden zwischen den einzelnen Virusstämmen in der Gattung Mimivirus ist manchmal die Gesamtgröße mit Filamenten, und manchmal der reine Kapsiddurchmesser angegeben.
Untersuchungen dieser Filamente von Klose et al. (2010) unter Verwendung eines Rasterkraftmikroskops ergeben, dass diese häufig an einer gemeinsamen Tragstruktur befestigt sind. Es konnte jedoch damals noch nicht herausgefunden werden, an welchen Teilen der Kapsidoberfläche diese Träger befestigt sind.[17] Jede Fibrille endet mit einer kleinen kugeligen Kappe aus einem Protein mit unbekannter Funktion.[15][17] Die Proteinfilamente erwiesen sich als resistent gegen Proteasen, außer sie wurden mit Lysozym behandelt. Die Filamente schienen deshalb mit Peptidoglycan beschichtet zu sein. Das war alles in guter Übereinstimmung mit der Tatsache, dass sich das Mimivirus durch die Gram-Methode anfärben lässt.[19]
Die Filamente spielen mit ihrer stark glykosylierten Oberfläche offenbar eine wichtige Rolle bei der Annäherung an die Wirtsamöben und der nachfolgenden Infektion.[19][15] Der Hauptbestandteil der Fasern ist das Protein R135 (neben L725 und L829). Seine Struktur ähnelt Proteinen aus der Familie der Glucose-Methanol-Cholin-Oxidoreduktasen (GMC-Oxidoreduktasen), die eine N-terminale FAD-Bindungsdomäne und eine C-terminale Substraterkennungsdomäne aufweisen. Das R135 am nächsten kommende Homolog ist eine Arylalkohol-Oxidase[20], die am biologischen Ligninabbau von Pflanzenzellwänden beteiligt ist. Somit könnte R135 an der Perforation der Zellwand ihrer natürlichen Wirte, insbesondere ligninhaltiger Algen, beteiligt sein.[15] Unter Laborbedingungen ist aber keines der drei genannten Proteine für die Infektiosität unbedingt erforderlich.[15]
Stargate
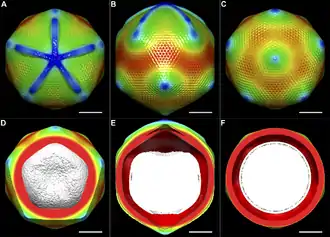
Auffällig ist die fünfeckige, sternförmige Struktur an einer der Ecken des Kapsids, das so genannte ‚Stargate‘ (auch star-gate,[21] englisch für „Sternentor“, deutsch auch ‚Sternstruktur‘ oder ‚Seesternstruktur‘ genannt).[22] Blickt man direkt auf diesen Eckpunkt (den Mittelpunkt des Sterns), so scheinen zwischen dessen Strahlen fünf dreieckige Flächen zu liegen. Die Strahlen haben eine Breite von ungefähr 50 nm, eine Dicke von 40 nm und eine Länge von 200 nm; sie erreichen fast die benachbarten Eckpunkte der ikosaedrischen Kapsids. Das Stargate ist nicht von Fibrillen bedeckt.[15] Das Vorhandensein dieser Struktur verändert die Geometrie des Kapsids, indem seine Geometrie von der idealen Ikosaederform abweicht: Tatsächlich verläuft bei genauer Betrachtung nur eine einzige Achse mit Fünfstrahlsymmetrie durch das Virion, die durch den Mittelpunkt des Sterns (genannt Scheitelpunkt) verläuft.[17][23]
Die Symmetrie des Kapsids wird unterschiedlich angegeben mit einer Triangulationszahl T=972–1141 oder T=1200.[6]
Da auf der Oberfläche der Sternstruktur keine hexagonal geordneten Vertiefungen zu beobachten sind, wird vermutet, dass es sich bei dieser um ein Protein handelt, das sich vom Hauptkapsidprotein unterscheidet.[17]
Das Stargate spielt eine besondere Rolle bei der Infektion der Wirtszelle: Während der Infektion öffnet sich der „Verschluss“ am Scheitelpunkt und es erfolgt die Freisetzung des viralen Kerns (mit DNA und vorgefertigten Proteinen) aus dem Kapsid in das Zytosol der Wirtszelle (per Phagocytose). Das ist der Grund, warum die Sternstruktur als „Sternentor“ (englisch stargate) bezeichnet wird.[24]
Nukleokapsid
Das Mimivirus hat mehrere morphologische Merkmale mit allen Mitgliedern der NCLDV-Virengruppe gemeinsam. Unmittelbar unter dem Kapsid des Mimivirus befinden sich beispielsweise zwei elektronendichte Schichten, die als Membranen gedeutet werden.[19] Unter diesen Membranen befindet sich eine etwa 7 nm dicke Proteinhülle, in der die lineare doppelsträngige DNA des Virus eingeschlossen ist. Dieser kondensierte Zentralkern des Virions erscheint unter dem Elektronenmikroskop als dunkler Bereich, das sogenannte ‚Nukleokapsid‘. In diesem Bereich befindet sich das große Genom des Virus, daneben auch mRNAs und vorgefertigte Proteine. Da alle anderen NCLDVs eine interne Lipidschicht besitzen, die den zentralen Kern umgibt, vermutet man das auch bei Mimivirus. Die Wände des Nukleokapsids liegen etwa 30 nm hinter den Wänden des Kapsids zurück, im Bereich der Sternstruktur (dem Stargate) ist die Oberfläche des Nukleokapsids zusätzlich abgesenkt.[17] Es wird angenommen, dass der Raum zwischen der Spitze der Sternstruktur und dem Nukleokapsid mit hydrolytischen Enzymen gefüllt ist, die für das Eindringen des Virus in die Wirtszelle erforderlich sind. Zwischen dem Kapsid und dem Nukleokapsid wurden interne Proteinstränge entdeckt, die anscheinend die gegenseitige räumliche Positionierung der beiden Teile zueinander stabilisieren.[19]
Genom
APMV hat damit im Vergleich zu den meisten anderen Viren ein großes und komplexes Genom, das aus einem einzelnen linearen DNA-Doppelstrang (dsDNA) besteht. Die Genomlänge des APMV Wildtyps (Ausgangsvariante Mimivirus M1) wurde von Raoult et al. (2004) mit 1.181.404 bp angegeben,[11] dieser Wert wurde durch Bäckström et al. (2019) leicht korrigiert auf 1.181.594 bp.[25] Das entspricht etwa 800 nm. Die fiberlose Variante Mimivirus M4 hat nur 0,993 Mbp, dazwischen liegen M2 mit 1.10 Mbp und M3 mit 1,10 Mbp.[26] Der GC-Gehalt von APMV liegt bei 28 %. Es gibt bei APMV 1260 Offene Leserahmen (ORFs, englisch open reading frames),[11] darunter vorhergesagt 979 kodierende Gene.[25][15][27] Dies geht weit über die Mindestausstattung von 4 Genen hinaus, die für ein Virus erforderlich sind, wie etwa bei den Phagen MS2 und Qβ.[28] Detaillierte Studien zum Genom korrigieren immer wieder Sequenzfehler und entdecken unter Umständen neue Leserahmen.[29]
Der Anteil von nichtcodierender DNA beträgt damit nur etwa 9,5 bis 10 %. Offene Leserahmen sind durch Lücken von ungefähr 157 Nukleotidpaaren getrennt. Zwei DNA-Abschnitte mit der Bezeichnung R (englisch right – rechts) und L (englisch left – links) kodieren ungefähr die gleiche Anzahl von Genen (450 bzw. 465, gemäß Daten von 2010). Der GC-Gehalt ist mit 28 % niedrig. In der Nähe der Enden des DNA-Moleküls wurden ‚Invertierte Wiederholungen‘ (englisch inverted repeats) mit 617 Nukleotidpaaren gefunden. Es wird vermutet, dass die gegenseitige Wechselwirkung dieser Stellen zur Bildung einer Q-Struktur führen kann – zirkuläre DNA mit zwei kleinen Fortsätzen.[29]
Bei der Analyse stieß man auf mindestens 21 Gene mit Homologie zu bekannten Proteinen, darunter solche, die man bis dato von keinem anderen Virus, sondern nur von zellulären Organismen kannte, inklusive Aminoacyl-tRNA-Synthetase.[11][9][30] 43 Gene sind homolog zu solchen anderer Riesenviren (NCLDVs).[15] Wie andere Riesenviren enthält Mimivirus mehrere Gene für den Zucker-, Lipid- und Aminosäurestoffwechsel. Es gab auch Stoffwechselgene, die zuvor in keinem anderen Virus gefunden wurden.[14]
Aus gereinigten Virionen konnten mehrere mRNA-Transkripte gewonnen werden. Wie schon bei anderen NCLDVs wurden insbesondere Transkripte für DNA-Polymerase, ein Kapsidprotein und ein TFII-ähnlicher Transkriptionsfaktor gefunden werden. Es wurden jedoch auch drei verschiedene Aminoacyl-tRNA-Synthetase-Transkripte und vier unbekannte mRNA-Moleküle gefunden, die für das Mimivirus spezifisch sind. Diese vorverpackten Transkripte können ohne virale Genexpression translatiert werden und sind wahrscheinlich für die Replikation von Mimivirus erforderlich. Andere DNA-Viren, wie das Humane Cytomegalievirus und das Herpes-simplex-Virus Typ 1, enthalten ebenfalls gepackte mRNA-Transkripte.[14]
Mimivirus ist eines der wenigen dsDNA-Viren, in deren Genom eine Intein-kodierende Sequenz nachgewiesen wurde. Inteine sind Proteindomäne, die ihre eigene Entfernung von einem Trägermolekül und die anschließende Verknüpfung der gebildeten Enden katalysieren. Eine solche Sequenz ist im Mimivirus-Gen für DNA-Polymerase B vorhanden.[31]
Auf Grund der außergewöhnlich komplexen genetischen Ausstattung des Virus stellt sich für einige Forscher die Frage neu, wo die Grenze zwischen belebter und unbelebter Natur verlaufe, also wie „Lebewesen“ zu definieren ist.
Vermehrungszyklus
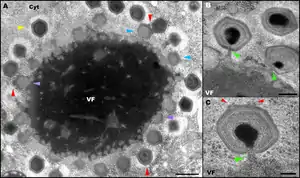
A) TEM-Aufnahme einer intrazellulären Virusfabrik, mit Viruspartikeln (Virionen) von Mimivirus in verschiedenen Montagestadien
- VF: Virusfabrik (Viroplasma)
- Cyt: Cytoplasma
- violette Pfeile: leere, noch faserlose Virionen im Anfangsstadium der Zusammensetzung, in unmittelbarer Nähe zur Peripherie der Virusfabrik auftretend
- blaue Pfeile: Teilweise montierte leere, faserlose Virionen
- gelbe Pfeile: reife, faserbedeckte Virionen, die sich jetzt weiter von der Virusfabrik entfernt befinden als die unreifen Partikel
- rote Pfeile: bei mehreren Viruspartikeln ist ein ‚Stargate’, das sich regelmäßig an der distalen Stelle der Fabrik befindet, zu erkennen
B) und C) TEM-Aufnahme der Mimivirus-Partikel
- grüne Pfeile: Mimivirus-Partikel im Stadium des ‚DNA-Packaging’ (DNA-Verpackung)
- rote Pfeile: zwei Kanten eines Stargates gegenüber der Stelle der DNA-Verpackung
Die Einzelheiten und die verschiedenen Stadien im Replikationszyklus von Mimivirus, wie die offensichtliche Bindung an die Zelloberfläche und den Eintritt in die Zelle, die Freisetzung des Viruskerns, die DNA-Replikation, die Transkription, die Translation, und schließlich den Zusammenbau und die Freisetzung von Tochter-Virionen, sind noch nicht ausreichend bekannt. Die Wissenschaftler haben jedoch den oben angegebenen allgemeinen Überblick anhand elektronenmikroskopischer Aufnahmen infizierter Zellen erstellt. Alle Stadien des Vermehrungszyklus verlaufen im Cytoplasma der Wirtszelle.[32]
Die Infektion der Amöbe mit einem Mimivirus erfolgt vermutlich nach folgendem Szenario:
- Die Mimivirus-Virionen ähneln in ihrer Größe und dem Vorhandensein charakteristischer Polysaccharide auf der Oberfläche Bakterien (siehe Gram-Färbung, Name). Sie werden daher von der Amöbe als Nahrung während eines Endozytoseprozesses absorbiert. Die Polysaccharide fungieren dabei als chemischer Rezeptor und leiten die Anlagerung ein. Als Ergebnis der Endozytose befinden sich die Virionen in Endosomen innerhalb der Zelle.
- Die Proteinfilamente werden in den Endosomen teilweise lysiert, wodurch das Kapsid mit der Endosomenmembran in Wechselwirkung treten kann.
- ~2 Stunden nach der Infektion: Das Kapsid öffnet sich im Bereich der Sternstruktur (Stargate), die innere Membran fusioniert mit der Endosomenmembran und der Inhalt des Kapsids wird in das Cytoplasma freigesetzt.
- Nachdem das Kernteilchen (der innere Teil des Nukleokapsids) ins Cytoplasma ausgetreten ist, beginnt aufgrund der Anwesenheit des viralen Transkriptionsapparats die Synthese der viralen mRNA. Diese mRNAs reichern sich im Inneren des Kernpartikels in Form von Granula an.[32] Äußerlich betrachtet scheint das Virus verschwunden und alles in der Zelle sieht normal aus (Dunkelphase, englisch eclipse phase).
- 4–5 Stunden nach der Infektion: Die virale DNA verlässt das Kernteilchen und wird entpackt, so dass die Replikation beginnen kann. Infolgedessen entsteht neben der leeren Hülle des Kernpartikels eine sogenannte „Virusfabrik“ – ein Ort für die Synthese der einzelnen Komponenten der Virionen und ihren anschließenden Zusammenbau. Wenn mehrere Viruspartikel in die Zelle gelangt sind, verschmelzen die von ihnen gebildeten „Fabriken“ beim Wachstum zu einer einzigen. Man erkennt jetzt kleine Ansammlungen in einigen Bereichen der Zelle.
- 6–9 Stunden nach der Infektion: Zusammensetzung (Assemblierung) der Kapside mit gleichzeitiger DNA-Packung an der Peripherie der „Virusfabriken“. Eine ungewöhnliche Eigenschaft des Mimivirus ist, dass DNA gepackt ist.[24] Die Mimivirus-Virionen werden in der Zelle deutlich sichtbar.
- 14–24 Stunden nach der Infektion: Die Amöbenzellen werden lysiert, d. h. sie platzen auf und die Virionen werden freigesetzt. Pro Wirtszelle werden so mehr als 300 Einheiten erzeugt.[29][14]
Mögliche Pathogenität
Es wurde spekuliert, dass Mimivirus Erreger bestimmter Formen von Lungenentzündung (Pneumonia) sein könnte. Dies beruht hauptsächlich auf indirekten Nachweisen in Form von Antikörpern gegen das bei Lungenentzündungspatienten entdeckte Virus.[33] Aufgrund der wenigen bisherigen Veröffentlichungen ist die Einstufung des Mimivirus als möglicher Krankheitserreger derzeit jedoch schwierig. Ein großer Teil der Fälle von Lungenentzündung verläuft ohne feststellbare Ursache.[34] Zwar wurde ein Mimivirus bei einer an Lungenentzündung leidenden Tunesierin isoliert,[35] und es gibt aus Zellkulturen Hinweise darauf, dass Mimivirus Makrophagen infizieren kann und darin repliziert wird.[36] So wurde unter experimentellen Bedingungen beobachtet, dass Mimivirus humane Makrophagen infizieren kann, d. h. via Phagocytose in die Zellen eindringen, und sich dort replizieren kann.[37][36] Außerdem wurden in mehreren Studien bei einer kleinen Anzahl von Patienten mit Lungenentzündung Antikörper gegen das Mimivirus gefunden.[33][38] Es wurde auch ein Einzelfall einer Lungenentzündung eines Laborassistenten beschrieben, der mit Kulturen dieses Virus arbeitete. Der Gehalt an Antikörpern gegen Mimivirus in seinem Blut war ebenfalls erhöht.[39] Das Vorhandensein von Antikörpern gegen das Virus an sich ist jedoch kein Hinweis auf seine Pathogenität. Es ist möglich, dass das Mimivirus einfach starke immunogene Eigenschaften aufweist, d. h. eine deutliche Immunantwort auslöst.[29] Auch war es in keinem der registrierten Fälle möglich, das Virus in seiner reinen Form aus Proben von Flüssigkeiten zu isolieren, die von Patienten erhalten wurden.[40]
Die Fiberproteine R135 und L829 wurden als Hauptantigene des Mimivirus identifiziert: Die faserlose Variante Mimivirus M4 zeigte jedoch keine Reaktivität mit Seren von menschlichen Patienten, was bestätigt, dass diese Proteine in M4 fehlen.[15]
Resistenzeigenschaft
Das Zamilon-Virus ist ein Satellitenvirus, das Mimiviren der Linien B und C befällt, nicht aber die Mimiviren der Linie A. Diese weisen nämlich eine MIMIVIRE, englisch mimivirus virophage resistance element, genannte Resistenz auf, die ähnlich funktioniert wie das CRISPR/Cas-System.[41][42]
Im Übrigen wurde nachgewiesen, dass es nicht nur einen Gentransfer zwischen den amöboid Wirten und Riesenviren als intrazellulären viralen Endocytobionten (Organismen, die in den Zellen anderer Organismen leben oder sich vermehren) gibt[43] sondern sogar zwischen den Viren und gleichzeitig vorhandenen bakteriellen Endocytobionten.[44][26][45]
Merkmale der Mimiviridae im Vergleich
| Virus | Aminoacyl-tRNA-Synthetase | Octocorallia-ähnliche MutS | Proteinfilamente (Länge) | Stargate[24] | Bekannter Virophage[46] | Cytoplasmische Virion-Fabrik |
Wirt |
|---|---|---|---|---|---|---|---|
| Megavirus chilensis | 7 (Tyr, Arg, Met, Cys, Trp, Asn, Ile) | ja | ja (75 nm) | ja | nein | ja | Acanthamöben (Unikonta, Amoebozoa) |
| Mamavirus ACMV | 4 (Tyr, Arg, Met, Cys) | ja | ja (120 nm) | ja | ja | ja | Acanthamöben (Unikonta, Amoebozoa) |
| Mimivirus ApMV (Wildtyp M1) | 4 (Tyr, Arg, Met, Cys) | ja | ja (120 nm) | ja | ja | ja | Acanthamöben (Unikonta, Amoebozoa) |
| Mimivirus M4 (bald/fiberless Variante) | 2 (Met, Cys) | nein | nein | ja | resistent | ja | Acanthamöben (Unikonta, Amoebozoa) |
| Cafeteria-roenbergensis-Virus | 1 (Ile) | ja | nein | nein | ja | ja | Phagotrophische Protozoen (Heterokonta, Stramenopiles) |
Systematik
Äußere Systematik
Die Gattung Mimivirus und einige andere – vom ICTV mit Stand März 2019 noch nicht bestätigte – genetisch ähnliche Gattungen und Spezies der Familie Mimiviridae (wie zum Beispiel „Mamavirus“, „Megavirus“ und „Moumouvirus“) bilden eine als Gruppe I bezeichnete Klade der Mimiviren im weiteren Sinn. Für diese wurde vorgeschlagen, sie als „Megamimivirinae“,[5] „Megavirinae“[4] oder auch „Mimivirinae“[3] in den Rang einer Unterfamilie zu erheben und so von anderen ebenfalls vorgeschlagenen Unterfamilien der Mimiviridae abzugrenzen, nämlich:
- Cafeteriavirus-Gruppe (Gruppe II)
- Klosneuvirus-Gruppe – als mögliche Unterfamilie „Klosneuvirinae“
Innerhalb der Mimiviridae-Gruppe I zeichnen sich die folgenden Linien ab:[4]
- Linie A: Mimivirus-Gruppe: Mimiviren im engeren Sinn
- Linie B: Moumouvirus-Gruppe (Moumouviren)
- Linie C: Courdo11-Gruppe mit „Megavirus chilensis“ (Spezies, dazu Courdo11-Virus)
- Tupanvirus-Gruppe (Tupanviren)
Da bisher (März 2019) vom ICTV lediglich die Gattung Mimivirus bestätigt wurde, bleibt zurzeit noch offen, welche Kandidaten für diese vorgeschlagene Unterfamilie dieser Gattung einmal zugeordnet werden, und für welche eigene Gattungen einzurichten sind. Sicher scheint aber, dass die Vertreter der Linie A in jedem Fall zur Gattung Mimivirus gehören.
Der von Wilson et al. 2017 beschriebene Kandidat „gvSAG AB-566-O17“ (vom NCBI als Spezies „Mimivirus AB-566-O17“ bezeichnet[47]) ist nach den Autoren (Fig. 2) mit APMV weitläufiger verwandt als CroV (aber näher als Vertreter der OLPG-Gruppe). Er ist daher keiner der obigen Gruppen zuzuordnen (insbesondere nicht der Gattung Mimivirus).[48]
Innere Systematik
Systematik der Linie A (Mimiviren im engeren Sinn):
- Gattung Mimivirus
- Spezies: Acanthamoeba polyphaga mimivirus (APMV)[49] (vom ICTV mit Stand März 2018 bestätigt) – Fundort: Bradford, England, UK[1] (informell Mimivirus s. s.[50])
- Mimivirus M1 (englisch wild type)[26] – Wildtyp (natürliche Ursprungsvariante) aus Bradford
- Mimivirus M1 und M2 – Zwischenformen zwischen M1 und M4 mit kürzeren Fibrillen als der Wildtyp[26]
- Mimivirus M4 (englisch bald/fiberless variant) – fiberlose Variante, wird nicht vom Sputnik-Virophagen befallen[15][26]
- Mimivirus Bombay (MVB, alias Bombay-Virus)[49][1][51][52][53][54] – Fundort: Mumbai, Indien
- Mimivirus shirakomae (alias Shirakomae-Virus)[53] – Fundort: Shirakoma Pond, Nagano, Japan[1][55]
- Mimivirus kasaii (alias Kasaii-Virus) – Fundort: Arakawa (Fluss), Tokio, Japan[1][55][53]
- Samba-Virus[49][56][51] – Fundort: Rio Negro, Brasilien[1][57][58][59]
- Mimivirus amazonia (alias Amazonia virus, Amazonian virus)[1][60][51] – Fundort: Rio Negro, Brasilien[1]
- Oyster virus – Fundort: Florianópolis, Brasilien[1][60]
- Kroon-Virus[61][60] – Fundort: Lagoa Santa, Brasilien[1]
- Das NCBI führt unter APMV noch das Saudi moumouvirus (SDMV) an,[62] verweist aber selbst als Referenz die Arbeit von Bajrai et al. (2016), die im Titel Saudi Moumouvirus, the First Group B Mimivirus Isolated from Asia[63] korrekt auf Gruppe d. h. Linie B (Moumouvirus-Gruppe) verweist.
- Spezies: „Acanthamoeba castellanii mamavirus“ (ACMV, alias Acanthamoeba castellanii mimivirus)[64][65][66] – Fundort: Paris, Frankreich[1]
- Spezies: „Hirudovirus“[67][68] – Fundort: Tunesien[1]
- Hirudovirus sangsue[53]
- Spezies: Acanthamoeba polyphaga mimivirus (APMV)[49] (vom ICTV mit Stand März 2018 bestätigt) – Fundort: Bradford, England, UK[1] (informell Mimivirus s. s.[50])
_Niemeyer_virus.jpg.webp)
.png.webp)
.png.webp)
- Spezies: „Niemeyer-Virus“ (NYMV)[71][69] – Fundort: Pampulha-See, Belo Horizonte, Brasilien (benannt zu Ehren des Architekten Oscar Niemeyer)[1][72]
- Spezies: „Mimivirus battle6“[73][51]
- Spezies: „Mimivirus battle7“[74][51]
- Spezies: „Mimivirus battle19“[75][51]
- Spezies: „Mimivirus battle27“[76][51]
- Spezies: „Mimivirus battle57“[77][51]
- Spezies: „Mimivirus battle66“[78][51]
- Spezies: „Mimivirus battle83“[79][51]
- Spezies: „Mimivirus battle86“[80][81]
- Spezies: „Mimivirus Cher“[82][81]
- Spezies: „Mimivirus dakar4“[83][51]
- Spezies: Mimivirus fauteuil (alias „Fauteuil-Virus“)[84][85][1][51] – vgl. Fauteuil virus FD[86]
- Spezies: „Mimivirus huitre A06“[87][51]
- Spezies: „Mimivirus lactour“ (alias „Mimivirus lactours“,[51] Lactours-Virus)[88][85] – vgl. Lactours virus LT2[89]
- Spezies: „Mimivirus lentille“ (alias „Lentille-Virus“, „Acanthamoeba polyphaga lentillevirus“)[90][85][51][66] – vgl. Lentille virus CL[91]
- Spezies: „Mimivirus longchamps“ (alias „Longchamps-Virus“)[92][85][1][51] – vgl. Longchamps virus FPL[93]
- Spezies: „Mimivirus marais“ (alias „Marais-Virus“)[94][85][51]
- Spezies: „Mimivirus montadette2“[95][81]
- Spezies: „Mimivirus pointerouge1“[96] – vgl. Pointerouge virus 1 (alias Pointe-Rouge 1 virus)[97][1][51] – Fundort: Marseille, Frankreich[1]
- Spezies: „Mimivirus pointerouge2“[98] – vgl. Pointerouge virus 2 (alias Pointe-Rouge 2 virus)[99][1][51] – Fundort: Marseille, Frankreich[1]
- Spezies: Mimivirus SR1 (alias „Mimivirus-like virus SR1“)[100] – Fundort: Serendah village, Malaiische Halbinsel – Wasserfall[65]
- Spezies: Mimivirus SR4 (alias „Mimivirus-like virus SR4“)[101] – Fundort: Serendah village, Malaiische Halbinsel – Mitte zw. Wasserfall und Dorf[65]
- Spezies: „Mimivirus SR9“ (alias „Mimivirus-like virus SR9“)[102] – Fundort: Serendah village, Malaiische Halbinsel – Mündung des Zuflusses in den See[65]
- Spezies: „Mimivirus T2“[103][51]
- Spezies: „Mimivirus T3“[104][51]
- Spezies: „Mimivirus terra2“ (alias Terra2-Virus)[105][85][51] – Fundort: Marseille, Frankreich[1]
- Terravirus2 TAO-TJA[81]
- Spezies: „Mimivirus univirus“[106][81]
Kladogramm
Vorgeschlagener phylogenetischer Baum der Gattung Mimivirus nach Abrahão et al. (2018), Fig. 4:[107]
| Mimivirus |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Anmerkung: Kursivschrift für vorgeschlagene Spezies.
Anmerkungen
- Das Material wurde von dieser Quelle kopiert, die unter einer Creative Commons Attribution 4.0 International License verfügbar ist.
Literatur
- Stefanie Reinberger: Revolution der Riesenviren. In: Spektrum der Wissenschaft. Spektrum der Wissenschaft Verlagsgesellschaft, Heidelberg 2012, S. 14–16.
- Graziele Oliveira, Bernard La Scola, Jônatas Abrahão: Giant virus vs amoeba: fight for supremacy. In: Virol J, 16, 126, 4. November 2019, doi:10.1186/s12985-019-1244-3, researchgate.net (PDF)
- Ana Cláudia dos S. P. Andrade, Thalita S. Arantes, Rodrigo A. L. Rodrigues, Talita B. Machado, Fábio P. Dornas, Melissa F. Landell, Cinthia Furst, Luiz G. A. Borges, Lara A. L. Dutra, Gabriel Almeida, Giliane de S. Trindade, Ivan Bergier, Walter Abrahão, Iara A. Borges, Juliana R. Cortines, Danilo B. de Oliveira, Erna G. Kroon, Jônatas S. Abrahão: Ubiquitous giants: a plethora of giant viruses found in Brazil and Antarctica. In: Virology Journal, Band 15, Nr. 22, 24. Januar 2018, doi:10.1186/s12985-018-0930-x
- Philippe Colson, Bernard La Scola, Anthony Levasseur, Gustavo Caetano-Anollés,& Didier Raoult: Mimivirus: leading the way in the discovery of giant viruses of amoebae. In: Nature Reviews Microbiology, Band 15, S. 243–254, 27. Februar 2017, doi:10.1038/nrmicro.2016.197
Weblinks
- Jean-Michel Claverie (Hrsg.): Bilder des Mimivirus. Structural & Genomic Information Laboratory CNRS UPR, Marseille
- Referenzsequenz NC_006450. NCBI
- David R. Wessner: Discovery of the Giant Mimivirus. Nature Masterclasses
- Mimivirus – an overview, auf: ScienceDirect, 2012
- Scinexx: Riesenviren an der Grenze zum Leben, vom 28. Februar 2018
- Andreas Jahn: Das Virus-Virus, auf: Spektrum.de vom 6. August 2008
- Biologie Seite: Mimivirus
- Iddo: Size matters. Life is live., auf: Byte Size Biology, Blog vom 1. Mai 2009, mit Bild vom sich öffnenden Stargate, identisch mit Zaubermann et al. (2008), Fig. 5.[24]
- Adrian De Novato: New study shines light on mysterious giant viruses. phys.org, 8. Mai 2020 (die Abbildung zeigt offenbar einen typischen Vertreter der Gattung Mimivirus mit Stargate (Öffnung für DNA) und Tegument (Hülle aus Fibrillen).
- Nicoletta Lanese: Giant viruses spew their DNA through a ‘stargate’. Now, scientists know what triggers them. LiveScience, 26. Mai 2020
Einzelnachweise
- Jan Diesend, Janis Kruse, Monica Hagedorn, Christian Hammann: Amoebae, Giant Viruses, and Virophages Make Up a Complex, Multilayered Threesome, in: Frontiers in Cellular and Infection Microbiology 7, Januar 2018, doi:10.3389/fcimb.2017.00527, via ResearchGate, Fig. 1 (NCLDVs und ‚Megavirales‘ werden in dieser Arbeit nicht ganz korrekt als ‚Familien‘ bezeichnet, gemeint sind ‚Gruppen‘).
- ICTV: ICTV Taxonomy history: Acanthamoeba polyphaga mimivirus, EC 51, Berlin, Germany, July 2019; Email ratification March 2020 (MSL #35)
- Christoph M. Deeg, Cheryl-Emiliane T. Chow, Curtis A. Suttle: The kinetoplastid-infecting Bodo saltans virus (BsV), a window into the most abundant giant viruses in the sea…, in: eLife Sciences 7, März 2018, doi:10.7554/eLife.33014
- Centre national de la recherche scientifique: List of the main “giant” viruses known as of today (March 2019) (PDF) Université Aix Marseille, März 2019.
- Frederik Schulz, Lauren Alteio, Danielle Goudeau, Elizabeth M. Ryan, Feiqiao B. Yu, Rex R. Malmstrom, Jeffrey Blanchard, Tanja Woyke: Hidden diversity of soil giant viruses. In: Nature Communications, volume 9, Article number: 4881 (2018), 19. November 2018, doi:10.1038/s41467-018-07335-2
- ViralZone: Mimivirus. ExPASy. Abgerufen am 8. Juli 2019.
- ICTV: Master Species List 2018b.v2. Abgerufen am 6. August 2019. MSL #34v
- Bernard La Scola, S. Audic, C. Robert, L. Jungang, X. de Lamballerie, M. Drancourt, R. Birtles, J. M. Claverie, D. Raoult: A giant virus in amoebae. In: Science, 299, 2003, S. 2033. PMID 12663918
- Laurie O'Keefe: Sizing Up Viruses. (PDF; 103 kB) The Scientist. Illustration zu Didier Raoult: Viruses Reconsidered, ebenda vom 28. Februar 2014
- Didier Raoult, S. Audic, C. Robert, C. Abergel, P. Renesto, H. Ogata, B. La Scola, M. Suzan, J. M. Claverie: The 1.2-megabase genome sequence of Mimivirus. In: Science, 306(5700), 19. November 2004, S. 1344–1350, doi:10.1126/science.1101485, PMID 15486256
- Discovery and Further Studies on Giant Viruses at the IHU Mediterranee Infection That Modified the Perception of the Virosphere. In: Viruses, 11(4), März/April 2019, pii: E312, doi:10.3390/v11040312, PMC 6520786 (freier Volltext), PMID 30935049
- H. Pearson: 'Virophage' suggests viruses are alive. In: Nature. 454, Nr. 7205, 2008, ISSN 0028-0836, S. 677–677. bibcode:2008Natur.454..677P. doi:10.1038/454677a.
- M Suzan-Monti, B La Scola, D Raoult: Genomic and evolutionary aspects of Mimivirus. In: Virus Research. 117, Nr. 1, April 2006, S. 145–155. doi:10.1016/j.virusres.2005.07.011. PMID 16181700.
- Thomas Klose, Dominik A. Herbst, Hanyu Zhu, Joann P. Max, Hilkka I. Kenttämaa, Michael G. Rossmann: A Mimivirus Enzyme that Participates in Viral Entry, in: Structure Band 23, Nr. 6, 2. Juni 2015, S. 1058–1065, doi:10.1016/j.str.2015.03.023
- ICTV: Mimiviridae - Figures. In: ICTV 9th Report (2011).
- Xiao C., Kuznetsov Y. G., Sun S., Hafenstein S. L., Kostyuchenko V. A., Chipman P. R., Suzan-Monti M., Raoult D., McPherson A., Rossmann M. G.: Structural studies of the giant mimivirus. In: PLoS Biol. 7, Nr. 4, 2009, S. e92. doi:10.1371/journal.pbio.1000092. PMID 19402750. PMC 2671561 (freier Volltext).
- Jean-Michel Claverie, Chantal Abergel, Hiroyuki Ogata: Mimivirus. In: Curr Top Microbiol Immunol. 328, 2009, S. 89–121. doi:10.1007/978-3-540-68618-7_3. PMID 19216436.
- Klose Thomas, Kuznetsov Y. G., Xiao C., Sun S., McPherson A., Rossmann M. G.: The three-dimensional structure of Mimivirus. In: Intervirology. 53, Nr. 5, 2010, S. 268–273. doi:10.1159/000312911. PMID 20551678. PMC 2895761 (freier Volltext).
- siehe auch Vanillylalkohol-Oxidase
- Chuan Xiao, Matthias G. Fischer, Duer M. Bolotaulo, Nancy Ulloa-Rondeau, Gustavo A. Avila & Curtis A. Suttle: Cryo-EM reconstruction of the Cafeteria roenbergensis virus capsid suggests novel assembly pathway for giant viruses. In: Nature Scientific Reports, Band 7, Nr. 5484, 14. Juli 2017, doi:10.1038/s41598-017-05824-w.
- James L. vaan Etten: Giant Viruses. aud: American Scientist, Band 89, Nr. 4, Juli/August 2011. Siehe insbes. Fig. 3.
- In der Physik ist dieses geometrische Prinzip als Symmetriebrechung (Brechung einer diskreten Symmetrie) bekannt.
- Autor=Nathan Zauberman, Y. Mutsafi, D. B. Halevy, E. Shimoni, E. Klein, C. Xiao, S. Sun, A. Minsky: Distinct DNA exit and packaging portals in the virus Acanthamoeba polyphaga mimivirus. In: PLoS Biol. 6, Nr. 5, 2008, S. e114. doi:10.1371/journal.pbio.0060114. PMID 18479185. PMC 2430901 (freier Volltext).
- Disa Bäckström, Natalya Yutin, Steffen L. Jørgensen, Jennah Dharamshi, Felix Homa, Katarzyna Zaremba-Niedwiedzka, Anja Spang, Yuri I. Wolf, Eugene V. Koonin, Thijs J. G. Ettema; Richard P. Novick (Hrsg.): Virus Genomes from Deep Sea Sediments Expand the Ocean Megavirome and Support Independent Origins of Viral Gigantism, in: mBio Vol. 10, Nr. 2, März–April 2019, S. e02497-18, PDF (PDF) doi:10.1128/mBio.02497-18, PMC 6401483 (freier Volltext), PMID 30837339, ResearchGate
- Mickaël Boyer, Saïd Azza, Lina Barrassi, Thomas Klose, A. Campocasso, I. Pagnier, G. Fournous, A. Borg, C. Robert, X. Zhang, C. Desnues, B. Henrissat, M. G. Rossmann, B. La Scola, D. Raoult: Mimivirus shows dramatic genome reduction after intraamoebal culture, in: Proc Natl Acad Sci (PNAS) USA 108(25), 21. Juni 2011, S. 10296–10301, doi:10.1073/pnas.1101118108, PMID 21646533, PMC 3121840 (freier Volltext)
- David M. Needham, Alexandra Z. Worden et al.: A distinct lineage of giant viruses brings a rhodopsin photosystem to unicellular marine predators, in: PNAS, 23. September 2019, doi:10.1073/pnas.1907517116, ISSN 0027-8424, hier: Supplement 1 (xlsx)
- Lansing M. Prescott: 2. (Hrsg.): Microbiology. Wm. C. Brown Publishers, Dubuque, IA 1993, ISBN 0-697-01372-3.
- Jean-Michel Claverie, Chantal Abergel: Mimivirus and its virophage. In: Annu Rev Genet. 43, Nr. 49–66, 2009. doi:10.1146/annurev-genet-102108-134255. PMID 19653859.
- Jean-Michel Claverie, Hiroyuki Ogata, Stéphane Audic, Chantal Abergel, Pierre-Edouard Fournier, Karsten Suhre: Mimivirus and the emerging concept of 'giant’ virus. In: Virus Research. 117, Nr. 1, 2006, S. 133–144. arxiv:q-bio/0506007. doi:10.1016/j.virusres.2006.01.008. PMID 16469402.
- Hiroyuki Ogata, Didier Raoult, Jean-Michel Claverie: A new example of viral intein in Mimivirus. In: Virol J. 2, Nr. 8, 2005. PMID 15707490. PMC 549080 (freier Volltext).
- Mutsafi Y., Zauberman N., Sabanay I., Minsky A.: Vaccinia-like cytoplasmic replication of the giant Mimivirus. In: Proc Natl Acad Sci USA (PNAS). 107, Nr. 13, 2010, S. 5978–5982. doi:10.1073/pnas.0912737107. PMID 20231474. PMC 2851855 (freier Volltext).
- B. La Scola, T. J. Marrie, J. P. Auffray, D. Raoult: Mimivirus in pneumonia patients. In: Emerg Infect Dis.. 11, Nr. 3, 2005, S. 449–452. PMID 1575756 3. PMC 3298252 (freier Volltext).
- Marrie TJ, Durant H, Yates L: Community-Acquired Pneumonia Requiring Hospitalization: 5-Year Prospective Study. In: Reviews of Infectious Diseases. 11, Nr. 4, 1989, S. 586–99. doi:10.1093/clinids/11.4.586. PMID 2772465.
- Hanene Saadi, Isabelle Pagnier, Philippe Colson, Jouda Kanoun Cherif, Majed Beji, Mondher Boughalmi, Saïd Azza, Nicholas Armstrong, Catherine Robert, Ghislain Fournous, Bernard La Scola, Didier Raoult: First isolation of Mimivirus in a patient with pneumonia. In: Clinical Infectious Diseases. 57, Nr. 4, August 2013, S. e127–34. doi:10.1093/cid/cit354. PMID 23709652.
- E. Ghigo, J. Kartenbeck, P. Lien, L. Pelkmans, C. Capo, J. L. Mege, D. Raoult: Ameobal pathogen mimivirus infects macrophages through phagocytosis. In: PLoS pathogens. Band 4, Nummer 6, Juni 2008, S. e1000087, doi:10.1371/journal.ppat.1000087, PMID 18551172, PMC 2398789 (freier Volltext).
- Vincent A., La Scola B., Papazian L.: Advances in Mimivirus pathogenicity. In: Intervirology. 53, Nr. 5, 2010, S. 304–309. doi:10.1159/000312915. PMID 20551682.
- P. Berger, L. Papazian, M. Drancourt, B. La Scola, J. P. Auffray, D. Raoult: Ameba-associated microorganisms and diagnosis of nosocomial pneumonia. In: Emerg Infect Dis. 12, Nr. 2, 2006, S. 248–255. PMID 16494750. PMC 3373093 (freier Volltext).
- D. Raoult, P. Renesto, P. Brouqui: Laboratory infection of a technician by mimivirus. In: Ann Intern Med. 144, Nr. 9, 2006, S. 702–703. PMID 16670147.
- M. J. Vanspauwen et al.: Infections with mimivirus in patients with chronic obstructive pulmonary disease. In: Respiratory Medicine. 106, Nr. 12, 2012, S. 1690–1694. doi:10.1016/j.rmed.2012.08.019.
- Anthony Levasseur, Meriem Bekliz, Eric Chabrière, Pierre Pontarotti, Bernard La Scola, Didier Raoult: MIMIVIRE is a defence system in mimivirus that confers resistance to virophage. In: Nature, 2016, doi:10.1038/nature17146.
- Ewen Callaway: CRISPR-like 'immune' system discovered in giant virus, in: Nature:News, vom 29. Juni 2016
- Nisrine Chelkha, Anthony Levasseur, Pierre Pontarotti, Didier Raoult et al.: A Phylogenomic Study of Acanthamoeba polyphaga Draft Genome Sequences Suggests Genetic Exchanges With Giant Viruses, in: Frontiers in Microbiology 9:2098, September 2018, doi:10.3389/fmicb.2018.02098
- Patrick L. Scheid: Free-Living Amoebae and Their Multiple Impacts on Environmental Health, in: Reference Module in Earth Systems and Environmental Sciences, 27. Februar 2018, doi:10.1016/B978-0-12-409548-9.10969-8, hier: Text im Anschluss an Fig. 8 (rechte Spalte)
- Eine Homologie zwischen CRISPR und MIMIVIRE scheint damit nicht unmöglich, bedarf aber sicher weiterer Untersuchungen.
- M. G. Fischer, C. A. Suttle: A Virophage at the Origin of Large DNA Transposons. In: Science. Band 332, Nr. 6026, 2011, S. 231–234, doi:10.1126/science.1199412, PMID 21385722.
- NCBI: Mimivirus AB-566-O17 (Acronym: gvSAG AB-566-O17) (Species)
- William H Wilson, Ilana C Gilg, Mohammad Moniruzzaman, Erin K Field, Sergey Koren, Gary R LeCleir, Joaquín Martínez Martínez, Nicole J Poulton, Brandon K Swan, Ramunas Stepanauskas, Steven W Wilhelm: Genomic exploration of individual giant ocean viruses. In: ISME Journal, 11(8), August 2017, S. 1736–1745, doi:10.1038/ismej.2017.61, PMC 5520044 (freier Volltext), PMID 28498373
- NCBI: Acanthamoeba polyphaga mimivirus (Spezies)
- Clara Rolland, Julien Andreani, Amina Cherif Louazani, Sarah Aherfi, Rania Francis, Rodrigo Rodrigues, Ludmila Santos Silva, Dehia Sahmi, Said Mougari, Nisrine Chelkha, Meriem Bekliz, Lorena Silva, Felipe Assis, Fábio Dornas, Jacques Yaacoub Bou Khalil, Isabelle Pagnier, Christelle Desnues, Anthony Levasseur, Philippe Colson, Jônatas Abrahão, Bernard La Scola: Discovery and Further Studies on Giant Viruses at the IHU Mediterranee Infection That Modified the Perception of the Virosphere. In: Viruses, 11(4), März/April 2019, pii: E312, doi:10.3390/v11040312, PMC 6520786 (freier Volltext), PMID 30935049
- Didier Raoult, Anthony Levasseur, Bernard La Scola: PCR Detection of Mimivirus, in: Emerging Infectious Diseases, Juni 2017, Band 23, Nr. 6, S. 1044–1045, doi:10.3201/eid2306.161896, PDF (PDF)
- Hansika Chhabra: Giant viruses found in water samples from Mumbai, in: BusinessLine: Science, Bangalore, 9. Mai 2019
- Sailen Barik: A Family of Novel Cyclophilins, Conserved in the Mimivirus Genus of the Giant DNA Viruses, in: Computational and Structural Biotechnology Journal, Band 16, Juli 2018, S. 231–236, doi:10.1016/j.csbj.2018.07.001
- Anirvan Chatterjee, Thomas Sicheritz-Pontén, Rajesh Yadav, Kiran Kondabagil: Genomic and metagenomic signatures of giant viruses are ubiquitous in water samples from sewage, inland lake, waste water treatment plant, and municipal water supply in Mumbai, India, in: Scientific Reports, Band 9, Nr. 3690, 6. März 2019, doi:10.1038/s41598-019-40171-y, PMID 30842490, PMC 6403294 (freier Volltext)
- Masaharu Takemura, Tatsuya Mikami, Shingo Murono: Nearly Complete Genome Sequences of Two Mimivirus Strains Isolated from a Japanese Freshwater Pond and River Mouth, in: Genome Announc. 4(6), November/Dezember 2016, e01378-16, doi:10.1128/genomeA.01378-16, PMC 5146454 (freier Volltext), PMID 27932662. Die beiden Viruslinien sind nac Angaben der Autoren eng mit Mimivirus Bombay verwandt, gehören also ebenfalls zur Spezies APMV.
- Rafael K Campos, Paulo V Boratto, Felipe L Assis, Eric RGR Aguiar, Lorena CF Silva, Jonas D Albarnaz, Fabio P Dornas, Giliane S Trindade, Paulo P Ferreira, João T Marques, Catherine Robert, Didier Raoult, Erna G Kroon, Bernard La Scola, Jônatas S Abrahão: Samba virus: a novel mimivirus from a giant rain forest, the Brazilian Amazon, in: Virology Journal 2014 11:95, doi:10.1186/1743-422X-11-95
- Jason R. Schrad, Jônatas S. Abrahão, Juliana R. Cortines and Kristin N. Parent: Structural and Proteomic Characterization of the Initiation of Giant Virus Infection (PDF) in: Cell vom 8. Mai 2020, doi:10.1016/j.cell.2020.04.032
- Jason R. Schrad, Jônatas S. Abrahão, Juliana R. Cortines, Kristin N. Parent: Boiling Acid Mimics Intracellular Giant Virus Genome Release (PDF) auf: bioRxiv vom 20. September 2019, doi:10.1101/777854 (Preprint)
- Mysterious Giant Viruses: Gargantuan in Size and Complexity, auf: SciTechDaily vom 9. Mai 2020, Quelle: Michigan State University
- Felipe L. Assis, Leena Bajrai, Jonatas S. Abrahao, Erna G. Kroon, Fabio P. Dornas, Kétyllen R. Andrade, Paulo V. M. Boratto, Mariana R. Pilotto, Catherine Robert, Samia Benamar, Bernard La Scola, Philippe Colson: Pan-Genome Analysis of Brazilian Lineage A Amoebal Mimiviruses, in: Viruses 7(7), 2015, S. 3483–3499, doi:10.3390/v7072782. Die hier abgehandelten Kandidaten stellen nach den Autoren eine Verwandtschaftsgruppe dar, wobei aus den anderen Quellen hervorgeht, dass Samba- und Kroon-Virus zur Spezies APMV gehören. Das gilt dann für die beiden anderen.
- Paulo Victor Miranda Boratto, Fábio Pio Dornas, Lorena Christine Ferreira da Silva, Rodrigo Araújo Lima Rodrigues, Graziele Pereira Oliveira, Juliana Reis Cortines, Betânia Paiva Drumond, Jônatas Santos Abrahão: Analyses of the Kroon Virus Major Capsid Gene and Its Transcript Highlight a Distinct Pattern of Gene Evolution and Splicing among Mimiviruses, in: J Virol. 92(2), 15. Januar 2018, e01782-17, doi:10.1128/JVI.01782-17, PMC 5752926 (freier Volltext), PMID 29118120
- NCBI: Saudi moumouvirus
- Leena H. Bajrai, Felipe L. de Assis, Esam I. Azhar, Priscilla Jardot, Catherine Robert, Jônatas Abrahão, Didier Raoult, Bernard La Scola: Saudi Moumouvirus, the First Group B Mimivirus Isolated from Asia, in: Front. Microbiol., 20. Dezember 2016, doi:10.3389/fmicb.2016.02029
- NCBI: Acanthamoeba castellanii mamavirus (Species)
- Tan Yeh Fong, Chai Ying Lim, Chun Wie Chong, Patricia Kim Chooi Lim, Ivan K. S. Yap, Pooi Pooi Leong, Kenny Voon: Isolation and Quantification of Mimivirus-Like and Marseillevirus-Like Viruses from Soil Samples in An Aboriginal (Orang asli) Village in Peninsular Malaysia, in: Intervirology 61(2), S. 1–4, August 2018, doi:10.1159/000491602, Medscape, PDF (PDF; 523 kB) Fig. 2 – SR1, SR4 und SR9 sind untereinander nahe verwandt, stehen aber den anderen Kandidaten der Linie A viel näher als dem Saudi moumouvirus (Linie B). Die Zuordnung zur Linie A ist daher wahrscheinlich.
-
Julien Guglielmini, Anthony C. Woo, Mart Krupovic, Patrick Forterre, Morgan Gaia: pnas.org, in: PNAS 116(39), 10./24. September 2019, S. 19585–19592, doi:10.1073/pnas.1912006116, PMID 31506349, Fig. 2. Dazu:
Julien Guglielmini, Anthony Woo, Mart Krupovic, Patrick Forterre, Morgan Gaia: Diversification of giant and large eukaryotic dsDNA viruses predated the origin of modern eukaryotes, auf: bioRxiv vom 29. Oktober 2018 (Preprint), doi:10.1101/455816 - NCBI: Hirudovirus strain sangsue
- Mondher Boughalmi, Isabelle Pagnier, Sarah Aherfi, Philippe Colson, Didier Raoult, Bernard La Scola: First Isolation of a Giant Virus from Wild Hirudo medicinalis Leech: Mimiviridae isolation in Hirudo medicinalis (PDF) in: Viruses 2013, 5, S. 2920–2930, doi:10.3390/v5122920
- Paulo V. M. Boratto, Thalita S. Arantes, Lorena C. F. Silva, Felipe L. Assis, Erna G. Kroon, Bernard La Scola, Jônatas S. Abrahão: Niemeyer Virus: A New Mimivirus Group A Isolate Harboring a Set of Duplicated Aminoacyl-tRNA Synthetase Genes. In: Front Microbiol., 2015; 6, S. 1256. doi:10.3389/fmicb.2015.01256, PMC 4639698 (freier Volltext), PMID 26635738.
- Gabriel Augusto Pires de Souza, Victória Fulgêncio Queiroz, Maurício Teixeira Lima, Erik Vinicius de Sousa Reis, Luiz Felipe Leomil Coelho, Jônatas Santos Abrahão: Virus goes viral: an educational kit for virology classes. In: Virology Journal, Band 17, Nr. 13, 31. Januar 2020, doi:10.1186/s12985-020-1291-9
- NCBI: Niemeyer virus (Species)
- Elton Alisson: New giant virus found in Brazil, bei: Agência FAPESP, Brasilien, 4. Juni 2014
- NCBI: Mimivirus battle6 (Species)
- NCBI: Mimivirus battle7 (Species)
- NCBI: Mimivirus battle19 (Species)
- NCBI: Mimivirus battle27 (Species)
- NCBI: Mimivirus battle57 (Species)
- NCBI: Mimivirus battle66 (Species)
- NCBI: Mimivirus battle83 (Species)
- NCBI: Mimivirus battle86 (Species)
- Anirvan Chatterjee, Thomas Sicheritz-Pontén, Rajesh Yadav, Kiran Kondabagil: Isolation and complete genome sequencing of Mimivirus bombay, a Giant Virus in sewage of Mumbai, India. In: Genomics Data, 9(C), Mai 2016, doi:10.1016/j.gdata.2016.05.013, Fig. 2
- NCBI: Mimivirus Cher (Species)
- NCBI: Mimivirus dakar4 (Species)
- NCBI: Mimivirus fauteuil
- Christelle Desnues, Bernard La Scola, Natalya Yutin, Ghislain Fournous et al.: Provirophages and transpovirons as the diverse mobilome of giant viruses, in: PNAS 109(44), 30. Oktober 2012, S. 18078–18083, doi:10.1073/pnas.1208835109
- NCBI: Fauteuil virus FD
- NCBI: Mimivirus huitre A06 (Species)
- NCBI: Mimivirus lactour (Species)
- NCBI: Lactours virus LT2 (Species)
- NCBI: Mimivirus lentille (Species)
- NCBI: Lentille virus CL (Species)
- NCBI: Mimivirus longchamps (Species)
- NCBI: Longchamps virus FPL (Species)
- NCBI: Mimivirus marais (Species)
- NCBI: Mimivirus montadette2 (Species)
- NCBI: Mimivirus pointerouge1 (Species)
- NCBI: Pointerouge virus 1 (Species)
- NCBI: Mimivirus pointerouge1 (Species)
- NCBI: Pointerouge virus 1 (Species)
- NCBI: Mimivirus SR1 (Species)
- NCBI: Mimivirus SR4 (Species)
- NCBI: Mimivirus SR9 (Species)
- NCBI: Mimivirus T2 (Species)
- NCBI: Mimivirus T3 (Species)
- NCBI: Mimivirus terra2 (Spezies)
- NCBI: Mimivirus univirus (Species)
- Jônatas Abrahão, Lorena Silva, Ludmila Santos Silva, Jacques Yaacoub Bou Khalil, Rodrigo Rodrigues, Thalita Arantes, Felipe Assis, Paulo Boratto, Miguel Andrade, Erna Geessien Kroon, Bergmann Ribeiro, Ivan Bergier, Herve Seligmann, Eric Ghigo, Philippe Colson, Anthony Levasseur, Guido Kroemer, Didier Raoult, Bernard La Scola: Tailed giant Tupanvirus possesses the most complete translational apparatus of the known virosphere. In: Nature Communications. 9, Nr. 1, 27. Februar 2018. doi:10.1038/s41467-018-03168-1.
{kind=link}