Irenolophus
Irenolophus ist eine Gattung aus der ausgestorbenen Familie der Deperetellidae, bei denen es sich um entfernte Verwandte der heutigen Tapire handelt. Die Gattung lebte im Unteren und Mittleren Eozän vor rund 52 bis 41 Millionen Jahren im heutigen Ostasien. Nachweise liegen aus der Inneren Mongolei vor. Sie umfassen einen vollständigen, aber stark zerdrückten Schädel mit Unterkiefer sowie weitere Gebissteile und einzelne Fußknochen. Es handelt sich um mittelgroße tapirartige Tiere, bei denen das Nasenbein noch relativ lang und der Naseninnenraum sehr klein war. Bei den Tapiren ist ersteres stark reduziert, letzterer deutlich vergrößert, beides entstand durch die Ausprägung des Rüssels. Weitere Merkmale finden sich in den einfach gestalteten Vormahlzähnen und den charakteristisch gebauten, mit zwei kräftigen Querleisten ausgestatteten Mahlzähnen. Die wissenschaftliche Einführung von Irenolophus erfolgte im Jahr 2019. Es werden momentan zwei Arten unterschieden.
| Irenolophus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
Schädel von Irenolophus, Holotypexemplar von I. qii | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unteres bis Mittleres Eozän | ||||||||||||
| 52 bis 41,1 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Irenolophus | ||||||||||||
| Bai, Meng, Mao, Zhang & Wang, 2019 | ||||||||||||
Merkmale
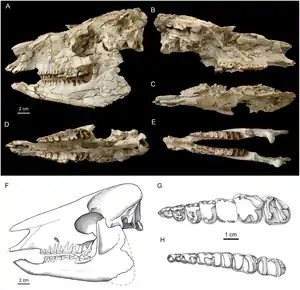
Irenolophus war ein mittelgroßer Vertreter der Tapiroidea. Die Gattung ist über einen vollständigen Schädel und mehrere Kieferreste überliefert. Der Schädel zeigt seitliche Verdrückungen, die durch Sedimentauflast entstanden, außerdem ist er stark zersplittert. Seine Gesamtlänge beträgt 24 cm, davon nahm der Schnauzenbereich rund 14 cm ein, er war damit gut 27 % länger als der Hirnschädelabschnitt. Insgesamt besaß der Schädel ein aufgewölbtes Profil. Der Mittelkieferknochen stand nicht nur mit dem Oberkiefer, sondern auch mit dem Nasenbein in Kontakt, was der Situation bei Heptodon entspricht, jedoch von Colodon abweicht, bei dem letzteres nicht zutrifft. Der Naseninnenraum war nur sehr kurz und endete wie bei Heptodon oberhalb der Lücke zwischen dem Eckzahn und dem vorderen Prämolaren. Bei Colodon und Lophialetes hingegen reichte er bis oberhalb der Molarenreihe zurück. Der Oberkiefer war hoch, das hier ausgebildete Foramen infraorbitale öffnete sich oberhalb des zweiten und dritten Prämolaren vergleichbar zu anderen Tapriverwandten, aber in einer niedrigeren Position als es etwa bei Heptodon und Colodon bekannt ist. Das Nasenbein war noch recht ausgedehnt und überragte nach vorn den hintersten Schneidezahn. Spätere Formen wie Colodon hatten deutlich kürzere Nasenbeine. Auf dem flachen Stirnbein erhoben sich zwei Knochenrippeln, die sich nach hinten zu einem deutlichen Scheitelkamm vereinten. Die Orbita war rund und recht groß, aber kleiner als in Lophioletes. Der hintere Teil des Schädels ist stark beschädigt, die Gelenkflächen am Hinterhauptsbein standen aber oben weit auseinander und unten eng beisammen. Der Gaumen auf der Schädelunterseite dehnte sich bis zum zweiten Molaren aus, in Übereinstimmung mit Heptodon. Der äußere Gehörgang öffnete sich nach unten. Am Unterkiefer bestand ein schlanker horizontaler Knochenkörper, der nur leicht nach hinten an Höhe zunahm. So betrug die Höhe unter dem zweiten Prämolaren 2,8, unter dem ersten Molaren 3,2 cm. Die Symphyse am vorderen Ende erstreckte sich bis zum ersten Prämolaren und war relativ schmal. Sie ragte vorn schräg nach oben. Im Gegensatz zum niedrigen Knochenkörper stieg der Kronenfortsatz bis zu 12 cm auf. Seine Vorderkante stand nahezu senkrecht zur Kauebene der Zähne. Der hintere Teil ist bei den meisten Unterkieferfunden beschädigt. Ein relativ vollständiges Exemplar maß 21 cm in der Länge.[1]
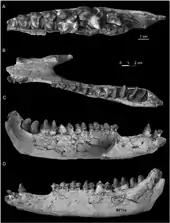
Das Gebiss bestand aus der vollständigen Bezahnung der Höheren Säugetiere mit insgesamt 44 Zähnen und folgender Zahnformel: . Das vordere Gebiss ist kaum bekannt, anhand der Alveolen war der hinterste Schneidezahn sowohl in der oberen als auch in der unteren Zahnreihe kleiner als die jeweils beiden vorderen. Der obere Eckzahn stand isoliert, ihn trennte eine kurzes Diastema vom dritten Schneidezahn und ein langes, bis zu 2,4 cm messendes vom ersten Prämolaren. Im Unterkiefer befand sich eine Zahnlücke nur zwischen Eckzahn und vorderstem Prämolaren. Der Eckzahn hatte in beiden Zahnreihen einen ovalen Querschnitt und eine wohl recht große Gestalt. Die Prämolaren waren nur wenig molarisiert, ähnelten den Mahlzähnen also nur wenig. Die Molaren besaßen einen strikt bilophodonten Bau mit zwei quergestellten Leisten, was typisch für die Tapirverwandtschaft ist. Markant für die Deperetellidae bildeten die beiden Leisten der oberen Molaren (Metaloph und Protoloph) zusammen ein U, wobei die vordere Leiste fast kontinuierlich in eine zusätzliche, und in die Struktur mit einbezogene Scherleiste, die Centrocrista, überging. Der Zusammenschluss war aber noch nicht so eindeutig ausgebildet wie bei Teleolophus. Der bei letzterer Gattung auf dem hintersten Mahlzahn vollständig verschwundene Metaconus war bei Irenolophus noch sichtbar. Auf den beiden vorderen Molaren bestand ebenfalls ein Metaconus, der entsprechend anderen Vertretern der Deperetellidae zungenwärts verschoben war. Auf den unteren Molaren verliefen die beiden Leisten (Protolophid und Hypolophid) nahezu parallel und waren etwas schräg gestellt, so dass sie zungenseitig nach hinten orientiert waren. Auf dem dritten Molaren fehlte das Hypoconulid, ein kleiner Nebenhöcker am hinteren Zahnende, der bei Lophialetes eine kleine dritte Leiste formte und bei Heptodon relativ schmal war. Die Prämolarenreihe war im Ober- und Unterkiefer jeweils rund 4 bis 4,7 cm lang, die Werte für die Molarenreihe lagen bei jeweils rund 5 bis 5,8 cm.[1]

Bemerkenswert an den aufgefundenen Handwurzelknochen ist, dass die Gelenkfazette des Hakenbeins zur Verbindung mit dem äußeren (fünften) Mittelhandknochen ausgesprochen groß war. Dies lässt darauf schließen, dass der äußere Fingerstrahl der Hand (Strahl V) noch nicht so stark reduziert war, wie dies etwa bei den heutigen Tapiren der Fall ist.[1]
Fossilfunde
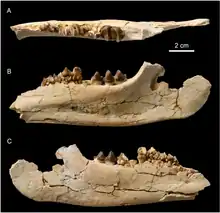
Das bisher bekannte Fossilmaterial von Irenolophus wurde zum größten Teil in der Arshanto-Formation, zu einem geringeren Teil auch in der Irdin-Manha-Formation entdeckt. Beide Gesteinseinheiten stellen wichtige Fossillagerstätten dar und haben für die ostasiatische Biostratigraphie zentrale Bedeutung, da einzelne Abschnitte nach ihnen benannt wurden. Sie sind im Erlian-Becken in der Inneren Mongolei aufgeschlossen, bedeutende Fundplätze finden sich südlich der Stadt Erlian. Erste Forschungsarbeiten erfolgten in der Region in den 1920er Jahren während mehrerer Expeditionen des American Museum of Natural History, jeweils geleitet von William Diller Matthew und Walter W. Granger. Die Arshanto-Formation erreicht eine Mächtigkeit von über 35 m. Sie liegt der Nomogen-Formation auf und beginnt basal mit gröberen fluviatil entstandenen und teils kreuzgeschichteten Sedimenten von grauer Färbung, die nach oben sukzessive in feinere Sandsteine übergehen. Der obere Abschnitt wird durch rötlich gefärbte tonige Ablagerungen gebildet. Die Irdin-Manha-Formation wiederum überdeckt die Arshanto-Formation. Ihre Mächtigkeit beträgt nur 10 bis 15 m. Sie zeigt sich als Serie von grau gefärbten Tonen, Sanden und Kiesen, in die zahlreiche Rinnen eingebetteten sind. Die Formationen gehören dem Unteren und Mittleren Eozän an, sie entstanden somit zwischen 52 und 41 Millionen Jahre vor heute (lokalstratigraphisch das Arshantum vor 52 bis 47 Millionen Jahren und das Irdinmanhum vor 47 bis 41 Millionen Jahren).[2][3]
Beide Gesteinseinheiten enthalten mehrere Fossilbereiche, die durch ihren generellen Fundreichtum herausragen. Allein unter den Unpaarhufern kommen bedeutende Reste vor. So sind aus der weiteren Tapirverwandtschaft (Tapiroidea) Heptodon aus der Arshanto- sowie Desmatotherium und Lophialetes aus der Irdin-Manha-Formation belegt. Ersterer entstammt auch die bedeutende Basalform Hyrachyus. In die weitere Nashornverwandtschaft (Rhinocerotoidea) gehören unter anderem Pappaceras und Forstercooperia aus der Gruppe der Indricotheriidae, die sich über beide Ablagerungseinheiten verteilen, beziehungsweise Rostriamynodon aus der Gruppe der Amynodontidae mit Resten aus der Irdin-Manha-Formation. Herausragend sind auch die Belege für die Brontotheriidae, da etwa Microtitan (Arshanto-Formation) sowie Protitan, Metatelmatherium und Gnathotitan (alle Irdin-Manha-Formation) von hier berichtet wurden. Darüber hinaus kommen zahlreiche Funde aus jüngeren Gesteinseinheiten vor, die das Obere Eozän und das Untere Oligozän abdecken.[4] Die Funde von Irenolophus streuen durch den größten Teil der Arshanto-Formation und finden sich vereinzelt noch in den unteren Partien der Irdin-Manha-Formation. Bisher geborgen wurden ein vollständiger Schädel mit Unterkiefer sowie einzelne Fußknochen vom Fundgebiet Huheboerhe. Sie lagen im Basisbereich der Arshanto-Formation und dienten zur Definition der Gattung Irenolophus. Darüber hinaus kamen verschiedene Unter- und Oberkiefer, isolierte Zähne und weitere isolierte Gliedmaßenfragmente aus den restlichen Bereichen der Gesteinseinheiten zu Tage. Sie verteilen sich auf verschiedene Fundgebiete, so neben Huheboerhe zusätzlich auch Wulanboerhe und Nuhertingboerhe.[1]
Systematik
Innere Systematik der Nashorn-Tapir-Verwandtschaft nach Bai et al. 2020[5]
|
Irenolophus ist eine Gattung aus der ausgestorbenen Familie der Deperetellidae. Die Gruppe gehört zur weiteren Verwandtschaft der Tapire (Tapiridae), den sogenannten Tapiroidea. Die Deperetellidae sind weitgehend auf das Eozän Asiens beschränkt. Sie bilden zusammen mit den Helaletidae und den Lophialetidae eine eher urtümliche Verwandtschaftsgemeinschaft früher tapirartiger Formen.[6][7] In einer weiteren Betrachtungsweise gehören die Tapiroidea und damit auch die Deperetellidae zu den Ceratomorpha, der gemeinsamen Gruppe der Tapire und Nashörner (Rhinocerotidae) beziehungsweise in die übergeordnete Gemeinschaft der Tapiromorpha. Traditionell wird die Tapir-Nashorn-Linie innerhalb der Ordnung der Unpaarhufer (Perissodactyla) als Schwestergruppe der Hippomorpha, der Pferdeverwandtschaft, betrachtet.[8][9]
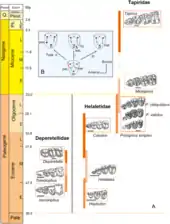
Ursprünglich wurden die Deperetellidae im Jahr 1965 von Leonard B. Radinsky etabliert. Sie zeichnen sich durch ihre vollständige Bezahnung der Höheren Säugetiere mit einem Diastema hinter dem Eckzahn, ihre nur teils molarisierten Prämolaren und ihre recht hochkronigen und extrem bilophodonten Molaren aus. Die beiden Leisten bilden auf den oberen Mahlzähnen zusammen mit dem Paraconus einen U-förmigen Verlauf, während der Metaconus in seiner Größe reduziert oder ganz verschwunden ist. Eine weitere Besonderheit der Deperetellidae findet sich in dem Molarisationsmodus der oberen Prämolaren, der etwas anders verlief als bei den Tapiren und einigen verwandten Gruppen sowie beim Großteil der Nashorngemeinschaft. Hierbei entstand die hintere Zahnleiste (Metaloph) nicht aus dem sich am hinteren Zahnrand neu herausbildenden Hypoconus, einem großen Zahnhöcker, sondern aus der zungen- und rückwärtigen Abwanderung einzelner bereits bestehender Höcker, namentlich der Protoconus und das Paraconule. Dies erinnert ein wenig an den Vorgang bei den Pferden.[10][1] Radinsky verwies 1965 mit Deperetella und Teleolophus nur zwei Gattungen in die Gruppe, denen nach und nach weitere zur Seite gestellt wurden wie etwa Pachylophus und Bahinolophus.[11][12] Die Herkunft der Deperetellidae ist nicht ganz eindeutig. Der vollständige Schädel von Irenolophus zeigt mit dem nur kurzen Naseninnenraum, dem niedrig sitzenden Foramen infraorbitale und dem Vorkommen des ersten Prämolaren sehr basale Eigenschaften, die den meisten Mitgliedern der Helaletidae mit Ausnahme von Heptodon fehlen. Die Zurücksetzung des Naseninnenraums und Verkürzung des Nasenbeins in späteren tapirartigen Formen hängen dabei offensichtlich mit der Ausprägung eines Rüssels zusammen.[13] Vorläufig wird daher davon ausgegangen, dass die Deperetellidae aus frühen helaletiden Tapirartigen hervorgingen. Innerhalb der Deperetellidae kann Irenolophus wiederum als basale Form angesehen werden. Der Schluss ergibt sich aus der relativ kurzen Prämolarenreihe gegenüber der Molarenreihe, da erstere bei späteren Formen verhältnismäßig an Länge zunimmt.[1]
Die Gattung Irenolophus wurde im Jahr 2019 durch Bin Bai und Forscherkollegen wissenschaftlich erstbeschrieben. Grundlage hierfür bildeten ein Schädel samt Unterkiefer und einige Handknochen aus dem Basisbereich der Arshanto-Formation im Fundegebiet Huheboerhe im Erlian-Becken in der Inneren Mongolei (Exemplarnummer IVPP V 25831). Der Gattungsname leitet sich von der Bezeichnung Iren für das Erlian-Becken und vom griechischen Wort λόφος (lophos) für „Hügel“ oder „Kamm“ her. Letzteres Wortteil bezieht sich auf die ausgeprägte Leistenbildung der Mahlzähne.[1]
Es können innerhalb der Gattung Irenolophus zwei Arten unterschieden werden:[1]
- I. qii Bai, Meng, Mao, Zhang & Wang, 2019
- I. primarius (Qi, 1987)
Die Art Irenolophus qii stellten Bai und Kollegen in ihrer Erstbeschreibung 2019 auf. Mit dem Artepitheton ehrten die Autoren den Forscher Tao Qi für seine verdienstvollen Arbeiten im Erlian-Becken.[1] Qi hatte bereits 1987 einen umfangreichen Aufsatz zur Säugetierfauna der Arshanto-Formation publiziert. In diesem führte er Teleolophus primarius als neuen Vertreter der Deperetellidae ein.[14] Das Fundmaterial dieser Form, verschiedene Gebiss- und Fußknochen, kommt ebenfalls im Huheboerhe und zusätzlich noch in Wulanboerhe vor, streut jedoch stärker durch die Arshanto-Formation und reicht bis in die liegenden Schichten der Irdin-Manha-Formation. Im Gegensatz zu anderen Angehörigen der Gattung Teleolophus zeigt Teleolophus primarius einige ursprünglichere Zahnmerkmale wie das nicht gegabelte Paralophid der unteren Prämolaren oder ein hier auch fehlendes Entoconid, einen großen Zahnhöcker. Bai und Kollegen führten daher Teleolophus primarius in die Gattung Irenolophus über. Zusätzlich vereinigten sie einige Fossilreste, die Radinsky 1965 unter Teleolophus cf medius erwähnt hatte und von den gleichen Lokalitäten vorliegen,[6] mit Irenolophus primarius. Im Vergleich zu Irenolophus qii zeigt Irenolophus primarius eine längere Prämolarenreihe bezogen auf die Molarenreihe (das Verhältnis Prämolaren- zu Molarenreihe beträgt bei I. qii 0.79, bei I. primarius 0,82). Eine möglicherweise dritte Art von Irenolophus könnte ein rechter Unterkieferast repräsentieren, der aus der Nuhertingboerhe-Region ebenfalls im Basisbereich der Arshanto-Formation zu Tage kam. Im Vergleich zur Typusart Irenolophus qii sind hier einzelne Abweichungen in der Zahnmorphologie und in der Position des Foramen mandibulae erkennbar. Da aber hier nur der letzte Molar vollständig ist und somit zu wenig Vergleichsmaterial vorliegt, wurde von der Aufstellung einer dritten Art vorerst abgesehen.[1]
Literatur
- Bin Bai, Jin Meng, Fang-Yuan Mao, Zhao-Qun Zhang und Yuan Qing Wang: A new early Eocene deperetellid tapiroid illuminates the origin of Deperetellidae and the pattern of premolar molarization in Perissodactyla. PLoS ONE 14 (11), 2019, S. e0225045, doi:10.1371/journal.pone.0225045
Einzelnachweise
- Bin Bai, Jin Meng, Fang-Yuan Mao, Zhao-Qun Zhang und Yuan Qing Wang: A new early Eocene deperetellid tapiroid illuminates the origin of Deperetellidae and the pattern of premolar molarization in Perissodactyla. PLoS ONE 14 (11), 2019, S. e0225045, doi:10.1371/journal.pone.0225045
- Jin Meng, Yuanqing Wang, Xijun Ni, K. Christopher Beard, Chengkai Sun, Qian Li, Xun Jin und Bin Bai: New Stratigraphic Data from the Erlian Basin: Implications for the Division, Correlation, and Definition of Paleogene Lithological Units in Nei Mongol (Inner Mongolia). American Museum Novitates 3570, 2007, S. 1–31
- Bin Bai, Yuan-Qing Wang, Fang-Yuan Mao und Jin Meng: New material of Eocene Helaletidae (Perissodactyla, Tapiroidea) from the Irdin Manha Formation of the Erlian Basin, Inner Mongolia, China and comments on Related Localities of the Huheboerhe Area. American Museum Novitates 3878, 2017, S. 1–44
- Bin Bai, Yuanqing Wang, Qian Li, Haibing Wang, Fangyuan Mao, Yanxin Gong und Jin Meng: Biostratigraphy and Diversity of Paleogene Perissodactyls from the Erlian Basin of Inner Mongolia, China. American Museum Novitates 3914, 2018, S. 1–60
- Bin Bai, Jin Meng, Chi Zhang, Yan-Zin Gong und Yuan-Qing Wang: The origin of Rhinocerotoidea and the phylogeny of Ceratomorpha (Mammalia, Perissodactyla). Communications Biology 3, 2020, S. 509, doi:10.1038/s42003-020-01205-8
- Leonard B. Radinsky: Early Tertiary Tapiroidea of Asia. Bulletin of the American Museum of Natural History 129 (2), 1965, S. 183–263
- Robert M. Schoch: A review of the Tapiroids. In: Donald R. Prothero und R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New-York 1989, S. 298–320
- Jerry J. Hooker und Demberelyin Dashzeveg: The origin of chalicotheres (Perissodactyla, Mammalia). Palaeontology 47 (6), 2004, S. 1363–1386
- Luke T. Holbrook und Joshua Lapergola: A new genus of Perissodactyl (Mammalia) from the Bridgerian of Wyoming, with comments on basal Perissodactyl phylogeny. Journal of Vertebrate Paleontology 31 (4), 2011, S. 895–901
- Luke Holbrook: The Identity and Homology of the Postprotocrista and its Role in Molarization of Upper Premolars of Perissodactyla (Mammalia). Journal of Mammalogy 22, 2015, S. 259–269, doi:10.1007/s10914-014-9276-3
- Yongsheng Tong und Yizhen Lei: Fossil tapiroids from the upper Eocene of Xichuan, Henan. Vertebrata PalAsiatica 22 (4), 1984, S. 269–280
- Takeshi Tsubamoto, Naoko Egi, Masanaru Takai, Chit Sein und Maung Maung: Middle Eocene ungulate mammals from Myanmar: A review with description of new specimens. Acta Palaeontologica Polonica 50 (1), 2005, S. :117–138 ()
- Leonard B. Radinsky: Origin and Early Evolution of North American Tapiroidea. Peabody Museum of Natural History Yale University Bulletin 17, 1963, S. 1–106.
- Tao Qi: The Middle Eocene Arshanto fauna (Mammalia) of Inner Mongolia. Annals of the Carnegie Museum 56, 1987, S. 1–73 ()