Paraceratherium
Paraceratherium (früher auch als Baluchitherium, Indricotherium oder Dzungariotherium beschrieben) ist eine Gattung aus der ausgestorbenen Familie der Indricotheriidae, die zu den Nashornartigen (Rhinocerotoidea) gehört und mit einem Gewicht von bis zu 20 Tonnen die größten landbewohnenden Säugetiere aller Zeiten stellte. Lediglich einige der größten Rüsseltiere könnten ähnliche Dimensionen erreicht haben. Weiterhin zeichnete es sich durch lange Beine, einen verlängerten Hals und einen vergleichsweise kleinen Schädel aus. Die Gattung lebte im Oligozän und im untersten Miozän vor etwa 34 bis rund 22 Millionen Jahren. Sie ist von zahlreichen, teils gut erhaltenen Fossilresten aus Ost-, Zentral- und Westasien bekannt, kam aber auch im heutigen Südosteuropa vor. Laut Untersuchungen der Backenzähne von Paraceratherium ernährte es sich hauptsächlich von weicher Pflanzenkost, dabei lebte es in teils offenen Landschaften, die unter subtropisch-warmem Klima bestanden. Die mitunter häufig genutzten Synonymnamen beruhen auf unabhängigen Beschreibungen von Fundmaterial aus unterschiedlichen Fundregionen, die überwiegend zu Anfang des 20. Jahrhunderts in recht kurzen Zeitabständen erstellt, aber schließlich im Jahr 1989 offiziell mit Paraceratherium vereint wurden. Die wissenschaftliche Entdeckung der Gattung begann Anfang des 20. Jahrhunderts. Der Name Paraceratherium leitet sich aus den griechischen Wörtern παρά (pará) für „neben“, κέρας (keras) für „Horn“ und θηρίον (thēríon) für „Tier“ ab und bezieht sich auf die nahe Verwandtschaft mit den Nashörnern.
| Paraceratherium | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion eines Paraceratherium | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oligozän bis Untermiozän | ||||||||||||
| 34 bis 22 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Paraceratherium | ||||||||||||
| Forster Cooper, 1911 | ||||||||||||
Merkmale
Habitus
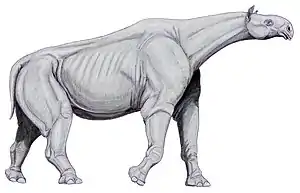
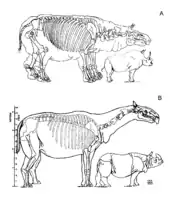
Paraceratherium war ein relativ langbeiniges und langhalsiges Tier mit einem kräftigen Körperbau und einem verhältnismäßig kleinen Kopf. Anhand der gefundenen Knochen wird für die größten Arten eine Kopf-Rumpf-Länge von 7,4 bis 8,7 m und eine Schulterhöhe von 4,5 bis 5 m angenommen (ohne Berücksichtigung der Bedeckung des Skelettes durch Weichteilgewebe), die Halslänge betrug rekonstruiert wohl rund 2 bis 2,5 m.[1] Das Gewicht erreichte dabei schätzungsweise 15 bis 20 Tonnen, weibliche Tiere waren möglicherweise deutlich kleiner.[2] Kleinere Arten wurden nur rund 6,2 m lang und besaßen ein Gewicht von etwa 7 bis 9 Tonnen.[3][4]
Schädel- und Gebissmerkmale
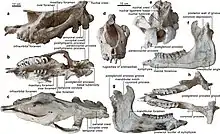
Ein vollständig erhaltener Schädel war bis zu 130 cm lang und an den Jochbeinbögen bis zu 61 cm breit, einige fragmentierte Stücke lassen noch größere Exemplare annehmen. Dabei war der Schädel ausgesprochen langgestreckt und im Bereich des Gehirnschädels schmal und niedrig. Allerdings besaß der Oberschädel in der Mitte eine leichte kuppelartige Aufwölbung.[2] In Relation zum gesamten Körper ist er verglichen mit heutigen Nashörnern eher klein.[5] Das Nasenbein zeigte eine nur schwache Entwicklung und wies keine Aufrauhungen auf, womit es belegt, dass die Gattung keine Hörner besaß. Das Hinterhauptsbein war schmal und lang und nicht so breit gebaut wie bei den heutigen Nashörnern. Es besaß lediglich einen schwach ausgeprägten Hinterhauptswulst, was auf eine weniger gut entwickelte Nackenmuskulatur schließen lässt – ein weiterer Hinweis auf die Abwesenheit eines Horns. Allerdings waren die Gelenkansätze am Hinterhaupt für die Wirbelsäule enorm groß.[6][2] Der Unterkiefer war lang und schmal, aber recht kräftig gebaut. Die Symphyse reichte bis zum dritten Prämolar und war damit äußerst lang und robust.[7][8]
Paraceratherium hatte ein teilweise reduziertes Gebiss mit folgender Zahnformel: .[9] Im vorderen Gebiss besaß Paraceratherium oben und unten jeweils ein Paar Schneidezähne, die eine dolchartige Form aufwiesen und mit einer Länge von 6 bis 7 cm kleinen Stoßzähnen ähnelten.[7] Dabei waren oben jeweils die zweiten Schneidezähne (I2) ausgebildet, die eher senkrecht oder leicht schräg nach vorn gerichtet im Kiefer saßen. Im Unterkiefer dagegen befanden sich die ersten Schneidezähne (I1) in horizontaler oder schräg aufsteigender Position. Ein weiteres kleines, äußeres Schneidezahnpaar konnte manchmal noch im Unterkiefer ausgebildet sein, zusätzlich kam im Oberkiefer noch ein rudimentär erhaltener Eckzahn vor.[10] Zwischen vorderem und hinterem Gebiss befand sich ein großes Diastema. Dass hintere Gebiss bestand im Oberkiefer aus vier Prämolaren und drei Molaren, im Unterkiefer fehlte der erste Prämolar. In Einzelfällen war auch im Oberkiefer der erste Backenzahn reduziert.[11] Die vorderen Prämolaren waren sehr klein, während die hinteren fast die Größe der folgenden Molaren erreichten. Die Molaren selbst ähnelten denen der Nashörner, besaßen aber weniger geschwungene Schmelzfalten. Der Zahnschmelz war aber mit 4 mm sehr dick. Allgemein zeichneten sich die Molaren durch moderat hohe (hypsodonte) Zahnkronen aus. Häufig wies der letzte Molar die größten Dimensionen auf und besaß Längen von über 10 cm. Allgemein waren die Backenzähne aber, bezogen auf die generelle Größe der Tiere, sehr klein, die Gesamtlänge der Backenzahnreihe betrug 40 cm und mehr.[2][12]

Körperskelett
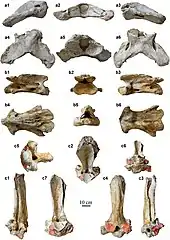
Die am Schädel ansetzenden Halswirbel erreichten eine enorme Größe und waren in der Länge gestreckt, was den langen Hals von Paraceratherium bewirkte. Die Länge der gesamten Halswirbelsäule übertraf die des Schädels um das anderthalb bis zweifache.[9] Die Lage der Gelenkflächen an den Wirbeln zeigt, dass der Kopf in der Regel horizontal gehalten wurde, dafür aber seitlich gut ausschwenkbar war. Außerdem waren die beiden ersten Halswirbel deutlich kürzer sowie breiter und schränkten dabei eine vertikale Bewegung ein. Aufgrund dessen war eine pferde- oder okapiähnliche dauerhafte Kopfhaltung mit nach oben angewinkeltem Hals kaum möglich. Die Dornfortsätze der ersten Brustwirbel waren extrem lang und indizieren dadurch einen kleinen Buckel direkt hinter dem Nacken, zudem aber auch eine Muskulatur, die kräftig genug war, den schweren Kopf zu halten. Seitlich an den Körpern der Wirbel vor dem Kreuzbein bestanden kleine, rundliche Aushöhlungen (Pleurocoele oder Luftsäcke), die denen der großen Sauropoden ähneln, es ist aber unbekannt, womit diese gefüllt waren.[13][2] Die Gliedmaßen waren sehr langgestreckt und setzten wie bei den heutigen Nashörnern leicht gewinkelt unter dem Körper an. Der Oberschenkelknochen erreichte Längen von 120 bis 150 cm, dieser besaß zudem im Gegensatz zu den nahe verwandten Nashörnern weniger stark ausgebildete Trochanter (Großer, Kleiner und Dritter), was möglicherweise auf die enorme Streckung des Gesamtknochens zurückzuführen ist und eine Parallele in der Entwicklung der Elefantenfemora findet. Das Schienbein war dagegen mit 80 bis 90 cm Länge deutlich kürzer. Generell war der Oberarmknochen mit 90 bis 99 cm kürzer als die Speiche, die teilweise über 120 cm Länge erreichte. Das Verhältnis dieser beiden Langknochen zueinander lässt auf eine Herkunft von schnellläufigen Vorahnen schließen. Sowohl die Vorder- als auch die Hinterbeine endeten in je drei Zehen, wobei der mittlere am größten war. Die Metapodien waren dabei seitlich teilweise verschmälert und markant verlängert – der mittlere konnte bis über 63 cm lang werden –, die Zehenknochen zusätzlich deutlich abgeplattet.[14][15][2]
Fundstellen
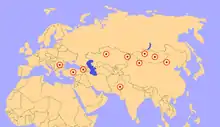
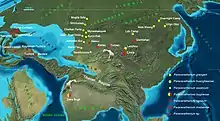
Bedeutende Fundstellen finden sich im ehemaligen Belutschistan im heutigen westlichen Pakistan. Von dort stammen auch die ersten Funde aus den Bugti-Bergen.[7][12] Sehr fundreiche Gebiete liegen des Weiteren bei Torghai im nordwestlichen Kasachstan, deren Fossilien zu den größten Vertretern von Paraceratherium gehören,[15] aber auch in anderen Landesteilen sind Reste dieser Tiergattung bekannt, so aus den Aktau-Bergen im Südosten.[16][17] Von herausragender Bedeutung ist vor allem das Tsagan-Nor-Becken in der Mongolei, wo die meisten Funde aus der Hsanda-Gol-Formation stammen und den bisher vollständigsten Schädel einschließen. Auch diese Funde repräsentieren teilweise sehr große Formen.[14] Die nördlichen und nordwestlichen Provinzen Chinas bergen überdies ebenfalls zahlreiche Fossilien, die aber auch andere Indricotherien-Formen umfassen. Genannt werden können hier etwa das Linxia- und Lanzhou-Becken in der Provinz Gansu sowie das Dsungarische Becken, das Hami-Becken und die Turpan-Senke in der autonomen Region Xinjiang.[18][8][19] In jüngerer Zeit wurden Paraceratherium-Funde auch aus der Türkei bekannt, die teilweise sehr großen Vertretern angehören, so etwa aus der Kızılırmak-Formation im Çankırı-Çorum-Becken im zentralen und aus der Güngörmez-Formation im Kağızman-Tuzluca-Becken im nordöstlichen Anatolien.[20][21] Die bisher westlichsten Fundpunkte liegen in Südosteuropa, wo Reste von Paraceratherium unter anderem in der Cuzăplac-Formation in Rumänien vorkommen,[22] aber auch aus Montenegro (Berane) und Bulgarien (Dragovištiza) wurden einzelne Knochen berichtet.[21]
Paläobiologie
Körpergröße
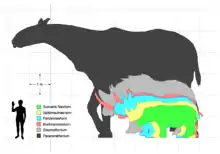
Ursprünglich wurde von einem Gewicht von bis zu 34 Tonnen ausgegangen, diese Werte beruhten aber auf Proportionsvergleichen mit heutigen Nashörnern und resultierten teilweise aus einem ersten, aber missglückten Rekonstruktionsversuch von Henry Fairfield Osborn aus dem Jahr 1923.[23] Diese Gewichtsangaben erwiesen sich aber nach neueren Studien als zu hoch, so dass heute von einem maximalen Gewicht von 15 bis 20 Tonnen, im Extremfall von 24 Tonnen ausgegangen wird, bei kleineren Exemplaren nur von 7 bis 9 Tonnen. Dabei beruhen diese Studien weitgehend auf dem Vergleich einzelner Knochen zueinander, vor allem des Schädels und der Gliedmaßen, und bezogen auch zahlreiche andere Tierarten mit ein.[3][24] Trotzdem müssen diese Gewichtsangaben nur als ungefähre Schätzwerte angesehen werden, da die Indricotherien in ihren Körperproportionen deutlich von anderen Unpaarhufern abweichen. Aufgrund dieser Werte gilt Paraceratherium aber letztendlich als das größte bekannte Landsäugetier aller Zeiten. Im Vergleich zum heutigen Afrikanischen Elefanten, der etwa 4 bis 6 Tonnen wiegt, ist Paraceratherium durchaus größer, der größte jemals geschossene Elefant wog jedoch immerhin rund 10 Tonnen und ist heute in der Smithsonian Institution ausgestellt. Die größten bekannten Rüsseltiere wiesen dem gegenüber ein Gewicht vergleichbar zu Paraceratherium auf. So wurde der Steppenelefant aus dem Pleistozän Eurasiens bis zu 4,5 m hoch und wog gut 14 Tonnen, ähnliche Angaben liegen für einige Vertreter der Gattung Palaeoloxodon vor. Deinotherium giganteum aus dem Miozän brachte es immerhin noch auf 4,3 m Schulterhöhe und 13 Tonnen Gewicht.[25][26] Möglicherweise stellen die von Paraceratherium erreichten 15 bis 20 Tonnen aufgrund der speziellen Nahrungsverwertung im Magen-Darm-Trakt die Obergrenze der Körpermasse bei landlebenden Säugetieren dar.[27] Die angegebenen Größenwerte setzen voraus, dass Paraceratherium über eine ausreichende Thermoregulation verfügt haben muss. Das subtropische Klima, in dem es überwiegend lebte, lässt somit annehmen, dass es eine nur geringe Fellbedeckung besaß, ähnlich den anderen sehr großen Säugern heute, da ein dichtes Fell die überschüssige Körperwärme nur unnötig speichert und schlecht ableitet. Weiterhin deuten die Knochen rund um den Ohreingang, vor allem der Processus mastoideus und der Processus paroccipitalis, die beide jenen heutiger Elefanten entsprechen, darauf hin, dass die Ohrmuschel möglicherweise deutlich größer gestaltet war als vergleichsweise bei den heutigen Nashörnern und so ebenfalls einen Beitrag zur Regulierung des Wärmehaushaltes leisten konnte.[4] Diese Interpretation wird aber teilweise auch skeptisch gesehen.[28]
Fortbewegung
Die meisten der heutigen extrem großen Säugetiere, wie die Elefanten, Nashörner und Flusspferde, besitzen Anpassungen an eine schwerfällige Fortbewegungsweise (graviportal), die sich aufgrund eines fehlenden Drucks durch Fressfeinde im ausgewachsenen Alter entwickelte, ihnen es aber ermöglicht, das hohe Körpergewicht zu tragen. Diese zeigen sich hauptsächlich bei den Gliedmaßen, die vor allem durch lange obere Abschnitte (Oberarm- und Oberschenkelknochen) und verkürzte untere, insbesondere der Hand- und Fußknochen, charakterisiert sind. Gerade bei den Metapodien (Mittelhand- und Mittelfußknochen) und den Phalangen (Finger- und Zehenglieder) sind diese Verkürzungen besonders deutlich erkennbar, die eine sehr schnelle Fortbewegung einschränken. Paraceratherium wies im Gegensatz dazu vergleichsweise kurze obere und lange untere Gliedmaßenabschnitte auf, vor allem an den vorderen Beinen, die auf seine Herkunft von den schnellläufigen (cursorialen) Hyracodontidae hinweisen. Als weiteres Anzeichen kann die Größenreduktion der seitlichen Strahlen der Vorder- und Hinterfüße gedeutet werden, da diese eine Tendenz zur Monodactylie erkennen lässt.[9] Im Gegensatz zu echten Fluchttieren wie etwa Pferden oder Rehen sind aber die Phalangen deutlich in ihrer Länge gekürzt und gleichen so denen der schwerfällig gebauten Nashörner und Elefanten, eine ähnliche Entwicklung ist auch bei den großen Sauropoden zu beobachten.[13] Daraus lässt sich schließen, dass auch Paraceratherium in einer sehr schnellen Fortbewegung eingeschränkt war.[4]
Ernährungsweise
Der Bau der Zähne, der lange Hals und die generelle Größe der Tiere mit hoher Kopfposition machen es wahrscheinlich, dass Paraceratherium auf weiche Pflanzennahrung mit geringem Kieselsäureanteil spezialisiert war und sich höchstwahrscheinlich von Blättern, Zweigen, Knospen oder Blüten ernährte (browsing). Mikroskopische Abschleifspuren an den Backenzähnen zeigen dabei ein typisches Muster, welches jenem der heutigen, auf weiche Pflanzennahrung spezialisierten Pflanzenfressern gleicht. Zudem ergaben Isotopenanalysen der Backenzähne, die sowohl an Funden aus China als auch aus Pakistan vorgenommen wurden, dass sich Paraceratherium ausgehend vom Verhältnis der Kohlenstoff-Isotopen zueinander fast ausschließlich von Blättern ernährte.[29][30] Seine Größe ermöglichte es ihm dabei, in den Baumkronen mittelhoher Bäume seiner Zeit zu weiden, ähnlich wie es bei den heutigen Giraffen in Afrika zu beobachten ist. Da bei zahlreichen rezenten auf Blattnahrung spezialisierten Tieren, wie beispielsweise bei den diversen sich so ernährenden Nashornarten, eine sehr bewegliche Oberlippe zur Unterstützung bei der Nahrungsaufnahme ausgebildet ist, kann eine solche auch bei Paraceratherium angenommen werden. Knöcherne Erhebungen als Muskelansatzstellen im Gesichtsschädel ebenso wie der weit nach hinten reichende Naseninnenraum weisen darauf hin, dass die Oberlippe sogar die Form eines kleinen, kurzen Rüssels, ähnlich den Tapiren, besessen haben könnte.[31][4]
Es ist zu vermuten, dass bei Paraceratherium analog zu den heutigen und nachgewiesen auch bei einigen ausgestorbenen Unpaarhufern die Verdauung im hinteren Darmtrakt stattfand (Enddarmfermentierer). Daraus lässt sich schließen, dass die Verwertung der aufgenommenen Nahrung schlechter erfolgte als bei den Wiederkäuern, wodurch es nötig für ein Tier wurde, täglich große Nahrungsmengen aufzunehmen. Im Vergleich zu der Menge an Futter, die ein heutiger Elefant pro Tag vertilgen muss, ist von einer mindestens ebenso großen bei Paraceratherium auszugehen.[4]
Paläolandschaft

Die Größe der Tiere deutet darauf hin, dass sie in offenen Waldlandschaften oder Baumsavannen zu Hause waren.[23] Neuere Untersuchungen an Fundstellen mit Paraceratherium-Resten belegen diese Vermutung.[20] Diese offenen Landschaften gehen mit Klimaveränderungen während des Oligozän einher, die zu kühleren und trockeneren Klimaten führten und so zur Öffnung der Landschaften beitrugen. Da Paraceratherium überwiegend Blattnahrung zu sich nahm, muss in solchen offenen Landschaften der Aktionsradius einzelner Tiere extrem groß gewesen sein, um die notwendige tägliche Nahrungsmenge zu erreichen. Ob diese Herden bildeten, ist unbekannt, die meisten Unpaarhufer leben solitär mit Mutter-Kalb-Beziehungen während der Aufzucht, nur einige Pferdearten bilden heute Herdenverbände, allerdings ist dies eine eher moderne Anpassung an Steppen- und Savannenlandschaften, die ab dem Miozän entstanden.[32] Ausgewachsene Vertreter von Paraceratherium hatten wohl aufgrund der Größe kaum natürliche Feinde, allerdings finden sich an einigen Knochen aus den Bugti-Bergen in Pakistan Bissspuren, die auf ein riesiges Krokodil, Astorgosuchus, zurückgeführt werden, das bis zu 11 m lang wurde und offensichtlich gelegentlich ein Tier angriff.[4]
Stammesgeschichte
Paraceratherium geht vermutlich auf das nur ponygroße Forstercooperia als Basisform der Indricotherien aus dem mittleren Eozän zurück.[33][34] Die darauf folgenden Gattungen Juxia und Urtinotherium aus dem späten Eozän und frühen Oligozän waren ebenfalls aufgrund der andersartigen Gebissmorphologie sehr urtümlich.[35] Im frühen Oligozän vor 37 Millionen Jahren ist dann auch erstmals Paraceratherium nachgewiesen, zu den frühesten Funden gehören jene von Nei-Monggol (China). Die Gattung war über weite Gebiete von West- bis Ostasien verbreitet und kam in einer späteren Phase auch im südöstlichen Europa vor. Im Übergang vom Oberen Oligozän zum Unteren Miozän vor rund 23 Millionen Jahren starb sie dann aus. Zu den stammesgeschichtlich jüngsten Nachweisen gehören jene von den Bugti-Bergen in Pakistan,[36][37] Altersangaben aus Lundo Chur, wo Ende der 1990er Jahre erneut Funde gemacht wurden, ergaben mit der Fission-Track-Datierung etwa 22,6 Millionen Jahre.[12] Dieses Aussterben ging möglicherweise mit weiteren klimatischen Abkühlungen einher, aber auch mit der Ankunft neuer, konkurrenzfähigerer Pflanzenfresser, etwa der Rüsseltiere, die zu jener Zeit Eurasien über die neuentstandene Landbrücke durch die Schließung der Tethys aus Afrika erreichten und möglicherweise, ähnlich den heutigen Elefanten, aufgrund ihrer Lebensweise die Landschaften grundsätzlich zu ändern vermochten.[38][4]
Forschungsgeschichte
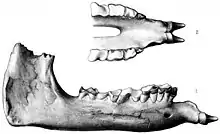
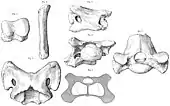
Der Name Paraceratherium wurde 1911 von Sir Clive Forster Cooper (1880–1947) eingeführt.[7] Die ersten veröffentlichten Knochen dieses Tieres, darunter ein hinterer Teil eines Unterkiefers und einzelne Oberkieferzähne, die während einer Expedition nach Dera Bugti (Bugti-Berge) in Belutschistan (Pakistan) entdeckt wurden, hatte bereits Guy Ellcock Pilgrim (1875–1943) im Jahr 1910 als Aceratherium bugtiense beschrieben. Weitere isolierte Oberkieferzähne wies Pilgrim der Art Bugtitherium grandincisivum zu, die vollständige Publikation des Materials erfolgte aber erst 1912.[39] Pilgrims Interesse an dieser Region war durch die Fossilienfunde von William Thomas Blanford (1832–1905) aus dem Jahr 1882 geweckt worden.[40] Die frühesten bekannten Funde stammten aber aus dem Jahr 1846, die ein Soldat namens Vickary fand, aufgrund ihres fragmentierten Charakters aber lange Zeit nicht zugeordnet werden konnten.[4] Forster Cooper leitete 1910 eine eigene Expedition in das Gebiet und fand einen fast vollständigen Unterkiefer, mehrere Wirbel und Langknochen. Auf Basis dieses Materials und unter Berufung auf Pilgrims erster kurzer Erwähnung veröffentlichte Forster Cooper die Erstbeschreibung von Paraceratherium bugtiense, wobei der Unterkiefer als Holotyp der Gattung dient. Während einer weiteren Expedition im Jahr 1911 in die Region, genauer nach Lundo Tschur, ebenfalls Belutschistan, entdeckte Forster Cooper neben einem weitgehend vollständigen auch drei stärker fragmentierte Schädel.[6] Die Fundstellen von Dera Bugti wurden erst in den der Mitte der 1990er Jahre wiederentdeckt, wobei die Originalfundstelle der heutigen Chitawara-Formation zugewiesen werden konnte.[41][12]
Der Gattungsname Indricotherium basiert auf einer Beschreibung von Alexei A. Borissiak (1872–1944) aus dem Jahr 1915, die anhand von Funden aus Torghai im nördlichen Turkestan (Kasachstan) erfolgte. Diese umfassten neben Gebissfragmenten einen Großteil der Wirbel und der Knochen des Bewegungsapparates.[42][15] Da die vollständige Artbenennung als Indricotherium asiaticum erst 1923 abgeschlossen war,[43] wurde dieser Name nicht anerkannt, da bereits im vorangegangenen Jahr M. Pavlova Indricotherium transouralicum anhand von Funden, darunter ein 63 cm langes Unterkieferfragment mit vollständiger hinterer Bezahnung, aus der gleichen Region beschrieben hatte.[44]
Weiteres umfangreiches Knochenmaterial wurde 1922 während der Second Central Asiatic Expedition of the American Museum of Natural History unter Leitung von Roy Chapman Andrews (1884–1960) und Walter W. Granger (1872–1942) im Tsagan-Nor-Becken in der Mongolei entdeckt, darunter ein Unterkiefer, ein nahezu vollständiger, aber in mehr als 360 Einzelteile[45] zerfallener, 129 cm langer Schädel sowie ein Oberarmknochen. Diese wurden im darauffolgenden Jahr von Henry Fairfield Osborn (1857–1935) als Baluchitherium grangeri beschrieben.[14] Der Gattungsname Baluchitherium war aber bereits 1913 von Forster Cooper aufgrund mehrerer sehr großer und seiner Meinung nach nicht zu Paraceratherium passender Wirbel eingeführt worden, nachdem der ursprünglich vorgesehene Name Thaumastotherium osborni für die von ihm neu beschriebene Art abgelehnt worden war, da die Gattung schon für ein Insekt aus der Gruppe der Schnabelkerfe vergeben worden war.[46][47]
Während die ersten Funde aus China in das Jahr 1922 in der Ordos-Region der Inneren Mongolei am Ufer des Gelben Flusses datieren, begannen systematische Untersuchungen erst Ende der 1950er, Anfang der 1960er Jahre und standen teilweise in Verbindung mit gemeinsamen Expeditionen zusammen mit sowjetischen Paläontologen. Dabei kamen bedeutende Fossilreste zu Tage, die alle aus dem nördlichen Teil Chinas, vor allem aus Xinjiang und aus der Inneren Mongolei stammen und mehrere Schädel umfassen, darunter auch ein 121 cm langes Exemplar aus dem Dzungaria-Becken, aber auch Zahnreste und Teile des postcranialen Skelettes einschließen. Diese Funde wurden der Gattung Dzungariotherium zugewiesen, einem sehr großen Vertreter.[48][49] Vor allem aus der Inneren Mongolei sind in jener Zeit zudem auch die ersten Überreste von Juxia und Urtinotherium entdeckt worden, zwei stammesgeschichtlich ältere Mitglieder der Indricotherien. Eine treibende Kraft hinter diesen Entdeckungen war der chinesische Paläontologe Chow Minchen (1918–1996).[4]
Europäische Funde kamen deutlich später zu Tage und sind insgesamt sehr selten. Bruchstücke von Oberkieferbackenzähnen stammen aus einem Kohlebergwerk südöstlich von Berane in Montenegro und wurden im Jahr 1957 veröffentlicht. Ihre Merkmale sprachen für einen Verweis zu Indricotherium,[50], ebenso wie bei einem letzten Prämolaren des Oberkiefers aus einer Tongrube bei Dragovištiza in Bulgarien.[51] Darüber hinaus kamen im Kreis Cluj in Rumänien einige Oberkieferbackenzähne zu Tage, die in ihrer Erstpublikation 1960 gleichfalls zu Indricotherium gestellt,[52] später aber mit Benaratherium gleichgesetzt wurden.[53][54] Diese Gattung kleinerer Vertreter der Indricotherien hatte fünf Jahre zuvor L. K. Gabunia unter Berufung auf einen rechten Unterkieferast aus Benara in Georgien eingeführt.[11] Ihr können insgesamt aber nur wenige Funde zugewiesen werden, so unter anderem einige Handwurzelknochen wie ein Mondbein, das bereits 1951 von der gleichen Fundstelle berichtet wurde.[55] Aufgrund des extrem geringen Fundmaterials ist die Stellung von Benaratherium aber sehr unsicher.[56]
Eine erste Rekonstruktion erfolgte 1923 von Osborn, die aber nur auf wenigen Funden aus der Mongolei basierte und dadurch einen zu nashornähnlichen Charakter trug mit deutlich zu kurzen Gliedmaßen und zu kurzem Hals. Noch im gleichen Jahr fertigte er eine erfolgreichere Rekonstruktion an, die er anhand umfangreicherer Funde aus der Mongolei und Belutschistans erstellte.[23] Ihr folgte der Rekonstruktionsversuch von Borissiak im Jahr darauf, den er basierend auf den Indricotherium-Funden durchführte und dem er mangels eines vollständigen Schädels den von Baluchitherium hinzufügte. Seine Rekonstruktion wich aber von der Osborns zweiter im Bezug auf die Körperproportionen ein wenig ab, kam dem realen Aussehen von Paraceratherium aber ebenfalls deutlich nahe.[5] Eine sehr umfassende und bis heute bildbestimmende Rekonstruktion ist jene von Granger und Gregory aus dem Jahr 1935, die mit Hilfe der mongolischen Funde erfolgte und der sich auch eine umfangreiche Beschreibung des Fundmaterials anschloss. Es ist dabei zu erwähnen, dass diese Rekonstruktion verschiedenes Fundmaterial einschließt, das von beiden Autoren in vier unterschiedliche Größenklassen eingeteilt wurde, da es von mehreren Individuen mit abweichender Körpergröße stammte.[57][2]
Systematik
Äußere Systematik
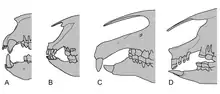
Als ein Gattungsvertreter aus der Überfamilie der Rhinocerotoidea war Paraceratherium mit den heutigen Nashörnern (Rhinocerotidae) verwandt, trug aber selbst keine Hörner. Bei seiner Entdeckung wurde die Tiergattung zunächst von Forster Cooper den Nashörnern zugewiesen,[7] von Borissiak stammt die Bezeichnung der Unterfamilie Indricotheriinae (Indricotherien), die er 1923 einführte und die er damals ebenfalls als Mitglied der Nashörner sah.[43] Im gleichen Jahr schlug Osborn die Bezeichnungen Baluchitheriinae beziehungsweise Paraceratheriinae vor, beide Benennungen haben aber keine Gültigkeit.[14] Im Jahr 1939 allerdings etablierte Borissiak den Familiennamen Indricotheriidae als Schwestertaxon der Nashörner, der in der Folgezeit aber kaum Anerkennung fand.[10] Fast 30 Jahre später wurde die Gruppe der Indricotherien 1967 aufgrund ihres komplexeren und teilweise anders strukturierten vorderen Gebisses aus den Rhinocerotidae ausgeschlossen und den Hyracodontidae zugewiesen. Die Hyracodontidae bildeten zu diesem Zeitpunkt innerhalb der Rhinocerotoidea das Schwestertaxon der Nashörner. Als Unterschied zwischen den beiden Gruppen wurde die Struktur des vorderen Gebisses herangezogen. So besitzen die echten Nashörner in der oberen Zahnreihe ein Paar plattenförmiger Schneidezähne, während im Unterkiefer spitze, dolchförmige Zähne ausgebildet sind (sogenannte „Meißel-Stoßzahn-Anordnung“). Die Hyracodontidae hingegen weisen ein relativ variables Vordergebiss auf. Die Indricotherien ähneln dabei den Nashörnern, zeigen im oberen Gebiss aber ebenfalls dolchartige Schneidezähne.[33] Dass ein am Vorderfuß ausgebildeter rudimentärer vierter Zeh bei basalen Indricotherien eine Stellung innerhalb der echten Nashörner befürwortete,[58] wurde weitgehend abgelehnt, da dieses Merkmal bei vielen urtümlichen Nashornartigen ebenfalls auftritt.[36] Die Ausgliederung der Indricotherien aus den Nashörnern wurde mehrheitlich begrüßt,[35] ist allerdings auch nicht ganz unumstritten.[10] Aufgrund der hohen Variabilität der Hyracodontidae wurde den Indicotherien in den 2010er Jahren ein eigenständiger Familienstatus zugewiesen.[59]
Innerhalb der Indricotheriidae ist die Eigenstellung von Indricotherium Gegenstand einer häufig geführten Diskussion. Während Baluchitherium schon sehr früh als identisch mit Indricotherium erkannt wurde[2] und beide letztendlich 1959 zu Indricotherium zusammengeführt wurden,[11] war die Beziehung zu Paraceratherium lange ungelöst. Bemerkenswert ist der Größenunterschied zwischen dem kleineren Paraceratherium und dem wesentlich größeren Indricotherium. Weitere Unterschiede betreffen die Form des Unterkiefers und des Schädels sowie den Zahnbau und die Zahnanzahl, vor allem bei den Schneidezähnen. So besaß Indricotherium zwei kleinere Schneidezähne zwischen den oberen großen dolchartigen Stoßzähnen, während dies bei Paraceratherium nicht der Fall war.[60] Auch die Stellung der unteren Schneidezähne unterscheidet sich in ihrer anatomischen Ausprägung bei beiden Formen, da jene von Indricotherium deutlich steiler aufgerichtet waren.[8] Trotz des geringen Fossilmaterials gerade dieser Körperpartien wurde deshalb schon in den 1960er und 1970er Jahren diskutiert, ob die bekannten Abweichungen auf unterschiedliche Arten oder Gattungen zurückzuführen sind.[33] Allerdings wurden beide Formen 1989 in einer Revision der Indricotherien durch Spencer G. Lucas und Jay C. Sobus als zu einer Gattung (Paraceratherium) gehörig zusammengeführt und die Unterschiede intraspezifisch als Geschlechtsdimorphismus angesehen, wonach Indricotherium möglicherweise die männliche und Paraceratherium die weibliche Form darstellen. Grundlage dafür waren neben morphologischen und metrischen Zahnvergleichen vor allem mehrere Schädel aus dem Turpan-Becken in China, die allesamt kaum Variationsunterschiede ergaben.[56] Trotzdem befürworten zahlreiche Forscher aufgrund der vorhandenen Unterschiede auf eine Trennung der beiden Formen.[8][20]
Innere Systematik der Familie der Indricotheriidae nach Wang et al. 2016[59]
|
Neben Paraceratherium gehören auch noch Pappaceras, Forstercooperia[61] Juxia[62] und Urtinotherium[63] zu den Indricotherien. Diese unterscheiden sich weitgehend in der höheren Anzahl der Zähne des vorderen Gebisses voneinander und sind allesamt kleiner. Das von Borissiak 1939 basierend auf Schädelfunden aus der Nähe des Aralsees beschriebene Aralotherium wurde ebenfalls 1959 bei der Gesamtbearbeitung des Fundmaterials der Indricotherien mit Paraceratherium gleichgestellt.[11] Ursprünglich wurde mit Dzungariotherium[48] (die Beschreibung basierte auf einem nahezu vollständigen Schädel aus Nordwestchina) noch eine weitere Gattung beschrieben, im Zuge der taxonomischen Überarbeitung 1989 jedoch in Paraceratherium mit eingeschlossen,[56] was aber nicht vollständig akzeptiert wird. Als markanter Unterschied wird unter anderem die Ausdehnung des Naseninnenraums genannt, der bei Dzungariotherium bis zur Orbita reichte, bei Paraceratherium jedoch deutlich davor endete.[8] Auch die Eigenständigkeit von Turpanotherium ist nicht ganz eindeutig, hierbei handelt es sich um eine vergleichsweise kleine Form.[19] Unklar bleibt zudem die taxonomische Stellung von Benaratherium aus Georgien, eingeführt von L. Gabunia 1955,[11] da diese Gattung bei der 1989 erfolgten Revision aufgrund zu weniger Funde nicht stärker berücksichtigt werden konnte.[56][20]
Innere Systematik
Innere Systematik von Paraceratherium nach Deng et al. 2021[19]
|
Zahlreiche unterschiedliche Arten wurden von Paraceratherium beschrieben, aufgrund der Mehrfachbenennung der Gattung überschneiden sich mehrere dieser Formen. So wurden Baluchitherium grangeri, Indricotherium transouralicum und Indricotherium asiaticum bzw. Paraceratherium bugtiense und Baluchitherium osborni bereits früh als synonyme Arten erkannt.[11] Im Folgenden werden die neun heute anerkannten Arten kurz aufgeführt, von denen die ersten vier durch die Revision der Gattung durch Lucas und Sobus 1989 anerkannt sind,[56] die weiteren fünf damals nicht berücksichtigt oder danach neu beschrieben wurden:
- P. bugtiense (Pilgrim, 1908, 1910, 1912), ursprünglich Aceratherium, stellt die Typusart von Paraceratherium dar und war ein kleinerer Vertreter, der etwa 7,4 Tonnen wog. Hauptsächlich nachgewiesen ist die Art in Pakistan.[6][3]
- P. transouralicum (Pavlova, 1922), ursprünglich Baluchitherium und Indricotherium, war eine große Art, Bullen wogen etwa 15 bis 20 Tonnen, Kühe wahrscheinlich nur 9 bis 11 Tonnen. Bedeutende Nachweisgebiete sind Kasachstan und die Mongolei. Neben P. bugtiense ist dies die am besten erforschte Art.[11][3]
- P. prohovori (Borissiak, 1939), ursprünglich Aralotherium, war ein kleinerer Vertreter, der ein Gewicht von rund 9 Tonnen aufwies und hauptsächlich im östlichen Kasachstan nachgewiesen wurde. Die Art besaß ein höheres Schädeldach und weicht mit einzelnen anatomischen Unterschieden im Unterkiefer ab.[3][8]
- P. orgosense (Qiu, 1973), ursprünglich Dzungariotherium, war eine große Art und erreichte etwa die Ausmaße von P. transouralicum, möglicherweise aber auch größere, da die Zähne bis zu 25 % größer sind als bei P. transouralicum, Gewichtsangaben gehen bei großen Individuen bis zu 24 Tonnen.[4][64] Sie ist unter anderem anhand eines nahezu vollständigen, 121 cm langen Schädelfundes aus dem Dsungarischen Becken in Xinjiang im Nordwesten Chinas bekannt und besaß sowohl im Ober- als auch im Unterkiefer jeweils ein Schneidezahnpaar, das deutlich in der Länge reduziert war, weiterhin seitlich verbreiterte Prämolaren und ein nahezu nicht vorhandenes Diastema.[48]
- P. zhajremensis Bayshashov, 1988 war eine große Art, möglicherweise größer als P. transouralicum. Sie ist durch mehrere Unterkieferfragmente aus dem Südosten Kasachstans bekannt, die morphologisch von den anderen Arten abweichen, vor allem im extrem tiefen Knochenkörper des Unterkiefers unterhalb des letzten Molaren.[16]
- P. sui Ye, Meng & Wu, 2003 stellte eine kleinere Art dar, war jedoch größer als P. bugtiense. Der bisher singuläre Unterkiefer aus Fuhai in Nordwestchina ähnelte stark dem der Typusart; die Symphyse des Unterkiefers ist aber deutlich nach unten gebogen.[8] Teilweise wird die Art auch zu Aralotherium gestellt.[9]
- P. yagouense Qiu, Wang & Deng, 2004 war eine kleine Art und in der Größe mit P. bugtiense vergleichbar. Sie unterscheidet sich von anderen Arten durch die Reduktion der oberen Schneide- und Eckzähne und höherkronigen Backenzähnen, allerdings beruht die Beschreibung auf einem Schädel eines nicht ausgewachsenen Tieres, der im Linxia-Becken in Gansu im Norden Chinas gefunden wurde.[65] Es besteht aber auch die Möglichkeit, dass P. yagouense zur 2007 neu eingeführten Gattung Turpanotherium gehört,[66] für die eine eventuell nähere Verwandtschaft mit Urtinotherium erwogen wird.[4]
- P. huangheense Li, Zhang, Li, Li & Xie, 2017, war eine große Art. Beschrieben wurde sie über Reste des Ober- und Unterkiefers aus dem Lanzhou-Becken in der chinesischen Provinz Gansu, nahe dem Gelben Fluss, worauf sich auch der Artname bezieht. Die Funde datieren in das Untere Oligozän und sind rund 31,5 Millionen Jahre alt. Die Art ähnelt P. bugtiense, war aber deutlich größer (Länge der unteren Backenzahnreihe 41,5 cm im Vergleich zu 32,5 cm). Unterschiede zu andern Formen bestehen vor allem in Details der Unterkiefer- und Zahnmorphologie.[67]
- P. linxiaense Deng, Lu, Wang, Flynn, Sun, He & Chen, 2021 repräsentiert eine große Form, die anhand eines 116 cm langen Schädels samt einem 82 cm langen Unterkiefer sowie einigen Teilen des Körperskeletts aus dem Linxia-Becken in Gansu beschrieben wurde. Das Alter der Funde liegt bei rund 26,5 Millionen Jahren. Die Beschreiber der Art sehen sie als vergleichsweise modern an, die sich unter anderem durch einen tieferen Naseninnenraum und ein höheres Hinterhaupt von älteren Formen absetzt.[19]
Chinesische Paläontologen unterscheiden darüber hinaus drei weitere Arten.
- P. tienshanense Chiu, 1962 war eine große Art, möglicherweise so groß wie P. transouralicum oder größer, ist aber nur von einem fragmentierten Unterkiefer bekannt, der aus dem Hami-Becken in Xinjiang stammt.[8][65] Der Unterkiefer wurde bereits 1959 vorgestellt[68] und drei Jahre später der neuen Art zugewiesen.[69] Mitunter wird die Art auch in der Gattung Dzungariotherium geführt.[9]
- P. lepidum Xu & Wang, 1978 war eine kleinere Art und erreichte die Maße von P. bugtiense, ist bisher aber nur von einem Schädelfund aus der Turpan-Senke in Xinjiang bekannt. Der Fund war assoziiert mit einer vollständigen Hals- und Brustwirbelsäule, die ein Tier von insgesamt 7,2 m Länge rekonstruieren lassen.[8][65][9]
- P. turfanense (Xu & Wang, 1978), stellt eine kleinere Art dar, die auch zu Dzungariotherium gestellt wird. Das Fundmaterial umfasst einen stark fragmentierten vorderen Schädel, einzelne Unterkiefer und Reste des postcranialen Skelettes aus der Turpan-Senke.[9]
Literatur
- Tao Deng, Xiaokang Lu, Shiqi Wang, Lawrence J. Flynn, Danhui Sun, Wen He und Shanqin Chen: An Oligocene giant rhino provides insights into Paraceratherium evolution. Communications Biology 4, 2021, S. 639, doi:10.1038/s42003-021-02170-6
- Walter W. Granger und William K. Gregory: Further notes on the gigantic extinct rhinoceros, Baluchitherium, from the Oligocene of Mongolia. Bulletin of the American Museum of Natural History 72, 1936, S. 1–73.
- Donald R. Prothero: Rhino giants: The palaeobiology of Indricotheres. Indiana University Press, 2013, ISBN 978-0-253-00819-0, S. 1–141.
- Zhan-Xiang Qiu und Ban-Yue Wang: Paracerathere fossils of China. Palaeontologia Sinica 193 (New Series C, 29), 2007, S. 1–396 (S. 247–386 in Englisch).
Einzelnachweise
- Michael P. Taylor und Mathew J. Wedel: Why sauropods had long necks; and why giraffes have short necks. PeerJ 1, 2013, S. e36.
- Walter W. Granger und William K. Gregory: Further notes on the gigantic extinct rhinoceros, Baluchitherium, from the Oligocene of Mongolia. Bulletin of the American Museum of Natural History 72, 1936, S. 1–73.
- Mikael Fortelius und John Kappelmann The largest land mammal ever imagined. Zoological Journal of the Linnean Society 107, 1993, S. 85–101.
- Donald R. Prothero: Rhino giants: The palaeobiology of Indricotheres. Indiana University Press, 2013, S. 1–141 ISBN 978-0-253-00819-0.
- Alexei A. Borissiak: Über die Unterfamilie Indricotheriinae Boriss. = Baluchitheriinae Osb. Zentralblatt für Mineralogie, Geologie und Paläontologie 18, 1924, S. 571–575.
- Clive Forster-Cooper: On the skull and dentition of Paraceratherium bugtiense: a genus of aberrant rhinoceros from the lower Miocene deposits of Dera Bugti. Philosophical Transactions of the Royal Society of London (B) 212, 1924, S. 369–394.
- Clive Forster-Cooper: Paraceratherium bugtiense, a new Genus of Rhinocerotidae from the Bugti Hills of Baluchistan - preliminary notice. The Anals and Magazine of Natural History 8, 1911, S. 711–716.
- Ye Jie, Meng Jin und Wu Wen: Discovery of Paraceratherium in the northern Junggar Basin of Xinjiang. Vertebrata Palasiatica 41 (3), 2003, S. 220–229.
- Zhan-Xiang Qiu und Ban-Yue Wang: Paracerathere fossils of China. Palaeontologia Sinica 193 (New Series C, 29), 2007, S. 1–396 (S. 247–386 in englisch).
- Demberelyin Dashzeveg: A new Hyracodontid (Perissodactyla, Rhinocerotoidea) from the Ergilin Dzo formation (Oligocene Quarry 1) in Dzamyn Ude, Eastern Gobi Desert, Mongolia. American Museum Novitates 3178, 1996, S. 1–12.
- Вера Громова: Гигантские носороги. Академия Наук СССР 71, Moskau 1959.
- Pierre-Olivier Antoine, S. M. Ibrahim Shah, Iqbal U. Cheema, Jean-Yves Crochet, Dario de Franceschi, Laurent Marivaux, Grégoire Métais and Jean-Loup Welcomme: New remains of the baluchithere Paraceratherium bugtiense (Pilgrim, 1910) from the Late/latest Oligocene of the Bugti hills, Balochistan, Pakistan. Journal of Asian Earth Sciences 24 (1), 2004, S. 71–77, doi:10.1016/j.jaes.2003.09.005.
- P. Martin Sander, Andreas Christian, Marcus Clauss, Regina Fechner, Carole T. Gee, Eva-Maria Griebeler, Hanns-Christian Gunga, J¨urgen Hummel, Heinrich Mallison, Steven F. Perry, Holger Preuschoft, Oliver W. M. Rauhut, Kristian Remes, Thomas T¨utken, Oliver Wings und Ulrich Witzel: Biology of the sauropod dinosaurs: the evolution of gigantism. Biological Reviews 86, 2011, S. 117–155.
- Henry Fairfield Osborn: Baluchitherium grangeri, a giant hornless rhinoceros from Mongolia. American Museum Novitates 78, 1923, S. 1–15.
- Алексей Алексеевич Борисяк: Остеология индрикотерия. Исвестя Императорскоӣ Академиӣ Наук 11, 1917, S. 287–299.
- Spencer George Lucas und Bolat U. Bayshashov: The giant rhinoceros Paraceratherium from the Late Oligocene at Aktau Mountain, southern Kazakhstan, and its biochronological significance. Neues Jahrbuch für Geologie und Paläontologie, Stuttgart 9, 1996, S. 539–548.
- Болат У. Байшашов: О гигантском носороге местонарождения Кызылжар (юго-восточныи Казахстан). Хабаршы Вестник 40 (1), 2009, S. 50–53.
- Chiu Chan-Siang: Giant rhinoceros from Loping, Yunnan, and discussion of the taxonomic characters of Indricotherium grangeri. Vertebrata Palasiatica 6 (1), 1962, S. 57–71.
- Tao Deng, Xiaokang Lu, Shiqi Wang, Lawrence J. Flynn, Danhui Sun, Wen He und Shanqin Chen: An Oligocene giant rhino provides insights into Paraceratherium evolution. Communications Biology 4, 2021, S. 639, doi:10.1038/s42003-021-02170-6.
- Pierre-Olivier Antoine, Levent Karadenizli, Gerçek Saraç und Sevket Sen: A giant rhinocerotoid (Mammalia, Perissodactyla) from the late Oligocene of north-central Anatolia (Turkey). Zoological Journal of the Linnean Society 152 (3), 2008, S. 581–592.
- Sevket Sen, Pierre-Olivier Antoine, Bakit Varol, Turban Ayyiidiz und Koray Sozeri: Giant rhinoceros Paraceratherium and other vertebrates from Oligocene and middle Miocene deposits of the Kağızman-Tuzluca Basin, Eastern Turkey. Naturwissenschaften 98 (5), 2011, S. 407–423.
- Vlad A. Codrea: Rinoceri și Tapiri Terțiari din România. Presa Universitara Clujeana, Cluj-Napoca, 2000, Französischer Abstract S. 145–147.
- Henry Fairfield Osborn: The extinct giant rhinoceros Baluchitherium of Western and Central Asia. Natural History, New York 23 (3), 1923, S. 208–228.
- Philip D. Gingerich: Predictions of Body by Browse to Save">Mass in mammalian species from bone lengths and diameters. Contributions from the Museum of Paleontology, University of Michigan, Ann Arbor 28 (4), 1990, S. 79–92.
- Per Christiansen: Body size in proboscideans, with notes on elephant metabolism. Zoological Journal of the Linnean Society 140, 2004, S. 523–549.
- Asier Larramendi: Shoulder height, body mass, and shape of proboscideans. Acta Palaeontologica Polonica 61 (3), 2016, S. 537–574.
- M. Clauss R. Frey, B. Kiefer, M. Lechner-Doll,·W. Loehlein,·C. Polster, G. E. Rössner·und W. J. Streich: The maximum attainable body size of herbivorous mammals: morphophysiological constraints on foregut, and adaptations of hindgut fermenters. Oecologia 136, 2003, S. 14–27.
- Pierre-Olivier Antoine: There were giants upon the earth in those days. Review of Prothero, Rhinoceros giants. Palaeovertebrata 38 (1), 2014, S. 1–3.
- C. Martin, I. Bentaleb und P.-O. Antoine: Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 2011, S. 19–29.
- Yang Wang und Tao Deng: A 25 m.y. isotopic record of paleodiet and environmental change from fossil mammals and paleosols from the NE margin of the Tibetan Plateau. Earth and Planetary Science Letters 236, 2005, S. 322–338.
- Luke T. Holbrook und Spencer George Lucas: A new genus of rhinocerotoid from the Eocene of Utah and the status of North American "Forstercooperia. Journal of Vertebrate Paleontology 17 (2), 1997, S. 384–396.
- Bruce J. MacFadden: Fossil Horses. Systematic, Paleobiology, and Evolution of the family Equidae. Cambridge University Press, 1992.
- Leonard B. Radinsky: A review of the Rhinocerotoid Family Hyracodontidae (Perissodactyla). Bulletin of the American Museum of Natural History 136 (1), 1967, S. 1–47.
- Bolat Uapovich Bayshashov und Spencer George Lucas: The giant rhinoceros Urtinotherium from the Upper Eocene of the Zaisan basin, Kazakhstan. Selevinia (Kazachstansky zoologichesky churnal – The Zoological Journal of Kazakhstan) 1–4, 2001, S. 185–187.
- Donald R. Prothero, Earl Manning und C. Bruce Hanson: The phylogeny of the rhinocerotoidea (Mammalia, Perissodactyla). Zoological Journal of the Linnean Society 87, 1986, S. 341–366.
- Donald R. Prothero, Claude Guérin und Earl Manning: The history of Rhinocerotoidea. In Donald R. Prothero und R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New-York, London, Oxford University Press, 1989, S. 321–340.
- Spencer George Lucas, Bolat U. Bayshashov, Lyubov A. Tyut'kova, Ayzhan K. Dzhamangaraeva und Bolat Zh. Aubekerov: Mammalian biochronology of the Paleogene-Neogene boundary at Aktau Mountain, Eastern Kazakhstan. Paläontologische Zeitschrift 71 (3–4), 1997, S. 305–314.
- P. V. Putshkov: “Proboscidean agent” of some Tertiary megafaunal extinctions. In: G. Cavarretta et al. (Hrsg.): The World of Elephants – International Congress. Consiglio Nazionale delle Ricerche. Rom, 2001, S. 611–616.
- Guy E. Pilgrim: The vertebrate fauna of the Gaj Series in the Bugti Hills and the Punjab. Memoirs of the Geological Survey of India (Palaeontologia Indica) New Series 4 (2), 1912, S. 1–6.
- Othenio Abel: Über die Entdeckung eines riesigen Säugetiers im unteren Miozän Asiens. Die Naturwissenschaften 15 (13. 4.), 1923, S. 284–286.
- Jean-Loup Welcomme, Mouloud Benammi, Jean-Yves Crochet, Laurent Marivaux, Gregoire Metais, Pierre-Olivier Antoine und Ibrahim Baloch: Himalayan Forelands: palaeontological evidence for Oligocene detrital deposits in the Bugti Hills (Balochistan, Pakistan). Geological Magazine 138 (4), 2001, S. 397–405.
- Алексей Алексеевич Борисяк: О зубномъ аппаратъ индрикотерия. Исвестя Императорскоӣ Академиӣ Наук 10, 1916, S. 343–350.
- Алексей Алексеевич Борисяк: О роде Indricotherium n. g. (сем. Rhinocerotidae). Записки Российской Академик Наукъ 35 (8), 1923, S. 1–128.
- M. Pavlova: Indricotherium transouralicum n. sp. provenant du district de Tourgay. Bulletin de la Societe des Naturalistes de Moscou, Section Geologique 31, 1922, 95–116.
- Othenio Abel: Weitere Entdeckungen von Resten des riesenhaften Rhinocerotiden Baluchitherium im Tertiär Innerasiens. Die Naturwissenschaften 1 (4. 1.), 1924, S. 14–17.
- Clive Forster-Cooper: Thaumastotherium osborni, a new genus of Perissodactyles from the Upper Oligocene deposits of the Bugti Hills of Baluchistan: preliminary notice. Annals and Magazine of Natural History (8) 12, 1913, S. 376–381.
- Clive Forster-Cooper: Correction of generic name Baluchitherium. Annals and Magazine of Natural History (B) 1, 1913, S. 504.
- Qiu Zhan-Xiang: A new genus of giant rhinoceros from oligocene of Dzungaria, Sinkang. Vertebrata Palasiatica 11 (2), 1973, S. 182–191.
- Qi Tao: A new species of Dzungariotherium (Perissodactyla, Mammalia). Vertebrata Palasiatica 27 (4), 1989, S. 301–305.
- Ž. Petronijević und Erich Thenius: Über den ersten Nachweis von Indricotherien (Baluchitherien, Rhinocerotidae, Mammalia) im Tertiär von Europa. Anzeiger der Mathematischen und Naturwissenschaftlichen Klasse, Österreichische Akademie der Wissenschaften (Wien) 9, 1957, S. 153–155.
- Ivan Nikolov und Kurt Heissig: Fossile Säugetiere aus dem Obereozän und Unteroligozän Bulgariens und ihre Bedeutung für die Paläogeographie. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 25, 1985, S. 61–70.
- Л. К. Габуня und О. Илиеску: О первой находке остатков гигантских носорогов из сем. Indricotheriidae в Румынии. Доклады Академии Наук СССР 130 (2), 1960, S. 425–427.
- L. K. Gabunia: Sur les mammiferes oligocenes du Caucase. Bulletin de la Societe Geologique de France 7, 1966, S. 857–869.
- Vlad A. Codrea und Nicolae Şuraru: New remains of indricotheriin (Perissodactyla, Mammalia) in the Lower Oligocene at Fildu de Jos (Sălaj District, Nw Transylvania). Romanian Journal of Paleontology 76, 1995, S. 81–87.
- Л. К. Габуня: О представителях семейства Indricotheriidae из Олигоценовых отложений Грузии. Доклады Академии Наук СССР 81 (6), 1951, S. 1135–1137.
- Spencer George Lucas und Jay C. Sobus: The systematics of Indricotheres. In: Donald R. Prothero und R. Schoch (Hrsg.): The evolution of Perissodactyls. New York: Oxford Univ. Press., 1989, S. 358–378.
- Walter W. Granger und William K. Gregory: A revised restoration of the skeleton of Baluchitherium, gigantic fossil rhinoceros of Central Asia. American Museum Novitates 787, 1935, S. 1–3.
- Kurt Heissig: The rhinocerotidae. In: Donald R. Prothero und R. M. Schoch (Hrsg.): The evolution of perissodactyls. New York, London, Oxford University Press, 1989, S. 399–417.
- Haibing Wang, Bin Bai, Jin Meng und Yuanqing Wang: Earliest known unequivocal rhinocerotoid sheds new light on the origin of Giant Rhinos and phylogeny of early rhinocerotoids. Scientific Reports 6, 2016, S. 39607, doi:10.1038/srep39607.
- Clive Forster-Cooper: The extinct Rhinoceroses of Baluchistan. Philosophical Transactions of the Royal Society of London (B) 223, 1934, S. 569–616.
- Horace Elmer Wood: Cooperia totadentata, a remarkable rhinoceros from the eocene of Mongolia. American Museum Novitates 1012, 1938, S. 1–22.
- Chow Minchen und Chiu Chan-Siang: An eocene giant rhinoceros Juxia sharamurenense. Vertebrata Palasiatica 8 (3), 1964, S. 264–268.
- Chow Minchen und Chiu Chan-Siang: New genus of giant rhinoceros from oligocene of inner Mongolia Urtinotherium incisivum. Vertebrata Palasiatica 7 (3), 1963, S. 230–239.
- Tao Deng: Late Cenozoic environmental changes in the Linxia basin (Gansu, China) as indicated by cenograms of fossil Mammals. Vertebrata Palasiatica 47 (4), 2009, S. 282–298.
- Qiu Zhan-Xiang, Wang Ban-Yue und Deng Tao: Indricothere (Perissodactyla, Mammalia) from Oligocene in Linxia Basin, Gansu, China. Vertebrata Palasiatica 42 (3), 2004, S. 177–192.
- Deng Tao, Qui Zhan-Xiang, Wang Ban-Yue, Wang Xiao-Ming und Hou Su-Kuan: Late Cenozoic biostratigraphy of the Linxia Basin, Northwestern China. In: Wang Xiaoming, Lawrence J. Flynn und Mikael Fortelius (Hrsg.): Fossil Mammals of Asia. Neogene biostratigraphy and chronology. Columbia University Press, New York, 2013, S. 243–270.
- Li Yong-Xiang, Zhang Yun-Xiang, Li Ji, Li Zhi-Chao und Xie Kun: New fossils of paraceratheres (Perissodactyla, Mammalia) from the Early Oligocene of the Lanzhou Basin, Gansu Province, China. Vertebrata Palasiatica 56 (4), 2017, S. 367–381, doi:10.19615/j.cnki.1000-3118.170922.
- Chow Minchen und Xu Yuxuan: Indricotherium from Hami Basin, Sinkiang. Vertebrata Palasiatica 3 (2), 1959, S. 93–98.
- Qiu Zhan-Xiang: Giant rhinoceros from Loping, Yunnan, and discussion of the taxonomic characters of Indricotherium grangeri. Vertebrata Palasiatica 6 (1), 1962, S. 58–71.
Weblinks
- The Paleobiology Database: Paraceratherium