Mammute
Die Mammute (Sing. das Mammut; Pl.: auch Mammuts), wissenschaftlicher Name Mammuthus (von frz. mammouth < russ.: мамонт mamont < vermutlich aus dem Waldnenzischen), bilden eine ausgestorbene Gattung der Elefanten. Sie entstand im Übergang vom Miozän zum Pliozän in Afrika und besiedelte in der darauf folgenden Zeit sowohl Europa als auch Asien und Nordamerika. Die letzten Vertreter der Mammute, die der weitaus bekanntesten Art, dem Wollhaarmammut (M. primigenius) angehören, starben erst vor rund 4000 Jahren auf der nordsibirischen Wrangelinsel aus.
| Mammute | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Lebendrekonstruktion eines Wollhaarmammuts im kanadischen Royal British Columbia Museum | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Pliozän bis Holozän | ||||||||||||
| 5,7 Mio. Jahre bis etwa 4.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Mammuthus | ||||||||||||
| Brookes, 1828 | ||||||||||||

Im November 2008 wurde in der Fachzeitschrift Nature die Genomsequenz des Wollhaarmammuts veröffentlicht.[1] Circa 70 Prozent der Erbinformation konnten entschlüsselt werden. Das Mammutgenom ist das erste Genom eines ausgestorbenen Tieres, das sequenziert wurde.
Lebensraum
Vor allem die während der letzten Kaltzeit lebenden Vertreter der Mammute (Mammuthus primigenius) waren an den Lebensraum der Kaltsteppe gebunden. Die üblicherweise als Mammutsteppe bezeichnete Vegetationsform besteht nur zu etwa 20 Prozent aus Gräsern, weitaus dominanter sind besonders proteinreiche Kräuter und Blüten wie Wegeriche, Beifuß, Schafgarbe, Chrysanthemen, Kuhschellen, Silberwurz und sogar einige Weiden.[2] Diese Tundra-ähnliche Kaltsteppe bildete mit den nahrhaften Kräutern die Grundlage zur Ernährung von Herden großer Landsäugetiere. Sie erstreckte sich ursprünglich vom westlichen Europa über die osteuropäische Tiefebene, das nördliche Sibirien bis nach Alaska und schloss auch einige damals trockengelegene Schelfgebiete ein, wie die Nordsee (Doggerland) und die Beringstraße. Neben dem Mammut gehörten zur damaligen Tierwelt die Saigaantilope, das Mufflon, der Steinbock, verschiedene Hirschartige (Rothirsch, Riesenhirsch, Elch, Ren) und Wildrinder (Wisent und Bison), aber auch das Wollnashorn, größere und kleinere Raubtiere wie Großkatzen (Schneeleopard, Höhlenlöwe), Bären (Höhlenbär, Braunbär) und Hunde (Wolf, Fuchs) und darüber hinaus zahlreiche Kleinsäuger zum üblichen Bild der Mammutsteppe. Nachfahren der Tierarten dieses Biotops leben heute vorwiegend in Hochgebirgen und arktischen Regionen.[3]
Dieser Lebensraum wurde außerdem von Populationen der frühen Jäger und Sammler bewohnt und genutzt.[3]
Merkmale
Allgemein und Körpergröße
Mammute waren große bis sehr große Säugetiere, die einen generell elefantenähnlichen Körperbau mit großem Kopf und säulenartigen Beinen aufwiesen. Die Größe war von Art zu Art unterschiedlich, das bekannte Wollhaarmammut (Mammuthus primigenius), das Endglied der Mammutentwicklung aus dem Mittleren und Oberen Pleistozän, erreichte eine Schulterhöhe von 2,8 bis 3,7 m und entsprach damit jener der heute lebenden Elefanten, das Gewicht variierte von 5 bis 8 t. Andere Arten konnten aber durchaus größer werden, so erreichten das Präriemammut (Mammuthus columbi) aus dem Pleistozän Nordamerikas und der Südelefant (Mammuthus meridionalis) aus dem Altpleistozän Eurasiens bis zu 4,2 m Schulterhöhe und ein Gewicht von gut 12 t. Der größte Vertreter der Mammute und eines der größten bekannten Rüsseltiere überhaupt war jedoch das Steppenmammut (Mammuthus trogontherii), das im Alt- und Mittelpleistozän in Eurasien lebte. Es wurde bis zu 4,5 m hoch, sein geschätztes Gewicht betrug 15 t.[4][5]
Innerhalb der Mammute kam es mehrfach durch Abtrennung kleinerer Gruppen von der Hauptpopulation der einzelnen Arten auf Inseln zur Ausbildung von Zwergformen. Nachgewiesen wurden Zwergformen auf Sardinien und Kreta, die vom Südelefanten abstammen. Eine Extremform ist dabei das Kreta-Zwergmammut (Mammuthus creticus), das nur etwa 1,1 m hoch und rund 310 kg schwer wurde.[6]
Schädel- und Gebissmerkmale
Charakteristisch für die Mammute waren kurze hohe Schädel, noch ausgeprägter als bei Elephas, die im Laufe ihrer Entwicklung immer höher wurden.[7] Die Stirnlinie verlief vorn steil und war abweichend von Loxodonta stark eingedellt. Das Stirnbein war nach hinten ausgezogen. Stirn- und Scheitelbein besaßen von vorn gesehen eine gewölbte Gestaltung, wobei eine sehr tiefe mittlere Eintiefung wie bei Elephas fehlte. Auch das Hinterhauptsbein war stark verlängert und wies tief liegende Gelenkansätze für die Halswirbelsäule auf, die nahezu auf der Höhe des Gaumenbeins lagen. Weitere besondere Charakteristika der Mammutschädel stellten die weit auseinander liegenden Augenfenster und vor allem im Gegensatz zu den rezenten Elefanten sehr eng stehenden, fast parallel verlaufenden Alveolen der oberen Stoßzähne. Die Knochen des Schädeldaches waren wie bei den heutigen Elefanten luftgefüllt, einerseits um das Gewicht zu reduzieren, andererseits um durch die damit verbundene Oberflächenvergrößerung eine größere Ansatzstelle für die massive Nacken- und Kiefermuskulatur zu gewährleisten.[8][9]
Das Gebiss war vergleichbar zu dem der rezenten Elefanten und bestand aus einem Paar Stoßzähne in der oberen Zahnreihe, die sich stammesgeschichtlich durch Hypertrophie aus den jeweils zweiten Schneidezähnen gebildet hatten, und aus drei Molaren je Kieferhälfte. Das Milchgebiss besaß zudem noch jeweils drei Prämolaren. Die Stoßzähne waren lang und deutlich gebogen, wobei die Krümmung im Laufe der Stammesgeschichte immer stärker wurde und in spiralartig geformte Stoßzähne mündete. Der längste bekannte Stoßzahn maß über die Krümmung 4,9 m und stammt von einem Präriemammut aus Post im US-Bundesstaat Texas. Die Stoßzähne steckten zu etwa einem Viertel ihrer Länge in den Alveolen.[5]
Das weitere Gebiss umfasste jeweils einen funktionalen Backenzahn je Kieferhälfte, der insgesamt fünfmal nach Abnutzung ausgetauscht werden konnte, also sechs Generationen umfasste. Dieser horizontale Zahnwechsel ist typisch für Elefanten und unterscheidet sich deutlich vom vertikalen Zahnwechsel der meisten Säugetiere. Generell waren die Backenzähne hochkronig (hypsodont), wobei die Hochkronigkeit im Laufe der Entwicklung zunahm. So überstieg die Höhe der Backenzähne später Mammutvertreter deren Breite teilweise um das Doppelte. Ein bedeutendes Merkmal stellte der lamellenartige Aufbau der Backenzähne dar, der durch enge Schmelzfalten erreicht wurde.[8] Die Anzahl der Lamellen der einzelnen Backenzähne, in der Regel der dritte und letzte Molar, hat taxonomischen Wert für die Bestimmung der Mammutarten. Die Anzahl der Schmelzfalten nahm im Laufe der Evolution der Mammute stark zu. Die frühesten afrikanischen Mammutformen wie Mammuthus subplanifrons oder Mammuthus africanavus besaßen 7 bis 9 beziehungsweise durchschnittlich 9 Schmelzfalten. Wies der Südelefant als erster eurasischer Mammutvertreter 13 bis 18 Schmelzlamellen auf, hatte das Steppenmammut bereits zwischen 17 und 23. Das Wollhaarmammut letztendlich besaß 21 bis 30 Schmelzlamellen. Diese späten Mammutformen stellen aufgrund der Morphologie der Backenzähne die am stärksten spezialisierten Elefanten überhaupt dar. Die Zunahme der Schmelzfalten ist ein Anzeichen für eine stärkere Anpassung an offene Landschaftsverhältnisse und eine steigende Spezialisierung auf die daraus resultierende Grasnahrung. Einher ging die Zunahme der Faltenanzahl mit einer Verdünnung des jeweiligen Zahnschmelzbandes der einzelnen Falten von 4 bis 5 auf 1 bis 2 mm.[8][10]
Systematik
Die Mammute haben sich im Übergang vom späteren Miozän zum frühen Pliozän in Afrika entwickelt und verbreiteten sich von dort aus bis Eurasien und Nordamerika.[8] Dabei spezialisierten sie sich zunehmend auf Grasnahrung und entwickelten Anpassungen an die Kälte. Laut molekulargenetischen Untersuchungen hatten sie sich bereits vor 6,7 Millionen Jahren von der Linie, die zum Asiatischen Elefanten (Elephas maximus) führte, abgetrennt.[11] Die ältesten bisher gefundenen Fossilreste von Mammuten sind etwa 5,7 Millionen Jahre alt und stammen aus der Landsenke von Afar in Äthiopien,[12] mit 5 Millionen Jahren nur wenig jünger sind Funde aus Langebaanweg in Südafrika. Diese Funde gehören der Art Mammuthus subplanifrons an. Der Übergang vom Miozän zum Pliozän zeichnete sich durch eine hohe Vielfalt an verschiedenen Rüsseltierformen in Afrika aus, zudem fällt in diese Zeit die Herausbildung der modernen Elefanten wie Loxodonta, Elephas oder eben Mammuthus. Problematisch bei diesen frühen Elefantenfunden ist dabei, dass diese überwiegend aus fragmentierten Zähnen und Gebissresten bestehen, zusammengehöriges Schädelmaterial ist selten. Die Zähne bestehen meist aus einer geringen Anzahl an Lamellen mit einer dicken Zahnschmelzschicht, durch diese ähnliche Gestaltung können noch Unterscheidungsschwierigkeiten zwischen den einzelnen frühen Formen auftreten.[8][13] Mammuthus subplanifrons überlebte sicher bis vor etwa drei Millionen Jahren. Etwa in diesem Zeitraum tauchte sein vermutlicher direkter Nachfolger Mammuthus africanavus in Nordafrika auf, der teilweise auch als das erste eindeutige Mitglied der Mammute angesehen wird. Möglicherweise war diese Art der Vorfahre des Südelefanten (Mammuthus meridionalis), der auch der erste in Eurasien nachgewiesene Mammutvertreter ist. Aus dem Südelefanten entwickelte sich vor etwa 750.000 Jahren das Steppenmammut (Mammuthus trogontherii), das sich über das nördliche Eurasien verbreitete und zum Vorfahren des Wollhaarmammuts wurde. Das Präriemammut (Mammuthus columbi) Nordamerikas hat sich wahrscheinlich ebenfalls aus dem Südelefanten entwickelt, der vor etwa 1,5 Millionen Jahren nach Amerika eingewandert ist. Einige Experten sehen auch einen Ursprung des Präriemammuts im Steppenmammut, welches bisher aber noch nicht in Nordamerika nachgewiesen ist. Das Präriemammut trat erstmals im Altpleistozän vor rund 1,2 Millionen Jahren auf.[14] Es bildet den Grundstock für einen Teil der amerikanischen Mammutpopulation. Im Jungpleistozän besiedelte dann auch das Wollhaarmammut Nordamerika, es bevorzugte aber gegenüber dem Präriemammut eher nördlichere Regionen. Wahrscheinlich kam es aber im Kontaktgebiet, etwa im nördlichen Bereich der heutigen USA, teilweise zur Vermischung der beiden Arten, da molekulargenetische Untersuchungen des Präriemammuts in dessen Genom einzelne Haplotypen des Wollhaarmammuts nachweisen konnten. Die Ursachen sind nicht vollständig geklärt, eventuell spielen Introgressions-Prozesse eine Rolle, wie sie vergleichbar auch zwischen dem Afrikanischen Elefanten (Loxodonta africana) und dem Waldelefanten (Loxodonta cyclotis) bekannt sind.[15]
Folgende Arten sind heute anerkannt:[16]
- Mammuthus subplanifrons (Osborn, 1928); Pliozän; Ost- und Zentralafrika
- Mammuthus africanavus (Arambourg, 1952); Pliozän bis Altpleistozän; Nordafrika
- Mammuthus meridionalis (Nesti, 1825) (Südelefant; einschließlich Mammuthus rumanus und Mammuthus gromovi); Pliozän bis Altpleistozän; Europa, Russland
- Mammuthus lamarmorai (Major, 1883); Jungpleistozän; Südeuropa
- Mammuthus creticus (Bate, 1907) (Kreta-Zwergmammut); Altpleistozän; Südeuropa
- Mammuthus trogontherii (Pohlig, 1885) (Steppenmammut); Alt- bis Mittelpleistozän; Eurasien
- Mammuthus primigenius (Blumenbach, 1799) (Wollhaarmammut); Mittelpleistozän Mittelholozän; Eurasien, Nordamerika
- Mammuthus columbi (Falconer, 1857) (Präriemammut; einschließlich Mammuthus hayi, Mammuthus imperator und Mammuthus jeffersoni); Alt- bis Jungpleistozän; Nord- und Zentralamerika
- Mammuthus exilis (Stock & Furlong, 1928) (Zwergmammut oder Kalifornisches Zwergmammut); Mittel- bis Jungpleistozän; Nordamerika
Im Allgemeinen meint man mit „Mammut“ das während der letzten Eiszeit in Eurasien und Nordamerika verbreitete Wollhaarmammut (Mammuthus primigenius). Irreführend ist der Umstand, dass der Gattungsname Mammut nicht etwa die Mammute bezeichnet, sondern einen mit diesen und den Elefanten nur entfernt verwandten Vertreter der Mammutiden (Mammutidae), einer urtümlicheren Gruppe von Rüsseltieren mit teilweise vier Stoßzähnen, die in der letzten Eiszeit ebenfalls behaarte Formen entwickelte. Beide Entwicklungslinien trennten sich bereits im Oberen Oligozän.[8] Aus forschungsgeschichtlicher Sicht ist anzumerken, dass der Gattungsname Mammut Blumenbach, 1799 für das Mitglied der heutigen Mammutidae wissenschaftlich eher eingeführt wurde als Mammuthus Brookes, 1828 für die bekannteren Mammute als Vertreter der Elefanten.[17]
Mammut und Mensch
Das Wollhaarmammut war eines der Jagdtiere der Menschen im Jungpleistozän, Knochen und Elfenbein dienten außerdem als Materialressourcen.[18] Dies ist durch zahlreiche Höhlenmalereien und eine Vielzahl von Mammutknochen-Anhäufungen in archäologischen Fundstellen des Aurignacien, Gravettien und Epigravettien dokumentiert. Bei der Fundstelle Judinowo in der Osteuropäischen Ebene[19] und auf der arktischen Kotelny-Insel wurden Hinweise gefunden, dass dort Mammuts von Menschen zumindest geschlachtet wurden.[20] Mammutknochenhäuser fanden sich in Meschyritsch, Mesyn, Dobranitschewka und dem Kiewer Kiew-Kyrill-Wohnplatz, (alle Ukraine) aus der Zeit des osteuropäischen Epigravettiens (entspricht zeitlich etwa dem Magdalénien Mitteleuropas)[21] – Mezyn ist auf 16.000 v. Chr. zu datieren.[22]
Aussterben
Die Diskussion, inwieweit Bejagung durch den Menschen im Sinne der „Overkill-Hypothese“ (siehe: Mensch und Präriemammut) zum Aussterben der eurasischen und nordamerikanischen Mammute beitrug, wird kontrovers geführt. Eine Studie von C. Johnson deutet darauf hin, dass das Aussterben des Wollhaarmammuts und anderer pleistozäner Arten mit einer rapiden Abnahme der Fruchtbarkeit einherging.[23] Eine zu geringe Reproduktionsrate sieht er bei einer Reihe von Großsäugern in Australien, Eurasien, Amerika und Madagaskar als Hauptursache des Aussterbens, während er die „Overkill-Hypothese“ (bei Johnson: „Blitzkrieg-Hypothese“) als ursächliches Szenario zurückweist. Da Arten mit zurückgehender Reproduktionsrate bei menschlicher Bejagung zusätzlichem Stress ausgesetzt sind, sei die Gleichzeitigkeit des Aussterbens mit der verstärkten Bejagung durch Jäger-und-Sammler-Populationen die logische Folge. Eine weitere, teils häufig vertretene Meinung sieht die Ursachen eher im Klimawandel am Ende der Eiszeit und den dadurch hervorgerufenen ökologischen Veränderungen (v. a. bodenbildungsbedingten Vegetationswandel). Das Einwirken des Menschen wird in dieser Sichtweise mitunter nur als Nebeneinfluss auf durch ökologische Faktoren bereits geschwächte Spezies eingeschätzt.[24] Für das Aussterben der Mammute wird daher vom dänischen Biologen Roy Weber der Universität Aarhus der Schwund der Kräuter in der Kaltsteppe und damit das Klima verantwortlich gemacht.[25] Ähnlich argumentiert eine Studie aus dem Jahr 2021. Anhand von Umwelt-DNA, also genetischen Resten der Fauna und Flora aus Bodenproben, aus dem Großteil des circumpolaren Gebietes kommt sie zu dem Schluss, dass der Rückgang der Mammutsteppe als nährstoffreiche Nahrungsgrundlage der Mammute zum Ende der letzten Kaltzeit verbunden mit wärmeren und feuchteren Klimabedingungen den Populationen der Rüsseltiere stark zusetzte. Der Einfluss des Menschen wird hier als weitgehend gering eingestuft.[26]
Bisher wurde angenommen, das Wollhaarmammut sei in Europa und Südsibirien bereits am Ende des Jungpleistozäns vor etwa 12.000 Jahren ausgestorben, nachdem es in der letzten Kaltphase („Jüngere Dryas“) am Ende der Weichsel-Eiszeit noch einmal bis Nordosteuropa vordringen konnte.[10][27][28][29] Neu datierte Altfunde der 1940er Jahre aus Russland zeigen aber, dass die letzten Mammute erst um 9250 v. Chr., also etwa 300 Jahre nach Beginn des Präboreal, aus Nordosteuropa verschwanden.[30] Nur wenig später verschwand die Art – nach dem heutigen Fossilreport zu urteilen – auch auf dem nordsibirischen Festland. Nur auf der ostsibirischen Wrangelinsel überlebten kleine Populationen bis etwa 2000 v. Chr.[31]
In Kontinentalalaska starb das Wollhaarmammut (Mammuthus primigenius) etwa um 12.000 v. Chr. aus,[24] während auf St. Paul, der nördlicheren der Pribilof-Inseln, eine Restpopulation der Wollhaarmammute der vormaligen Beringia-Landmasse[32] isoliert bis etwa 3600 v. Chr. bestand.[33][34] Auf den im Spätpleistozän nur durch einen schmalen Kanal von Kalifornien getrennten Kanalinseln (Kalifornien) überlebte eine als Mammuthus exilis[35] bezeichnete Inselverzwergung des Präriemammuts (Mammuthus columbi) bis etwa 11.000 v. Chr.[36] möglicherweise sogar bis 10.200 v. Chr.[37] – die kontinentale Form des Präriemammuts (Mammuthus columbi) starb ebenfalls ungefähr zu dieser Zeit aus.
Entdeckung erhaltener Körper
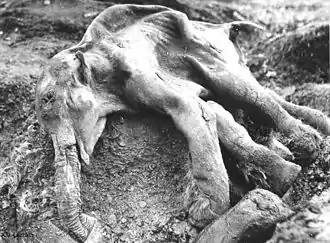
Im asiatischen Teil Russlands werden immer wieder nahezu vollständig im Eis eingeschlossene gut erhaltene Mammutkörper gefunden. Entdeckt werden diese meist durch den kilometerweit wahrnehmbaren beißenden Moschus- und Verwesungsgeruch, sobald Teile des Tierkörpers durch Auftauen freigelegt wurden. Neben der raschen Verwesung sorgen auch Aasfresser dafür, dass solche bis dahin über Jahrtausende ununterbrochen gefrorenen Kadaver oft schon binnen Wochen vollständig zerstört werden. Im Mai 2013 wurde auf den Ljachow-Inseln im Arktischen Ozean von russischen Wissenschaftlern ein äußerst gut erhaltenes älteres weibliches Mammut entdeckt, aus dessen Kadaver eine Probe flüssigen Blutes gewonnen werden konnte. Dies erhöht laut Angaben der Wissenschaftler die Chancen auf ein erfolgreiches Klonen von Mammuten.[38]

Fundorte und Museen
Zu wichtigen Fundorten, die viel zur Analyse der Lebensweise der Mammute beigetragen haben, gehören die Teergruben von Rancho La Brea und die Bechan Cave, eine Höhle, die vor 15.000 Jahren über eine Dauer von 1.500 Jahren von Präriemammuten genutzt wurde.
Überreste in großen Mengen wurden zudem auf den Neusibirischen Inseln gefunden, die einen Gutteil ihrer Entdeckungsgeschichte russischen Händlern verdanken, die Überreste von Mammuten, insbesondere deren elfenbeinerne Stoßzähne, suchten.
Sieben vollständige Mammutskelette sind im Mammutheum Siegsdorf, ein großes im Südostbayerischen Naturkunde- und Mammut-Museum Siegsdorf im Chiemgau und ein Teilskelett im Museum für Ur- und Frühgeschichte in Eichstätt zu besichtigen. Weitere, mehr oder weniger vollständige Mammutskelette befinden sich beispielsweise in Stuttgart, Münster (Fundort: Ahlen), Bottrop, Darmstadt, Halle an der Saale und Sangerhausen. Die größte Fundstätte in der Schweiz ist in Niederweningen, wo die Funde in einem extra dafür eingerichteten Mammutmuseum zu sehen sind.[39]
Im Juni 2009 wurde in einer Kohlegrube im serbischen Kostolac in unmittelbarer Nähe der Ausgrabungsstätte des ehemaligen römischen Legionärslagers und Stadt Viminatium das nahezu unversehrte Skelett eines Steppenmammuts (Mammuthus trogontherii) entdeckt. Der Fund datiert in das Alt- bis Mittelpleistozän und ist zwischen 1 Million und 400.000 Jahre alt. Es handelt sich um ein männliches Tier mit einem Alter von über 60 Jahren. Die Auswertung des Fundes übernahm ein Wissenschaftlerteam um Adrian M. Lister.[40] An derselben Lokalität wurde 2012 ein wissenschaftlich bedeutender Mammutfriedhof freigelegt; er barg mehrere Mammut-Skelette, die stark disartikuliert, also in nicht mehr ursprünglichem anatomischen Zusammenhang waren. Diese stammen aber aus Lössschichten des Mittleren Pleistozäns und sind radiometrischen Daten zufolge rund 192.000 Jahre alt.[41]
Schließlich wurde im Juni 2014 auf dem Gelände des Archäologieparkes Viminatium mit den Kostolac-Mammut-Fossilien der erste sogenannte „Mammut-Park“ in Europa eröffnet. Neben dem Steppenmammut sind noch vier weitere Mammutskelette ausgestellt. Das Kostolacer Mammut ist damit eines der wenigen Mammutfossilien, welches direkt an seinem Fundort ausgestellt ist.[42]
Handel mit Mammutskeletten
Schon aufgrund der enormen Größe befinden sich nur wenige vollständige Mammutskelette in Privatbesitz. 2016 wurde ein 3,5 Meter hohes und über 5,5 Meter langes Skelett, das ein Fossiliensammler aus 270 Knochen zusammengesetzt hatte, für 120.000 € versteigert.[43]
Etymologie
Die Bezeichnung Mammut ist seit dem 17. Jahrhundert in Europa verbreitet. Der Name wurde möglicherweise durch den Amsterdamer Bürgermeister Nicolaas Witsen (1641–1717) eingeführt,[44] der im Jahre 1692 einen Reisebericht nach Nordostsibirien veröffentlichte.[45] Das Wort, das im Russischen und in einigen älteren europäischen Quellen auch Mamont (мамонт) lautet, stammt aus einer sibirischen Sprache. Als ein mögliches Ausgangswort wurde die waldnenzische (waldjurakische) Bezeichnung „jěaŋ-ŋammurəttaə“ („Erdfresser“) identifiziert.[46]
Bilder
 Rekonstruktion (Naturhistorisches Museum Basel)
Rekonstruktion (Naturhistorisches Museum Basel) Skelett
Skelett Molar eines Mammuts, Blick auf die Kaufläche
Molar eines Mammuts, Blick auf die Kaufläche Querschnitt durch einen Mammutstoßzahn. Die Risse in den äußeren Schichten entstehen durch den Feuchtigkeitsverlust, der nach Ausgrabungen eintritt.
Querschnitt durch einen Mammutstoßzahn. Die Risse in den äußeren Schichten entstehen durch den Feuchtigkeitsverlust, der nach Ausgrabungen eintritt. Mammut-Kleinplastik aus dem Aurignacien (Vogelherdhöhle)
Mammut-Kleinplastik aus dem Aurignacien (Vogelherdhöhle)
Siehe auch
Literatur
- Vadim Evgenievič Garutt: Das Mammut. Mammuthus primigenius (Blumenbach). Nachdruck der 1. Auflage von 1964. Westarp Wissenschaften, Hohenwarsleben 2004, ISBN 3-89432-171-7.
- Ulrich Joger (Hrsg.): Mammuts aus Sibirien. Begleitbuch zur Ausstellung im Hessischen Landesmuseum Darmstadt vom 20. Oktober 1994 bis 19. Februar 1995. Hessisches Landesmuseum Darmstadt, Darmstadt 1994, ISBN 3-926527-34-X.
- Adrian Lister, Paul Bahn: Mammuts. Die Riesen der Eiszeit. Thorbecke, Sigmaringen 1997, ISBN 3-7995-9050-1.
- Richard Stone: Mammut – Rückkehr der Giganten? Franckh-Kosmos, Stuttgart 2003, ISBN 3-440-09520-7.
- Peter D. Ward: Ausgerottet oder ausgestorben? Warum die Mammuts die Eiszeit nicht überleben konnten. Birkhäuser, Basel 1998, ISBN 3-7643-5915-3.
- Reinhard Ziegler: Das Mammut (Mammuthus primigenius Blumenbach) von Siegsdorf bei Traunstein (Bayern) und seine Begleitfauna. In: Münchner Geowissenschaftliche Abhandlungen. Reihe A: Geologie und Paläontologie. 26, 1994, ISSN 0177-0950, S. 49–80.
Einzelnachweise
- W. Miller u. a.: Sequencing the nuclear genome of the extinct woolly mammoth. In: Nature. 456, 2008, S. 387–390. (online)
- Eske Willerslev, John Davison, Mari Moora, Martin Zobel, Eric Coissac, Mary E. Edwards, Eline D. Lorenzen, Mette Vestergård, Galina Gussarova, James Haile, Joseph Craine, Ludovic Gielly, Sanne Boessenkool, Laura S. Epp, Peter B. Pearman, Rachid Cheddadi, David Murray, Kari Anne Bråthen, Nigel Yoccoz, Heather Binney, Corinne Cruaud, Patrick Wincker, Tomasz Goslar, Inger Greve Alsos, Eva Bellemain6, Anne Krag Brysting, Reidar Elven, Jørn Henrik Sønstebø, Julian Murton, Andrei Sher, Morten Rasmussen, Regin Rønn, Tobias Mourier, Alan Cooper, Jeremy Austin, Per Möller, Duane Froese, Grant Zazula, Francois Pompanon, Delphine Rioux, Vincent Niderkorn, Alexei Tikhonov, Grigoriy Savvinov, Richard G. Roberts, Ross D. E. MacPhee, M. Thomas P. Gilbert, Kurt H. Kjær, Ludovic Orlando, Christian Brochmann, Pierre Taberle: Fifty thousand years of Arctic vegetation and megafaunal diet. In: Nature. Band 506, 2014, S. 47–51 doi:10.1038/nature12921.
- Wighart von Koenigswald: Lebendige Eiszeit. Klima und Tierwelt im Wandel. Stuttgart 2002.
- Per Christiansen: Body size in proboscideans, with notes on elephant Metabolism. In: Zoological Journal of the Linnean Society. 140, 2004, S. 523–549.
- Adrian Lister, Paul Bahn: Mammuts – Die Riesen der Eiszeit. Sigmaringen 1997.
- Victoria L. Herridge, Adrian M. Lister: Extreme insular dwarfism evolved in a mammoth. In: Proceedings of the Royal Society. series B, 2012. doi:10.1098/rspb.2012.0671
- Adrian M. Lister, Andrei V. Sher, Hans van Essen, Guangbiao Wei: The pattern and process of mammoth evolution in Eurasia. In: Quaternary International. 126–128, 2005, S. 49–64.
- William J. Sanders, Emmanuel Gheerbrant, John M. Harris, Haruo Saegusa, Cyrille Delmer: Proboscidea. In: Lars Werdelin, William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley/ London/ New York 2010, S. 161–251.
- Vincent J. Maglio: Four new species of Elephantidae from the Plio-Pleistocene of northwestern Kenya. In: Breviora. 341, 1970, S. 1–43.
- Ralf-Dietrich Kahlke: Die Entstehungs-, Entwicklungs- und Verbreitungsgeschichte des Oberpleistozänen Mammuthus-Coelodonta-Faunencomplexes in Eurasien (Großsäuger). In: Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft. 546, 1994, S. 1–64.
- Nadin Rohland, Anna-Sapfo Malaspinas, Joshua L. Pollack, Montgomery Slatkin, Paul Matheus, Michael Hofreiter: Proboscidean Mitogenomics: Chronology and Mode of Elephant Evolution Using Mastodon as Outgroup. In: PLOSBiology. 5, August 2007, S. 1663–1671.
- Jon E. Kalb, David J. Froehlich, Gordon L. Bell: Palaeobiogeography of late Neogene African and Eurasian Elephantoidea. In: Jeheskel Shoshani, Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford/ New York/ Tokyo 1996, S. 117–123.
- William J. Sanders, Yohannes Haile-Selassie: A New Assemblage of Mid-Pliocene Proboscideans from the Woranso-Mille Area, Afar Region, Ethiopia: Taxonomic, Evolutionary, and Paleoecological Considerations. In: Journal of Mammal Evolution. 19, 2012, S. 105–128.
- Spencer G. Lucas, Guillermo E. Alvarado: Fossil Proboscidea from the Upper Eozoic of Central America: Taxonomy, evolutionary and paleobiogeographic significance. In: Revista Geológica de América Central. 42, 2010, S. 9–42.
- Jacob Enk, Alison Devault, Regis Debruyne, Christine E. King, Todd Treangen, Dennis O’Rourke, Steven L. Salzberg, Daniel Fisher, Ross MacPhee, Hendrik Poinar: Complete Columbian mammoth mitogenome suggests interbreeding with woolly mammoths. In: Genome Biology. 12, 2011, S. R51. (online)
- Karol Schauer: Anmerkungen und Quellenangaben zur Evolutionstafel der Proboscidea in Afrika und Asien. In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale 2010, S. 630–650.
- Jeheskel Shoshani, Pascal Tassy: Advances in proboscidean taxonomy & classification, anatomy & physiology, and ecology & behavior. In: Quaternary International. 126–128, 2005, S. 5–20.
- S. Gaudzinski, E. Turner, A. P. Anzidei, E. Àlvarez-Fernández, J. Arroyo-Cabrales, J. Cinq-Mars, V. T. Dobosi, A. Hannus, E. Johnson, S. C. Münzel, A. Scheer, P. Villa: The use of Proboscidean remains in every-day Palaeolithic life. In: Quaternary International. 126–128, 2005, ISSN 1040-6182, S. 179–194. doi:10.1016/j.quaint.2004.04.022.
- Mietje Germonpré, Mikhail Sablin, Gennady Adolfovich Khlopachev, Galina Vasilievna Grigorieva: Possible evidence of mammoth hunting during the Epigravettian at Yudinovo, Russian Plain. In: Journal of Anthropological Archaeology. 27, Nr. 4, 2008, S. 475–492. doi:10.1016/j.jaa.2008.07.003.
- Jeanne Timmons: Scientists Find Mammoth Seemingly Butchered by Humans on Arctic Island, auf: Gizmodo vom 11. Dezember 2020
- I. G. Pidoplichko: Upper palaeolithic dwellings of mammoth bones in the Ukraine. In: BAR international series. 712, 1998.
- AMS-Holzkohlendaten 14,610 ± 60 BP (GifA 80147/SacA-11486), 14,600 ± 60 yr BP (GifA 80148/SacA-11487); L. Marquer, V. Lebreton, T. Otto, H. Valladas, P. Haesaerts, E. Messager, D. Nuzhnyi, S. Péan: Charcoal scarcity in Epigravettian settlements with mammoth bone dwellings: the taphonomic evidence from Mezhyrich (Ukraine). In: Journal of Archaeological Science. 39 (1), 2012, S. 109–120. doi:10.1016/j.jas.2011.09.008
- C. N. Johnson: Determinants of loss of mammal species during the Late Quaternary ‘megafauna’ extinctions: life history and ecology, but not body size. In: Proceedings of the Royal Society of London B 269. 2002, S. 2221–2227.
- Daniel Mann, Pamela Groves, Michael Kunz, Richard Reanier, Benjamin Gaglioti: Ice-age megafauna in Arctic Alaska: extinction, invasion, survival. In: Quaternary Science Reviews. Volume 70, Juni 2013, S. 91–108.
- videnskab.dk
- Yucheng Wang, Mikkel Winther Pedersen, Inger Greve Alsos, Bianca De Sanctis, Fernando Racimo, Ana Prohaska, Eric Coissac, Hannah Lois Owens, Marie Kristine Føreid Merkel, Antonio Fernandez-Guerra, Alexandra Rouillard, Youri Lammers, Adriana Alberti, France Denoeud, Daniel Money, Anthony H. Ruter, Hugh McColl, Nicolaj Krog Larsen, Anna A. Cherezova, Mary E. Edwards, Grigory B. Fedorov, James Haile, Ludovic Orlando, Lasse Vinner, Thorfinn Sand Korneliussen, David W. Beilman, Anders A. Bjørk, Jialu Cao, Christoph Dockter, Julie Esdale, Galina Gusarova, Kristian K. Kjeldsen, Jan Mangerud, Jeffrey T. Rasic, Brigitte Skadhauge, John Inge Svendsen, Alexei Tikhonov, Patrick Wincker, Yingchun Xing, Yubin Zhang, Duane G. Froese, Carsten Rahbek, David Bravo Nogues, Philip B. Holden, Neil R. Edwards, Richard Durbin, David J. Meltzer, Kurt H. Kjaer, Per Möller, Eske Willerslev: Late Quaternary dynamics of arctic biota from ancient environmental genomics. In: Nature., 2021, doi:10.1038/s41586-021-04016-x.
- A. J. Stuart: Mammalian extinctions in the late Pleistocene of Northern Eurasia and North America. In: Biological Reviews. 66, 1991, S. 453–562.
- A. J. Stuart: Late Pleistocene megafaunal extinctions; a European perspective. In: R. D. E. MacPhee (Hrsg.): Extinctions in Near Time: Causes, Contexts and Consequences. Kluwer Academic/Plenum Publishers, New York 1999.
- Anthony Stuart, Leopold Sulerzhitsky, Lyobov Orlova, Yaroslav Kuzmin, Adrian Lister: The latest woolly mammoths (Mammuthus primigenius Blumenbach) in Europe and Asia: a review of the current evidence. In: Quaternary Science Reviews. 21 (14–15), 2002, S. 1559–1569. doi:10.1016/S0277-3791(02)00026-4
- Tscherepowez-Mammut AMS-Knochendatum: 9760 ± 40 BP (GIN-8885c); A. J. Stuart: The extinction of woolly mammoth (Mammuthus primigenius) and straight-tusked elephant (Palaeoloxodon antiquus) in Europe. In: Quaternary International. 126–128, 2005, S. 171–177.
- Anthony J. Stuart, Leopold D. Sulerzhitsky, Lyobov A. Orlova, Yaroslav V. Kuzmin, Adrian M. Lister: The latest woolly mammoths (Mammuthus primigenius Blumenbach) in Europe and Asia: a review of the current evidence. In: Quaternary Science Reviews. 21 (14–15), 2002, S. 1559–1569. doi:10.1016/S0277-3791(02)00026-4
- J. Enk, D. Yesner, Kr. Crossen, D. Veltre, D. O'Rourke: Phylogeographic analysis of the mid-Holocene Mammoth from Qagnax Cave, St. Paul Island, Alaska. In: Palaeogeography, Palaeoclimatology, Palaeoecology. 273 (1–2), 2009, S. 184–190. doi:10.1016/j.palaeo.2008.12.019
- Kr. Crossen, D. Yesner, D. Veltre, R. Graham: 5,700-year-old mammoth remains from the Pribilof Islands, Alaska: last outpost of north american megafauna. In: Geological Society of America Abstracts with Programs. 37/7, 2005, S. 463. (online)
- Russell W. Graham, Soumaya Belmecheri, Kyungcheol Choy, Brendan J. Culleton, Lauren J. Davies, Duane Froese, Peter D. Heintzman, Carrie Hritz, Joshua D. Kapp, Lee A. Newsom, Ruth Rawcliffe, Émilie Saulnier-Talbot, Beth Shapiro, Yue Wang, John W. Williams, Matthew J. Wooller: Timing and causes of mid-Holocene mammoth extinction on St. Paul Island, Alaska. In: Proceedings of the National Academy of Sciences. online veröffentlicht am 1. August 2016. doi:10.1073/pnas.1604903113
- Donald Johnson: The origin of Island Mammoths and the quaternary land bridge history of the Northern Channel Islands, California. Quaternary Research 10 (2), 1978, S. 204–225, doi:10.1016/0033-5894(78)90102-3.
- AMS-Datierung Mammutknochen 11030 ± 50 BP (CAMS 168100); Larry Agenbroad, John Johnson, Don Morris, Thomas Stafford: Mammoths and humans as late pleistocene contemporaries on Santa Rosa Island. In: Dave Garcelon, Catherin Schwemm (Hrsg.): Proceedings of the Sixth California Islands Symposium, Ventura, California, December 1–3, 2003. National Park Service technical publication, CHIS-05-01 (Arcata/Calif.: Institute for Wildlife Studies 2005).
- konventionelles Datum von vergesellschafteter Holzkohle 10290 ± 100 BP (AA-1268); Adrian Wenner, John Cushing, Elmer Noble, Marla Daily: Mammoth radiocarbon dates from the Northern Channel Islands, California. In: Martin Rosen, Lynne Christenson, Timothy Gross (Hrsg.): Proceedings of the Society for California Archaeology. Volume 4, 1991. Papers presented at the annual meeting of the Society for California Archaeology (San Diego/California 1991), S. 221–226.
- Russian scientists make rare find of 'blood’ in mammoth, phys.org, 29. Mai 2013, abgerufen am 29. Mai 2013.
- mammutmuseum.ch
- Adrian M. Lister, Vesna Dimitrijević, Zoran Marković, Slobodan Knežzević, Dick Mol: A skeleton of ‘steppe’ mammoth (Mammuthus trogontherii (Pohlig)) from Drmno, near Kostolac, Serbia. In: Quaternary International. 276–277, 2012, S. 129–144. doi:10.1016/j.quaint.2012.03.021
- Vesna Dimitrijević, Nemanja Mrdjić, Miomir Korać, Seimi Chu, Dejan Kostić, Mladen Jovičić und Bonnie A. B. Blackwell: The latest steppe mammoths (Mammuthus trogontherii (Pohlig)) and associated fauna on the Late Middle Pleistocene steppe at Nosak, Kostolac Basin, Northeastern Serbia. In: Quaternary International. 379, 2015, S. 14–27. doi:10.1016/j.quaint.2015.06.025
- Nemanja Tomić, Slobodan B. Marković, Miomir Korać, Nemanja Mrđić, Thomas A. Hose, Djordjije A. Vasiljević, Mladen Jovičić und Milivoj B. Gavrilov: Exposing mammoths: From loess research discovery to public palaeontological park. In: Quaternary International. 372, 2015, S. 142–150. doi:10.1016/j.quaint.2014.12.026
- Mammutskelett für 120.000 Euro versteigert. orf.at 11. Oktober 2016, abgerufen 17. Oktober 2016.
- J. Augusta, Z. Burian: Das Buch von den Mammuten. Artia Verlag, 1962, S. 24.
- Nicolaas Witsen: Noord en Oost Tartarije. Amsterdam, 1692.
- Kluge: Etymologisches Wörterbuch der deutschen Sprache. 23., erweiterte Auflage. Bearbeitet von Elmar Seebold. Berlin/ New York 1999, ISBN 3-11-016392-6.
Weblinks
- LWL-Museum für Naturkunde, Westfälisches Landesmuseum mit Planetarium
- Mammutheum
- Mammutmuseum Niederweningen
- Naturkundemuseum Siegsdorf
- Universität Tübingen Älteste Darstellung eines Mammuts
- Das Mammut vom Vogelherd Eines der ältesten Kunstwerke der Menschheit aus Mammutelfenbein geschnitzt
- Expedition Mammut: Zeittafel auf Spiegel Online