Sukzession (Biologie)
Unter Sukzession (lateinisch succedere „nachrücken“, „nachfolgen“) versteht man die natürliche Rückkehr der für einen Standort typischen Pflanzen-, Tier- und Pilzgesellschaften (Biozönose), die sich nach einer Störung aufgrund der vorherrschenden Umweltfaktoren (vor allem Klima und Bodenart) dort wieder einstellt. Diese sukzessive Entwicklung führt von einem mehr oder minder gestörten oder veränderten Ausgangsstadium (zum Beispiel durch Sturmschäden, Lawinen, Vulkanausbrüche usw. oder diverse vom Menschen verursachte Veränderungen der Natur wie etwa Rodung oder Übernutzung) im Extremfall vom vegetationsfreien Boden („Initialstadium“) über verschiedene Zwischenstadien zu einem (über eine längere Zeit) stabilen Endstadium (Klimaxgesellschaft), sofern keine erneuten Störungen auftreten.

In der Umgangssprache werden Flächen, die früher genutzt und seither über einen längeren Zeitraum sich selbst überlassen wurden, als Sukzessionsflächen bezeichnet. Sukzession wird dann häufig mit Verbuschung gleichgesetzt.
Die natürliche Sukzession lässt sich mit dem Heilungsprozess eines Lebewesens vergleichen.
Ablauf


Rechts: Bildung einer geschlossenen Pflanzendecke.
Sukzession kann in allen Ökosystemen stattfinden. Ausgangspunkt sind neu entstandene (z. B. freigelegte Felsoberflächen, frische Sanddünen), gestörte (z. B. Wälder nach Waldbrand oder Sturmschaden) oder genutzte (z. B. Wiesen, Heiden) Lebensräume. Aber auch in natürlichen Lebensräumen kann Sukzession stattfinden, wenn sich die Lebensbedingungen ändern (z. B. Klimawandel). Folgt die Lebensgemeinschaft wie in diesem Fall passiv sich graduell ändernden abiotischen Umweltbedingungen, spricht man auch von allogener (exogener) Sukzession.
Die Lebensgemeinschaft selbst stellt dabei aber oft eine wesentliche Ursache bzw. einen Antrieb für den weiteren Verlauf der Sukzession dar. Etablierte Lebensgemeinschaften können die Standortfaktoren selbst verändern, z. B. durch Bodenbildung (Pedogenese) oder beim Verlanden eines Sees durch abgestorbene Pflanzenstreu der Röhrichtarten. Neu hinzukommende Arten können weiteren Arten die Besiedlung erleichtern, oder sie können sie (durch Konkurrenz) verdrängen. Eine solche Sukzession, die durch die Einwirkung der Organismen einer Lebensgemeinschaft selbst vorangetrieben wird, wird autogene (endogene) Sukzession genannt.
Es gibt drei mögliche Schlüsselprozesse bei einer Sukzession[1]
- Förderung: Vorgänger fördern Nachfolger, indem sie den Lebensraum und Standort zu deren Gunsten verändern, z. B. indem Pflanzen mit hohem Nährstoffbedarf erst durch die Akkumulation von Nährstoffen durch Pflanzenstreu anderer Arten im Boden gedeihen können. Arten späterer Sukzessionsstadien sind dann auf Pionierarten als Erstbesiedler angewiesen.
- Toleranz: Arten können sich ansiedeln, ohne von vorher dort wachsenden Arten zu profitieren. Dies kommt z. B. vor, wenn später aufkommende Arten ein niedrigeres Niveau von essentiellen Ressourcen tolerieren können als Erstbesiedler. Sie können dann in einen Lebensraum eindringen, obwohl dieser durch Individuen der Erstbesiedler bereits besetzt ist. Letztlich führt dies zur Verdrängung der Erstbesiedler.
- Hemmung: Spätere Arten können sich nur trotz der Frühbesiedler etablieren. Solange diese vital sind, hindern sie die Folgearten an der Etablierung.
Wesentlicher Faktor ist oft Konkurrenz: konkurrenzschwache Pioniere (r-Strategen) werden so durch konkurrenzstarke Nachfolger (K-Strategen) ersetzt.
Das Verständnis über die Sukzession lässt sich z. B. in Landschaftsplanung und Naturschutz anwenden, wenn es darum geht, den gestörten Naturhaushalt – als Lebensgrundlage für den Menschen – wiederherzustellen und langfristig zu sichern.
Die Annahme eines (einzigen) Ökologischen Gleichgewichts, auf das sich Ökosysteme nach Störungen durch Sukzession hinentwickeln würden, wurde in der Ökologie durch empirische Studien in Frage gestellt und ist neueren Modellen dynamischer und multipler Gleichgewichte gewichen.
Sukzessionsdauer
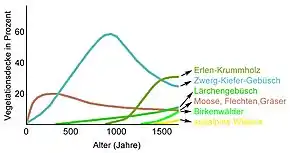
Einige Beispiele für die Dauer der Sukzession: Für Mitteleuropa kann eine Sukzessionsdauer von 300 bis 600 Jahren von unbelebten Flächen bis hin zum natürlichen Wald angenommen werden.[2] Der Piedmont-Laubwald des östlichen Appalachen-Plateaus benötigt mindestens 40 Jahre, um etwa nach Waldbränden oder Hurricans wieder seine ursprüngliche Arten-Zusammensetzung zu erreichen.[3] Die strauchartige Vegetation aus Feld-Beifuß der Binnendünen in der Oberrheinebene (zum Beispiel die Oftersheimer Dünen) kehrt bereits nach mindestens 10 Jahren wieder zu ihrem ursprünglichen Zustand zurück (sofern der Boden nicht verändert wurde).[4] In Süddeutschland benötigt ein Baggersee rund 30 Jahre bis zur Ausbildung eines typischen Seenbiotopes.[5]
Die Dauer der (primären) Sukkzession ist von verschiedenen Faktoren abhängig: Insbesondere das Großklima, die Bodenart und die Entfernung zu den Quellen für die Wiederbesiedlung (Samen und/oder Wurzeln noch im Boden oder von entfernten Standorten) sind entscheidend. So ist es naheliegend, dass ein trockenes Kontinentalklima, in dem die Klimaxvegetation eine baumlose, relativ artenarme Grassteppe ist, nach einem Brand bereits nach wenigen Jahren wieder im ursprünglichen Zustand ist, während es auf Lava-Bims im borealen Gebirgsklima über 1500 bis 2000 Jahre dauern kann (siehe Grafik), bevor sich wieder die heute zu erwartende subalpine Waldrandvegetation einstellt – um zwei Extrembeispiele zu nennen. Im feucht-subtropischen Klima Südjapans dauert die Sukzession eines ähnlichen Waldrandstandortes an Vulkanhängen nur 700 Jahre.[6] Bei der tropischen Vulkaninsel Krakatau, die 1883 nahezu vollkommen zerstört wurde, dauerte es gerade einmal 50 Jahre, bis dort wieder ein Sekundärwald stand.[7] Die zyklisch wiederkehrende Regeneration eines tropischen Regenwaldes bis zu einem vollständigen Primärwald dauert wiederum Jahrhunderte.
Je länger eine Sukzessionsabfolge dauert, desto unsicherer ist ihre Rekonstruktion, da innerhalb solch langer Zeiträume der normale Klimawandel zu veränderten Bedingungen und damit anderen Klimaxzuständen führt als zu Beginn der Sukzession. So wird etwa diskutiert, ob die heutigen Lorbeerwälder der zentralen Anden eher ein Sukzessionsstadium zu einem reinen Araukarienwald sind, denn eine Schlusswaldgesellschaft aus Araukarien und sommergrünen Scheinbuchen.[8]
Typen und Stadien
In der Botanik werden unterschieden:
- „primäre“ Sukzession: Diese setzt auf bisher unbesiedelten Standorten ein.
- „sekundäre“ Sukzession: Ausgangspunkt sind hier bereits besiedelte und von den vorkommenden Arten veränderte Standorte. Das können vorhandene Vegetationsbestände sein, z. B. Sukzession von Wiese zu Wald bei Nutzungsaufgabe. Aber auch bei vegetationslosen Standorten sind hier bereits Bodenentwicklungen abgelaufen (z. B. Humusgehalt, Nährstoffanreicherung) und im Boden ist meist eine Samenbank (Diasporenbank) vorhanden. Sekundärsukzession tritt ebenfalls bei Störung einer Gemeinschaft auf, die den Boden intakt ließ. Häufig entwickelt sich dann das gestörte Gebiet wieder in seinen ursprünglichen Zustand zurück.
Im Falle einer Sukzession auf unbewachsenem Boden ist folgende Abfolge typisch:
Initialstadium, Folgestadien, Klimaxstadium

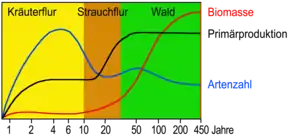
Im Initialstadium erschließen Pionierarten das unbesiedelte Gebiet. Ist es aufgrund einer Störung durch Menschen oder aufgrund menschlicher Nutzung entstanden, spricht man auch von Ersatzgesellschaften. Arten solcher Pioniergesellschaften müssen über effektive Mechanismen zur Fernverbreitung verfügen (z. B. windverbreitete Samen). Oft besitzen sie im Gegensatz zu Arten der Klimaxgesellschaften eine größere Toleranz gegenüber extremen Standortfaktoren. Im Initialstadium und in frühen Stadien der Sukzession herrschen Arten mit hohem Fortpflanzungspotenzial, die r-Strategen, vor (benannt nach dem Reproduktionsfaktor r in der logistischen Gleichung). r-Strategen reproduzieren sich rasch und in großer Zahl. Im Lauf der Zeit wandern weitere Arten ein, die sich langsamer ausbreiten und die meist konkurrenzschwachen Pionierarten verdrängen. Die Pionierarten verändern außerdem die Standortfaktoren, z. B. durch Akkumulation (Ansammlung) von Stickstoff, anderen Nährstoffen und Humus, verändern Wasserhaushalt und Klima, wirken auf den Boden (siehe Pedogenese) und auf die Fauna (Gesamtheit der Tierarten) ein.

Auf Grund der veränderten Standortfaktoren sind nun wiederum andere Arten in der Lage, das veränderte Biotop zu besiedeln. Diese Arten sind anspruchsvoller (Klima, Wasser, Nährstoffe etc.) und besitzen eine höhere Produktivität. In Folgestadien setzen sich deshalb mehr und mehr die K-Strategen durch (benannt nach der Lebensraumkapazität K in der logistischen Gleichung). K-Strategen vermehren sich weniger stark, haben also weniger Nachkommen. Diese besitzen aber ein höheres Durchsetzungsvermögen im Überlebenskampf und verdrängen die Pionierarten. Auch die neuen Arten verändern ihrerseits die Standortfaktoren, und der Prozess geht weiter, eine nächste, anspruchsvollere und produktivere Gesellschaft übernimmt das Regiment. Die Artenvielfalt nimmt dabei im Anfangsstadium zu, bevor sie wieder deutlich zurückgeht, um sich am Ende auf einem mittleren Level einzupendeln.[9]
Das Klimaxstadium ist erreicht, wenn sich die Artzusammensetzung nicht mehr oder nur noch sehr geringfügig verändert. Klassisch wird davon ausgegangen, dass das Klimaxstadium auch die höchste Primärproduktion an Biomasse besitzt, die auf einem Standort möglich ist. Das Klimaxstadium verwertet seine Ressourcen am effektivsten (wären noch nutzbare Ressourcen frei, könnten sie ja von hinzukommenden Arten verwertet werden. Damit ginge die Sukzession weiter, der Endpunkt wäre noch nicht erreicht).
Klimaxgesellschaften/Klimaxstadien
Der Klimax-Begriff in der Botanik geht auf Frederic Edward Clements zurück.[10] In seiner ursprünglichen Fassung ging er von einer für jede Klimazone einheitlichen Pflanzengemeinschaft aus, die sich bei genügend langer Entwicklungszeit schließlich überall und auf allen Standorten durchsetzen würde („Monoklimax“). In der modernen ökologischen Fachdiskussion wird der Begriff meist relativiert und mit abgewandelter Bedeutung verwendet. In der Regel geht man davon aus, dass sich stark unterschiedliche Standorte nicht völlig nivellieren würden („Polyklimax“). Außerdem werden (z. B. zyklische) Veränderungen auch im Klimaxstadium als Möglichkeit angenommen.
Die Klimaxvegetation entspricht (weitgehend) dem durch den Vegetationskundler Reinhold Tüxen eingeführten Begriff der „potenziellen natürlichen Vegetation“. (Unterschied: Dauerhafte Standortveränderungen im Zuge der Sukzession werden nicht berücksichtigt).
Nach vorherrschender Auffassung ist die Klimaxvegetation in Mitteleuropa außer auf Extrem- und Ausnahmestandorten ein weitestgehend geschlossener Wald. Auf den meisten Standorten handelt es sich um einen pflanzenartenarmen (Buchen)-Wald. Nur auf Sonderstandorten finden sich andere Klimaxgesellschaften. Moore bilden meistens Hochmoore als Klimaxstadium; Hochgebirgslagen, Watten bilden wiederum andere Klimaxstadien. Weitere Ausnahmen bilden u. a. azonale Waldgesellschaften, wie z. B. Auwälder, Bruchwälder.
Zu beachten ist dabei allerdings, dass manche Gesellschaften einer natürlichen Dynamik unterliegen (z. B. Flussauen mit regelmäßigen Hochwassern). Hier wird aufgrund dieser (natürlichen) Störungen die Klimaxvegetation unter Umständen nie erreicht. Diskutiert werden auch fortdauernde Lichtungen im Klimaxwald aufgrund (natürlicher) Störungen durch große Pflanzenfresser (vgl. Megaherbivorenhypothese).
Regressive Sukzession
Neben der oben geschilderten „normalen“ (oder „progressiven“) Sukzession auf eine höher organisierte „Klimaxgesellschaft“ hin kann es in Ausnahmefällen zu einer rückwärts gerichteten „regressiven“ Sukzession kommen, bei der hochentwickelte Lebensgemeinschaften durch einfacher strukturierte ersetzt werden. Dies ist z. B. der Fall, wenn nach einer Waldzerstörung (z. B. nach Waldbrand) der Boden so stark beeinträchtigt oder erodiert worden ist, dass die Entwicklung anschließend in eine völlig andere Richtung abläuft und nicht der Ausgangszustand, sondern eine einfacher strukturierte Lebensgemeinschaft (z. B. ein Gebüsch oder eine Felsheide) folgt.
Mosaik-Zyklus-Theorie
Die Mosaik-Zyklus-Theorie (auch Mosaik-Zyklus-Konzept) geht auf Kurt Michael Zukrigl zurück und wurde in den letzten Jahrzehnten vor allem durch Hermann Remmert propagiert.[11] Sie geht anstelle eines einheitlichen Klimaxstadiums von einer mosaikartigen Struktur aus, in der unterschiedlichste Sukzessionsstadien nebeneinander existieren.
Siehe auch
Literatur
- Hermann Remmert: Das Mosaik-Zyklus-Konzept und seine Bedeutung für den Naturschutz. 1991.
Weblinks
Einzelnachweise
- Joseph H. Connell; Ralph O. Slatyer (1977): Mechanisms of Succession in Natural Communities and Their Role in Community Stability and Organization. American Naturalist 111 (982): 1119-1144.
- Volker Beer: Klima und Vegetation. Klimaänderungen im 21. Jahrhundert, pdf, abgerufen am 21. Februar 2022, S. 10.
- Miller & Levine Biology: Lesson Overview 4.3 Succession, pdf, Seite 4, abgerufen am 21. Februar 2022.
- Arbeitsgemeinschaft Natur- und Umweltschutz Bruchsal e.V.: Sandbiotope Bruchsal 2022, abgerufen am 21. Februar 2022.
- Johann Senner, Deborah Graf, Aleina Döll: Landschaftsplan 2035 VG Stockach, Vorentwurf vom 11. November 2019, Planstatt Senner, Überlingen 2019. S. 52.
- Sergei Grishin u. Roger del Moral: Dynamics of Forests after catastrophic eruptions of Kamtchatka´s Volcanoes, in I. M. Turner, C. H. Diong, S. S. L. Lim u. P. K. L. Ng (Hrsg.): Biodiversity and the Dynamics of Ecosystems, DIWPA Serie, Ausgabe 1, S. 133–146, pdf, abgerufen am 20. Februar 2022. S. 139–140, 145.
- Dietmar Kalusche: Ökologie in Zahlen. Eine Datensammlung in Tabellen mit über 10.000 Einzelwerten.. 2. Auflage, Springer Spektrum, Berlin, Heidelberg 2016, ISBN 978-3-662-47986-5. S. 154–155.
- Jörg S. Pfadenhauer und Frank A. Klötzli: Vegetation der Erde. Springer Spektrum, Berlin/Heidelberg 2014, ISBN 978-3-642-41949-2. S. 3, 385.
- Dieter Heinrich, Manfred Hergt: Atlas zur Ökologie. Deutscher Taschenbuch Verlag, München 1990, ISBN 3-423-03228-6. S. 72–73.
- F. E. Clements: Nature and Structure of the Climax. (PDF; 1,8 MB) In: The Journal of Ecology. 1936, 24(1): 252–284.
- Hermann Remmert: Das Mosaik-Zyklus-Konzept und seine Bedeutung für den Naturschutz. 1991.