Cruziana
Cruziana ist der Name einer Spurenfossil-Gattung (Ichnogenus) des Paläozoikums und frühen Mesozoikums. Bei Cruziana im strengen, exklusiven Sinn, handelt sich um eine bandartige bilateralsymmetrische Fortbewegungs- oder kombinierte Fortbewegungs- und Fressspur (Repichnion), die entweder direkt auf oder knapp unterhalb der Sedimentoberfläche eines Gewässers (zumeist eines Meeres) erzeugt wurde. Im weiteren, inklusiveren Sinn umfasst Cruziana auch Fortbewegungs- sowie Ruhespuren (Cubichnia), die oft den Spurengattungen Rusophycus oder Isopodichnus zugewiesen werden. Verursacher von Cruziana und Rusophycus waren Trilobiten, aber sehr wahrscheinlich auch andere Arthropoden. Isopodichnus ist mit relativer Sicherheit nie von Trilobiten verursacht worden.

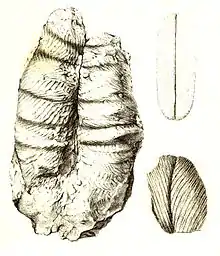
Aufgrund ihrer charakteristischen Morphologie, wie sie in typischer Überlieferung als Relief an Schichtunterseiten, d. h. als Ausfüllung der eigentlichen Spur, auftritt, werden Cruziana, Rusophycus, Isopodichnus und noch einige weitere, ähnliche Spurenfossilien auch als Bilobiten bezeichnet.
Forschungsgeschichte
Der Name Cruziana wurde 1842 durch den französischen Naturforscher Alcide Dessalines d’Orbigny in einem Teilband seines Werkes Voyage dans l’Amerique méridionale geprägt. Unter den Namen Cruziana rugosa und C. furcifer beschrieb er darin Fossilien aus dem Caradoc (spätes Ordovizium) von Liriuni nahe Cochabamba in Bolivien.[1] Im Glauben, es handele sich um Körperfossilien, ordnete d’Orbigny Cruziana unter anderem zusammen mit Trilobiten bei den Gliedertieren („animaux articulés“) ein.[2] Ursprünglich hatte er für die Gattung den Namen „Bilobites“ gewählt, und in den Bildunterschriften der zur Beschreibung gehörigen, doch bereits vorher veröffentlichten[3] Abbildungen wird dieser Name auch tatsächlich verwendet. Nachfolgend hatte d’Orbigny jedoch erfahren, dass der Name „Bilobites“ bereits vom US-Amerikaner James Ellsworth DeKay für fossile Muscheln benutzt wurde, und änderte den Namen zu Ehren des südamerikanischen Unabhängigkeitskämpfers Andrés de Santa Cruz in Cruziana.[2] Allem Anschein nach nutzte DeKay „Bilobites“ aber gar nicht im Sinne eines Gattungsnamens, sondern schlicht als englische Pluralform des Wortes „Bilobit“.[3] Allerdings hatte bereits 1831 der US-Amerikaner Constantine Samuel Rafinesque-Schmaltz einen Trilobiten unter dem Namen Bilobites beschrieben (heute ein Nomen dubium),[3] weshalb die Umbenennung dennoch sinnvoll war. Die informellen Bezeichnungen „Bilobit“ bzw. „Bilobiten“ werden bisweilen heute noch als Sammelbegriff für Spuren mit Cruziana-ähnlicher Morphologie genutzt (siehe Abschnitte Rusophycus und Isopodichnus).
Erhaltung und Morphologie
Cruziana ist meistens an Unterseiten von Sandsteinbänken als sogenanntes konvexes Hyporelief erhalten. Die Sandsteinbank repräsentiert im Regelfall eine Sturmablagerung (Tempestit), die an der Sedimentoberfläche erzeugte Spuren ausgefüllt hat. In einigen Fällen können im Sediment erzeugte Cruziana vorher, d. h. während des Sturms, durch die Tätigkeit grundberührender Wellen freigelegt worden sein. In seltenen Fällen ist Cruziana auch als Epirelief, d. h. an Schichtoberseiten, erhalten.[4]
Als Hyporelief weist Cruziana die typische bilobate Morphologie aus, die Namensgebend für den ursprünglich ausgewählten Gattungsnamen „Bilobites“ war, d. h. sie besteht aus zwei gleich großen, parallelen, meist relativ breiten und dicht beieinander liegenden Wülsten („Loben“), die durch eine zentrale, durchgehende, meist relativ schmale Furche voneinander getrennt sind. Die Wülste weisen eine Striemung auf, die fischgrätenartig entweder schräg oder quer zur zentralen Furche verläuft.[5][6]
Hierbei können grob zwei Subtypen unterschieden werden: die bandartigen „Cruziana Sensu stricto“ und die kurzen, ovalen („kaffebohnenförmigen“) oder tropfenförmigen Formen, die meist Rusophycus genannt werden. Die Breite einer Spur beträgt typischerweise mehr als einen und deutlich weniger als zehn Zentimeter. Die bandartigen Spuren können mehrere Meter Länge erreichen und verlaufen entweder gerade oder in relativ weiten Bögen. Kreuzen sich mehrere solcher Bögen auf einer Schichtfläche, wird auch von „Schleifen“ (engl.: loops) gesprochen.
Interpretation
Cruziana sensu stricto wird interpretiert als Fortbewegungs- oder kombinierte Fortbewegungs- und Fressspur von Trilobiten sowie anderen, morphologisch und ökologisch ähnlichen Arthropoden.
Der berühmte deutsche Palichnologe Adolf Seilacher postuliert, dass Trilobiten Cruziana erzeugten, während sie sich horizontal halb eingegraben im Sediment fortbewegten und dieses gleichzeitig nach organischen Partikeln durchkämmten (Seilacher beschreibt dieses Verhalten mit dem Wort plowing, „Pflügen“).[7] Die Loben seien dabei das Ergebnis des „Schaufelns“ von Sediment zur Mitte der Spur hin, unter den Rumpf des Trilobiten. Die Striemung auf den Loben gehe dabei auf die Tätigkeit der als Lauf- und Grabwerkzeuge funktionierenden Teile (Endopodite) der Arthropodenbeine im Sediment zurück. Die Bewegung der Endopodite sei dabei medio-posterior gewesen, d. h., ihre rumpffernen (distalen) Enden vollzogen auch eine Bewegung zum Körperende hin, während sie zur Längsachse des Körpers hin angezogen wurden. Nach dieser Hypothese zeigte das „V“, das von der Querstriemung auf den Loben meist gebildet wird, mit der offenen Seite in Bewegungsrichtung des Tieres. Nahrungspartikel, die von den Exopoditen aus dem Sediment „gesiebt“ und zur Nahrungsrinne an der Bauchseite geschaufelt wurden, seien von den Coxae, den proximalsten (rumpfnächsten) Beingliedern, in die Kopfregion zur Mundöffnung weitergereicht worden. Dass Cruziana, anders als es bei Weidespuren im engeren Sinn (Pascichnia) der Fall ist, keine Mäander, sondern gerade Bänder oder weite Bögen bildet, liege im Körperbau der Trilobiten begründet, der das Laufen enger Kurven nicht gestattete.[7]
Einige Cruziana-Ichnospezies weisen neben den Querstriemen auch eine deutlich feiner ausgebildete Längsstriemung auf, die als Streifspuren (engl.: „brushings“) der federartigen Kiemen, die an den Exopoditen der Trilobitenbeine saßen, interpretiert werden.[7]
Es gilt als unstrittig, dass die Striemungsmuster, einschließlich der „Brushings“, sowie weitere spezielle Merkmale der verschiedenen Cruziana-Ichnospezies, in engem Zusammenhang mit der Anatomie, insbesondere der Beine, des erzeugenden Arthropoden stehen. Dieser Zusammenhang wird auch als „Fingerprinting“ bezeichnet.[5]
Erzeuger
Oft wird Cruziana vereinfachend als „Trilobitenspur“ bezeichnet. In zahlreichen Fällen mag dies stimmen, und in einigen Fällen ist sogar eine enge stratigraphische und räumliche Assoziation von Spuren und Trilobiten nachgewiesen, beispielsweise zwischen der Ichnospezies Cruziana semiplicata und der Trilobitengattung Maladioidella im Oberkambrium des Oman (Andam-Formation) und, in relativ enger Assoziation, auch in gleich alten Schichten in Spanien und Wales.[8] Jedoch tritt Cruziana auch in Ablagerungen auf, die jünger sind oder einem anderen Ablagerungsmilieu entstammen als die, in denen Körperfossilien von Trilobiten auftreten. So ist Cruziana auch in Süßwasserablagerungen[9] und in triassischen Sedimenten[10] oder beidem[11] nachgewiesen.
Eine aktuo-paläontologische Arbeit zeigt, dass Cruziana-artige Spuren auch von dem Rückenschaler (Notostraca) Triops, dem „Urzeitkrebs“, erzeugt werden, und zwar, anders als Seilacher dies für Trilobiten vermutet, direkt auf der Sedimentoberfläche[12] (zu Cruziana in Süßwassersedimenten siehe auch Isopodichnus und Vorkommen).
Rusophycus
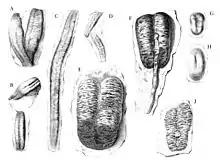


Seilacher betrachtet seit den 1970er Jahren alle größeren bilobaten Spuren mit Querstriemung, unabhängig davon ob bandartig oder nicht, als Cruziana.[5][7] Andere Bearbeiter unterscheiden jedoch bandartige Fortbewegungsspuren („Cruziana sensu strictu“) von nicht-bandartigen, ovalen („kaffebohnenförmigen“) bilobaten Ruhespuren (Cubichnia), die sie in das Ichnogenus Rusophycus einordnen. Seilacher fasst die bandartigen Formen lediglich informell unter dem Attribut ‚cruzianaeform‘ zusammen, und die ovalen kurzen bezeichnet er als ‚rusophyciform‘.
Der Grund für Seilachers Synonymisierung besteht darin, dass er Cruziana nicht nur als Fortbewegungs-, sondern vor allem auch als Fress- bzw. Weidespur (Pascichnion) betrachtet und Rusophycus nicht nur als Ruhespur, sondern auch als horizontal stationäre Fressspur.[7] Vor allem die Ichnospezies Rusophycus leiferikssoni (bzw. Cruziana leiferikssoni), die einen verhältnismäßig tiefen Bau repräsentiert, lässt diese Sicht plausibel erscheinen.[7][13] Für den kaum weniger bedeutenden Palichnologen Richard Bromley ist diese Synonymisierung jedoch unzulässig, da Cruziana und Rusophycus offensichtlich auf unterschiedliche Verhaltensweisen des Spurenerzeugers (horizontale Bewegung vs. ortsfestes „Graben“) zurückzuführen sind, was klar eine Zuordnung zu verschiedenen Spurengattungen rechtfertige.[14]
Interessanterweise repräsentiert das Fossilmaterial, auf Basis dessen die Spurengattung Cruziana seinerzeit errichtet wurde,[2] tatsächlich eher Formen, die nicht als bandartig, sondern als oval bis kaffebohnenförmig anzusprechen sind und damit dem aktuellen Konzept der Gattung Rusophycus näher kommen als dem aktuellen Konzept der Gattung Cruziana (sensu strictu). Rusophycus selbst wurde im Jahr 1852 vom US-Amerikaner James Hall in der Annahme errichtet, es handle sich um Pflanzen.[15] Das Typusmaterial aus der Clinton-Gruppe (frühes Silur) von Oneida County im US-Bundesstaat New York, umfasst sowohl die ovalen Formen, die dem tatsächlichen heutigen Verständnis der Gattung entsprechen, als auch die bandartigen Formen, die heute stets als Cruziana bezeichnet werden. Daher wurden lange Zeit die Namen Cruziana und Rusophycus sowie Bilobites von verschiedenen Bearbeitern für die gleichen Fossilien verwendet, bis in den 1950ern ausgerechnet durch Seilacher jene Trennung der beiden Gattungskonzepte vorgenommen wurde,[5] die heute von den meisten Palichnologen, nicht aber von Seilacher, akzeptiert wird.
Isopodichnus

Bilobate Spuren mit höchstens einem Zentimeter Breite, die in kontinentalen Sedimenten des höheren Paläozoikums und der Trias auftreten, werden traditionell unter dem Gattungsnamen Isopodichnus zusammengefasst. Da dieses traditionelle Konzept der Spurengattung aber sowohl cruzianaeforme bandartige Spuren als auch rusophyciforme „Ruhespuren“ beinhaltet, also Spuren die durch jeweils unterschiedliche Tätigkeiten entstanden sind, jedoch durch jeweils die gleichen Tätigkeiten entstanden sind, auf die auch Cruziana bzw. Rusophycus zurückgehen, werden sie von Bromley bei den beiden letztgenannten Spurengattungen eingeordnet. Bromley und Kollegen liefern ferner überzeugende Hinweise darauf, dass die triassischen Formen von „Isopodichnus“ von Kleinkrebsen aus der Gruppe der Rückenschaler (Notostraca) erzeugt wurden.[11] Jüngere Arbeiten weisen auch permische Isopodichnus der Tätigkeit dieser Kleinkrebse zu.[16]
Vorkommen
Cruziana (einschließlich Rusophycus) ist, bei insgesamt weltweiter Verbreitung, ein typisches Spurenfossil flachmariner Sandsteine des Paläozoikums, kommt aber bisweilen auch in flachmarinen Kalksteinen vor.[4]
Große, „robuste“ Cruziana sind bis in die Trias nachgewiesen, bislang jedoch ausschließlich in marinem Milieu.[10] In terrestrischem Milieu treten nur die kleinen, sowohl cruzianaeformen als auch rusophyciformen, traditionell „Isopodichnus“ genannten Spuren auf. Diese sind u. a. aus dem devonischen kontinentalen Old-Red-Sandstein Schottlands,[17] aus oberkarbonischen Schmelzwasserseesedimenten der Agua-Colorada-Formation (Argentinien)[18] und Seaham-Formation (New South Wales, Australien),[19] aus der Abo-Formation (Unterperm von New Mexico)[9] und aus dem „Saxon“ der permo-mesozoischen südwestlichen Umrandung des französischen Zentralmassivs (Lodève-Becken)[16] sowie aus diversen triassischen Ablagerungen, unter anderem dem deutschen Buntsandstein[11][20] und aus der Obertrias der Newark-Supergruppe[21] bekannt.
Ein relativ häufiges Auftreten von Cruziana sensu strictu und Rusophycus in tiefmarinen Ablagerungen ist aus dem Kambrium und Ordovizium von Ost-Kanada vermeldet worden. Post-ordovizische tiefmarine Vorkommen dieser Spurengattungen sind hingegen sehr selten.[22]
Cruziana-Ichnostratigraphie
Cruziana ist das erste Spurenfossil, auf dem basierend ein detailliertes biostratigraphisches Schema entwickelt wurde. Maßgeblich an dessen Entstehung beteiligt war einmal mehr Adolf Seilacher. Forciert wurde die Entwicklung unter anderem dadurch, dass kontinentale und randmarine Sandsteine der sogenannten nubischen Fazies, die große Teile der paläozoisch-mesozoischen Abfolgen in den alten Sedimentbecken Nordafrikas und des Nahen Ostens ausmachen, wichtige potenzielle Speichergesteine für Erdöl- und Erdgas sind. Die Cruziana-Ichnostratigraphie gestattet zumindest für die älteren, vollmarinen, körperfossilarmen Schichtglieder dieser Abfolgen eine grobe relative Alterseinstufung, was die Erkundung der geologischen Verhältnisse in diesen Sedimentbecken erleichtert. Aufgrund der geographischen Isolierung der Nordkontinente Laurentia („Paläo-Nordamerika“), Baltica („Paläo-Europa“) und Sibiria („Paläo-Mittelsibirien“) im frühen Paläozoikum ist der Anwendungsbereich dieses Schemas aber auf Gondwana (Südamerika, Afrika, Indien, Australien) und perigondwanische Terrane (wie Avalonia oder Süd-China) beschränkt.[23] Jedoch scheint auch für Baltica die relative Datierung von Sedimentgesteinen mit Hilfe von Cruziana zumindest im höheren Kambrium und unteren Ordovizium möglich.[24]
Die Eignung von Cruziana als Leitfossil beruht auf der relativ hohen Diversität der Striemungsmuster auf den Loben der Spuren, wodurch zahlreiche Cruziana-Ichnospezies oder sogar -Subspezies unterschieden werden können, die jeweils nur in einem relativ kurzen stratigraphischen Intervall vorkommen.[23] Da die Striemungsmuster als „Fingerprinting“ der Spurenerzeuger gelten, liegt hier eine relativ enge Korrelation zwischen der stratigraphischen Abfolge der Ichnospezies und dem evolutiven Wandel der Anatomie der Erzeuger vor. Folglich handelt es sich bei der Cruziana-Ichnostratigraphie um eine Art „Stellvertreter-Biostratigraphie“. Ein ähnlich enger Zusammenhang zwischen der Neuentstehung und dem Aussterben von Arten im Laufe der Entwicklungsgeschichte und dem Wandel in der Spurenüberlieferung existiert bei Tetrapoden und ihren fossilen Fährten (siehe dazu auch Fährtensandstein). Eine große Schwäche des Schemas ist, dass viele der darin enthaltenen Cruziana-Arten bislang nur aus einer einzigen Lokalität bekannt sind.
Die untenstehende Tabelle zeigt die aktuelle Version der Cruziana-Ichnostratigraphie.[23] Es ist zu beachten, dass, wie von Seilacher mittlerweile allgemein praktiziert, cruzianaeforme und rusophyciforme Spuren mit identischem „Fingerprint“ in der gleichen Ichnospezies vereint sind. Ichnospezies, die in mehr als einer Lokalität vorkommen, sind mit einem Sternchen gekennzeichnet.
| Karbon | |
|---|---|
| Unterkarbon | Oberkarbon |
|
|
| Devon | ||
|---|---|---|
| Unterdevon | Mitteldevon | Oberdevon |
|
|
|
| Silur | |
|---|---|
| Untersilur | Obersilur |
|
|
| Ordovizium | |
|---|---|
| Unterordovizium | Oberordovizium |
|
|
| Kambrium | ||
|---|---|---|
| Unterkambrium | Mittelkambrium | Oberkambrium |
|
|
|
Weblinks
Einzelnachweise
- Sven O. Egenhoff, Bernd Weber, Oliver Lehnert, Jörg Maletz: Biostratigraphic precision of the Cruziana rugosa group: a study from the Ordovician succession of southern and central Bolivia. Geological Magazine. Bd. 144, Nr. 2, 2007, S. 289–303, doi:10.1017/S0016756807003093.
- Alcide Dessalines d’Orbigny: Voyage dans l’Amerique méridionale (le Brésil, la République orientale de l’Uruguay, la République Argentine, la Patagonie, la République du Chili, la République de Bolivia, la République du Pérou), exécuté pendant les années 1826, 1827, 1828, 1829, 1830, 1831, 1832, et 1833. 3. Band, 4. Teil, Paris/Strasbourg 1842, S. 30 (archive.org).
- G. Winston Sinclair: The Generic Name Bilobites. In: Journal of Paleontology. Bd. 25, Nr. 2, 1951, S. 228–231, JSTOR 1299805.
- Olev Vinn: Cruziana traces from the Late Silurian (Pridoli) carbonate shelf of Saaremaa, Estonia. In: Estonian Journal of Earth Sciences. Bd. 63, Nr. 2, 2014, S. 71–75, doi:10.3176/earth.2014.06.
- D. Christopher A. Stanley, Ron K. Pickerill: Systematic ichnology of the Late Ordovician Georgian Bay Formation of Southern Ontario, eastern Canada. In: Royal Ontario Museum Life Sciences Contributions. Bd. 162, 1994, doi:10.5962/bhl.title.53483, S. 10 ff.
- Guillermo F. Aceñolaza, Juan Pablo Milana: Remarkable Cruziana beds in the Lower Ordovician of the Cordillera Oriental, NW Argentina. In: Ameghiniana. Bd. 42, Nr. 3, 2005, scielo.org.
- Adolf Seilacher: Trilobite Burrows. In: Trace Fossils Analysis. Springer-Verlag, Berlin/Heidelberg 2007, ISBN 978-3-540-47225-4, S. 31 ff.
- R. A. Fortey, A. Seilacher: The trace fossil Cruziana semiplicata and the trilobite that made it. In: Lethaia. Bd. 30, Nr. 2, 1997, S. 105–112, doi:10.1111/j.1502-3931.1997.tb00450.x.
- Nicholas J. Minter, Spencer G. Lucas: The arthropod trace fossil Cruziana and associated ichnotaxa from the lower Permian Abo Formation, Socorro County, New Mexico. In: Virgil Lueth, Spencer G. Lucas, Richard M. Chamberlin (Hrsg.): Geology of the Chupadera Mesa. New Mexico Geological Society Fall Field Conference Guidebook. Bd. 60, 2009, S. 291–298 (PDF 8,80 MB).
- John-Paul Zonneveld, S. George Pemberton, Thomas D. A. Saunders, Ronald K. Pickerill: Large, Robust Cruziana from the Middle Triassic of Northeastern British Columbia: Ethologic, Biostratigraphic, and Paleobiologic Significance. In: PALAIOS. Bd. 175, Nr. 5, 2002, S. 435–448, doi:10.1669/0883-1351(2002)017<0435:LRCFTM>2.0.CO;2 JSTOR 3515719.
- Richard G. Bromley, Ulla Asgaard: Triassic freshwater ichnocoenoses from Carlsberg Fjord, East Greenland. In: Palaeogeography, Palaeoclimatology, Palaeoecology. Bd. 28, 1979, S. 39–80, doi:10.1016/0031-0182(79)90112-3.
- Grzegorz Sadlok: New data on the trace fossil, Cruziana semiplicata (Furongian, Wiśniówka Sandstone Formation, Poland): origin, ethology and producer. In: Annales Societatis Geologorum Poloniae. Bd. 84, 2014, S. 35–50, asgp.pl.
- M. Gabriela Mángano, Luis A. Buatois: Rusophycus leiferikssoni Bergströn en la Formación Campanario implicancias en la paleoecología de planicies mareales cámbricas. Asociación Paleontológica Argentina, Publicación Especial. Bd. 9 (IV Reunión Argentina de Icnología y II Reunión de Icnología del Mercosur), 2003, S. 65–84 (apaleontologica.org.ar).
- Richard G. Bromley: Spurenfossilien: Biologie, Taphonomie und Anwendungen. Springer, Berlin/Heidelberg 1999, ISBN 978-3-540-62944-3, S. 178
- James Hall: Paleontology of New York. Volume 2. Containing Descriptions of the Organic Remains of the Lower Middle Division of the New York System (Equivalent in Part to the Middle Silurian Rocks of Europe). C. van Benthuysen, Albany NY 1852, S. 23 (archive.org)
- G. Gand, J. Garric, J. Schneider, H. Walter, J. Lapeyrie, C. Martin, A. Thiery: Notostraca trackways in Permian playa environments of the Lodève basin (France). In: Journal of Iberian Geology. Bd. 34, Nr. 1, 2008, S. 73–108 (online)
- N. H. Trewin, M. F. Thirlwall: Old Red Sandstone. In: Nigel H. Trewin (Hrsg.): The Geology of Scotland. 4th Edition. The Geological Society, London 2002, ISBN 1-86239-105-X, S. 228, 236
- Luis A. Buatois, M. Gabriela Mángano: Trace fossils from a carboniferous turbiditic lake: Implications for the recognition of additional nonmarine ichnofacies. In: Ichnos. Bd. 2, Nr. 3, 1993, S. 237–258, doi:10.1080/10420949309380098 (alternativer Volltextzugriff: Researchgate).
- Martin F. Glaessner: Palaeozoic arthropod trails from Australia. In: Paläontologische Zeitschrift. Bd. 31, Nr. 1–2, 1957, S. 103–108, doi:10.1007/BF02988968.
- J. E. Pollard: Isopodichnus, related arthropod trace fossils and notostracans from Triassic fluvial sediments. In: Transactions of the Royal Society of Edinburgh: Earth Sciences. Bd. 76, Nr. 2–3, 1985, S. 273–285, doi:10.1017/S026359330001049X
- David L. Fillmore, Michael J. Szajna, Spencer G. Lucas, Brian W. Hartline, Edward L. Simpson: Ichnology of a Late Triassic Lake Margin: The Lockatong Formation, Newark Basin, Pennsylvania. New Mexico Museum of Natural History Bulletin 76. Albuquerque 2017 (GoogleBooks; eine kleinere aber deutlich schlechtere Version wird bei New Mexico Digial Collections vorgehalten), S. 18, 39 f.
- Ron K. Pickerill: Deep-water marine Rusophycus and Cruziana from the Ordovician Lotbinière Formation of Quebec. In: Atlantic Geology. Bd. 31, Nr. 2, 1995, S. 103–108, hil.unb.ca.
- Adolf Seilacher: Cruziana Stratigraphy. In: Trace Fossils Analysis. Springer-Verlag, Berlin/Heidelberg 2007, ISBN 978-3-540-47225-4, S. 187 ff.
- Dirk Knaust: Cambro-Ordovician trace fossils from the SW-Norwegian Caledonides. In: Geological Journal. Bd. 39, Nr. 1, 2004, S. 1–24, doi:10.1002/gj.941.