Elasmotherium
Elasmotherium ist eine ausgestorbene Gattung der Nashörner, die im nördlichen Eurasien vorkam und zur Gruppe der Elasmotheriini gehört, einer Entwicklungslinie der Nashörner mit in der Regel nur einem Horn. Die Gattung trat erstmals im Oberen Miozän auf und überlebte mindestens bis zum Beginn des Jungpleistozäns. Vermutlich starb sie während der letzten Kaltzeit aus. Ihr Verbreitungsgebiet umfasste die Steppengebiete und Flussuferlandschaften Osteuropas sowie Zentral- und Ostasiens.
| Elasmotherium | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Elasmotherium caucasicum | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberes Miozän bis Jungpleistozän | ||||||||||||
| ? Mio. Jahre bis 50.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Elasmotherium | ||||||||||||
| Fischer, 1808 | ||||||||||||
Elasmotherium stellt einen der größten bekannten Vertreter der Nashörner dar. Die Tiere waren massig gebaut und an die offenen Regionen angepasst. Ein herausragendes Merkmal findet sich in der kuppelartigen Knochenaufwölbung auf dem Stirnbein des Schädels, auf der ein einzelnes Horn aufsaß. Aufgrund fehlender Fossilerhaltung ist dabei unklar, wie massiv dieses Horn ausgebildet war. Zudem besaßen die Tiere das in ihrer Zahnanzahl am stärksten reduzierte Gebiss aller Nashörner. Die Zähne waren extrem hochkronig und auf der Kauoberfläche durch auffällig gefalteten Zahnschmelz charakterisiert. Im gesamten Gebissaufbau kann Elasmotherium somit als äußerst spezialisiert angesehen werden, was vor allem für die stammesgeschichtlich jüngsten Angehörigen der Gattung zutrifft.
Über die Lebensweise ist wenig bekannt. Die Tiere lebten wahrscheinlich in offenen Landschaften, die mit wasserreichen Gebieten durchsetzt waren. Dort ernährten sie sich wohl von Sumpfpflanzen. Insgesamt treten die einzelnen Arten von Elasmotherium im Vergleich zu anderen, gleichzeitig bestehenden Nashornformen wie dem Wollnashorn eher selten im Fossilbericht auf. Die Gattung wurde erstmals im Jahr 1808 anhand eines Unterkiefers beschrieben. Ihre Zugehörigkeit zu den Nashörnern wurde erst später erkannt. Der Gattungsname setzt sich aus den griechischen Wörtern έλασμα (elasma) für „Platte“ und θηρίον (thēríon) für „Tier“ zusammen und bezieht sich auf die besondere Struktur der Zähne. Es werden heute fünf Arten anerkannt.
Merkmale
Elasmotherium gehört zu den am wenigsten untersuchten fossilen Nashorngattungen.[1] Insgesamt handelte es sich um sehr robuste und große Nashörner. Die Gliedmaßen waren im Verhältnis zum Körper relativ lang.[2] Die Vorderbeine endeten wie bei den rezenten Nashörnern in je drei Zehen (Metacarpus II – IV), wobei die mittlere am größten war. Ein vierter Zeh (Metacarpus V) war lediglich rudimentär ausgebildet. Auch die Hinterbeine besaßen je drei Zehen. Der weitere Körperbau war relativ kräftig: So erreichten die Dornfortsätze an den Brustwirbeln, an denen die massive Muskulatur zur Stabilisierung des Kopfes ansetzte, Längen von über 50 cm.[3][4]

Der Schädel war ebenfalls sehr massiv und groß mit einem robusten Unterkiefer. Die Breite erreichte in der Regel weniger als 50 % der Länge, so dass er eher schmal wirkte. Der Gesichtsschädel übertraf den hinteren Schädelabschnitt an Länge. Das Hinterhauptbein war lang und ausladend und besaß einen stark ausgebildeten Knochenkamm. Das in Seitenansicht deutlich gestreckte und dadurch gewinkelte Hinterhaupt bewirkte, dass das Tier den Kopf schräg nach unten hielt, ähnlich wie es heute beim Breitmaulnashorn (Ceratotherium simum) der Fall ist.[5][6] Bezüglich der beiden Geschlechter ist zumindest für die späten Vertreter von Elasmotherium ein Sexualdimorphismus nachweisbar, der bei Kühen zu einem schlankeren Schädelbau, mit einem weniger stark ausgeprägten Hinterhauptkamm führte.[7][3] Das Nasenbein war gegenüber dem breiten Hinterhaupt schlank gebaut, wodurch der Schädel in der Aufsicht deutlich keilartig erschien. Dazu korrespondierte eine am Unterkiefer langgezogene und schmale Symphyse. Weiterhin war die Nasenscheidewand beim erwachsenen Tier vollständig verknöchert. Ihre Breite betrug vorn 3,6 cm, verjüngte sich aber nach hinten. Das Merkmal kommt bei den heute lebenden Nashornarten nicht vor und ist fossil sonst nur bei der Gattung Coelodonta, hier vor allem beim Wollnashorn (Coelodonta antiquitatis), sowie teilweise von einigen Vertretern der Gattung Stephanorhinus bekannt. Da Elasmotherium aber mit den beiden Gattungen nicht in direkter Linie miteinander verwandt ist, muss sich dieses Charakteristikum unabhängig voneinander entwickelt haben (konvergente Evolution).[3][8][3][9]
Markant war eine dom- oder kuppelartige Erhöhung auf dem Stirnbein, die den Schädel um rund 15 cm überragte und einen Durchmesser von 25 bis 35 cm hatte. Bei weiblichen Individuen zeigte sie sich weniger prominent als bei männlichen. Die Erhöhung stellte die Hornbasis dar, was an zahlreichen kleinen Eintiefungen für Blutkanälchen ersichtlich wird und was der Oberfläche eine raue Struktur verlieh. Die Knochenwand selbst war relativ dünn und variierte von 0,8 bis 1,6 cm in der Dicke. Das Innere der Struktur wurde von der vergrößerten Stirnhöhle ausgefüllt, die mit dem Nasenraum in Verbindung stand. Das Horn war höchstwahrscheinlich wie bei allen Nashörnern aus Keratin aufgebaut, aufgrund einer fehlenden Fossilüberlieferung ist über sein Aussehen nichts bekannt. Die Ausmaße der Hornbasis ließen einige Autoren an ein massives und großes Horn denken. Eine umfassende Analyse aus dem Jahr 2021 kommt dagegen zu einem anderen Schluss. Insgesamt unterscheidet sich die Oberfläche der kuppelartigen Erhöhung bei Elasmotherium von den anderen horntragenden Nashörnern, deren Hornansatzstelle blumenkohlartig aufgeraut ist. Vielmehr ähnelt sie dem mit Blutkanälchen durchzogenen knöchernen Hornunterbau bei zahlreichen Hornträgern. Dadurch kann angenommen werden, dass die kuppelartige Erhöhung bei Elasmotherium eher von einem dünnen Keratinmantel überzogen war, der primär als Schutz für die dünne Knochenwand diente. Da bei den Schädeln männlicher Individuen die kräftigeren Aufwölbungen mit intensiveren Aufrauhungen verbunden sind, als dies bei den weiblichen der Fall ist, war bei ersteren der Keratinschutz möglicherweise massiver gewesen. Insgesamt unterscheidet sich bei Elasmotherium die Lage der Kuppel auf der Stirn deutlich von den rezenten einhörnigen Nashörnern wie dem Panzernashorn (Rhinoceros unicornis), die ihr Horn auf der Nase tragen.[5] Zusätzlich traten aber bei Elasmotherium am vorderen Ende des Nasenbeins zusätzlich aufgeraute Flächen auf. Hier ist ein weiteres, leicht erhabenes Keratinschild denkbar.[9]
Auch das Gebiss von Elasmotherium zeigte charakteristische Eigenschaften, da es stark reduziert war und lediglich aus zwei Vormahlzähnen und drei Mahlzähnen je Kieferbogen bestand. Entsprechend lautet die Zahnformel für das Dauergebiss: . Das Vorhandensein der Alveolen der Schneidezähne im allerdings rudimentären Zustand lässt annehmen, dass Schneidezähne im Milchgebiss der Jungtiere noch ausgebildet waren. Das Fehlen der Schneidezähne im Dauergebiss wurde durch flache plattenartige Knochenbildungen am Ober- und Unterkiefer ausgeglichen, die möglicherweise lippenartige Bildungen darstellten und als Organ zum Zupfen von Gras gedeutet werden, ähnlich wie es bei den rezenten afrikanischen Nashörnern der Fall ist.[5][3] Bemerkenswert sind die Molaren, die eine deutlich rechteckige Form mit markant gefaltetem Zahnschmelz besaßen, wobei die Faltenlinien bei den jüngeren Vertretern zusätzlich noch einen sinusförmigen Verlauf aufwiesen. Die starke Faltung führte zur Namensgebung der Gattung. Zudem waren die Molaren äußerst hochkronig und wurzellos (euhypsodont). Die extreme Hochkronigkeit ist die stärkste bisher bekannte Spezialisierung in der Familie der Nashörner und der Ordnung der Unpaarhufer und wird unter den Säugetieren lediglich von einigen Vertretern der Nagetiere übertroffen.[10][11][3]

Lebensweise

Elasmotherium ist eines der größten bekannten Säugetiere des Pliozäns und des Pleistozäns in Eurasien. Aufgrund seines massigen Körpers dürfte es somit an produktive, nährstoffreiche Landschaften angepasst sein, etwa an langgrasige Steppen oder an Uferrandgebiete. Die hochkronigen Zähne mit dem auffallend gefalteten Zahnschmelz verweisen auf stark abrasives Nahrungsmaterial. Häufig werden derartige Zähne als eine Anpassung an Grasnahrung gedeutet. Da Gräser weniger Nährstoffe enthalten, muss ein Tier dementsprechend mehr fressen, was den Abrieb der Zähne beschleunigt. Dies wird zusätzlich noch durch einen hohen Kieselsäuregehalt verstärkt, was bei ausschließlich grashaltiger Nahrung die hohen Zahnkronen erforderlich macht.[6] Gegen eine vollständige Spezialisierung auf Grasnahrung spricht aber die schmale Schnauze von Elasmotherium. Diese war gut geeignet, um in feuchtem Untergrund nach Nahrungspflanzen zu suchen. Die Ansicht wird auch dadurch unterstützt, dass zumindest die späten Vertreter von Elasmotherium in weiten Teilen ihres Verbreitungsareals, so etwa im wasserreichen Südsibirien, in Auwäldern der Flussufer oder an Seeufern vorkamen. Dadurch ist es wahrscheinlich, dass sich die Tiere von Sumpfpflanzen ernährten, deren untere Abschnitte mit Erde angereichert waren und so einen hohen Mineralgehalt aufwiesen, der wiederum die Abrasion der Backenzähne bewirkte. Bei wurzelreicher Nahrung, die eventuell mitsamt der Erde aus dem Boden gerissen wurde, verstärkte sich dieser Effekt noch. Einige frühe Formen weisen aber ein Abnutzungsmuster an den Zähnen auf, die eine gemischte Pflanzenkost aus harten und weichen Komponenten annehmen lassen.[12] Insgesamt verweist die rekonstruierte tiefe Haltung des Kopfes auf eine überwiegende Nahrungssuche am Boden, bei der möglicherweise auch das vordere Keratinschild eingesetzt wurde.[9] Unterstützt wird die Ansicht einer überwiegend bodennahen Nahrungsressource von Isotopenanalysen, die für Elasmotherium extrem hohe Werte an Stickstoff und Kohlenstoff ergaben. Die Werte liegen außerhalb der Variation von anderen Nashörnern wie dem Wollnashorn und sind vergleichbar mit denen einiger lokaler Populationen der heutigen Saigas. Dadurch kann unter anderem eine verstärkte Konsumierung von Fuchsschwanzgewächsen und/oder von Wurzeln angenommen werden.[13] Im Gegensatz zu einigen anderen postulierten Meinungen[14][15] war Elasmotherium jedoch nicht an eine semi-aquatische Lebensweise angepasst. Gegen eine derartige Lebensweise sprechen die Körperproportionen allgemein und die langen Beine speziell. Diese stellen zumeist Anpassungen an offene Graslandschaften dar. Offenbar bewegten sich die Tiere auf den Wanderungen von ihren Nahrungsplätzen in den Auen- oder Uferrandgebieten durch angrenzende oder dazwischen liegende Steppenlandschaften. Somit kann der Lebensraum der Nashorngattung als offene Landschaft mit zahlreichen eingeschlossenen Wasserbiotopen angenommen werden.[16][3]
Über die weitere Lebensweise ist nur wenig bekannt. Die Lage des Horns auf der Stirn lässt vermuten, dass es nicht oder nur in geringem Maße in Dominanz- oder Territorialkämpfen eingesetzt wurde, im Gegensatz zu heutigen Nashörnern. Da sich bei heutigen Nashörnern das (größere) Horn auf der Nase befindet, kann es wesentlich effektiver verwendet werden, da über die Nackenmuskulatur bei der Bewegung des Kopfes mehr Wucht oder Schwung auf das Horn übertragen wird. Deshalb geht man davon aus, dass Kämpfe zwischen einzelnen Vertretern von Elasmotherium hoch ritualisiert waren. Darüber hinaus besaß das Horn selbst aufgrund seiner Lage wohl keine Funktion bei der Nahrungssuche, wie es bei heutigen Nashörnern teilweise der Fall ist.[5] Jedoch ist für die von Hohlräumen durchsetzte kuppelartige Erhöhung eine derartige Aufgabe durchaus wahrscheinlich. Durch die Verbindung mit der Nasenhöhle könnte so ein Erweiterungsraum für die Geruchsorgane bestanden haben, was das Auffinden der Nahrung erleichterte.[17][3] Als weitere Interpretation wird ein Resonanzraum für die Kommunikation angedacht, wodurch Bullen vermochten, mit Kühen in Kontakt zu treten oder ihren Revieranspruch gegen Konkurrenten auszudrücken.[9]
Natürliche Feinde hatte Elasmotherium aufgrund seiner Größe wahrscheinlich nicht. Allerdings weist der Schädel von Atyrau (früher Gurjew) eine runde Schädelverletzung im Stirnbereich auf, die jedoch zu Lebzeiten des Tiers wieder verheilt war. Zudem wird vermutet, dass die Tiere aufgrund ihrer relativ nördlichen Verbreitung möglicherweise ein Fell besessen hatten.[3]
Systematik
|
Verwandtschaftsverhältnis der Nashörner basierend auf genetischen Daten nach Liu et al. 2021[19]
|
Elasmotherium ist eine Gattung aus der Familie der Nashörner. Innerhalb der Familie gehört sie zur Unterfamilie der Rhinocerotinae und zur Tribus der Elasmotheriini. Die Tribus schließt Nashörner mit einem allgemein großen bis sehr großen Körperbau und extrem hohen Zahnkronen ein. Bezugnehmend auf letzteres Merkmal stellen die Elasmotherien zumindest in ihrer späten Entwicklungsphase die am stärksten spezialisierten Nashornvertreter dar. Sie bilden außerdem die Schwestergruppe der Rhinocerotini, zu denen wiederum die heute lebenden Nashornarten gestellt werden. Beide Nashornlinien können eine lange eigenständige Entwicklungsgeschichte vorweisen, da sich ihre Trennung molekulargenetischen Daten zufolge bereits im Paläogen vollzogen hatte. Der ermittelte Zeitraum reicht je nach Untersuchung vom Mittleren Eozän vor etwa 47 Millionen Jahren bis zum Unteren Oligozän vor 31 Millionen Jahren.[13][20][19] Als nächster Verwandter von Elasmotherium ist Sinotherium aufzufassen, beide Formen werden von einigen Wissenschaftlern zur Untertribus der Elasmotheriina zusammengefasst. Ihr charakteristisches Merkmal findet sich in der Hornbildung auf der Stirn. Den Elasmotheriina gegenüber stehen die Iranotheriina, bei denen es sich um stammesgeschichtlich ältere Elasmotherien mit einem Nasalhorn handelt. Zusätzlich weisen letztere generell nicht ganz so hochkronige Backenzähne und einen weniger gefalteten Zahnschmelz auf. Diese Untertribus werden neben dem namengebenden Iranotherium unter anderem Ningxiatherium, Parelasmotherium oder Hispanotherium beigeordnet. Die Unterscheidung von Elasmotheriina und Iranotheriina erfolgte erstmals durch den ungarischen Paläontologen Miklós Kretzoi (1907–2005) im Jahr 1943.[21][1]
Gegenwärtig sind fünf Arten aus der Gattung Elasmotherium anerkannt. Dazu gehören das weitgehend obermiozäne Elasmotherium primigenium, das ins Obere Pliozän datierende Elasmotherium chaprovicum, die beiden im Altpleistozän auftretenden Formen Elasmotherium caucasicum beziehungsweise Elasmotherium peii und das vom Mittel- bis Jungpleistozän lebende Elasmotherium sibiricum. Einige Autoren reduzieren die Artanzahl unter Verweis auf metrische Merkmale am Skelettmaterial auf nur zwei Arten, zumeist Elasmotherium caucasicum und Elasmotherium sibiricum, wobei die anderen beiden Arten als synonym zu ersterer angesehen werden,[7] allerdings wird dies häufig abgelehnt.[22]
Arten
Elasmotherium primigenium
Der bisher älteste und ursprünglichste Vertreter wird als Elasmotherium primigenium bezeichnet. Die Form wurde im Jahr 2021 von Sun Danhui und Kollegen anhand eines über 52 cm langen Teilschädels aus Dingbian in der chinesischen Provinz Shaanxi wissenschaftlich eingeführt. Der Fund gehört in das Obere Miozän. Auffallend sind die weniger deutliche Faltung des Zahnschmelzes und ein relativ gerade verlaufendes Ectoloph, eine Zahnschmelzleiste auf der Wangenseite der Mahlzähne. Wie auch die späteren Formen besaß Elasmotherium primigenium eine massive Hornbasis, die 18 cm breit sowie 24 cm lang war und sich gut 16 cm aufwölbte. Das ausgezogene Hinterhauptsbein gibt an, dass die Tiere ihren Kopf sehr tief trugen. Dies und die hochkronigen Zähne lassen eine überwiegende grashaltige Nahrung annehmen. Dadurch lebte die Art wohl in offenen Landschaften Ostasiens, die damals von frühen Pferdeformen wie Hipparion dominiert wurden. In seinen Merkmalen steht Elasmotherium primigenium vermittelnd zwischen den stammesgeschichtlich älteren Sinotherium und den jüngeren Arten von Elasmotherium.[23]
Elasmotherium chaprovicum
Elasmotherium chaprovicum stellt den nächstälteren Angehörigen der Nashorngattung dar, welcher im Oberen Pliozän vor 3,6 bis 2,6 Millionen Jahren im Gebiet des Schwarzen Meeres verbreitet war. Von dieser Art sind bisher nur wenige Knochen und Zähne bekannt. Sie gehört dem Chapri-Faunenkomplex an und ist mit frühen Vertretern des Südelefanten, dem Etruskischen Nashorn (Stephanorhinus etruscus) sowie Paracamelus vergesellschaftet. Typusfundstellen sind hier Chapri und Liwenzowka (beide Ukraine).[24][25] Anfangs wurde sie zur Art Elasmotherium caucasicum gezählt,[17][26] doch wurden schnell Unterschiede zu dieser Art erkannt.[16][27] Diese betreffen vor allem den Zahnapparat; so sind die Zähne in ihrer Größe mit denen von Elasmotherium caucasicum vergleichbar und deutlich größer als die des Elasmotherium sibiricum, unterscheiden sich jedoch von ersteren im Aufbau und der Struktur des Zahnschmelzes. Weiterhin sind die Gliedmaßen deutlich kürzer als die der kaukasischen Variante und erreichen die Maße des sibirischen Vertreters, sind aber deutlich massiver als bei diesem. Da aber bisher zu wenig Fundmaterial vorliegt, ist eine Abgrenzung zurzeit schwierig. Erstmals beschrieben wurde Elasmotherium chaprovicum von Anna Konstantinowna Schwyrjowa 2004 anhand von Zähnen aus Liwenzowka.[24]
Elasmotherium caucasicum
Elasmotherium caucasicum war der größte Vertreter der Gattung und erreichte eine Länge von 5 bis 5,2 m.[3] Das rekonstruierte Gewicht von 5 t ist vergleichbar mit dem heutiger ausgewachsener Asiatischer Elefanten, womit die Nashornart zu den größten des Pleistozäns gehörte.[28] Die Art ist aber bislang lediglich über einzelne Skelettelemente bekannt, darunter Zähne und Langknochen – vollständige Skelette wurden bisher nicht gefunden. Allgemein war der Körperbau ähnlich dem des Elasmotherium sibiricum. Die Gliedmaßen waren jedoch kräftiger gebaut und wesentlich länger, außerdem zeigten sie gegenüber dem sibirischen Verwandten noch einige altertümliche Merkmale. Im Vergleich mit den Arm- und Beinknochen anderer Nashörner fallen jene des Elasmotherium caucasicum allein durch die enorme Größe auf. So sind jene des Breitmaulnashorns rund 50 % kleiner und 30 % schmaler.[2] Die Zähne charakterisierten sich durch eine etwas geringere Kronenhöhe und ihre allgemeine Größe, während der Zahnschmelz zwar deutlich gefaltet war, die Faltenlinien aber noch nicht einen so markant sinusartigen Verlauf hatten, wie es beim späteren Elasmotherium sibiricum der Fall war. Bemerkenswert ist der letzte Vormahlzahn, der deutlich massiv ausfiel, was vermuten lässt, dass diese Art noch nicht so stark an ein Leben in offenem Gelände angepasst war wie das spätere Elasmotherium sibiricum.[16]
Die Nashornart trat in Zentralasien und im Schwarzmeergebiet auf und ist im jüngeren Altpleistozän nachgewiesen, sie kam hier vor 1,1 bis 0,8 Millionen Jahren vor. Ein relativ dichtes Nachweisgebiet befindet sich auf der Taman-Halbinsel zwischen Schwarzem Meer und dem Asowschen Meer.[2][16] Allein hier liegen wenigstens 230 Fossilfunde von Elasmotherium caucasicum vor, die klassische Fundstelle Sinaya Balka allein barg rund elf Individuen verschiedener Altersstufen. Eine Skelettrekonstruktion zusammengesetzt aus verschiedenen Individuen weist eine Schulterhöhe von 2,4 und eine Länge von 4,2 m auf. Die kaukasische Form bildete einen Bestandteil des Taman-Faunenkomplexes, der weiterhin den Südelefanten und den Taman-Bison umfasst.[25][12] Weitere Funde stammen unter anderem aus Salcia (Moldawien). Auch in Kasachstan ist die Nashornart belegt, so mit einigen Zähnen vom Dorf Lebjazhje am Irtysch.[29]
Beschrieben wurde die Art 1914 von Alexei Alexejewitsch Borissiak.[30] Als Grundlage der Beschreibung dienten mehrere große und morphologisch vom Elasmotherium sibiricum abweichende Zähne eines Individuums aus Sinaya Balka auf der Taman-Halbinsel am Asowschen Meer.[16]
Elasmotherium peii
Die Art Elasmotherium peii wurde zuerst von einigen Fundstellen des Nihewan-Beckens im Norden der chinesischen Provinzen Hebei und Shanxi sowie vom Huáng Hé im Süden von Shanxi bekannt,[31][2][32] später kamen noch Reste aus dem östlichen Europa dazu.[22] Es handelte sich um einen kleineren Vertreter, der hauptsächlich von Zähnen und einigen Extremitätenknochen belegt ist. Er erschien erstmals am Ende des Oberen Pliozäns in Ostasien, wo er unter anderem im Nihewan-Becken mit frühen Vertretern des Wollnashorns (Coelodonta nihowanensis) innerhalb des Nihewan-Faunenkomplexes auftrat. Im frühen Altpleistozän vor 1,6 Millionen Jahren verschwand er aber wieder.[16] Beschrieben wurde die Art im Jahr 1958 von Chow Minchen unter Berufung auf eine obere linke Zahnreihe mit den erhaltenen Zähnen vom zweiten Milchprämolar über den dritten dauerhaften Prämolaren bis zum dritten dauerhaften Molaren. Im gleichen Aufsatz etablierte Chow mit Elasmotherium inexpectatum noch eine weitere eigenständige Art anhand eines einzelnen oberen Molars.[31] Sie wird heute aber zu Elasmotherium peii gestellt. Einige der Funde waren bereits in den 1930er und 1940er Jahren von Teilhard de Chardin in Nordchina gesammelt worden.[33][3]
Zu Beginn des 21. Jahrhunderts hoben einige Wissenschaftler unter Berufung auf Analysen an den Zähnen und an den vorher nicht näher dokumentierten Langknochen den Artstatus von Elasmotherium peii auf und sahen die Form als identisch mit Elasmotherium caucasicum an.[16][2] Neuere, sehr gut erhaltene Funde, etwa Zähne und Fußknochen unter anderem aus Shanshenmiaozui und Heitugou im Nihewan-Becken, ermöglichten im weiteren Verlauf, beide Vertreter wieder auf Artebene zu trennen. Demnach besitzt Elasmotherium peii weniger hochkronige Zähne, ein dickeres Zahnschmelzmuster und deutlich ausgebildete Zahnwurzeln.[32]
Elasmotherium sibiricum
Die bekannteste und bisher am häufigsten aufgefundene Art stellt Elasmotherium sibiricum dar. Die Tierart erreichte eine Länge von 4 bis 4,5 m (ohne Schwanz), bei einer Widerristhöhe von 2 m. Allein der Schädel maß 95 cm.[17][3] Mit einem errechneten Körpergewicht von 4 t[28] war sie etwas kleiner als das stammesgeschichtlich ältere Elasmotherium caucasicum. Weitere Unterschiede zu dieser Art bestehen darin, dass Elasmotherium sibiricum im Verhältnis zum Körper längere Beine hatte. Auch war die Zahnreihe deutlich kürzer ausgebildet, wobei der letzte Prämolar relativ grazil wirkte. Die Zähne besaßen stark gefalteten Zahnschmelz mit deutlich geschwungenen Faltenlinien und waren deutlich hochkroniger. In Relation zu Elasmotherium caucasicum waren die Zähne aber merklich kleiner.[24][29][16]
Das Verbreitungsgebiet von Elasmotherium sibiricum reichte vom Schwarzen Meer bis in das östliche Kasachstan. Die südlichste Verbreitungsgrenze lag ungefähr beim 44. Breitengrad im heutigen Usbekistan, während die Art im Norden bis nach Sibirien an die Mündung der Kama nahe der Stadt Irbit zum 57. Breitengrad vorstieß. Insgesamt sind bisher über 60 Fundstellen bekannt, von denen etwa die Hälfte in Kasachstan liegt. Dabei wurden meist nur einzelne Knochen und Zähne gefunden – bis heute gibt es kein vollständiges Skelett dieser Tierart, so dass einzelne Skelettelemente immer noch unbekannt sind. Das bisher am vollständigsten erhaltene Skelett stammt aus Selenokumsk (Russland) im nördlichen Kaukasusvorland, wo es 1966 in einer Ziegeleigrube am rechten Ufer der Kuma gefunden wurde. Das Skelett, dessen Reste auf einer Fläche von 10 m² verstreut lagen, umfasst einen vollständigen Schädel, Teile der Wirbelsäule, Rippen und Vorder- und Hintergliedmaßen. Nur 200 m entfernt kamen 1968 zwei weitere Individuen dieser Nashornart zum Vorschein. Bereits 1964 war bei Stawropol, ebenfalls im Süden Russlands gelegen, ein relativ vollständiges Skelett gefunden worden, welches später im dortigen Museum als Skelettmontage ausgestellt wurde. Einer der umfangreichsten Fundkomplexe von Elasmotherium sibiricum fand sich bereits 1938 am rechten Ufer des Großen Karaman, einem Nebenfluss der Wolga. Er besteht aus mehreren Individuen, darunter ein teilweise vollständiges Skelett. In relativer Nähe dazu liegt die Fundstelle Irgis 1 am Ufer des Großen Irgis, ebenfalls ein Nebenfluss der Wolga. Dort fanden sich Überreste von wenigstens sieben Individuen aller Altersstufen. Die geologisch-paläontologischen Daten sprechen für einen abrupten Tod der Tiere bei rascher Abdeckung der Kadaver. Analysen der Abnutzungsspuren der Zähne verweisen darauf, dass die Tiere in ihrer letzten Lebensphase von der allgemeinen Grasnahrung auf eine weiche Pflanzenkost wechselten. Die ungewöhnliche Ernährungsumstellung erklären die Wissenschaftler mit einem harschen Dsud, verbunden mit kräftigem Schneefall. Die dadurch unzugängliche Grasnahrung zwang die Tiere auf Blätter und Zweige auszuweichen, was zu Mangelernährung und letztendlich zu ihrem Tod führte.[34] Des Weiteren gibt es mehrere Schädelfunde: Ein Schädel stammt von der linken Uferterrasse der Sakmara bei Tscheljabinsk (Russland), allein vier weitere wurden bei Lutschka nahe der Wolgamündung (ebenfalls Russland) gefunden und zwei sind von kasachischen Fundstellen bei Atyrau und bei Nur-Sultan (bis 2019 Astana) überliefert.[4][17][7][29]
Erstmals aufgetreten ist Elasmotherium sibiricum im Mittelpleistozän, es ging vermutlich aus Elasmotherium caucasicum hervor. Vor allem in Zentralasien war es Bestandteil des Koshkurgan-Faunenkomplexes, welcher unter anderem den Südelefanten, den Schoetensack-Bison, das Waldnashorn sowie das Mosbacher Pferd umfasst.[35][29] In Osteuropa gehört es dem spätmittelpleistozänen Chazar-Faunenkomplex des Wolga-Don Gebietes an und kommt hier zusammen mit dem Steppenmammut und dem Waldnashorn vor.[25][36] Der jüngste Fund in Osteuropa stammt von der Wolga-Insel Choroschewski, die mit der Fertigstellung des Saratower Stausees geflutet wurde, und datiert in die Mikulino-Warmzeit (Synonym für die Eem-Warmzeit; vor 128.000 bis 115.000 Jahren).[37][3] Der dort gefundene Oberkiefermolar weist zudem einige Besonderheiten auf, die aufgrund fehlender, ähnlich gearteter Vergleichsfunde als pathologische Veränderungen angesehen werden.[37] In seinen östlichsten Verbreitungsgebieten überlebte Elasmotherium sibiricum aber länger. So kamen Funde aus der Höhle Smelowskaja II im südwestlichen Sibirien zu Tage, sie sind mit dem Wollhaarmammut und dem Wollnashorn vergesellschaftet und weisen radiometrischen Daten zufolge ein Alter von mehr als 50.000 Jahren auf. Weitere spätpleistozäne Knochenreste sind aus Batpak und Voronovka belegt, ebenfalls in Südwestsibirien.[38][36][39] Das Alter von Funden nahe dem Ort Kozhamzhar in der Region Pavlodar in Kasachstan, darunter neben einem Schädel von Elasmotherium sibiricum auch Reste des Steppen- und des Wollhaarmammuts, wurde auf 27.500 bis 29.000 Jahre datiert und könnte somit den Zeitpunkt des Aussterbens deutlich gegenüber den bis dahin angenommenen Daten verschieben.[40] Es bestehen allerdings methodische Probleme bei der Messung, so dass die Werte vorerst als unsicher gelten müssen.[41] Im Vergleich dazu ergaben 19 analysierte Knochenreste aus dem westlichen Asien und östlichen Europa Radiocarbon-Daten zwischen 50.000 und 36.520 Jahren. Demnach wäre Elasmotherium sibiricum noch deutlich vor dem Maximum der letzten Kaltzeit verschwunden, sein Aussterben rückt aber relativ nahe an den Höhepunkt der Quartären Aussterbewelle heran.[13]
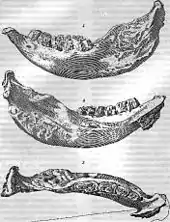
Die wissenschaftliche Erstbeschreibung der Gattung Elasmotherium mit der Art Elasmotherium sibiricum erbrachte der deutsche Zoologe Gotthelf Fischer von Waldheim (1771–1853). Sie erschien 1808 als kurzer Aufsatz, der im Jahr darauf noch einmal in einem größeren Rahmen mit den gleichen Abbildungen veröffentlicht wurde.[42] Die Grundlage der Beschreibung bildete ein Unterkiefer aus dem Naturkundemuseum in Moskau, welcher zur Fossiliensammlung der Prinzessin Jekaterina Daschkowa gehörte. Daschkowa hatte die Sammlung drei Jahre zuvor der Universität Moskau geschenkt mit dem ausdrücklichen Wunsch, sie der Öffentlichkeit zugänglich zu machen.[43] Der Unterkiefer gilt als Holotyp, während ein wesentlich vollständigerer, 97 cm langer und bereits 1877 aufgefundener Schädel von Lutschka als Lectotyp angesehen wird.[7] Ein ebenfalls in der Erstbeschreibung vorgestellter Zahn von Elasmotherium ging im Jahr 1812 verloren.[43] Im Laufe der Forschungsgeschichte gab es verschiedene Bezeichnungen für die Art Elasmotherium sibiricum:[17]
- Elasmotherium fischerii Desmarest, 1820
- Elasmotherium keyserlingii Fischer von Waldheim, 1842
- Stereocerus galli Duvernoy, 1855
- Enigmatherium stavropolitanum Pavlova, 1916.
Elasmotherium fischerii ist eine von Anselme Gaëtan Desmarest im Jahr 1820 benutzte Bezeichnung, die sich auf Fischer von Waldheims Beschreibung stützt.[44] Sie wurde im 19. Jahrhundert mehrfach gebraucht,[45] ist aber aufgrund der späteren Benennung ungültig. Die Form Elasmotherium keyserlingii bezieht sich auf Alexander Graf Keyserling und beruht auf einem Zahn vom Kaspischen Meer, den Keyserling im Jahr 1841 von einem kirgisischen Fürsten geschenkt bekam. Fischer von Waldheim etablierte dann im folgenden Jahr den Namen zu Ehren von Keyserling,[46][43] Johann Friedrich von Brandt vereinte ihn aber 1864 wieder mit Elasmotherium sibiricum.[45] Die Beschreibung von Enigmatherium stavropolitanum basiert auf einem einzelnen Zahn aus dem nördlichen Kaukasusgebiet, der jedoch keine Merkmalsunterschiede zu Elasmotherium sibiricum aufweist, weswegen diese Bezeichnung nicht mehr verwendet wird. Problematisch ist die Art Stereocerus galli. Sie geht auf ein hinteres Schädelfragment aus der Naturaliensammlung des Arztes Franz Joseph Gall (1758–1828) zurück, dessen Herkunft mit „Rheintal“ angegeben war.[47] Dies führte zu der Annahme, Elasmotherium sibiricum wäre bis nach Mittel- oder Westeuropa verbreitet gewesen.[48] Jedoch ist die tatsächliche Herkunft dieser forschungsgeschichtlich frühen Sammlungsfunde unklar, ebenso wie bei ähnlich alten Elasmotherium-Fossilien aus Ungarn oder Italien.[17][33][3]
Stammesgeschichte
Ursprung und Entwicklung
Die Gattung Elasmotherium erschien erstmals im Oberen Miozän Asiens und entwickelte sich möglicherweise aus Sinotherium, das gleichfalls vom Oberen Miozän bis zum Unteren Pliozän vorkam. Beide Gattungen bilden eine Klade, die – zusammen mit ihrer Schwesterklade Parelasmotherium und Ningxiatherium und dem weiter außen stehenden Iranotherium – zur Tribus der Elasmotheriini gehört, die meistens der Unterfamilie Rhinocerotinae,[49] von einigen Wissenschaftlern aber auch einer eigenständigen Unterfamilie, den Elasmotheriinae, zugewiesen wird.[50][1] Alle genannten Nashörner zeichnen sich durch einen großen Körperbau, durch eine Tendenz zur Hochkronigkeit der Zähne mit stark gefaltetem Zahnschmelz und durch eine fehlende vordere Bezahnung aus. Diese Merkmale waren bei den frühesten Vertretern der Elasmotheriini im Mittleren Miozän noch nicht ausgebildet und kristallisierten sich erst im späten Miozän und frühen Pliozän heraus.[50] Dabei gehen diese Veränderungen, insbesondere die zunehmende Körpergröße und die Zahnmorphologie, mit einer stärkeren Ausbreitung offener Landschaften durch die weiter zunehmende Aridisierung des Klimas und der damit verbundenen Nahrungsumstellung auf Gräser einher. Eine deutliche Parallele ist dabei in der Entwicklung der eng verwandten Gattungen Stephanorhinus mit dem Steppen- und dem Waldnashorn (Stephanorhinus hemitoechus und Stephanorhinus kirchbergensis) und Coelodonta mit dem Wollnashorn (Coelodonta antiquitatis) während des Pleistozäns festzustellen, allerdings gehören diese zur Gruppe der Rhinocerotini und somit nicht in den unmittelbaren Verwandtschaftskreis von Elasmotherium. Auch hier kam es infolge der Versteppung der Landschaften zu einer Nahrungsumstellung und somit zu einer zunehmenden Hochkronigkeit der Backenzähne, die jeweils in Elasmotherium und in Coelodonta ihren Höhepunkt erreichte (bei Elasmotherium jedoch noch stärker ausgebildet war).[1][11][28] Möglicherweise war diese starke Spezialisierung auch ursächlich verantwortlich für das vergleichsweise frühe Aussterben von Elasmotherium, da die bisher vorliegenden Isotopenanalysen auf eine wenig flexible Ernährungsweise hindeuten. Demnach waren die Tiere wohl nur mäßig befähigt, auf die stärkeren Klimaschwankungen der letzten Kaltzeit verbunden mit einem wechselnden Angebot an Nahrungspflanzen zu reagieren. Verstärkt wurde der Aussterbeprozess durch die geringe Populationsdichte und die bei Nashörnern übliche lange Generationsfolge.[13]
Die Entwicklung des Hornes von Elasmotherium
Elasmotherium stellt das Endglied der Entwicklung der Elasmotherien dar und ist durch ein einzelnes, auf der Stirn aufragendes Horns charakterisiert, das zudem auf einer großen domartigen Erhebung steht. Lange Zeit war unklar, wie dieses Horn und dessen knöcherner Unterbau evolutiv entstanden war. Die meisten stammesgeschichtlichen Vorgänger von Elasmotherium besaßen ein einzelnes Horn auf der Nase und eine flache oder gerade verlaufende Stirnlinie. Einzig bei Sinotherium war aufgrund des nur fragmentiert überlieferten Schädels die Ausbildung des Hornes bisher nicht bekannt gewesen, anhand anatomischer Merkmale des Schädels gehörte der Nashornvertreter aber zu den nächsten Verwandten von Elasmotherium. Durch den Fund eines Teilschädels aus dem Linxia-Becken im Jahr 2012 ist nun von allen moderneren Elasmotherien die Schädelmorphologie weitgehend bekannt, dadurch lässt sich die Hornentwicklung innerhalb dieser Nashorngruppe besser nachvollziehen. Sie begann bei vergleichsweise kleinen Vertretern mit einem ebenfalls kleinen Horn auf der Nasenspitze, wie dies bei Hispanotherium aus dem Übergang vom Unteren zum Mittleren Miozän vor 18 Millionen Jahren zu erkennen ist. Anhand der Fossilien lässt sich die langsame Zunahme der Körpergröße der Tiere, aber auch der Hörner durch die Ausdehnung der aufgerauten Ansatzstellen nachvollziehen. Im Laufe der stammesgeschichtlichen Entwicklung der Elasmotherien kam es auch zu einer leichten Rückverlagerung des vorderen Hornes, dass nun nicht mehr direkt auf der Nasenspitze, sondern etwas weiter hinten auf dem Nasenbein saß: Bekannt ist dies vor allem bei der spätmiozänen Form Parelasmotherium, die vor 10 bis rund 5 Millionen Jahren lebte. Bei Sinotherium befand sich die Hornbasis am Übergang vom Nasen- zum Stirnbein; eine zusätzlich weitere, deutlich kleinere Hornbasis lag direkt dahinter und war leicht abgetrennt. Beide zeigten aber schon einen deutlich kuppelartigen Aufbau, wie er auch von Elasmotherium aus dem Plio- und dem Pleistozän bekannt ist. Bei diesem erhob sich die Hornbasis aber direkt auf der Stirn. Bei der Entwicklung zu Elasmotherium muss es somit zu einer Fusion dieser beiden kuppelartigen Erhebungen gekommen sein. Auf der Hornbasis von Elasmotherium ist eine leichte Knochennaht erkennbar, deren Ursprung bisher nicht erklärbar war, jedoch ihren Ursprung wohl in der Verschmelzung der beiden bei Sinotherium getrennten Erhebungen haben könnte.[51]
Parallel zur Rückwärtsverschiebung des Hornes kam es bei den Elasmotherien auch zu einer Überformung des Nasenbereiches. Elasmotherium besitzt eine vollständig verknöcherte Nasenscheidewand, ebenso, wie Sinotherium; bei älteren Elasmotherien ist dies weitgehend nicht bekannt. Lediglich Ningxiatherium aus dem späten Miozän weist ein im vorderen Drittel verknöchertes Septum auf. Ob diese Verwachsungen mit der Rückverlagerung des Hornes zusammenhängen, ist unbekannt. Zur Stabilisierung der Stirn und des hinteren Nasenbereiches durch die Auflast des massiven Horns besaß Sinotherium zahlreiche querverlaufende knöcherne Rippen am Gaumenbereich. Verknöcherte Nasenscheidewände treten bei Nashörnern ansonsten wiederum nur in der Coelodonta-Stephanorhinus-Linie auf, die besonders deutlich beim Wollnashorn und beim Steppennashorn ausgebildet sind.[51]
Forschungsgeschichte

Als Fischer von Waldheim 1808 die Gattung Elasmotherium einführte, stand ihm nur ein Unterkiefer zur Verfügung. Er brachte die neue Form daher noch nicht mit einem ausgestorbenen Nashorn in Verbindung, sondern verglich sie mit anderen, heute nicht mehr existenten Säugetierarten, die nach seiner Meinung unter den rezenten Vertretern keine Verwandten besäßen. Dazu gehörten unter anderem das Amerikanische Mastodon (ein urtümliches Rüsseltier), Megatherium und Megalonyx (zwei bodenlebende Faultiere) oder Palaeotherium (ein Pferdeverwandter).[42] In der nachfolgenden Zeit blieb die Beziehung von Elasmotherium ohne zusätzliche Funde weiter unklar. Richard Owen führte die Gattung daher in seinem bedeutenden Werk Odontography, das von 1840 bis 1845 herausgebracht wurde, lediglich innerhalb der Huftiere.[52] Allerdings vermutete Johann Jakob Kaup bereits 1840 unter Einbeziehung des Schädelfragments aus der Sammlung von Franz Joseph Gall, dass es sich bei Elasmotherium um einen Vertreter der Nashörner handelt.[47] (Das gleiche Schädelfragment benutzte Georges Louis Duvernoy zur Aufstellung von Stereocerus galli, das er ebenfalls den Nashörnern zuwies,[53] die Form war aber nicht allgemein anerkannt.) Dem schloss sich Johann Friedrich von Brandt fast ein Viertel Jahrhundert später mit einer ähnlichen Argumentation an,[45] doch erst der 1877 bei Lutschka aufgefundene Schädel ermöglichte es Brandt, Elasmotherium eindeutig zu den Nashörnern zu verweisen. Er legte 1878 eine umfassende Beschreibung des Schädels vor, in der er nicht nur die Stirnkuppel als besondere Bildung hervorhob, sondern auch Mutmaßungen über Aussehen und Größe des Tieres äußerte.[54] Im gleichen Jahr veröffentlichte der russische Wissenschaftler Alexander Fjodorowitsch Brandt, ein Verwandter Johann Friedrichs, eine erste bildliche Rekonstruktion anhand der bisherigen Funde von Elasmotherium, die vor allem ein massives Stirnhorn beinhaltete.[3]
In der Folgezeit wurde Elasmotherium in Ermangelung an umfassenden Skelettmaterial häufig in einer äußeren Gestalt ähnlich dem Wollnashorn allerdings mit nur einem Stirnhorn interpretiert. Konstantin K. Fljorow rekonstruierte die Tiere in den 1950er Jahren als Steppenbewohner, die auf der Suche nach Nahrung den Boden mit ihrer Schnauze aufwühlten. Der Kopf wurde von einem Horn dominiert, das kappenartig auf der domartigen Schädelaufwölbung aufsaß. Den Stirndom interpretierte Fljorow zudem als erweitertes Geruchsorgan, dass bei der Suche nach Nahrung behilflich war. Etwa im gleichen Zeitraum sah V. A. Teryaev Elasmotherium als flusspferdartiges Tier, das Sümpfe bewohnte. In seiner Lebendrekonstruktion ähnelte es nicht mehr einem Nashorn, die kuppelartige Aufwölbung auf der Stirn war hornlos, zusätzlich bestanden die Vorderfüße aus vier Zehen, vergleichbar den Tapiren, um besser im sumpfigen Untergrund voranzukommen. Teryaev begründete seine Interpretation mit dem häufigen Auffinden von Fossilfunden von Elasmotherium in ehemaligen Flusslandschaften. Er sah auch die Aufwölbung als zu grazil an, um eine tragende Funktion auszuüben.[14][15] In jüngerer Zeit konnten Teryaevs Ansichten aus anatomischer Sicht meist revidiert werden.[33][3]
Elasmotherium und der Mensch
Ob das riesige Nashorn Beutetier früher menschlicher Jäger- und Sammlergruppen war, wie etwa des Homo erectus oder des Neandertalers, ist unklar, an archäologischen Fundstellen scheint es jedoch bisher nur selten vorzukommen. Die klassische Fundstelle Sinaya Balka auf der Taman-Halbinsel erbrachte neben dem Typusexemplar von Elasmotherium caucasicum und weiteren Individuen auch zusätzlich Steinartefakte des Altpaläolithikums.[12] Von einigen Travertinfundstellen im Süden von Kasachstan, etwa Koshkurgan und Shoktas, liegen ebenfalls altpläolithische Steinartefakte gemeinsam mit Resten von Elasmotherium sibiricum vor. Die Funde sind etwas jünger als jene von Sinaya Balka und datieren in das frühe Mittelpleistozän.[35]

Eher problematisch ist die Interpretation einiger charakteristischer Nashornporträts der jungpaläolithischen Höhlenkunst wie etwa aus der Höhle Rouffignac als Darstellung von Elasmotherium.[55] In der Regel beziehen sich diese Annahmen auf individuelle Nashornzeichnungen mit nur einem Horn. Unter der Voraussetzung, dass die Kunstdarstellungen des Jungpaläolithikums höchstwahrscheinlich nicht der getreuen Wiedergabe realistischer Objekte dienten, können alle Nashorndarstellungen dem Wollnashorn zugewiesen werden. Zudem fehlen bisher eindeutige Belege, dass Elasmotherium sowohl räumlich als auch zeitlich so weit verbreitet war.[17][3]
Teilweise wird Elasmotherium auch mit den Legenden um das vermeintliche Einhorn in Verbindung gebracht. Ein Wegbereiter dafür war Alexander Fjodorowitsch Brandt, der in seinem populären Artikel aus dem Jahr 1878 über Erzählungen einiger ostsibirischer Völker über die Erlegung riesiger schwarzer, einhörniger Bullen berichtete. Die ersten schriftlichen Zeugnisse über das Fabeltier finden sich in der griechischen Antike. Bereits zuvor wurden einhornartige Wesen auf Siegeln in den Städten Mohenjo-Daro und Harappa der Indus-Kultur abgebildet, die etwas mehr als 4000 Jahre alt sind. Unklar ist allerdings, ob sie tatsächlich Einhörner darstellen oder dies nur durch ihre strikte Seitendarstellung suggerieren. Elasmotherium als Prototyp des Einhorns kann aber nur dann eine Berechtigung haben, wenn eine lückenlose mündliche Überlieferung aus der Zeit seiner Existenz bis in jüngere zeitliche Abschnitte bestanden hätte.[3]
Literatur
- Анна К. Швырева: Исқопаемые носороги зласмотерии. Stawropol, 1995, S. 1–104.
- Anna K. Schvyreva: On the importance of the representatives of the genus Elasmotherium (Rhinocerotidae, Mammalia) in the biochronology of the Pleistocene of Eastern Europe. Quaternary International 379, 2015, S. 128–134.
- Vadim V. Titov, Vera S. Baigusheva und Roman S. Uchytel’: The experience in reconstructing of the head of Elasmotherium (Rhinocerotidae). Russian Journal of Theriology 20 (2), 2021, S. 173–182, doi:10.15298/rusjtheriol.20.2.06
- Vladimir Zhegallo, Nikolay Kalandatze, Andrey Shapovalov, Zoya Bessudnova, Natalia Noskova und Ekaterina Tesakova: On the fossil rhinoceros Elasmotherium (Including the collections of the Russian Academy of sciences). Cranium 22 (1), 2005, S. 17–40.
Einzelnachweise
- Deng Tao: A new elasmothere (Perissodactyla, Rhinocerotidae) from the late Miocene of the Linxia Basin in Gansu, China. Geobios 41, 2008, S. 719–728
- Deng Tao und Zheng Ming: Limb bones of Elasmotherium (Rhinocerotidae, Perissodactyla) from Nihewan (Hebei, China). Vertebrata Palasiatica 43, 2005, S. 110–121
- Vladimir Zhegallo, Nikolay Kalandatze, Andrey Shapovalov, Zoya Bessudnova, Natalia Noskova und Ekaterina Tesakova: On the fossil rhinoceros Elasmotherium (Including the collections of the Russian Academy of sciences). Cranium 22 (1), 2005, S. 17–40
- E. I. Belyaeva: About the hyoideum, sternum and metacarpale V bones of Elasmotherium sibiricum Fischer (Rhinocerotidae). Journal of the Palaeontological Society of India 20, 1977, S. 10–15
- Paul Mazza und A. Azzaroli: Ethological inferences on Pleistocene rhinoceroses of Europe. Rendiconti Lincei 4, (2) 1993, S. 127–137
- Jan van der Made und René Grube: The rhinoceroses from Neumark-Nord and their nutrition. In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale, 2010, S. 382–394
- Ekaterina Petrova: New data about the skull of the Elasmotherium sibiricum. In: Abstraktband: Quaternary stratigraphy and paleontology of the Southern Russia: connections between Europe, Africa and Asia. 2010 annual meeting INQUA-SEQS, Rostov-on-Don, Russia, June 21–26, 2010. Rostow am Don, 2010, S. 121–122
- Esperanza Cerdeño: Cladistic analysis of the family Rhinocerotidae (Perissodactyla). American Museum Novitates 3143, 1995, S. 1–25
- Vadim V. Titov, Vera S. Baigusheva und Roman S. Uchytel’: The experience in reconstructing of the head of Elasmotherium (Rhinocerotidae). Russian Journal of Theriology 20 (2), 2021, S. 173–182, doi:10.15298/rusjtheriol.20.2.06
- Mikael Fortelius: Ecological aspects of dental functional morphology in the Plio-Pleistocene rhinoceroses of Europe. In: B. Kurtén (Hrsg.): Teeth: Form, function and evolution. New York, 1982, S. 163–181
- Donald R. Prothero, Claude Guérin und Earl Manning: The history of Rhinocerotoidea. In Donald R. Prothero und R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New-York, 1989, S. 321–340
- V. S. Baigusheva, G. I. Timonina und V. V. Titov: Some biological features of Elasmotherium caucasicum (Mammalia, Rhinocerotidae) from the Late Early Pleistocene of Eastern Europe. In: Femke Holwerda, Anneke Madern, Dennis Voeten, Anneke van Heteren, Hanneke Meijer und Natasja den Ouden (Hrsg.): XIV Annual Meeting of the European Association of Vertebrate Palaeontologists 6-10 July 2016, Haarlem, The Netherlands Programme and Abstract Book. Haarlem, 2016, S. 43
- Pavel Kosintsev, Kieren J. Mitchell, Thibaut Devièse, Johannes van der Plicht, Margot Kuitems, Ekaterina Petrova, Alexei Tikhonov, Thomas Higham, Daniel Comeskey, Chris Turney, Alan Cooper, Thijs van Kolfschoten, Anthony J. Stuart und Adrian M. Lister: Evolution and extinction of the giant rhinoceros Elasmotherium sibiricum sheds light on late Quaternary megafaunal extinctions. Nature Ecology & Evolution 3, 2019, S. 31–38, doi:10.1038/s41559-018-0722-0
- В. А. Теряев: Был ли Elasmotherium трехпалым? Ежегодник 8, 1930, S. 77–82
- В. А. Теряев: Геологическое положение горболобого носорога (зласмотерия). Советская Геология 34, 1948, S. 81–89
- N. G. Noskova: Elasmotherians – evolution, distribution and ecology. In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche (Rom) 2001, S. 126–128
- Baldyrgan S. Kožamkulova: Elasmotherium sibiricum und sein Verbreitungsgebiet auf dem Territorium der UdSSR. Quartärpaläontologie 4, 1981, S. 85–91
- Oscar Sanisidro, María Teresa Alberdi und Jorge Morales: The First Complete Skull of Hispanotherium matritense (Prado, 1864) (Perissodactyla, Rhinocerotidae) from the Middle Miocene of the Iberian Peninsula. Journal of Vertebrate Paleontology, 32 (2), 2012, S. 446–455
- Shanlin Liu, Michael V. Westbury, Nicolas Dussex, Kieren J. Mitchell, Mikkel-Holger S. Sinding, Peter D. Heintzman, David A. Duchêne, Joshua D. Kapp, Johanna von Seth, Holly Heiniger, Fátima Sánchez-Barreiro, Ashot Margaryan, Remi André-Olsen, Binia De Cahsan, Guanliang Meng, Chentao Yang, Lei Chen, Tom van der Valk, Yoshan Moodley, Kees Rookmaaker, Michael W. Bruford, Oliver Ryder, Cynthia Steiner, Linda G.R. Bruins-van Sonsbeek, Sergey Vartanyan, Chunxue Guo, Alan Cooper, Pavel Kosintsev, Irina Kirillova, Adrian M. Lister, Tomas Marques-Bonet, Shyam Gopalakrishnan, Robert R. Dunn, Eline D. Lorenzen, Beth Shapiro, Guojie Zhang, Pierre-Olivier Antoine, Love Dalén und M. Thomas P. Gilbert: Ancient and modern genomes unravel the evolutionary history of the rhinoceros family. Cell 184 (19), 2021, S. 4874–4885, doi:10.1016/j.cell.2021.07.032
- Ashot Margaryan, Mikkel-Holger S. Sinding, Shanlin Liu, Filipe Garrett Vieira, Yvonne L. Chan, Senthilvel K. S. S. Nathan, Yoshan Moodley, Michael W. Bruford und M. Thomas P. Gilbert: Recent mitochondrial lineage extinction in the critically endangered Javan rhinoceros. Zoological Journal of the Linnean Society 190 (1), 2020, S. 372–383, doi:10.1093/zoolinnean/zlaa004
- Miklós Kretzoi: Gobitherium n. g. (Mamm., Rhinoc.). Földtany Közlony 73, 1943, S. 268–271 ()
- Anna K. Schvyreva: On the importance of the representatives of the genus Elasmotherium (Rhinocerotidae, Mammalia) in the biochronology of the Pleistocene of Eastern Europe. Quaternary International 379, 2015, S. 128–134
- Danhui Sun, Tao Deng und Qigao Jiangzuo: The most primitive Elasmotherium (Perissodactyla, Rhinocerotidae) from the Late Miocene of northern China. Historical Biology: An International Journal of Paleobiology, 2021, doi:10.1080/08912963.2021.1907368
- В. В. Титов: Крупные млекопитающие позднего плиоцена Зеверо-Восточного Приазовья. Rostow am Don, 2008
- Vera Baigusheva und Vadim Titov: Pleistocene large mammals associations of the Sea of Azov and adjacents regions. In: Abstraktband: Quaternary stratigraphy and paleontology of the Southern Russia: connections between Europe, Africa and Asia. 2010 annual meeting INQUA-SEQS, Rostov-on-Don, Russia, June 21–26, 2010. Rostow am Don, 2010, S. 24–27
- Vera S. Bajgusheva und Vadim V. Titov: Results of the Khapry Faunal Unit revision. In: L. Maul und R. D. Kahlke (Hrsg.): Late Neogene and Quaternary biodiversity and evolution: Regional developments and interregional correlations. Conference Volume. 18th International Senckenberg Conference. VI International Palaeontological Colloquium in Weimar. Konferenz in Weimar, 25.–30. April, 2004. Terra Nostra, Schriften der Alfred-Wegener-Stiftung (Berlin), 2004
- Vitaliy Logvynenko: The development of Late Pliocene to early Middle Pleistocene large mammal fauna of Ukraine. In: L. Maul und R. D. Kahlke (Hrsg.): Late Neogene and Quaternary biodiversity and evolution: Regional developments and interregional correlations. Conference Volume. 18th International Senckenberg Conference. VI International Palaeontological Colloquium in Weimar. Konferenz in Weimar, 25.–30. April, 2004. Terra Nostra, Schriften der Alfred-Wegener-Stiftung (Berlin), 2004
- Esperanza Cerdeño: Diversity and evolutionary trends of the the family Rhinocerotidae (Perissodactyla). Palaeo 141, 1998, S. 13–34
- Piruza Tleuberdina, Gulzhan Nazymbetova: Distribution of Elasmotherium in Kazakhstan. In: Abstraktband: Quaternary stratigraphy and paleontology of the Southern Russia: connections between Europe, Africa and Asia. 2010 annual meeting INQUA-SEQS, Rostov-on-Don, Russia, June 21–26, 2010. Rostow am Don, 2010, S. 171–173
- А. Борисякъ: О зубномъ аппарате Elasmotherium caucasicum n. sp.. Исвестя Императорскоӣ Академиӣ Наук 6 (9), 1914, S. 555–584
- Chow Minchen: New elasmotherine rhinoceroses from Shansi. Vertebrata Palasiatica 2 (2/3), 1958, S. 132–142
- Tong Haowen, Wang Fagang, Zheng Min und Chen Xi: New Fossils of Stephanorhinus kirchbergensis and Elasmotherium peii from the Nihewan Basin. Vertebrata Palasiatica 33 (3), 2014, S. 369–388
- Анна К. Швырева: Исқопаемые носороги зласмотерии. Stawropol, 1995, S. 1–104
- Florent Rivals, Natalya E. Prilepskaya, Ruslan I. Belyaev und Evgeny M. Pervushov: Dramatic change in the diet of a late Pleistocene Elasmotherium population during its last days of life: Implications for its catastrophic mortality in the Saratov region of Russia. Palaeogeography, Palaeoclimatology, Palaeoecology 556, 2020, S. 109898, doi:10.1016/j.palaeo.2020.109898
- Anatoly P. Derevianko, Valery T. Petrin, Zhaken. K. Taimagambetov und Marcel Otte: Early Palaeolithic assemblages in travertine, Southern Kazakhstan (a variant of an adaptation model). Anthropologie 36, 1998, S. 137–164
- Diana Pushkina: The Pleistocene easternmost distribution in Eurasia of the species associated with the Eemian Palaeoloxodon antiquus assemblage. Mammal Review 37 (3), 2007, S. 224–245
- А. А. Хромов: Зуб эласмотерия с нетипичной из среднеплейстоценовых отложений Саратовского поволжья. In: В. М. Подбина, С. А. Родыгин, Н. И. Савина und Г. М. Татьянин: Эволюция жизни на Земле. Материалы II Международного симпозиума 12-15 ноября 2001 г. (Evolution of life on the earth. Proceedings of the II International symposium 12. - 15. November 2001) Tomsk, 2001, S. 534–535
- Pavel Kosintsev: Relict mammal species of the Middle Pleistocene in Late Pleistocene Fauna of Western Siberia south. In: Abstraktband: Quaternary stratigraphy and paleontology of the Southern Russia: connections between Europe, Africa and Asia. 2010 annual meeting INQUA-SEQS, Rostov-on-Don, Russia, June 21–26, 2010. Rostow am Don, 2010, S. 78–79
- P. A. Kosintsev: Elasmotherium (Elasmotherium sibiricum Fischer, 1808): new data on the period of existence and geographic range. In: A. V. Borodin, E. A. Markova und T. V. Strukova (Hrsg.): The Quaternary of the Urals: global trends and Pan-European Quaternary records. International conference INQUA-SEQS 2014 Ekaterinburg, Russia, September 10–16, 2014. Ekaterinenburg, 2014, S. 67–68
- Andrei Valerievich Shpansky, Valentina Nurmagambetovna Aliyassova und Svetlana Anatolievna Ilyina: The Quaternary Mammals from Kozhamzhar Locality (Pavlodar Region, Kazakhstan). American Journal of Applied Sciences 13 (2), 2016, S. 189–199 ()
- Paula Jo Reimer und Svetlana Vladimirovna Svyatko: Comment on Shpansky et al. 2016, ‘The Quaternary Mammals from Kozhamzhar Locality (Pavlodar Region, Kazakhstan). American Journal of Applied Sciences 13 (4), 2016, S. 477–478
- Gotthelf Fischer von Waldheim: Sur l'Elasmotherium et le Trogontherium deux animaux fossiles et insonnus de la Russie. Mémoires de la Société des naturalistes de Moscou 2, 1809, S. 251–268 ()
- Gotthelf Fischer von Waldheim: Nachtrag zur Geschichte des Elasmotheriums. Bulletin de la Société impériale des naturalistes de Moscou 15, 1842, S. 457–461 ()
- Anselme Gaëtan Desmarest: Mammalogie ou description des espèces des Mammifères. Paris, 1820, S. 1–555 (S. 546) ()
- Johann Friedrich von Brandt: Observationes de elasmotherii reiquiis. Mémoires de l'Académie impériale des sciences de St.-Pétersbourg 7, 8 (4), 1864, S. 1–34 ()
- Alexander Graf Keyserling: Beobachtung eines Elasmotherium. Bulletin de la Société impériale des naturalistes de Moscou 15, 1842, S. 454–457 ()
- Johann Jakob Kaup: Über das Elasmotherium. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrafakten-Kunde, 1840, S. 453–456 ()
- Mikael Fortelius und Kurt Heissig: The phylogenetic relationships of the Elasmotheriini (Rhinocerotidae, Mamm.). Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 29, 1989, S. 227–233
- Kurt Heissig und Oldřich Fejfar: Die fossilen Nashörner (Mammalia, Rhinocerotidae) aus dem Untermiozän von Tuchorice in Nordwestböhmen. Sborník Národního Muzea v Praze. Acta Musei Nationalis Pragae (series B, Natural History) 63 (1), 2007, S. 19–64
- Pierre-Oliver Antoine: Middle Miocene elasmotheriine Rhinocerotidae from China and Mongolia: taxonomic revision and phylogenetic relationships. The Norwegian Academy of Science and Letters - Zoologica Scripta 32, 2003, S. 95–118
- Tao Deng, ShiQi Wang und SuKuan Hou: A bizarre tandem-horned elasmothere rhino from the Late Miocene of northwestern China and origin of the true elasmothere. Chinese Science Bulletin 2012, S. 1–7
- Richard Owen: Odontography; or, A treatise on the comparative anatomy of the teeth; their physiological relations, mode of development, and microscopic structure, in the vertebrate animals. Hippolyte Bailliere, London, 1840–1845, S. 1–655 (S. 587) ()
- Georges Louis Duvernoy: Nouvelles études sur les rhinocéros fossiles. Archives du Muséum d'Histoire Naturelle 7, 1855, S. 1–482 (S. 125–130) ()
- Johann Friedrich von Brandt: Zur Geschichte der allmälig gewonnenen Kenntniss des Elasmotheriums. Mémoires de l'Académie impériale des sciences de St.-Pétersbourg 7, 26 (6), 1878, S. 1–31 ()
- W. T. Schaurte: Darstellung eines Elasmotherium in der Felsmalerei von Rouffignac. Natur und Museum 94 (9), 1964, S. 354–356