Na Dương (Fossillagerstätte)
Na Dương ist eine bedeutende Fossillagerstätte im nordöstlichen Vietnam. Sie liegt im gleichnamigen Becken nahe der Grenze zu China. Dieses ist durch seinen Reichtum an Steinkohle bekannt, die in einem Tagebau gefördert wird. Das Becken verdankt seine Entstehung einer ausgedehnten Verwerfungszone, die durch die Kollision des Indischen Subkontinentes mit der asiatischen Landmasse verursacht wurde. Die Aufschlüsse im Tagebau von Na Dương zeigen eine Abfolge aus Kohle, Tonsteinen und Schluffsteinen. Sie lassen sich zwei Gesteinseinheiten zuweisen, die beide fossilführend sind. Die Fossilien umfassen Reste der Pflanzen- und Tierwelt. Letztere sind mit Wirbellosen, Fischen, Reptilien und Säugetieren vertreten. Der Großteil der Landwirbeltiere entstammt einer basalen Kohlelage.
Die Gesteinsablagerungen entstanden unter den Bedingungen eines stehenden Gewässers. Dieses wurde von Weichtieren, Fischen, Schildkröten und Krokodilen besiedelt. Im Umland wuchs ein dichter Wald, worauf verschiedene erhaltene Baumstümpfe hinweisen, ebenso wie Blattreste. Möglicherweise stand der Wald mit einer Sumpflandschaft in Verbindung. Hier lebten diverse Huftiere. Der Lebensraum war wahrscheinlich einem tropischen Klima ausgesetzt. Absolute Altersangaben sind für die Ablagerungen von Na Dương nicht verfügbar. Die Zusammensetzung der Flora und Fauna spricht aber für eine Einstufung in das Obere Eozän bis Untere Oligozän, was einem Zeitraum von vor 38 bis 28 Millionen Jahren entspricht.
Fossilfunde sind aus Na Dương bereits seit dem Ende der 1970er Jahre bekannt. Intensivere und zumeist international orientierte Arbeiten erfolgten aber weitgehend erst mit dem Beginn des 21. Jahrhunderts. Dazu gehören auch Vorortuntersuchungen der Senckenberg Gesellschaft für Naturforschung, in deren Zuge das bisher umfassendste Material geborgen wurde.
Geographische Lage
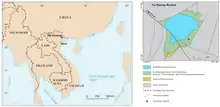
Die Fossillagerstätte Na Dương liegt nahe dem gleichnamigen Hauptort des Bezirks Lộc Bình im Nordosten von Vietnam. Der Bezirk wiederum gehört zur Provinz Lạng Sơn, deren Hauptstadt befindet sich rund 20 km nordwestlich von Na Dương. Geographisch bildet Na Dương einen Teil des Na-Dương-Beckens, das rund 45 km² groß ist und eine leicht längliche Form mit einer von Nordnordost nach Südsüdwest orientierten Achse besitzt. Es wird vom Fluss Kỳ Cùng entwässert. Das Na-Dương-Becken zeichnet sich vor allem durch seinen Reichtum an Steinkohle aus. Der Rohstoff wird im Tagebaubetrieb unmittelbar nördlich des Hauptortes erschlossen und findet in dem angegliederten Kraftwerk Na Dương Verwendung. Der Tagebauaufschluss markiert den Südostrand des Beckens.[1][2][3]
Geologie
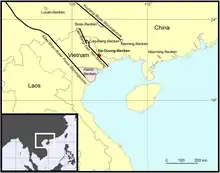
Aus geologischer Sicht liegt das Na-Dương-Becken nahe der Grenze zwischen der Indochina- und der Südchinesischen Mikroplatte (Terrane). Charakterisiert wird die Region durch eine große, von Nordwest nach Südost verlaufende Verwerfungszone, die allgemein unter der Bezeichnung Ailao Shan-Red River fault zone bekannt ist. Diese erstreckt sich über eine Länge von 1000 km von Tibet kommend bis zum Golf von Tonkin. Seine Entstehung verdankt der Verwerfungsgürtel der Kollision des Indischen Subkontinentes mit Asien im Verlauf des Eozäns vor rund 50 bis 40 Millionen Jahren, teilweise wird aber auch ein deutlich jüngerer, im Miozän ansetzender Ursprung diskutiert.[4] Unmittelbar verbunden mit der Ailao Shan-Red River fault zone sind einige kleinere, parallel orientierte Bruchzonen, von denen sich das Cao Bang-Tien Yen fault system in rund 160 km nördlicher Entfernung befindet und sich über eine Distanz von rund 230 km verfolgen lässt. Entlang letzteren Verwerfungssystems haben sich im weiteren Verlauf mehrere Beckenstrukturen herausgeformt. Diese beginnen im Nordwesten mit dem Cao-Bằng-Becken und setzen sich nach Südosten mit dem Thất-Khê-Becken, dem Lạng-Sơn-Becken und dem Na-Dương-Becken fort. Die Bildung der Becken geht auf Dehnungsprozesse in den Verwerfungszonen zurück, die durch tektonische Spannungen hervorgerufen wurden. Sie können daher als pull-apart basins („Ausziehbecken“) angesehen werden.[5][6][2][3][7]
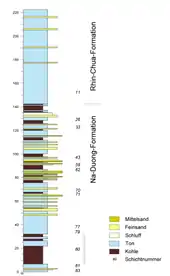
Die Basis des Beckens bilden triassische und kreidezeitliche Ablagerungen terrigenen Ursprungs, die sich weitgehend aus Siliziklastika zusammensetzen. Ihre Mächtigkeit beträgt rund 500 m. Darauf folgen Sedimente mit känozoischer Zeitstellung, deren Dicke mit rund 570 m nahezu der des älteren Untergrundes entspricht. Es lassen sich zwei Ablagerungseinheiten differenzieren. Zuunterst befindet sich die Na-Duong-Formation, überdeckt wird sie von der Rhin-Chua-Formation. Die Na-Duong-Formation wird Bohrkernerkundungen zufolge im Beckenzentrum bis zu 240 m mächtig. Davon sind im Tagebau des Na-Dương-Beckens die oberen 140 m aufgeschlossen. Die randliche Lage des Tagebaus im Süden des Beckens bedingt, dass die einzelnen Schichten der Gesteinseinheit teilweise in bis zu einem Winkel von 29° nach Norden einfallen. Der aufgeschlossene Bereich besteht aus einer Wechsellagerung von Steinkohle und bräunlichen bis gräulichen, mergeligen Ton-, Schluff- und feinkörnigen Sandsteinen, gelegentlich kommen auch gröberkörnige Sandsteine vor. Vor allem die Sandsteine bilden mitunter massive Bänke. Im stärker ausgewitterten Zustand verlieren die Sedimente ihren Karbonatgehalt, ändern aber nicht ihre Färbung. Die Steinkohle wiederum tritt als teils bitumenartiges, teils schieferartiges Substrat auf. Der mächtigste Flöz, der in der lokalen Gliederung die Schichtnummer 80 erhielt, befindet sich basal in der aufgeschlossenen Folge und zeigt sich als fein geschichtet mit teils hohem Anteil an Eisensulfiden, überwiegend Pyrit. Nach oben hin wird das Flöz durch gelbliche schluffige bis sandige Tonsteine unterbrochen. Die Kohle enthält rund 18,5 % an Feuchtigkeit.[1] Ihre Hauptbestandteile3 sind Mazerale der Vitrinitgruppe, allen voran Telinit und Collotelinit.[6][2][3][7]
Die sich im Hangenden anschließende Rhin-Chua-Formation wird bis zu 300 m mächtig und ist im Aufschlussgebiet des Tagebaus in ihren unteren 80 m freigelegt. Sie enthält im Gegensatz zur Na-Duong-Formation keine Kohle und besteht überwiegend aus bräunlichen mergeligen Tonsteinen, zwischen die sich im oberen Aufschlussbereich dünne Lagen aus feinkörnigen Sandsteinen mit symmetrischen Rippelmarken schieben. Der Übergang zwischen beiden Gesteinseinheiten wird je nach Autor als konkordant[3] oder als diskordant[6] beschrieben. Die Tonsteine erhalten im verwitterten Zustand durch sekundäre Hämatitbildung einen rostfarbenen Farbcharakter.[6][2][3][7]
Fossilfundstellen
Das Fossilmaterial der Na-Duong-Formation umfasst sowohl pflanzliche wie auch tierische Reste. Pflanzen sind wiederum als Pollen, aber auch als Makroreste in Form von Blättern, Samen oder Baumstümpfen überliefert, ebenso kommen Harze vor. Die Fauna setzt sich aus Wirbellosen und Wirbeltieren zusammen. Erstere schließen überwiegend Weichtiere ein, letztere Fische, Reptilien und Säugetiere.[2][3]
Die Funde streuen durch das gesamte Profil. Es lassen sich aber einige größere Fundbereiche eingrenzen. Die untere mächtige Kohleschicht mit der Schichtnummer 80 bildet das primäre Fossillager für Wirbeltiere. Dort kommen aber auch pflanzliche Reste vor. Der größere Teil der Fossilien entstammt direkt dem Basisabschnitt am Übergang von der unterlagernden Tonsteinschicht zur schieferigen Kohle. Die im oberen Teil des Kohleflözes auftretenden schmalen Tonsteinbänder enthalten wiederum versteinerte Baumstämme in höherer Anzahl zuzüglich einiger reichhaltiger Blätterakkumulationen. Weiter im Hangenden treten lokal zusätzliche Fundkonzentrationen auf, die zumeist Baumstämme wie in Schicht 43 und 26 beinhalten. In den mergeligen Tonsteinen sind zudem Blätter auffindbar, deren Erhaltungszustand mitunter so gut ist, dass ihre Cuticula überliefert ist.[2][3]
Auch in der Rhin-Chua-Formation sind Fossilien in Form von Mollusken und Fischen dokumentiert, darüber hinaus kommen Pflanzen vor. Sie streuen hier ebenfalls durch größere Abschnitte des Profils, sind aber teilweise an Bioturbationen gebunden.[2][3]
Funde
Flora
Die Flora besteht sowohl aus Mikro- wie auch aus Makroresten. Zu den Mikroresten gehören die Pollen, die sich in nahezu allen Abschnitten der Na-Duong- und der Rhin-Chua-Formation finden. Über diese ließen sich unter anderem Nacktsamer wie Tannen, Hemlocktannen, Stechtannen, Kiefern oder Taubenbäume nachweisen. Bedecktsamer sind mit Hickory, Gänsefüßen, Knöterichen, Walnüssen, Stechpalmen, Eichen, Weiden, Ulmen und Zelkoven, aber auch mit Lithocarpus, Diplopanax und Myrica vertreten. Zu den Gefäßsporenpflanzen wiederum zählen Farne wie Hypolepis, Pteris und Pyrrosia, zusätzlich auch die Bärlappe.[7] Makroreste liegen in Form von Blättern, Samen und Stämmen vor. Die basale Kohleschicht 80 enthält zahlreiche kalzifizierte Baumstämme, die einen Durchmesser von bis zu einem Meter haben können, bisher aber weitgehend unidentifiziert sind. Ebenso kommen Stammreste von Königsfarngewächsen vor. In den unterlagernden Schichten 81 und 83, aber auch in hangenden Schichten treten Samen von Lotosblumen auf. Möglicherweise können diesen auch zahlreiche Rhizome aus den gleichen Fundschichten zugeordnet werden. Vor allem in den sandigen Tonsteinlagen im oberen Abschnitt der basalen Kohle sind häufig Blattreste erhalten. Auffallenderweise zeigen diese zu einem größeren Teil gezackte oder gezähnte Ränder. Einige ähneln Eotrigonobalanus aus der Gruppe der Buchengewächse. Andere wiederum entsprechen den Bauhinien aus der Gruppe der Hülsenfrüchtler. Daneben sind Farne vertreten, wie etwa Pronephrium. Aus zahlreichen Lagen konnten honigfarbene Harzreste von bis zu 2 cm Länge geborgen werden, die spektroskopisch in etwa mit Shorea aus der Gruppe der Flügelfruchtgewächse übereinstimmen.[3]
Wirbellose
Die Wirbellosen beschränken sich weitgehend auf Weichtiere. In der Na-Duong-Formation kommen sie weitgehend nur in den mergeligen Tonsteinen vor, in den Kohleschichten haben sich die Gehäuse dort möglicherweise zersetzt. Ein größerer Teil der Funde wird durch die Fluss- und Teichmuscheln gestellt. Deren Gehäusehälften sind in der Regel noch artikuliert oder liegen geöffnet in Schmetterlingsposition. An Gattungen konnten Nodularia und Cuneopsis aus der Gruppe der Unionini sowie Cristaria aus der Gruppe der Anodontini bestimmt werden. Alle drei sind noch heute in Flüssen und Seen Ost- und Südostasiens präsent. Des Weiteren kommen zahlreiche Schneckenschalen vor, die wahrscheinlich zu Sumpfdeckelschnecken gehören, aber zu stark fragmentiert sind. Als Abdrücke erhielten sich einzelne Gehäuse von Tarebia, eine Angehörige der Kronenschnecken in der Rhin-Chua-Formation. Von dort und auch aus der Na-Duong-Formation wurde zudem die Gattung Brotia aus der Gruppe der Pachychilidae berichtet.[2][8][3][9]
Fische
Fische sind häufig, aber zumeist nur über isolierte Knochen oder Schlundzähne überliefert. Es lassen sich Reste von zwei größere Gruppen unterscheiden, die Amiidae und die Karpfenartigen. Das Fundmaterial der Amiidae, zu denen der heutige Kahlhecht gehört, ist zu stark fragmentiert, um eine genauere taxonomische Zuordnung vorzunehmen. Anhand der Größe einiger Wirbel erreichten die längsten Individuen wohl Ausmaße von 50 cm. Die Amiidae sind in der Na-Duong-Formation besonders häufig. An Karpfenartigen treten unter anderem Barben und Bitterlinge sowie Angehörige der Cultrinae und der Xenocypridinae hervor. Die Barben sind mit mehreren Arten nachgewiesen, die sich anhand ihrer vorderen Zähne unterscheiden lassen. Von diesen ist eine nur aus der Na-Duong-Formation bekannt, die anderen kommen in der Rhin-Chua-Formation vor. Letzteres gilt auch für alle anderen Formen der Karpfenartigen. Eine Besonderheit bilden kleine, nur 3 mm große Schlundzähne, die der Gattung Planktophaga zugewiesen werden. Sie steht in der Verwandtschaft der Marmorkarpfen, die heute im südlichen Ostasien vorkommen, und bildet den ältesten Fossilbeleg dieser Gruppe. Im Gegensatz zu den rezenten Marmorkarpfen waren aber die Vertreter der Gattung Planktophaga wesentlich kleiner.[3]
Reptilien

Es liegen bisher rund 150 Reste an Reptilien vor, die fast ausschließlich aus der basalen Kohelschicht 80 stammen. Ein Drittel nehmen Krokodile ein. Die Panzerechsen kommen sowohl als vollständige oder teilerhaltene Skelette als auch als einzelne Schädel oder isolierte Knochen vor. Zusätzlich sind auch Koprolithen aufgefunden worden. Es können drei Morphotypen unterschieden werden: eine langschnauzige große, bis zu 6 m lange Form, die den Echten Krokodilen nahesteht, eine breitschnauzige Form der Alligatoren mit rund 2 m Gesamtlänge und ein singulärer, 50 cm langer Schädel eines langschnauzigen Gavials. Der Gavial zeigt ein homodontes Gebiss, was ihn wohl als spezialisierten Fischfresser auszeichnet. Die krokodil- und alligatorartigen Formen weisen hingegen eine heterodonte Bezahnung auf. Bei beiden Formen lassen recht breite hintere Zähne auf hartschalige Nahrung schließen. Der Alligator wurde als Orientalosuchus beschrieben, basierend auf rund 30 Individuen, wodurch nicht nur der Schädel, sondern auch Teile des Körperskelettes relativ gut bekannt sind.[10][2][3]

Das restliche Material entfällt auf Schildkröten. Überliefert sind ausschließlich die Panzer. Diese zeigen starke Pressungserscheinungen, so dass das Oberteil (Carapax) teilweise direkt auf dem Unterteil (Plastron) auflagert. Schädel oder Teile des Bewegungsapparates fehlen. Die Schildkröten sind relativ divers mit möglicherweise rund einem halben Dutzend an Arten, die sich aus Altwelt-Sumpfschildkröten und aus Weichschildkröten zusammensetzen. Bisher wurde lediglich von ersterer Gruppe die Gattung Banhxeochelys genauer bestimmt, die rund 30 Funde auf sich vereint. Vertreten sind hierbei sowohl ausgewachsene Individuen als auch Jungtiere. Die Panzerlänge der ausgewachsenen Tiere variiert zwischen 28 und 31 cm. Insgesamt zeigt Banhxeochelys eine recht hohe Variabilität, die einerseits auf das Lebensalter, andererseits auch auf individuelle Besonderheiten zurückzuführen ist.[11][2][3]
Säugetiere
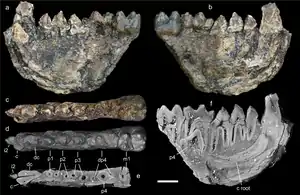
Unter den Säugetieren, die ebenfalls weitgehend in der Schicht 80 aufgefunden wurden, heben sich vor allem die Huftiere hervor. Mit Epiaceratherium wurde ein recht urtümlicher Vertreter der Nashörner entdeckt, der ansonsten eher aus dem westlichen Eurasien bekannt ist. Belegt ist er über einen einzelnen Schädel. Als typisch kann das Fehlen des unteren Eckzahns und hintersten Schneidezahns gewertet werden. Mit mehreren Schädeln, Unterkiefern und einzelnen isolierten Zähnen sowie einigen postcranialen Skelettelementen sind die Anthracotheriidae dokumentiert. Diese bilden eine ausgestorbene Gruppe der Paarhufer mit einer möglichen näheren Verwandtschaft zu den Flusspferden. Wie die heutigen Flusspferde lebten die Tiere wohl auch semi-aquatisch. Mit Anthracokeryx, Bothriogenys und Bakalovia sind bisher wenigstens drei Gattungen aus Na Dương identifiziert. Die Bestimmung von letzterer ist aber nicht eindeutig, da auch ein Verweis der Fossilreste zu Elomeryx möglich ist. Deren Fossilreste schließen mehrere Gebissreste ein, von denen rund ein halbes Dutzend nicht ausgewachsene Individuen repräsentieren.[3][12] Abseits der Huftiere wurde ein Unterkiefer zu Anthradapis verwiesen, eine Form der Primaten aus der Gruppe der Adapiformes. Das gut 5 kg schwere Tier gehört in den Verwandtschaftskreis weiterer südostasiatischer Vertreter. Von diesen unterscheidet sich aber Anthradapis in der Beibehaltung des vorderen Prämolaren, so dass er wohl einer eigenen Entwicklungslinie angehört.[13]
Altersstellung
Der Entstehungszeitraum für die Na-Duong- und die Rhin-Chua-Formation wurde in der forschungsgeschichtlichen Vergangenheit unterschiedlich bewertet. Die meisten Überlegungen basieren auf geologischen und biostratigraphischen Erwägungen. Vor allem Ende des 20. und Anfang des 21. Jahrhunderts nahm man für die Na-Duong-Formation eine Ablagerung in einer Zeitspanne vom Oligozän bis Oberen Miozän an, für die Rhin-Chua-Formation entsprechend vom Oligozän bis Pliozän. Grundlage hierfür bildet die Entstehung der Becken im Zuge der Herausformung der Ailao Shan-Red River fault zone und der mit ihr verbundenen sekundären Verwerfungsstrukturen. Da diese wiederum aus der Kollision des Indischen Subkontinentes mit der asiatischen Landmasse vor rund 50 bis 40 Millionen Jahren resultierten, ist dieses Ereignis als unterer Alterswert anzusehen. Für die Becken entlang des Cao Bang-Tien Yen fault system wurde eine zeitlich versetzte Entstehung angenommen, die im Nordwesten begann und sich kontinuierlich nach Südosten fortsetzte.[6] Nachfolgende Analysen fanden dafür aber nur wenig Bestätigung.[7]
Die bisher einzigen absoluten Altersdatierungen aus den Sandsteinen der Na-Duong-Formation erfolgten Anfang der 2000er Jahre und verwendeten Zirkon-Mineralien. Die mit Hilfe der Uran-Blei-Methode ermittelten Werte lagen bei durchschnittlich 185 Millionen Jahren, was einerseits als deutlich zu alt für den Entstehungszeitraum der Gesteinseinheit zu bewerten ist, andererseits als Ressource der Sedimentation älteres Material aus weiter nördlich gelegenen Regionen annehmen lässt.[14] Getätigte paläomagnetische Messungen erbrachten, dass die Mineralien eines Großteils der Na-Duong-Formation und eines Teils der Rhin-Chua-Formation nach ihrer Einbettung erneut magnetisiert worden waren und so keine verwertbaren Aussagen liefern. Lediglich die obersten 40 m der Rhin-Chua-Formtion ließen jeweils einzelne Phasen einer inversen und einer reversen Polarisierung erkennen. Die Abschnitte sind allerdings zu kurz, um sie bestimmten Ereignissen der bestehenden Magnetostratigraphie zuzuweisen.[3]
Aus genannten Gründen ist daher derzeit eine gesicherte Altersdatierung nur über die Biostratigraphie zu erbringen. Hierzu bestehen aber etwas unterschiedliche Ansätze. Die Pflanzengemeinschaft zeigt Ähnlichkeiten zu anderen fossilen Floren in der Region, wie sie beispielsweise für die Shangcun-Formation des Maoming-Beckens und für das Nanning-Becken, beide im südlichen China, beschrieben wurden. Als stratigraphische Anzeiger können hier die Hemlocktannen und Knöteriche gesehen werden, deren erstes Auftreten in das Untere Oligozän vor 34 bis 28 Millionen Jahren fällt.[15] Vergleichbares ist für einige Farne wie Hypolepis und Pyrrosia oder für die Riemenblumengewächse zu sagen.[2][7] Etwas abweichend davon stehen die Aussagen der Säugetierstratigraphie. Der urtümliche Nashornvertreter Epiaceratherium ist in Eurasien seit dem Oberen Eozän belegt und hielt sich dort noch bis zum Oligozän. Der aus der Na-Duong-Formation vorliegende Angehörige zeigt relativ ursprüngliche Merkmale, was ihn mit den älteren Formen aus dem westlichen Eurasien verbindet. Ebenso werden für die Angehörigen der Anthracotheriidae Anthracokeryx und Bothriogenys eher primitive Charakteristika angegeben, die sie mit den obereozänen Verwandten in Ost- und Südostasien teilen. Dadurch könnte zumindest die wirbeltierführende Schicht aus dem basalen Kohleflöz der Na-Duong-Formation noch in das ausgehende Eozän gehören und somit ein Alter von 38 bis 34 Millionen Jahren aufweisen.[3][12]
Landschaftsrekonstruktion
Die Na-Duong-Formation entstand innerhalb eines aus Sümpfen und Trockenland bestehenden Ökosystems. Das wasserreiche Milieu wird unter anderem durch die Tonsteine, die Steinkohle und die Sandsteine angezeigt. Erstere beiden entstanden unter Bedingungen von Stillgewässern, letztere sind auf Bäche und kleinere Flüsse zurückzuführen. Für das Hauptkohleflöz mit der Schichtnummer 80 ist von einer Bildung in einem See mit anoxischen Bedingungen auszugehen. Er nahm den südlichen Teil des heutigen Na-Dương-Beckens ein. Abgeleitet wird dies aus dem weitgehenden Fehlen der Kohle in den nördlichen Beckenbereichen. Die in den oberen Abschnitten des Kohleflözes eingeschalteten schluffigen bis sandigen Tonsteine verweisen auf einen größeren Materialeintrag durch Fließgewässer, das auch einen größeren Anteil an pflanzlichen Resten mit sich führte. Diese Fließgewässer gehörten vermutlich zu einem größeren Flusssystem, das zur damaligen Zeit vom Gebiet des heutigen Tibets über das des Jangtsekiang in das Südchinesische Meer entwässerte.[14][16] Die in dem See auftretende Fauna bestand aus Fossilverwandten des Kahlhechts und Karpfenartigen, verschiedenen Schildkröten und Krokodilen. Wahrscheinlich waren auch Weichtiere anwesend und bildeten die Nahrungsgrundlage vieler Fische und Schildkröten, doch haben sich ihre Gehäuse in dem mit Huminsäure angereicherten Wasser zersetzt. Die Fische dienten wiederum den gavialartigen Krokodilen als Nahrung, während die Schildkröten und möglicherweise auch die Landsäugetiere von den alligator- und krokodilartigen Panzerechsen erlegt wurden. Darauf deutet bei ersteren zumindest ein singulärer Massenfund an Panzern hin, die auf einer Fläche von 2,5 m² lagen und Bissmarken von Krokodilszähnen aufweisen.[2][11] Vor und nach der Bildung des Hauptkohleflözes bestand der See allerdings wohl eher als flacher Tümpel, der von Lotosblumenwiesen durchsetzt war und in dem Sumpfdeckelschnecken und kleinere Barben lebten. Außerhalb des Sees kamen sumpfartige Landschaften oder Überflutungsflächen vor, worauf die sehr dünnen Kohleschichten und einzelne Tonsteinlagen mit pedogenetischen Überprägungen hindeuten. Diese Areale waren wohl mit Wald bedeckt. Zahlreiche Harzklumpen lassen hier vor allem Flügelfruchtgewächse annehmen. Unverlagerte Baumstümpfe geben Stammdurchmesser von 7 bis 50 cm an. Die individuellen Abstände zueinander betragen 2 bis 5 m, was zu einer Baumdichte von rund 600 je Hektar führt. Die Kronenhöhe lag möglicherweise bei 35 m. Die Werte stimmen in etwa mit denen vergleichbarer heutiger Landschaften auf Borneo und Sumatra überein. Die Wälder bewohnten verschiedene Großsäuger, die aus Nashörnern, weiteren Huftieren wie den Anthracotheriidae und Primaten bestanden. Sie ernährten sich überwiegend blätterfressend. Während die Nashörner wohl reine Landbewohner waren, lebten die Anthracotherien aufgrund ihres flusspferdartigen Körperbaus semi-aquatisch. Eventuell fielen sie auch den größeren Krokodilen zum Opfer, worauf das übermäßig häufige Auftreten von Jungtieren bei Bakalovia hinweist.[6][3][7]
Die unteren Abschnitte der Rhin-Chua-Formation gehen auf ein limnisches Bildungsmilieu zurück. Das Vorkommen von Greigit in einigen Bereichen ist wahrscheinlich auf phasenweise vorherrschende anoxische Bedingungen zurückzuführen. Diese Sedimentlagen sind weitgehend fossilfrei. In anderen wiederum kommen verschiedene Weichtiere und Karpfenartige vor, so unter anderem eine Fossilform der Marmorkarpfen. Dadurch sind variable Sauerstoffverhältnisse für den Seegrund anzunehmen, was eventuell mit schwankenden Wassertiefen verbunden war. Zahlreiche Rippelmarken in den Sandsteinlagen geben in der jüngsten aufgeschlossenen Phase des Sees eine stark abnehmende Wassertiefe wieder. Die hier überlieferte Fauna ist reichhaltig an Weichtieren wie Sumpfdeckelschnecken sowie Fluss- und Teichmuscheln. Teilweise bilden diese schillartige Lagen. Ebenso kommen Fische vor, hier werden allerdings die bisherigen Karpfenartigen weitgehend durch größere Barben ersetzt.[6][3][7]
Die Rekonstruktion des Paläoklimas erfolgte auf unterschiedliche Weise unter Heranziehung von sowohl Pollenanalysen als auch Makroresten. Das Pollenspektrum ist relativ artenarm, was teilweise auf den hohen Verwitterungsgrad zurückgeführt werden kann, setzt sich aber aus Nackt- und Bedecktsamern sowie Gefäßsporenpflanzen zusammen. Einige der Nacktsamer wie die Tannen oder die Hemlocktannen und auch der Bedecktsamer wie die Ulmen und die Weiden verweisen auf eher warm-gemäßigte Klimaverhältnisse. Andere Formen sind dem gegenüber an eher tropische Klimate angepasst, wozu unter anderem einige Farne wie Pteris oder Hartriegelartige wie die Tupelogewächse zählen. Ein vergleichbar gemischtes Bild geben die Makrofossilien wieder. Einzelne unidentifizierte Blätter weisen gezackte Ränder auf, was eher typisch für Pflanzen aus gemäßigten Gebieten ist. Dem gegenüber können Pflanzen wie die Flügelfruchtgewächse als Indikatoren für tropisch-warme Bereiche herangezogen werden. Deren heutige Vertreter kommen in Südostasien und im südlichen Ostasien vor und keimen nicht bei Temperaturen unterhalb von 15 °C. Die Regionen ihrer Verbreitung sind durch eine Jahresdurchschnittstemperatur von 20 °C und einer monatlichen Minimaltemperatur nicht unter 10 °C gekennzeichnet. Ähnliches kann zu den Bauhinien gesagt werden. Unter den Wirbeltieren lassen sich die Krokodile als Anzeiger für warme Klimaverhältnisse heranziehen. Heutige Krokodile sind nur in Landschaften verbreitet, deren Jahresdurchschnittstemperaturen nicht unter rund 14 °C und im kältesten Monat nicht unter 5 °C liegen. Die großen Ausmaße der Krokodile aus dem Na-Dương-Becken verweisen darauf, dass diese Angaben wohl eher Minimalwerte darstellen. Des Weiteren fehlen den Ablagerungen des Na-Dương-Beckens charakteristisch oxidierte Paläoböden oder Karbonatausfällung, die auf stärkere Bodenbildungsprozesse zurückzuführen wären. Rückschließend daraus war die Landschaft wohl feuchten Klimabedingungen ausgesetzt, die keinen jahreszeitlichen Schwankungen unterlagen.[3][7]
Vergleich mit regional und überregional bedeutenden Fundstellen
Entlang des Cao Bang-Tien Yen fault system sind weitere Becken ausgebildet, die ebenfalls Fossilien enthalten, jedoch nicht den Reichtum des Na-Dương-Beckens erreichen. Von Bedeutung ist hier das Cao-Bằng-Becken in der gleichnamigen nordvietnamesischen Provinz, das sich rund 120 km nordwestlich befindet und gut 65 km² Fläche einnimmt. Die ebenfalls kohleführenden Schichten werden teilweise auch der Na-Duong-Formation zugewiesen, ebenso bestehen aus palynologischer Sicht Ähnlichkeiten, so dass von einer vergleichbaren Zeitstellung ausgegangen werden kann. Die geologischen Daten verweisen hier analog zum Na-Dương-Becken auf einen See mit angrenzenden Überschwemmungsflächen. Um so auffälliger ist, dass sich die Weichtier- und Fischfauna beider Becken unterscheiden mit nur wenigen sich überschneidenden Formen. An Landwirbeltieren wurde im Cao-Bằng-Becken bisher lediglich ein Krokodil nachgewiesen, das vermutlich dem Sunda-Gavial nahesteht.[6][2][8][17] Das Hang-Mon-Becken in der nordvietnamesischen Provinz Sơn La nahe der Grenze zu Laos steht mit der Son La fault zone in Verbindung, die südwestlich parallel zur Ailao Shan-Red River fault zone verläuft. Es ist nur wenige Quadratkilometer groß und seit Ende der 1970er Jahre als Fossillagerstätte bekannt. Die Ablagerungen datieren in das Untere Miozän vor gut 23 bis 21 Millionen Jahren. Angezeigt wird dies unter anderem durch die Wirbeltierfauna, die mit Nashörnern wie Protaceratherium und Bugtirhinus, Wiederkäuern wie Dorcatherium und Dorcabune oder Schweineartigen wie Hyotherium deutlich moderner wirkt.[2][18]
Das Maoming-Becken in der südchinesischen Provinz Guangdong zeichnet sich durch seine außerordentliche lange Ablagerungssequenz aus, die insgesamt 2700 m an paläogenen Sedimenten verteilt auf sechs Gesteinseinheiten einschließt.[19] Die generelle Ähnlichkeit der Pollenflora der in das Oligozän datierenden Shangcun-Formation mit der des Na-Dương-Beckens wurde bereits erwähnt. Hier ist jedoch besonders die Youganwo-Formation hervorzuheben, die sich im Oberen Eozän herausbildete. An Wirbeltieren wurden neben Schildkröten und Krokodilen auch verschiedentlich Säugetiere belegt. Neben einzelnen Funden aus der entfernteren Verwandtschaft der Nashörner wie Cadurcodon, einem gut 1,4 t schweren Vertreter der Amynodontidae, kam hier mit Maobrontops auch ein Angehöriger der Brontotheriidae zu Tage, der nur etwa halb so schwer wurde. Daneben trat eine Form der säbelzahnigen Nimravidae auf, die mit den Katzen näher verwandt sind.[20][21][22]
Eine der wichtigsten Fundstellen in gesamt Südostasien ist die Pondaung-Formation im zentralen Myanmar. Die zahlreichen Fossilaufschlüsse sind bereits seit dem Beginn des 20. Jahrhunderts bekannt, besonders intensive Untersuchungen fanden in den 1990er und 2000er Jahren statt. Nach Analysen mittels der Fission-Track-Methode gehört die Gesteinseinheit mit einem Alter von rund 37 Millionen Jahren dem Oberen Eozän an.[23] In der überaus reichhaltigen Fauna sind neben Fischen und Reptilien wie Schlangen, Schildkröten, Eidechsen und Krokodilen einige wenige Vögel und vor allem Säugetiere präsent. Hier treten unter anderem die Huftiere recht formenreich auf. Zu den Unpaarhufern gehören etwa das urtümliche Nashorn Teletaceras oder verschiedene Brontotherien wie Sivatitanops und Bunobrontops, Amynodonten wie Paramynodon und Chalicotherien wie Eomoropus.[24][25] Die Paarhufer wiederum können archaische Formen wie Magwetherium oder Pakkokuhyus vorweisen, ersteres gehört zur Gruppe der Diacodexeidae, letzteres möglicherweise zu den Dichobunidae.[26][27] Die Anthracotheriidae sind unter anderem mit Anthracokeryx und Anthracotherium belegt.[23][12] Daneben tritt auch ein breiteres Spektrum an Primaten auf, darunter Paukkaungia aus der Gruppe der Adapiformes oder Ganlea und Pondaungia aus der Gruppe der Affen.[28][29] Beutegreifer wiederum werden durch mehrere Vertreter der Hyaenodonta und der Raubtiere angezeigt.[30][31]
Kurz soll noch das Krabi-Becken in der gleichnamigen Provinz im südlichen Thailand erwähnt werden. Die paläogenen, kohleführenden Ablagerungen gehören aus magnetostratigraphischer Sicht in den Übergang vom Oberen Eozän zum Unteren Oligozän. Die mehr als 30 bekannten Säugetierarten werden durch die Paarhufer dominiert, hier allen voran die Anthracotherien mit Anthracotherium, Anthracokeryx und Bothriogenys. Unter den Primaten sind Siamopithecus und Krabia erwähnenswert, die Beziehungen zu Pondaungia aufweisen und den frühen Affen angehören, während Muangthanhinius in seinen genauen Verwandtschaftsverhältnissen uneindeutig ist, aber wohl den Adapiformes nahesteht.[32][33][34]
Forschungsgeschichte
Im Vergleich zu den anderen Becken entlang des Cao Bang-Tien Yen fault system ist das Na-Dương-Becken relativ gut untersucht. Zu verdanken ist dieser Umstand dem Tagebaubetrieb zur Erschließung der Steinkohle, mehreren weiteren natürlichen Aufschlüssen und zahlreichen Bohrungen zur Erkundung des Untergrundes. Untersuchungen reichen teilweise bis in die 1970er Jahre zurück und wurden in dieser Zeit überwiegend von vietnamesischen Wissenschaftlern getätigt. Den Schwerpunkt bildeten dabei neben sedimentologischen auch paläobotanische Fragestellungen. Im Ergebnis davon wurde zumeist eine relativ junge neogene Bildung des Beckens angenommen.[6]
Anfang der 2000er Jahre kam es zu einer Intensivierung der Arbeiten im Na-Dương-Becken. Hierzu gehören einzelne Untersuchungen zur Entstehung des Cao Bang-Tien Yen fault system, das bis zu diesem Zeitpunkt und im Gegensatz zur weiter südlich gelegenen Ailao Shan-Red River fault zone nur wenig im Zentrum wissenschaftlichen Interesses gestanden hatte, und der damit verbundenen Beckenstrukturen.[5] Federführende Arbeiten wurden hier vor allem von Anna Wysocka von der Universität Warschau durchgeführt. Ausgehend von ähnlichen geologischen Strukturen entlang der Ailao Shan-Red River fault zone,[35][36] widmete sie sich folgend den weiter nördlich gelegenen Becken des Cao Bang-Tien Yen fault system. In mehreren Kampagnen, die bis 2016/2017 anhielten, fokussierte sie sich wiederum auf verschiedene sedimentologische und palynologische Aspekte und kam zu dem Schluss einer deutlich älteren zeitlichen Entstehung des Na-Dương-Beckens.[6][7]
Nahezu zeitgleich begannen Untersuchungen der Senckenberg Gesellschaft für Naturforschung in Verbindung mit der Eberhard Karls Universität Tübingen in mehreren Becken im Norden Vietnams unter Leitung von Madelaine Böhme, darunter auch das Na-Dương-Becken. Bereits in der ersten Untersuchungsphase von 2008 bis 2009 konnten dabei dort reiche und bis dahin unbekannte Fossillager in den Kohleablagerungen der Na-Duong-Formation entdeckt werden. Weitere Tätigkeiten vor Ort fanden im Jahr 2012 statt. Die Arbeiten führten zur Beschreibung mehrerer neuer Fossilformen und präzisierten die Alterstellung.[2][3]
Literatur
- Madelaine Böhme, Jérôme Prieto, Simon Schneider, Nguyen Viet Hung, Do Duc Quang und Dang Ngoc Tran: The Cenozoic on-shore basins of Northern Vietnam: Biostratigraphy, vertebrate and invertebrate faunas. Journal of Asian Earth Sciences 40 (2), 2011, S. 672–687, doi:10.1016/j.jseaes.2010.11.002
- Madelaine Böhme, Manuela Aiglstorfer, Pierre-Olivier Antoine, Erwin Appel, Philipe Havlik, Grégoire Métais, Laq The Phuc, Simon Schneider, Fabian Setzer, Ralf Tappert, Dang Ngoc Tran, Dieter Uhl und Jérôme Prieto: Na Duong (northern Vietnam) – an exceptional window into Eocene ecosystems from Southeast Asia. Zitteliana A 53, 2013, S. 120–167
- Anna Wysocka, Phan Dong Pha, Ewa Durska, Urszula Czarniecka, Do Van Thang, Anna Filipek, Nguyen Quoc Cuong, Dang Minh Tuan, Nguyen Xuan Huyen, Hoang Van Tha und Radosław Staniszewski: The Na Duong Basin (North Vietnam): A key for understanding Paleogene basin evolution in relation to the left-lateral Cao Bang-Tien Yen Fault. Journal of Asian Earth Sciences 195, 2020, S. 104350, doi:10.1016/j.jseaes.2020.104350
Einzelnachweise
- Nguyen Duc Thao: Development strategy and the application of circulating fluidized bed combustion in power generation projects of Vietnam National Coal Corporation (VINACOAL). Electricity Supply Industry in Transition: Issues and Prospect for Asia, 14-16 January 2004, S. 16-24–16-34 ()
- Madelaine Böhme, Jérôme Prieto, Simon Schneider, Nguyen Viet Hung, Do Duc Quang und Dang Ngoc Tran: The Cenozoic on-shore basins of Northern Vietnam: Biostratigraphy, vertebrate and invertebrate faunas. Journal of Asian Earth Sciences 40 (2), 2011, S. 672–687, doi:10.1016/j.jseaes.2010.11.002
- Madelaine Böhme, Manuela Aiglstorfer, Pierre-Olivier Antoine, Erwin Appel, Philipe Havlik, Grégoire Métais, Laq The Phuc, Simon Schneider, Fabian Setzer, Ralf Tappert, Dang Ngoc Tran, Dieter Uhl und Jérôme Prieto: Na Duong (northern Vietnam) – an exceptional window into Eocene ecosystems from Southeast Asia. Zitteliana A 53, 2013, S. 120–167
- M. P. Searle: Role of the Red River Shear zone, Yunnan and Vietnam, in the continental extrusion of SE Asia. Journal of the Geological Society, London, 163, 2006, S. 1025–1036
- Manuel Pubellier, Claude Rangin, Phung Van Phach, Bui Cong Que, Doan The Hung und Chan Lung Sang: The Cao Bang-Tien Yen Fault: Implications on the relationships between the Red River Fault and the South China Coastal Belt. Advances in Natural Sciences 4 (4), 2003, S. 347–361
- Anna Wysocka: Sedimentary environments of the Neogene basins associated with the Cao Bang–Tien Yen Fault, NE Vietnam. Acta Geologica Polonica 59 (1), 2009, S. 45–69
- Anna Wysocka, Phan Dong Pha, Ewa Durska, Urszula Czarniecka, Do Van Thang, Anna Filipek, Nguyen Quoc Cuong, Dang Minh Tuan, Nguyen Xuan Huyen, Hoang Van Tha und Radosław Staniszewski: The Na Duong Basin (North Vietnam): A key for understanding Paleogene basin evolution in relation to the left-lateral Cao Bang-Tien Yen Fault. Journal of Asian Earth Sciences 195, 2020, S. 104350, doi:10.1016/j.jseaes.2020.104350
- Thomas A. Neubauer, Simon Schneider, Madelaine Böhme und Jérôme Prieto: First records of freshwater rissooidean gastropods from the Palaeogene of Southeast Asia. Journal of Molluscan Studies 78, 2012, S. 275–282, doi:10.1093/mollus/eys011
- Simon Schneider, Madelaine Böhme und Jérôme Prieto: Unionidae (Bivalvia; Palaeoheterodonta) from the Palaeogene of northern Vietnam: exploring the origins of the modern East Asian freshwater bivalve fauna. Journal of Systematic Palaeontology 11 (3), 2013, S. 337–357, doi:10.1080/14772019.2012.665085
- Tobias Massonne, Davit Vasilyan, Márton Rabi und Madelaine Böhme: A new alligatoroid from the Eocene of Vietnam highlights an extinct Asian clade independent from extant Alligator sinensis. PeerJ 7, 2019, S. e7562, doi:10.7717/peerj.7562
- Rafaella C. Garbin, Madelaine Böhme und Walter G. Joyce: A new testudinoid turtle from the middle to late Eocene of Vietnam. PeerJ 7, 2019, S. e6280, doi:10.7717/peerj.6280
- Stéphane Ducrocq, Mouloud Benammi, Olivier Chavasseau, Yaowalak Chaimanee, Kantapon Suraprasit, Phan Dong Pha, Vu Le Phuong, Phung Van Phach und Jean-Jacques Jaeger: New Anthracotheres (Cetartiodactyla, Mammalia) from the Paleogene of Northeastern Vietnam: Biochronological Implications. Journal of Vertebrate Paleontology 35 (3), 2015, S. e929139, doi:10.1080/02724634.2014.929139
- Olivier Chavasseau, Yaowalak Chaimanee, Stéphane Ducrocq, Vincent Lazzari, Phan Dong Pha, Mana Rugbumrung, Jérôme Surault, Dang Minh Tuan und Jean-Jacques Jaeger: A new primate from the late eocene of Vietnam illuminates unexpected strepsirrhine diversity and evolution in Southeast Asia. Scientific Reports 9, 2019, S. 19983, doi:10.1038/s41598-019-56255-8
- Long van Hoang, Fu-Yuan Wu, Peter D. Clift, Anna Wysocka und Anna Swierczewska: Evaluating the evolution of the Red River system based on in situ U-Pb dating and Hf isotope analysis of zircons. Geochemistry, Geophysics, Geosystems 10 (11), 2009, S. Q11008, doi:10.1029/2009GC002819
- Alexei B. Herman, Robert A. Spicer, Galina N. Aleksandrova, Jian Yang, Tatiana M. Kodrul, Natalia P. Maslova, Tewresa E. V. Spicer, Gang Chen und Jian-Hua Jin: Eocene–early Oligocene climate and vegetation change in southern China: Evidence from the Maoming Basin. Palaeogeography, Palaeoclimatolology, Palaeoecology 479, 2017, S. 126–137
- Peter D. Clift, Andrew Carter, Anna Wysocka, Long Van Hoang, Hongbo Zheng und Nikki Neubeck: A Late Eocene-Oligocene Through-Flowing River Between the Upper Yangtze and South China Sea. Geochemistry, Geophysics, Geosystems 21 (7), 2020, S. e2020GC009046, doi:10.1029/2020GC009046
- Anna Wysocka, Phan Dong Pha, Ewa Durska, Urszula Czaniecka, Do Van Thang, Anna Filipek, Nguyen Quoc Cuong, Dang Minh Tuan, Nguyen Xuan Huyen und Hoang Van Tha: New data on the continental deposits from the Cao Bang Basin (Cao Bang-Tien Yen Fault Zone, NE Vietnam) – biostratigraphy, provenance and facies pattern. Acta Geologica Polonica 68 (4), 2018, S. 689–709, doi:10.1515/agp-2018-0037
- Jérôme Prieto, Pierre-Olivier Antoine, Jan van der Made, Grégoire Métais, Laq The Phuc, Quý Trương Quan, Simon Schneider, Dang Ngoc Tran, Davit Vasilyan, Luong The Viet und Madelaine Böhme: Biochronological and palaeobiogeographical significance of the earliest Miocene mammal fauna from Northern Vietnam. Palaeobiodiversity and Palaeoenvironments 98, 218, S. 287–313, doi:10.1007/s12549-017-0295-y
- G. N. Aleksandrova, T. M. Kodrul und J. H. Jin: Palynological and Paleobotanical Investigations of Paleogene Sections in the Maoming Basin, South China. Stratigraphy and Geological Correlation 23 (3), 2015, S. 300–325
- Alexander Averianov, Ekaterina Obraztsova, Igor Danilov, Pavel Skutschas und Jianhua Jin: First nimravid skull from Asia. Scientific Reports 6, 2016, S. 25812 doi:10.1038/srep25812
- Alexander Averianov, Igor Danilov, Jianhua Jin und Yingyong Wang: A new amynodontid from the Eocene of South China and phylogeny of Amynodontidae (Perissodactyla: Rhinocerotoidea). Journal of Systematic Palaeontology 15, 2017, S. 927–945
- Alexander Averianov, Igor Danilov, Wen Chen und Jianhua Jin: A new brontothere from the Eocene of South China. Acta Palaeontologica Polonica 63 (1), 2018; S. 189–196 doi:10.4202/app.00431.2017
- Takehisa Tsubamoto, Masanaru Takai, Naoko Egi, Nobuo Shigehara, Soe Thura Tun, Aye Ko Aung, Aung Naing Soe und Tin Thein: The Anthracotheriidae (Mammalia; Artiodactyla) from the Eocene Pondaung Formation (Myanmar) and comments on some other anthracotheres from the Eocene of Asia. Paleontological Research 4, 2002, S. 363–384 ()
- Patricia A. Holroyd und Russell L. Ciochon: Bunobrontops savagei: A new genus and species of brontotheriid perissodactyl from the Eocene Pondaung fauna of Myanmar. Journal of Vertebrate Paleontology 20 (2), 2000, S. 408–410
- Patricia A. Holroyd, Takehisa Tsubamoto, Naoko Egi, Russell L. Ciochon, Masanaru Takai, Soe Thura Tun, Chit Sein und Gregg F. Gunnell: A Rhinocerotid Perissodactyl from the Late Middle Eocene Pondaung Formation, Myanmar. Journal of Vertebrate Paleontology 26 (2), 2006, S. 491–494
- Stéphane Ducrocq, Aung Naing Soe, Chit Sein, Vincent Lazzari, Yaowalak Chaimanee, Xavier Valentin und Jean-Jacques Jaeger: First record of a diacodexeid artiodactyl in the middle Eocene Pondaung Formation (Myanmar). Paläontologische Zeitschrift 90, 2016, S. 611–618, doi:10.1007/s12542-016-0283-y
- Stéphane Ducrocq: Pakkokuhyus and Progenitohyus (Artiodactyla, Mammalia) from the Eocene of Southeast Asia are not Helohyidae: paleobiogeographical implications. Paläontologische Zeitschrift 93 (8), 2019, S. 105–113, doi:10.1007/s12542-018-0425-5
- K. Christopher Beard, Laurent Marivaux, Soe Thura Tun, Aung Naing Soe, Yaowalak Chaimanee, Wanna Hitoon, Bernard Marandat Htun Htun Aung und Jean-Jaques Jaeger: New Sivaladapid Primates from the Eocene Pondaung Formation of Myanmar and the Anthropoid Status of Amphipithecidae. Bulletin of Carnegie Museum of Natural History·39, 2007, S. 67–76
- K. Christopher Beard, Laurent Marivaux, Yaowalak Chaimanee, Jean-Jacques Jaeger, Bernard Marandat, Paul Tafforeau, Aung Naing Soe, Soe Thura Tun und Aung Aung Kyaw: A new primate from the Eocene Pondaung Formation of Myanmar and the monophyly of Burmese amphipithecids. Proceedings of the Royal Society B 276, 2009, S. 3285–3294, doi:10.1098/rspb.2009.0836
- Louis de Bonis, Floreal Solé, Yaowalak Chaimanee, Aung Naing Soe, Chit Sein, Vincent Lazzari, Olivier Chavasseau und Jean-Jacques Jaeger: New hyaenodonta (Mammalia) from the middle Eocene of Myanmar. Comptes Rendus Palevol 17 (6), 2018, S. 357–365
- Takehisa Tsubamoto, Naoko Egi, Masanaru Takai, Nobuo Shigehara, Hisashi Suzuki, Takeshi Nishimura, Hiroaki Ugai, Maung Maung, Chit-Sein, Soe Thura Tun, Aung Naing Soe, Aye Ko Aung, Tin Thein, Thaung-Htike und Zin-Maung-Maung-Thein: A summary of the Pondaung fossil expeditions. Asian Paleoprimatology 4, 2006, S. 1–66
- Laurent Marivaux, Yaowalak Chaimanee, Paul Tafforeau und Jean-Jacques Jaeger: New strepsirrhine primate from the late Eocene of Peninsular Thailand (Krabi Basin). American Journal of Physical Anthropology 130 (4), 2006, S. 425–434, doi:10.1002/ajpa.20376
- Yaowalak Chaimanee, Olivier Chavasseau, Vincent Lazzari, Adélaïde Euriat und Jean-Jacques Jaeger: A new Late Eocene primate from the Krabi Basin (Thailand) and the diversity of Palaeogene anthropoids in southeast Asia. Proceedings of the Royal Society B 280, 2013, S. 20132268, doi:10.1098/rspb.2013.2268
- Mouloud Benammi, Yaowalak Chaimanee, Jean-Jacques Jaeger, Varavudh Suteethorn und Stéphane Ducrocq: Eocene Krabi basin (southern Thailand): Paleontology and magnetostratigraphy. Geological Society of America Bulletin 113 (2), 2001, S. 265–273
- Anna Wysocka und Anna Świerczewska: Alluvial deposits from the strike-slip fault Lo River Basin (Oligocene/Miocene), Red River Fault Zone, north-western Vietnam. Journal of Asian Earth Sciences 21, 2003, S. 1097–1112
- Anna Wysocka und Anna Świerczewska: Tectonically-controlled sedimentation of Cenozoic deposits from selected basins along the Vietnamese segment of the Red River Fault Zone. Acta Geologica Polonica 55 (2), 2005, S. 131–145
Weblinks
- Senckenberg: mehrere Pressemitteilungen: Vorläufer des europäischen Nashorns in Vietnam entdeckt vom 12. März 2014, Marmorkarpfen: Von 5 auf 150 Zentimeter in 37 Millionen Jahren vom 12. März 2014, Fossiler Krokodilverwandter entdeckt vom 8. Januar 2020
- Universität Tübingen: Vietnam, Universität Tübingen, Präsentation der Forschungsarbeiten