Edmontosaurus
Edmontosaurus ist eine Gattung von ornithopoden Dinosauriern aus der Gruppe der Hadrosauridae. Sie lebte in der späten Oberkreide (spätes CampanA und Maastricht) von Nordamerika. Edmontosaurus ist eine der häufigsten und am besten bekannten Dinosauriergattungen sowohl der USA als auch weltweit.[1]
| Edmontosaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
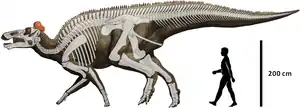
Skelett- und Lebendrekonstruktion von Edmontosaurus regalis | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (spätes CampaniumA und Maastrichtium) | ||||||||||||
| ca. 73 bis 66 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Edmontosaurus | ||||||||||||
| Lambe, 1917 | ||||||||||||
| Arten | ||||||||||||
| ||||||||||||
Etymologie
Die Gattung Edmontosaurus bzw. ihre Typusart Edmontosaurus regalis wurden 1917 vom kanadischen Paläontologen Lawrence Lambe anhand zweier Skelette mit Schädel beschrieben, die im Jahr 1912 von den Brüdern George und Levi Sternberg am Red Deer River in Alberta entdeckt und aufgesammelt wurden.[2] Benannt ist die Gattung nach der Edmonton-Formation, den Schichten, in denen die Funde gemacht wurden (heute als Horseshoe-Canyon-Formation der Edmonton-Gruppe ausgewiesen). Die Edmonton-„Formation“ ist wiederum nach der Hauptstadt Albertas benannt.
Merkmale

Skelett


Oben: Blick auf die Innenseite des Kiefers. Die vertikalen Reihen mit den an dieser Seite rautenförmigen, gekielten Zähnen (durch die Fossilisation schwarz gefärbt) sind gut zu erkennen.
Edmontosaurus ist ein großwüchsiger Hadrosauride ohne auffällige knöcherne Schädelauswüchse. Die Länge des Schädels beträgt bei ausgewachsenen Individuen mehr als einen Meter. Mit einer geschätzten Gesamtlänge von maximal 13 Metern und einem Gewicht von maximal 4 Tonnen war Edmontosaurus einer der größten Vertreter der Hadrosauriden.
Der Schädel von Edmontosaurus besitzt eine langgezogene, am vorderen Ende hadrosauriertypisch entenschnabelartig verbreiterte Schnauzenpartie (Rostrum). Der „Oberschnabel“ wird überwiegend vom Prämaxillare gebildet. Der vordere (orale) Rand des Prämaxillare ist nach hinten „umgeschlagen“ und bildet eine Art knöcherne „Lippe“ (engl. reflected margin of premaxilla). Das Rostrum zeichnet sich überdies durch eine sehr große äußere Nasenöffnung aus, die in eine Mulde eingesenkt ist (circumnariale Fossa). In ihrer Größe erinnert die Nasenöffnung an das Präorbitalfenster, das viele Dinosaurier und geologisch ältere Archosaurier aufweisen (vgl. → Triapsider Schädel). Jedoch ist das überwiegend vom Maxillare und Lacrimale gerahmte Präorbitalfenster bei den „höheren“ Ornithopoden generell reduziert.[3] Die vordere Hälfte der Kiefer ist zahnlos, die hintere mit einer Batterie aus bis zu 300 gekielten Zähnen bestückt, von denen aber immer nur ca. 100 aktiv genutzt wurden.
Von anderen kamm- und hornlosen Hadrosauriden mit großer Nasenöffnung unterscheidet sich Edmontosaurus darin, dass die Prämaxillarlippe besonders ausgeprägt ist und sich an den seitlichen (lateralen) Rändern des „Schnabels“ relativ weit nach hinten zieht. Ein weiteres Alleinstellungsmerkmal ist, dass die circumnariale Fossa, in ihrer hinteren unteren (caudoventralen) Partie besonders tief ist. Speziell kennzeichnend ist zudem eine taschenartige Aushöhlung des ventralen, an der hinteren Umrandung der Augenhöhle beteiligten Astes des PostorbitaleB und zwar an der Seite, die zur Augenhöhle weist (engl. postorbital pocket). Eine ähnliche Tasche, jedoch flacher, besitzt auch die an der Umrandung der Augenhöhle beteiligte Partie des Präfrontale. Zudem ist das Frontale an der Umrandung der Augenhöhle beteiligt.[4]
Das postcraniale Skelett unterscheidet sich, abgesehen von seiner Größe, nicht wesentlich von dem anderer Hadrosauriden oder Iguanodontiden. So sind u. a. die Vordergliedmaßen kürzer als die Hintergliedmaßen und die drei (gewichtstragenden) Finger bzw. Zehen der Extremitäten enden in hufartigen Klauen.
Weichteile

In einigen Fällen zeigen Exemplare von Edmontosaurus die Erhaltung von Weichgeweben, die normalerweise nicht fossil überliefert werden (siehe unten). Daher ist relativ viel über die Weichteilanatomie dieser Gattung bekannt. Anhand eines dieser Exemplare konnte zumindest bei Edmontosaurus regalis ein fleischiger Scheitelkamm („Hahnenkamm“) nachgewiesen werden.[5] Funde mit zweidimensionaler, schattenhafter Erhaltung von Haut zeigen, dass bei Edmontosaurus entlang der Wirbelsäule ein segmentierter Rückenkamm verlief, während andere Hadrosauriden einen durchgehenden Kamm besaßen.[6] Die Finger bzw. Vorderzehen waren von außen nicht einzeln zu sehen, sondern in einer Art „Fleisch-und-Hautfäustling“ eingebettet. Dies wurde früher als Schwimmhäute interpretiert. Heute wird eher davon ausgegangen, dass es sich um eine Anpassung handelt, die es dem Tier ermöglichte, sein Körpergewicht besser auf dem Untergrund zu verteilen.[7]
Lebensweise
| |
|---|---|
| Skelettrekonstruktion eines Individuums von Edmontosaurus annectens aus der Hell-Creek-Formation von Montana im Denver Museum of Nature and Science. Der unvollständige Dornfortsatz eines der vorderen Schwanzwirbel (siehe Markierung) wird als das Resultat einer erfolgreich abgewehrten Attacke eines großen Theropoden, vermutlich Tyrannosaurus rex, gedeutet.[8][9] (für Nahaufnahmen der mutmaßlichen Bissstelle siehe hier) |

{kind=link}
Edmontosaurus dürfte eine ähnliche Lebensweise gehabt haben, wie sie für andere Hadrosauriden angenommen wird. Die Zahnbatterien weisen große, durch Abnutzung erzeugte Kauflächen auf und stellen eine konvergente Entwicklung zu den flächigen Backenzähnen heutiger pflanzenfressender Säugetiere dar. Sie zeigen, dass die Nahrung durch intensives Kauen zerkleinert wurde.C Die Nahrung bestand aus höherwüchsiger Bodenvegetation sowie aus Blättern und Nadeln von Sträuchern und den untersten Bereichen der Baumkronen.[10] Alle Funde von Edmontosaurus entstammen Sedimenten, die ihrer speziellen Ausbildung (Fazies) zufolge in stark bewaldeten, relativ meeresnahen Bereichen von Küstentiefländern oder in Flussdeltas abgelagert wurden.D Es ist nicht auszuschließen, dass dies auf eine eng an Gewässer gebundene Lebensweise hindeutet.[10]
Wie für die meisten Ornithopoden wird für Edmontosaurus eine überwiegende Fortbewegung auf vier Beinen (Quadrupedie) angenommen. Zweibeinige Fortbewegung (Bipedie) erfolgte wahrscheinlich nur im Fluchtfall. Als Hauptfressfeinde von Edmontosaurus gelten große Tyrannosauriden: Albertosaurus für die Vertreter im späten Campan und frühen Maastricht, Tyrannosaurus für die Vertreter im späten Maastricht.[11]
Systematik
Äußere Systematik
Mit seinem kamm- und hornlosen Schädel, dem sehr breiten Schnabel und der großen äußeren Nasenöffnung ist Edmontosaurus ein typischer Vertreter der Hadrosaurinae, einer Unterfamilie, die traditionell innerhalb der Familie Hadrosauridae ausgehalten und den kamm- oder horntragenden Lambeosaurinae gegenübergestellt wird.E
Taxonomische Geschichte
_(20333747874).jpg.webp)



Zwar fällt die Benennung der Gattung Edmontosaurus durch Edward Lambe im Jahre 1917 noch in eine relativ frühe Phase der nordamerikanischen Dinosaurierforschung, jedoch wurden Vertreter dieser Gattung – nach heutigem Konzept – schon im späten 19. Jahrhundert entdeckt und unter anderen Namen beschrieben oder erwähnt. Der früheste dokumentierte Fund mit taxonomischer Relevanz ist ein Skelett aus den „Laramie-Schichten von Dakota“ (heute Hell-Creek-Formation[4]), das von Edward Drinker Cope im Jahr 1883F unter dem Namen Diclonius mirabilis ausführlich beschrieben wird. Er weist dieses Material eigentlich der bereits 1856 von Joseph Leidy beschriebenen Art Trachodon mirabilis zu, nutzt aber den von ihm 1876 geprägten Gattungsnamen DicloniusG unter der Annahme, Leidy hätte den Namen Trachodon in der Zwischenzeit verworfen.[12] Im Jahr 1892 publiziert Othniel Marsh die Beschreibung eines Hadrosauriden aus den „Laramie-Schichten“ von Wyoming (heute Lance-Formation[13]), den er Claosaurus annectens nennt.[14] Die Gattung Claosaurus war von ihm selbst zwei Jahre zuvor anhand eines Skelettes aus den „Pteranodon-Schichten“ (heute Smoky-Hill-Subformation, Ober-Coniac bis Unter-Campan[15]) der Niobrara-Formation von Kansas errichtet worden, das er 1872 unter dem Namen „Hadrosaurus agilis“ beschrieben hatte.[16] In einer Revision der bis dahin bekannten „Trachodontiden“ (d. h. Hadrosauriden) von Nordamerika stellt John Bell Hatcher 1901 aufgrund der relativ großen geographischen und stratigraphischen Distanz zwischen den „Pteranodon-Schichten“ von Kansas und den „Laramie-Schichten“ von Wyoming und South Dakota die Art Claosaurus annectens in Leidys Gattung Trachodon. Ebenso verfährt er mit Copes „Diclonius mirabilis“.[16] Im Jahr 1913, vier Jahre vor der Erstbeschreibung von Edmontosaurus regalis, veröffentlicht Lambe einen Aufsatz über die Vorderextremität eines Hadrosauriden aus der gleichen Gegend, in der auch das Typusmaterial von E. regalis aufgesammelt wurde. Ein Vergleich der Zähne veranlasst Lambe – unter Vorbehalt – diesen Fund der von ihm 1902 beschriebenen Art Trachodon marginatus zuzuweisen. Elf Jahre später beschreibt Charles Whitney Gilmore basierend auf demselben Exemplar eine neue Art unter dem Namen Thespesius edmontoni, wobei die Gattung Thespesius bereits 1856 von Leidy aufgestellt wurde. Im Jahre 1926 wird durch Charles M. Sternberg, den Bruder der beiden Entdecker des Typusmaterials von E. regalis, unter dem Namen Thespesius saskatchewanensis aus der „Lance-Formation“ (heute Frenchman-Formation[10]) von Saskatchewan das jüngste Typusmaterial eines „Edmontosauriers“ beschrieben.
In einer umfassenden Revision der nordamerikanischen Hadrosauriden, veröffentlicht im Jahre 1942 von Richard Swann Lull und Nelda E. Wright, werden fünf Arten der Hadrosaurinae in die eigens errichtete Gattung Anatosaurus gestellt, darunter Claosaurus annectens als Typus sowie Thespesius edmontoni und Thespesius saskatchewanensis. Auch Copes „Diclonius mirabilis“ ordnen sie Anatosaurus zu. Allerdings konstatieren sie, gestützt auf die relativ große stratigraphische Distanz zwischen Leidys originalen Fundstücken von Trachodon mirabilis („Judith River formation“) und dem von Cope aus den „Laramie-Schichten“ beschriebenen Material, dass es sich bei den beiden nicht um die gleiche Art handeln kann. Folglich gründen sie auf Copes „Laramie“-Material die neue Art Anatosaurus copei. Edmontosaurus bleibt vorerst monotypisch.[17]
Nachdem Lull und Wright (1942) bereits eine relativ starke Ähnlichkeit zwischen den Anatosaurus-Arten und Edmontosaurus regalis feststellten, geht der sowjetische Paläontologe Anatoli Konstantinowitsch Roshdestwenski im Jahr 1968 in einer Übersichtsarbeit zu kasachischen Hadrosauriden einen Schritt weiter und erklärt Anatosaurus zu einem jüngeren Synonym von Edmontosaurus.[18]
In einer Revision der Hadrosauridae stimmt Michael Brett-Surman im Jahr 1989 mit Roshdestwenskis Synonymisierung überein und reduziert zudem die Anzahl der Edmontosaurus-Arten auf eine: E. annectens. Er unterscheidet lediglich zwei unterschiedlich große Morphotypen (regalis und annectens, mit regalis als dem größeren), die seiner Ansicht nach Ausdruck eines Sexualdimorphismus sein könnten.[19] Für „Anatosaurus“ copei errichtet er jedoch die neue Gattung Anatotitan, die er als „den Gipfel der Evolution der Hadrosaurinae“H bezeichnet. Ferner werden in dieser Arbeit die Gattungen Thespesius und Trachodon zu Nomina dubia erklärt,[19] eine Ansicht, die heute allgemein akzeptiert ist.
In den Kapiteln zu den Hadrosauriden der beiden Auflagen des Kompendiums The Dinosauria wird die Synonymie zwischen Anatosaurus und Edmontosaurus bestätigt. Allerdings werden drei Edmontosaurus-Arten unterschieden: Edmontosaurus regalis, E. annectens (einschl. „Thespesius edmontoni“) und E. saskatchewanensis. Hierbei wird in der ersten Auflage (1990) Anatotitan copei als eigenständiges Taxon geführt,[10] in der zweiten Auflage (2004) jedoch als Synonym von E. annectens.[20]
Im Jahr 2011 stellen Nicolás Campione und David Evans schließlich im Ergebnis einer morphometrischen Analyse fest, dass sich in der späten Oberkreide Nordamerikas doch nur zwei Edmontosaurus-Arten unterscheiden lassen: Edmontosaurus regalis und E. annectens. „Anatotitan copei“ liege demnach innerhalb des Variationsspektrums von E. annectens und repräsentiere die ältesten und größten Individuen dieser Spezies.[4] Außerdem liege „Thespesius edmontoni“ innerhalb des Variationsspektrums von E. regalis und sei somit kein Synonym von E. annectens, wie von anderen Bearbeitern[10][20] postuliert.
Überreste eines Hadrosauriden aus dem Liscomb-Bonebed (spätes Campan/frühes Maastricht) der Prince-Creek-Formation im Norden Alaskas, die zunächst als Edmontosaurus sp. klassifiziert worden waren,[4][21][22] sind 2015 als die neue Gattung und Art Ugrunaaluk kuukpikensis beschrieben worden,[23] wobei andere Autoren an der ursprünglichen Klassifikation festhalten.[24]
Anerkannte Arten
Der jüngsten Revision[4] zufolge enthält die Gattung Edmontosaurus zwei Arten:
- Edmontosaurus regalis Lambe, 1917 (Generotypus), spätes CampanA von Alberta
- syn. Thespesius edmontoni Gilmore, 1924
- syn. Anatosaurus edmontoni (Gilmore, 1924)
- syn. Anatosaurus edmontonensis (Gilmore, 1924)
- Edmontosaurus annectens (Marsh, 1892), Maastricht von Montana, North Dakota, South Dakota, Wyoming, Alberta und Saskatchewan
- syn. Claosaurus annectens Marsh, 1892
- syn. Trachodon annectens (Marsh, 1892)
- syn. Anatosaurus annectens (Marsh, 1892)
- syn. Thespesius saskatchewanensis C. M. Sternberg, 1926
- syn. Anatosaurus saskatchewanensis (C. M. Sternberg, 1926)
- syn. Edmontosaurus saskatchewanensis (C. M. Sternberg, 1926)
- syn. „Diclonius mirabilis“ sensu Cope, 1883
- syn. Anatosaurus copei Lull & Wright, 1942
- syn. Anatotitan copei (Lull & Wright, 1942)
E. regalis ist hierbei speziell charakterisiert durch eine besonders breite, „geschwollen“ aussehende Prämaxillarlippe (vgl. Merkmale der Gattung) und eine nach unten (ventrad) ausgedehnte vordere Partie des Nasale. Des Weiteren ist die vordere Schnauzenpartie, zwischen Schnauzenspitze und vorderem Ende der Nasenöffnung kürzer als bei E. annectens. Auch ist, anders als bei E. annectens, die hintere obere (caudodorsale) Partie der circumnarialen Fossa besonders ausgeprägt und ragt nach oben (dorsad) über den Schnauzenrücken hinaus. Eine weitere Besonderheit von E. regalis ist die stärker ausgeprägte „Postorbital Pocket“, mit einer horizontalen, gesimsartigen Kontaktfläche des dorsalen Astes des Jugale für den ventralen Ast des Postorbitale.[4]
Fossilmumien

Edmontosaurus gehört zu den wenigen Dinosauriern, von denen nicht ausschließlich Knochen bekannt sind, sondern auch in größerem Umfang fossilisierte Weichteile, bis hin zu mehr oder weniger vollständig und dreidimensional erhaltenen Individuen. Letztgenannte Form der Erhaltung wird landläufig als „Mumie“ bezeichnet, jedoch handelt es sich im Gegensatz zu echten Mumien nicht wirklich um erhaltenes organisches Material. Bei einem Teil der „Mumien“ ist das Weichgewebe durch Sediment ersetzt worden. Hierbei bildete zunächst das Sediment, das den verschütteten toten Dinosaurier umgab, vorzugsweise ein feinkörniger Sand, die Hautoberfläche als Negativabdruck ab. Durch die Stoffwechselaktivität zersetzender Bakterien bzw. einer daraus resultierenden Mineralisation wurde dieses Sediment im unmittelbaren Umfeld des verwesenden Kadavers stabilisiert. In dem derartig verfestigten Sand konnte der zunehmend zerfallende Kadaver einen Hohlraum hinterlassen. Anschließend drang in diesen Hohlraum lockerer Sand ein und füllte ihn aus, einschließlich der Hautabdrücke in seiner Wandung.[25] Bei anderen „Mumien“ ist offenbar das Weichgewebe selbst bis hinunter auf Zellebene mineralisiert worden.[26]
Bereits das Typusexemplar von „Diclonius mirabilis“ soll großflächig „Hautabdrücke“ gezeigt haben, die jedoch bei den Grabungsarbeiten zerstört wurden.[7] Eine relativ vollständige „Mumie“ von Edmontosaurus wurde erstmals im Jahre 1908 aus der Lance-Formation in Wyoming geborgen. Henry Fairfield Osborn veröffentlichte 1912 eine detaillierte Beschreibung dieser sogenannten Trachodon-Mumie.[27] Schon 1910 wurde eine zweite „Mumie“ in Wyoming ausgegraben[7] und anschließend von der Senckenbergischen Naturforschenden Gesellschaft erworben. Sie ist noch heute Teil der Ausstellung ihres Museums in Frankfurt am Main (siehe → Edmontosaurus-Mumie im Senckenberg Naturmuseum).
Im Jahr 1999 wurde in der Hell-Creek-Formation in den Badlands im Südwesten von North Dakota eine beinahe vollständig erhaltene „Mumie“ entdeckt, die wahrscheinlich zu einem Vertreter der Gattung Edmontosaurus gehört (in der ersten wissenschaftlichen Publikation klassifiziert als cf. Edmontosaurus sp.[26]), und auf den Namen „Dakota“ getauft.[28] Es handelt sich um einen der am vollständigsten erhaltenen Dinosaurierfunde überhaupt. Die Analyse des einzigartigen Fundes mittels Computertomographie führte zu vorläufigen neuen Erkenntnissen bezüglich der Muskelmasse und der Längenberechnung von Dinosauriern im Allgemeinen und gab Anlass zu neuen Hypothesen über Hautbeschaffenheit und Färbung.[29]
Obwohl sie relativ unvollständig ist und nur aus einem unvollständigen Kopf sowie einer Hals- und einer kleinen Rückenpartie besteht, sorgte eine Mumie aus der Wapiti-Formation (spätes Campan/frühes Maastricht) von Alberta, die der Art Edmontosaurus regalis zugeordnet wird, für einiges Aufsehen: Das hintere obere Schädeldach weist eine kuppelartige Struktur auf, die als Überrest eines fleischigen Scheitelkamms, in etwa vergleichbar mit einem Hahnenkamm, gedeutet wird. Es ist der erste derartige Nachweis für einen Nicht-Vogel-Dinosaurier. Zwar sind Kopfanhänge bei Hadrosauriden seit Langem bekannt, insbesondere und in beachtlicher Größe bei den Vertretern der Lambeosaurinae, jedoch sind dies alles knöcherne Schädelauswüchse.[5]
Anmerkungen
- A Hinsichtlich der stratigraphischen Stellung der ältesten Funde oder vielmehr ihrer Fundhorizonte, speziell der unteren Horseshoe-Canyon-Formation, finden sich in der Literatur abweichende Angaben. In jüngeren Arbeiten wird die untere Horseshoe-Canyon-Formation oft in das jüngste Campan gestellt, wobei die Bearbeiter die Campan-Maastricht-Grenze bei 70,6 mya ziehen.[30][31] Entsprechend fielen die geologisch ältesten Funde von Edmontosaurus ebenfalls ins Campan. Nach der aktuellen internationalen Zeitskala liegt die Grenze allerdings bei 72,1 mya[32] und damit die Horseshoe-Canyon-Formation fast vollständig im Maastricht.
- D Beispielsweise enthält die Horseshoe-Canyon-Formation, d. h. die Schichten, aus denen das Typusmaterial von Edmontosaurus regalis stammt, zahlreiche Kohleflöze.[30]
- F Tatsächlich benennt Cope bereits in den Jahren 1871 und 1874 die Arten Trachodon atavus aus New Jersey bzw. Agathaumas milo aus Colorado, die von Weishampel & Horner (1990)[10] und Horner et al. (2004)[20] als Synonyme von Edmontosaurus regalis geführt werden, jedoch handelt es sich dabei um sehr unvollständige, kaum bekannte Stücke.
- G Typus der Gattung ist „Diclonius pentagonus“, eines von Copes „berühmten“ Formtaxa, die auf isolierten, schlecht erhaltenen und damit faktisch undiagnostischen Zähnen fußen.[19]
Einzelnachweise
- Peter Dodson: American Dinosaurs. In: Philip J. Currie, Kevin Padian: Encyclopedia of Dinosaurs. Academic Press, San Diego u. a. 1997, ISBN 0-12-226810-5, S. 10–13.
- Lawrence M. Lambe: A new genus and species of crestless hadrosaur from the Edmonton Formation of Alberta. The Ottawa Naturalist. Bd. 31, Nr. 7, 1917, S. 65–73 (archive.org).
- Lawrence M. Witmer: The Evolution of the Antorbital Cavity of Archosaurs: A Study in Soft-Tissue Reconstruction in the Fossil Record with an Analysis of the Function of Pneumaticity. Society of Vertebrate Paleontology Memoir. Bd. 3 (Journal of Vertebrate Paleontology, Bd. 17, Supplementum Nr. 1), 1997, doi:10.1080/02724634.1997.10011027, S. 61 ff.
- Nicolás E. Campione, David C. Evans: Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae): Implications for Latest Cretaceous Megaherbivore Diversity in North America. PLoS ONE. Bd. 6, Nr. 9, 2011, e25186, doi:10.1371/journal.pone.0025186.
- Phil R. Bell, Federico Fanti, Philip J. Currie, Victoria M. Arbour: A Mummified Duck-Billed Dinosaur with a Soft-Tissue Cock’s Comb. Current Biology. Bd. 24, Nr. 1, 2014, S. 70–75, doi:10.1016/j.cub.2013.11.008 (alternativer Volltextzugriff: ResearchGate).
- John R. Horner: A “segmented” epidermal tail frill in a species of hadrosaurian dinosaur. Journal of Paleontology. Bd. 58, Nr. 1, 1984, S. 270–271 (JSTOR 1304751, alternativer Volltextzugriff: ResearchGate).
- Phil R. Bell: A review of hadrosaur skin impressions. In: David A. Eberth, David C. Evans: Hadrosaurs. Indiana University Press, Bloomington IN 2014, S. 572–590, ISBN 978-0-253-01385-9
- Kenneth Carpenter: Evidence of predatory behavior by carnivorous dinosaurs. In: B. P. Perez-Moreno, T. J. Holtz, J. L. Sanz, J. Mortalla (Hrsg.): Aspects of Theropod Paleobiology. GAIA – Revista de Geosciências. Bd. 15, 1998, S. 135–144.
- Robert A. DePalma, David A. Burnham, Larry D. Martin, Bruce M. Rothschild, Peter L. Larson: Physical evidence of predatory behavior in Tyrannosaurus rex. Proceedings of the National Academy of Science of the United States of America. Bd. 110, Nr. 31, 2013, S. 12560–12564, doi:10.1073/pnas.1216534110, PMC 3732924 (freier Volltext).
- David B. Weishampel, John R. Horner: Hadrosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. University of California Press, Berkeley CA u. a. 1990, ISBN 0-520-06726-6, S. 534–561.
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs. Princeton University Press, Princeton NJ 2010, ISBN 978-0-691-13720-9, S. 297 f.
- Edward Drinker Cope: On the Characters of the Skull in the Hadrosauridæ. Proceedings of The Academy of Natural Sciences of Philadelphia. Bd. 35, 1883, S. 97–107 (BHL).
- Brent H. Breithaupt: Lance Formation. In: Philip J. Currie, Kevin Padian: Encyclopedia of Dinosaurs. Academic Press, San Diego u. a. 1997, S. 394–395.
- Othniel Charles Marsh: Notice of new reptiles from the Laramie formation. American Journal of Science. Bd. 43, 1892, S. 449–453 (archive.org).
- Donald E. Hattin: Stratigraphy and Depositional Environment of Smoky Hill Chalk Member, Niobrara Chalk (Upper Cretaceous) of the Type Area, Western Kansas. Kansas Geological Survey Bulletin. Bd. 225, 1982 (online).
- John Bell Hatcher: The genera and species of the Trachodontidæ. Annals of the Carnegie Museum. Bd. 1, 1901, S. 377–386 (BHL).
- Richard S. Lull, Nelda E. Wright: Hadrosaurian Dinosaurs of North America. Geological Society of America Special Papers. Bd. 40, 1942, doi:10.1130/SPE40-p1.
- Anatoli K. Roshdestwenski: Die Hadrosaurier Kasachstans [Гадрозавры Казахстана]. In: Leonid P. Tatarinow (Hrsg.): Jungpaläozoische und mesozoische Amphibien und Reptilien der UdSSR [Верхнепалеозойские и мезозойские земноводные и пресмыкающиеся СССР]. Moskau, 1968, S. 97–144 (russisch).
- Michael Keith Brett-Surman: A revision of the Hadrosauridae (Reptilia: Ornithischia) and their evolution during the Campanian and Maastrichtian. PhD-Thesis, George Washington University, Graduate School of Arts and Sciences, Washington, D.C. 1989 (online), S. 75–96.
- John R. Horner, David B. Weishampel, Catherine A. Forster: Hadrosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. Zweite Auflage. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 438–463.
- Roland A. Gangloff, Anthony R. Fiorillo: Taphonomy and paleoecology of a bonebed from the Prince Creek Formation, North Slope, Alaska. PALAIOS. Bd. 25, Nr. 5, 2010, S. 299–317, doi:10.2110/palo.2009.p09-103r.
- Hirotsugu Mori, Patrick Druckenmiller, Gregory Erickson, Albert Prieto-Márquez: Cranial ontogeny of Edmontosaurus: implications for the taxonomic status of the Prince Creek Formation species (lower Maastrichtian, northern Alaska). Journal of Vertebrate Paleontology, 73rd annual meeting of the Society of Vertebrate Paleontology, Los Angeles, October 30 – November 2 2013, Program and Abstracts, S. 180 (PDF 9,3 MB, kompletter Abstract-Band; Poster als PDF bei ResearchGate).
- Hirotsugu Mori, Patrick S. Druckenmiller, Gregory M. Erickson: A new Arctic hadrosaurid from the Prince Creek Formation (lower Maastrichtian) of northern Alaska. Acta Palaeontologica Polonica (im Druck), 2015, doi:10.4202/app.00152.2015.
- Hai Xing, Jordan C. Mallon, Margaret L. Currie: Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae. PLoS ONE. Bd. 12, Nr. 4, 2017, e0175253, doi:10.1371/journal.pone.0175253, S. 30 f.
- Kenneth Carpenter: How to Make a Fossil: Part 2 - Dinosaur Mummies and Other Soft Tissue. Journal of Paleontological Sciences. Bd. 1, 2007, JPS.C.07.0002 (PDF 1,9 MB)
- Phillip L. Manning, Peter M. Morris, Adam McMahon, Emrys Jones, Andy Gize, Joe H. S. Macquaker, George Wolff, Anu Thompson, Jim Marshall, Kevin G. Taylor, Tyler Lyson, Simon Gaskell, Onrapak Reamtong, William I. Sellers, Bart E. van Dongen, Mike Buckley, Roy A. Wogelius: Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA). Proceedings of the Royal Society B. Bd. 276, 2009, S. 3429–3437, doi:10.1098/rspb.2009.0812, PMC 2817188 (freier Volltext).
- Henry Fairfield Osborn: Integument of the iguanodont dinosaur Trachodon. Memoirs of the American Museum of Natural History, New Series. Bd. 1, Nr. 2, 1912, S. 30–54 (online)
- Blake Nicholson: Workers Uncovering Mummified Dinosaur. National Geographic News, 18. März 2008.
- Dinosaurier so gut erhalten wie Mumie. Spiegel Online, 3. Dezember 2007.
- David A. Eberth: A revised stratigraphic architecture and history for the Horseshoe Canyon Formation (Upper Cretaceous), southern Alberta plains. AAPG International Conference and Exhibition, Calgary, Canada, 12–15 September 2010 (PDF 240 kB).
- David A. Eberth, David C. Evans, Donald B. Brinkman, François Therrien, Darren H. Tanke, Loris S. Russell: Dinosaur biostratigraphy of the Edmonton Group (Upper Cretaceous), Alberta, Canada: evidence for climate influence. Canadian Journal of Earth Sciences. Bd. 50, Nr. 7, 2013, S. 701–726, doi:10.1139/cjes-2012-0185.
- International Commission on Stratigraphy: International Chronostratigraphic Chart (v2015/01).
- Hadrosaur chewing mechanism visualization using 3D animation. Video auf dem YouTube-Kanal von Alex Tirabasso, veröffentlicht am 20. Januar 2012, ursprünglich Bestandteil einer peer-reviewten Online-Publikation, dort aber anscheinend in einem zu alten Format vorliegend, vgl. Natalia Rybczynski, Alex Tirabasso, Paul Bloskie, Robin Cuthbertson, Casey Holliday: A Three-Dimensional Animation Model of Edmontosaurus (Hadrosauridae) for Testing Chewing Hypotheses. Palaeontologia Electronica. Bd. 11, Nr. 2, 2008, Artikel-Nr. 11.2.9A. (online).
- Albert Prieto-Márquez: Phylogeny and Historical Biogeography of Hadrosaurid Dinosaurs. PhD-Thesis, Florida State University, College of Arts and Science, Tallahassee FL 2008 (online), S. 6–42.
{kind=link}
Weblinks
- Edmontosaurus in der Paleobiology Database