Tyrannosauroidea
Die Tyrannosauroidea ist eine Überfamilie der Coelurosaurier (Coelurosauria) innerhalb der theropoden Dinosaurier. Sie umfasst die Tyrannosauridae, zu denen etwa der bekannte Tyrannosaurus rex gehört, sowie die basalen, also die am Beginn der Entwicklungsreihe stehenden Arten. Tyrannosauroiden lebten auf dem nördlichen Superkontinent Laurasia und tauchten erstmals im Mitteljura vor etwa 168 Millionen Jahren auf. In der Oberkreide waren sie die dominierenden großen Prädatoren der Nordhalbkugel. Ihre Fossilien wurden in Nordamerika, Europa und Asien entdeckt.
| Tyrannosauroidea | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Albertosaurus im Royal Tyrrell Museum in Alberta | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mitteljura bis Oberkreide (Bathonium bis Maastrichtium)[1] | ||||||||||||
| 168,3 bis 66 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Tyrannosauroidea | ||||||||||||
| Brown, 1914 | ||||||||||||
Tyrannosauroiden waren wie die meisten Theropoden bipede Fleischfresser. Die Gruppe ist durch zahlreiche gemeinsam abgeleitete Merkmale (Synapomorphien) charakterisiert, die sich besonders in den Schädel- und Beckenknochen finden. Früh in ihrer Evolution waren Tyrannosauroiden kleine Prädatoren mit langen, dreifingrigen Armen. Arten der Oberkreide wurden wesentlich größer und schlossen einige der größten landlebenden Fleischfresser mit ein, die jemals existiert haben – die meisten dieser späten Arten hatten jedoch proportional kleine Arme mit nur zwei Fingern. Primitive Federn sind von Dilong bekannt, einem frühen Tyrannosauroiden aus China, sowie von Yutyrannus und waren bei anderen Tyrannosauroiden vielleicht ebenfalls vorhanden. Viele Arten trugen auffällige Knochenkämme in verschiedenen Formen, die vielleicht der Zurschaustellung dienten.
Merkmale
Die Körpergröße variierte zwischen einzelnen Gattungen beträchtlich, im Laufe der Evolution wurde die Gruppe jedoch immer größer. Frühe Tyrannosauroiden waren noch relativ kleine Tiere[2] – so maßen ausgewachsene Exemplare von Dilong 1,6 Meter[3] und von Guanlong drei Meter Länge.[4] Gattungen der Kreidezeit wurden größer; so war ein noch nicht geschlechtsreifer Eotyrannus über vier Meter[5] und ein noch nicht ausgewachsener Appalachiosaurus mehr als sechs Meter lang.[2] Tyrannosauriden der Oberkreide wiederum erreichten teils gigantische Ausmaße. Albertosaurus und Gorgosaurus wurden beide etwa neun Meter lang, und Tyrannosaurus war mit über zwölf Metern Länge und vielleicht über 6400 Kilogramm Gewicht die größte bekannte Gattung.[2]

Die Schädel früher Tyrannosauroiden waren lang, niedrig und leicht gebaut und ähneln damit denen anderer Coelurosaurier, während spätere Formen wesentlich höhere und massivere Schädel hatten. Trotz dieser Unterschiede können bestimmte Schädelmerkmale (Synapomorphien) in allen bisher bekannten Tyrannosauroiden gefunden werden: So war das Zwischenkieferbein (Prämaxilla) sehr hoch, was eine stumpfe Schnauze zur Folge hatte – ein Merkmal, das sich konvergent auch bei den Abelisauriden entwickelt hat. Das paarige Nasenbein war verschmolzen, leicht nach oben gewölbt und auf der Oberseite meist sehr rau, oft rindenartig strukturiert. Die Zähne des Zwischenkieferbeins am vorderen Teil des Oberkiefers waren kleiner und anders geformt als die restlichen Zähne und wiesen einen „D“-förmigen Querschnitt auf. Der Unterkiefer aller Tyrannosauroiden außer Guanlong wies einen ausgeprägten Kamm am Surangulare auf, der sich seitlich direkt unter dem Kiefergelenk erstreckte.[2][3][4]
Tyrannosauroiden hatten wie die meisten anderen Theropoden „S“-förmig gebogene Hälse und lange Schwänze. Frühe Gattungen hatten dreifingrige, lange Arme, die bei Guanlong 60 Prozent der Länge der Hinterbeine erreichten.[4] Die langen Arme waren für die Gruppe mindestens bis in die Unterkreide charakteristisch, wie Eotyrannus zeigt,[5] bei Appalachiosaurus sind sie jedoch nicht erhalten geblieben.[6] Die Arme späterer Tyrannosauroiden waren deutlich kürzer – besonders bei Tarbosaurus aus der Mongolei, dessen Oberarmknochen (Humerus) lediglich ein Viertel der Länge des Oberschenkelknochens (Femur) erreichte.[2] Der dritte Finger hat sich im Laufe der Evolution ebenfalls zurückgebildet: Während er bei dem basalen Guanlong noch nicht reduziert war,[4] war er bereits bei Dilong deutlich kleiner als die anderen beiden Finger.[3] Eotyrannus hatte noch immer drei funktionale Finger an jeder Hand;[5] Tyrannosauriden hatten jedoch nur noch zwei Finger, obwohl Rudimente eines dritten Fingers bei einigen Exemplaren nachgewiesen wurden.[7] Wie bei den meisten anderen Coelurosauriern war der zweite Finger der größte.
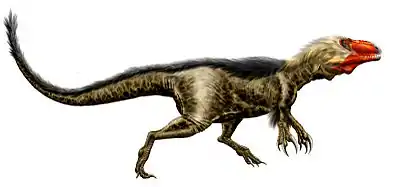
Charakteristische Merkmale in den Beckenknochen schließen eine Kerbe am unteren Ende des Darmbeins (Os ilium), einen deutlich begrenzten vertikalen Kamm auf dem Darmbein, der sich von der Hüftgelenkpfanne (Acetabulum) nach oben erstreckte, und den vergrößerten Ende des Schambeins (Os pubis) mit ein, das „T“-förmig zu beiden Seiten verlängert und mehr als halb so lang war wie der eigentliche Schaft des Schambeins.[2] Diese Merkmale finden sich bei allen bekannten Tyrannosauroiden, einschließlich der basalen Gattungen Guanlong[4] und Dilong.[3] Das Schambein ist in Aviatyrannis und Stokesosaurus unbekannt; jedoch zeigen beide Gattungen für Tyrannosauroiden typische Merkmale im Darmbein.[8] Die Hinterbeine aller Tyrannosauroiden hatten wie bei den meisten Theropoden vier Zehen, obwohl die erste Zehe (der Hallux) nicht den Boden berührte. Die Hinterbeine der Tyrannosauroiden waren im Verhältnis zur Körpergröße länger als bei fast allen anderen Theropoden und zeigen Proportionen, die für schnell laufende Tiere charakteristisch sind; so waren das Schienbein (Os Tibia) und die Mittelfußknochen verlängert.[2] Diese Proportionen bestanden selbst in dem größten bekannten Exemplar von Tyrannosaurus,[9] obwohl diese Tiere vielleicht gar nicht rennen konnten.[10] Die obere Hälfte des dritten (d. h. mittleren) Mittelfußknochens der Tyrannosauriden war zwischen den beiden anderen Mittelfußknochen „eingequetscht“ und hatte nur geringen Anteil an der Kontaktfläche von Mittelfuß und Fußwurzel – eine Struktur, die als Arctometatarsus bekannt ist. Der Arctometatarsus wurde auch bei Appalachiosaurus nachgewiesen,[6] es ist aber nicht klar, ob er auch bei Eotyrannus oder Dryptosaurus vorhanden war.[11] Diese Struktur findet sich bei Ornithomimiden, Troodontiden und Caenagnathiden,[12] war aber bei basalen Tyrannosauroiden wie Dilong nicht vorhanden, was auf konvergente Evolution schließen lässt.[3]
Systematik
Tyrannosaurus wurde zusammen mit der Familie Tyrannosauridae von Henry Fairfield Osborn im Jahr 1905 benannt.[13] Der Name leitet sich aus den altgriechischen Wörtern τύραννος/tyrannos („Tyrann“) und σαῦρος/sauros („Echse“) ab. Der Name der Überfamilie Tyrannosauroidea veröffentlichte erstmals der britische Paläontologe Alick Walker im Jahr 1964.[14] Die Endung -oidea, die für gewöhnlich für Überfamilien im Tierreich verwendet wird, ist aus dem altgriechischen εἶδος/eidos („Form“) abgeleitet.[15]
Wissenschaftler verstanden die Tyrannosauroidea meistens als Taxon, das die Tyrannosauridae und ihre unmittelbaren Vorfahren mit einschließt.[14][16] Seitdem die phylogenetische Systematik in die Paläontologie eingeführt wurde, erhielt die Gruppe jedoch genaue Definitionen. Die erste wurde von Paul Sereno im Jahr 1998 aufgestellt und sieht die Tyrannosauroidea als ein Taxon, das alle Spezies mit einschließt, die näher mit Tyrannosaurus rex verwandt waren als mit neornithen Vögeln.[17] Um die Gruppe noch exklusiver zu beschreiben, definierte Thomas Holtz sie im Jahr 2004 neu, um alle Spezies mit einzuschließen, die näher mit Tyrannosaurus rex als mit Ornithomimus velox, Deinonychus antirrhopus oder Allosaurus fragilis verwandt waren.[2] Sereno veröffentlichte im Jahr 2005 eine neue Definition, nach der alle Spezies der Tyrannosauroidea angehören, die näher mit Tyrannosaurus rex als mit Ornithomimus edmontonicus, Velociraptor mongoliensis und Troodon formosus verwandt waren.[18]





Klassifikation
Überfamilie Tyrannosauroidea
- ?Iliosuchus (Mitteljura, England)
- Aviatyrannis (Oberjura, Portugal)
- Stokesosaurus (Oberjura, westliche Vereinigte Staaten)
- Dilong (Unterkreide, östliches China)
- Eotyrannus (Unterkreide, England)
- Moros (Unterkreide, westliche Vereinigte Staaten)[19]
- Raptorex (Unterkreide, östliches China)
- Xiongguanlong (Unterkreide, China)
- Yutyrannus (Unterkreide, China)
- Suskityrannus (Mittelkreide, Vereinigte Staaten)[20]
- Timurlengia (Mittelkreide, Usbekistan)[21]
- ?Bagaraatan (Oberkreide, Mongolei)
- Dryptosaurus (Oberkreide, östliche Vereinigte Staaten)
- Alectrosaurus (Oberkreide, Mongolei)
- Appalachiosaurus (Oberkreide, östliche Vereinigte Staaten)
- ?Labocania (Oberkreide, westliches Mexiko)
- Bistahieversor (Oberkreide, westliche Vereinigte Staaten)
- Familie Proceratosauridae[22]
- Kileskus (Mitteljura, zentrales Russland)
- Proceratosaurus (Mitteljura, England)
- Guanlong (Oberjura, westliches China)
- Sinotyrannus (Unterkreide, östliches China)
- Familie Tyrannosauridae
- Albertosaurus (Oberkreide, westliche Vereinigte Staaten)
- ?Alioramus (Oberkreide, Mongolei)
- Daspletosaurus (Oberkreide, westliche Vereinigte Staaten)
- Dynamoterror (Oberkreide, New Mexico)[23]
- Gorgosaurus (Oberkreide, westliche Vereinigte Staaten)
- Lythronax (Oberkreide, westliches Nordamerika)[24]
- Qianzhousaurus (Oberkreide, China)[25]
- Tarbosaurus (Oberkreide, Mongolei)
- Teratophoneus (Oberkreide, Vereinigte Staaten)
- Tyrannosaurus (Oberkreide, westliche Vereinigte Staaten)
- Zhuchengtyrannus (Oberkreide, China)
Phylogenetik
Im 20. Jahrhundert galten Tyrannosauriden für gewöhnlich als Vertreter der Carnosauria, welche nach damaligem Gesichtspunkt nahezu alle großen Theropoden mit einschlossen.[26][27] Innerhalb dieser Gruppe wurden die Allosauriden oft für die Vorfahren der Tyrannosauriden gehalten.[16][28] In den frühen 1990er-Jahren begannen kladistische Analysen, die Tyrannosauridae innerhalb der Coelurosauria einzuordnen,[29][12] wobei erste Vermutungen schon in den 1920er-Jahren geäußert wurden.[30][31] Tyrannosauriden werden heute allgemeingültig als große Coelurosaurier betrachtet.[2][4][32][33][34][35][36]
Im Jahr 1994 gruppierte Holtz die Tyrannosauroiden zusammen mit den Elmisauriden, den Ornithomimosauriern und den Troodontiden in eine neue Gruppe innerhalb der Coelurosauria, die er Arctometatarsalia nannte. Diese Gruppe basiert auf dem Arctometatarsus, einer Struktur in den Mittelfußknochen, bei der der zweite Mittelfußknochen von vorne betrachtet von den restlichen beiden Mittelfußknochen teilweise verdeckt wird.[12] Basale Tyrannosauroiden wie Dilong zeigten jedoch kein Arctometatarsus, was darauf schließen lässt, dass sich dieses Merkmal in verschiedenen Gruppen mehrmals unabhängig voneinander (konvergent) entwickelt hat.[3] Daher wurde die Gruppe Arctometatarsalia verworfen und von den meisten Paläontologen nicht mehr verwendet, wobei die Tyrannosauroidea heute meistens als basale Gruppe der Coelurosauria außerhalb der Maniraptoriformes klassifiziert wird.[2][33][35] Eine neue Analyse kam zu dem Schluss, dass die Familie Coeluridae das Schwestertaxon der Tyrannosauroidea gewesen sein könnte.[36]
Der ursprünglichste Tyrannosauroide, der von vollständigem Skelettmaterial bekannt ist, ist Guanlong.[4] Andere frühe Gattungen schließen Stokesosaurus und Aviatyrannis mit ein, sind aber nur von weit weniger vollständigem Material bekannt.[8] Der besser bekannte Dilong ist etwas moderner als Guanlong und Stokesosaurus.[3][4] Dryptosaurus galt lange als schwierig einzuordnen, wird heute jedoch als basaler Tyrannosauroide geführt, der etwas ursprünglicher war als Eotyrannus und Appalachiosaurus.[2][6][37] Alectrosaurus, eine wenig bekannte Gattung aus der Mongolei, ist definitiv ein Tyrannosauroide, seine genauen Verwandtschaftsbeziehungen sind jedoch unklar.[2] Andere Taxa wurden von manchen Autoren als mögliche Tyrannosauroiden vorgeschlagen, einschließlich Bagaraatan, Labocania und eine Gattung, die fälschlicherweise dem Chilantaisaurus zugeordnet wurde, „C.“ maortuensis.[2] Siamotyrannus aus der Unterkreide Thailands wurde ursprünglich als früher Tyrannosauride beschrieben,[38] gilt heute jedoch als Mitglied der Carnosauria.[33][39] Iliosuchus zeigt einen vertikalen Kamm auf dem Darmbein, was ein Merkmal der Tyrannosauroidea ist. Diese Gattung könnte daher tatsächlich der früheste bekannte Vertreter dieser Überfamilie sein, was jedoch nur durch weitere Knochenfunde bestätigt werden kann.[8][39]
Verbreitung
Die frühesten bekannten Tyrannosauroiden lebten im Mitteljura und schließen Guanlong aus dem nordwestlichen China,[4] Stokesosaurus aus den westlichen USA und Aviatyrannis aus Portugal mit ein. Einige Fossilien, die derzeit Stokesosaurus zugeschrieben werden, gehören vielleicht zu Aviatyrannis – die Dinosaurierfaunen von Portugal und Nordamerika waren zu dieser Zeit tatsächlich sehr ähnlich. Falls Iliosuchus aus dem Mitteljura Englands ein Tyrannosauroide war, wäre er die früheste bekannte Gattung; dies könnte darauf hindeuten, dass die Tyrannosauroiden ihren Ursprung in Europa hatten.[8]
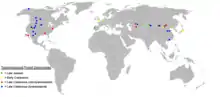
Tyrannosauroiden der Unterkreide fand man ebenfalls in allen drei nördlichen Kontinenten. Eotyrannus aus England[5] und Dilong aus dem nordöstlichen China[3] sind die beiden einzigen benannten Gattungen aus dieser Zeit; des Weiteren sind Zähne des Zwischenkieferbeins aus der Cedar-Mountain-Formation in Utah (Vereinigte Staaten)[40] und aus der Tetori-Gruppe in Japan bekannt.[41] „Chilantaisaurus“ maortuensis aus der Dashuigou-Formation der Inneren Mongolei in China wird manchmal ebenfalls als ein früher Tyrannosauroide der Unterkreide betrachtet.[2]
In Europa sind Tyrannosauroiden seit der Mittelkreide aus der Fossilüberlieferung verschwunden, was ein lokales Aussterben auf diesem Kontinent vermuten lässt.[2] Zähne und mögliche Körperfossilien aus der Mittelkreide sind aus der nordamerikanischen Dakota-Formation,[40] sowie aus Formationen in Kasachstan, Tadschikistan und Usbekistan bekannt.[42] Die ersten unzweifelhaften Überreste von Tyrannosauriden stammen aus dem Campanium (späte Oberkreide) von Nordamerika und Asien. Diese Familie wird in zwei Unterfamilien unterteilt, wobei die Albertosaurinen lediglich in Nordamerika nachgewiesen wurden, die Tyrannosaurinen aber auf beiden Kontinenten entdeckt wurden.[2] Fossilien von Tyrannosauriden wurden auch in Alaska entdeckt, das vielleicht mit einer Landbrücke mit Asien verbunden war, über die sich die Faunen von Nordamerika und Asien austauschen konnten.[43] Die Tyrannosauroiden Alectrosaurus und vielleicht Bagaraatan werden außerhalb der Tyrannosauridae klassifiziert, lebten mit diesen in Asien aber zur gleichen Zeit, während sie in Nordamerika fehlten.[2] Das östliche Nordamerika wurde seit der Mittelkreide durch einen Meerarm, dem Western Interior Seaway, vom westlichen Teil des Kontinents isoliert. Da Tyrannosauriden im östlichen Teil des Kontinents fehlten, wird vermutet, dass diese Familie erst entstand, nachdem der Meerarm das Land geteilt hatte. Dies erlaubte basalen Tyrannosauroiden wie Dryptosaurus und Appalachiosaurus im östlichen Nordamerika bis an das Ende der Oberkreide zu überleben.[6]
Paläobiologie
Federn
Lange faserige Strukturen sind zusammen mit den Skeletten zahlreicher Coelurosaurier erhalten geblieben, die aus der Unterkreide der Yixian-Formation und weiteren Formationen aus Liaoning (China) stammen.[44] Diese Strukturen werden für gewöhnlich als „Protofedern“ interpretiert, homolog zu den verästelten Federn moderner Vögel,[45][46] obwohl auch andere Hypothesen aufgestellt wurden.[47] Ein im Jahr 2004 beschriebenes Skelett von Dilong ist das erste bekannte Beispiel für Protofedern bei Tyrannosauroiden. Ähnlich den Daunen moderner Vögel waren die von Dilong bekannten Protofedern verästelt, es handelte sich jedoch nicht um Konturfedern. Möglicherweise dienten sie der Wärmeisolierung.[3]
Dass Protofedern bei basalen Tyrannosauroiden gefunden wurden, ist nicht überraschend, da Federn als ein charakteristisches Merkmal der Coelurosaurier angesehen werden. Sie finden sich sowohl bei anderen basalen Gattungen wie Sinosauropteryx[45] als auch bei moderneren Gattungen.[44] Seltene Hautabdrücke großer Tyrannosauriden zeigen jedoch keine Hinweise auf Federn, sondern stattdessen eine schuppige Haut.[48] Es ist möglich, das Protofedern an Teilen des Körpers anwesend waren, die nicht durch Hautabdrücke überliefert worden sind. Alternativ könnten die Protofedern bei großen Tyrannosauriden verloren gegangen sein, da das kleine Fläche-Volumen-Verhältnis dieser Tiere eine Wärmeisolierung überflüssig macht – ähnlich wie bei heutigen Großsäugern wie Elefanten.[3] Eine Ausnahme ist hier Yutyrannus, bei dem bis zu 20 Zentimeter lange Filamente gefunden wurden, die als Federn interpretiert werden.[49]
Kopfkämme
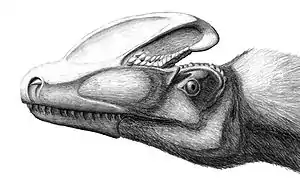
Knochenkämme wurden auf den Schädeln vieler Theropoden gefunden, einschließlich zahlreicher Tyrannosauroiden. Das spektakulärste Beispiel ist Guanlong, bei dem das paarige Nasenbein einen einzigen, großen Kamm unterstützt, der auf der Mittellinie des Schädels von der Schnauze bis zur Hinterseite verlief. Der Kamm war von großen Öffnungen durchzogen, die sein Gewicht reduzierten.[4] Weniger auffällig waren die Kopfornamente von Dilong, der aus zwei niedrigen, parallelen Kämmen bestanden, die auf jeder Seite des Schädels verliefen und von Nasenbein und Tränenbein unterstützt wurden. Die Kämme waren direkt hinter den Nasenlöchern nach innen gebogen, was eine „Y“-förmige Struktur ergab.[3] Das verschmolzene paarige Nasenbein der Tyrannosauriden war oft sehr rau strukturiert. Alioramus, ein möglicher Tyrannosauride aus der Mongolei, zeigte eine einzelne Reihe mit fünf auffälligen knöchernen Beulen auf dem Nasenbein; eine ähnliche Reihe mit wesentlich kleineren Beulen ist von Appalachiosaurus sowie von einigen Funden von Daspletosaurus, Albertosaurus und Tarbosaurus bekannt.[6] Das auf dem Tränenbein sitzende Horn fehlt bei Tarbosaurus und bei Tyrannosaurus, welche stattdessen eine halbmondförmige Haube hinter jedem Auge auf den Postorbitalknochen besaßen.[2] Die Kopfkämme der Tyrannosauroiden dienten wahrscheinlich der Zurschaustellung – vielleicht zur Arterkennung oder zur Balz.[2]
Weblinks
- Tyrannosauroidea (Memento vom 25. Januar 2007 im Internet Archive) in The Theropod Database (englisch)
Einzelnachweise
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs, 2010. ISBN 978-0-691-13720-9, S. 99–110 Online
- Thomas R. Holtz Jr.: Tyrannosauroidea. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 111–136.
- Xing Xu, Mark A. Norell, Xuewen Kuang, Xiaolin Wang, Qi Zhao, Chengkai Jia: Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. In: Nature. Bd. 431, Nr. 7009, 2004, S. 680–684, doi:10.1038/nature02855, PMID 15470426.
- Xing Xu, James M. Clark, Catherine A. Forster, Mark A. Norell, Gregory M. Erickson, David A. Eberth, Chengkai Jia, Qi Zhao: A basal tyrannosauroid dinosaur from the Late Jurassic of China. In: Nature. Bd. 439, 2006, S. 715–718, doi:10.1038/nature04511.
- Stephen Hutt, Darren Naish, David M. Martill, Michael J. Barker, Penny Newberry: A preliminary account of a new tyrannosauroid theropod from the Wessex Formation (Cretaceous) of southern England. In: Cretaceous Research. Bd. 22, Nr. 2, 2001, ISSN 0195-6671, S. 227–242, doi:10.1006/cres.2001.0252.
- Thomas D. Carr, Thomas E. Williamson, David R. Schwimmer: A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama. In: Journal of Vertebrate Paleontology. Bd. 25, Nr. 1, 2005, ISSN 0272-4634, S. 119–143, doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2.
- Elizibeth D. Quinlan, Kraig Derstler, Mercedes M. Miller: Anatomy and function of digit III of the Tyrannosaurus rex manus. In: The Geological Society of America. Abstracts with Programs. Bd. 39, Nr. 6, 2007, ISSN 0016-7592, S. 77, Zusammenfassung.
- Oliver W. M. Rauhut: A tyrannosauroid dinosaur from the Upper Jurassic of Portugal. In: Palaeontology. Bd. 46, Nr. 5, 2003, ISSN 0031-0239, S. 903–910, doi:10.1111/1475-4983.00325.
- Christopher A. Brochu, Richard A. Ketcham: Osteology of Tyrannosaurus rex. Insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull (= Society of Vertebrate Paleontology. Memoir. 7, ISSN 1062-161X = Journal of Vertebrate Paleontology. Bd. 22, Nr. 4, Supplement). Society of Vertebrate Paleontology, Northbrook IL 2002.
- John R. Hutchinson, Mariano Garcia: Tyrannosaurus was not a fast runner. In: Nature. Bd. 415, Nr. 6875, 2002, S. 1018–1021, doi:10.1038/4151018a, Digitalisat (PDF; 279,67 KB).
- Kenneth Carpenter, Dale Russell, Donald Baird, Robert Denton: Redescription of the holotype of Dryptosaurus aquilunguis (Dinosauria: Theropoda) from the Upper Cretaceous of New Jersey. In: Journal of Vertebrate Paleontology. Bd. 17, Nr. 3, 1997, S. 561–573, doi:10.1080/02724634.1997.10011003.
- Thomas R. Holtz Jr.: The phylogenetic position of the Tyrannosauridae: implications for theropod systematics. In: Journal of Paleontology. Bd. 68, Nr. 5, 1994, ISSN 0022-3360, S. 1100–1117, Zusammenfassung.
- Henry Fairfield Osborn: Tyrannosaurus and other Cretaceous carnivorous dinosaurs. In: Bulletin of the American Museum of Natural History. Bd. 21, Article 14, 1905, S. 259–265, Digitalisat.
- Alick D. Walker: Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs. In: Proceedings of the Royal Society of London. Series B: Biological Sciences. Bd. 248, Nr. 744, 1964, ISSN 0080-4649, S. 53–134, doi:10.1098/rstb.1964.0009.
- Henry George Liddell, Robert Scott: A Lexicon abridged from Liddell and Scott's Greek-English Lexicon. 24th edition, carefully revised throughout. Ginn, Boston 1891 (Reprinted edition. Clarendon Press, Oxford 2010, ISBN 978-0-19-910207-5).
- José F. Bonaparte, Fernando E. Novas, Rodolfo A. Coria: Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia (= Contributions in Science. Nr. 416, ISSN 0459-8113). Natural History Museum of Los Angeles County, Los Angeles CA 1990.
- Paul C. Sereno: A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. In: Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen. Bd. 210, Nr. 1, 1998, ISSN 0077-7749, S. 41–83.
- Paul C. Sereno: Stem Archosauria. (Nicht mehr online verfügbar.) In: TaxonSearch Version 1.0. 2005, archiviert vom Original am 26. Dezember 2007; abgerufen am 28. Juli 2014 (englisch). Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis.
- Lindsay E. Zanno, Ryan T. Tucker, Aurore Canoville, Haviv M. Avrahami, Terry A. Gates & Peter J. Makovicky: Diminutive fleet-footed tyrannosauroid narrows the 70-million-year gap in the North American fossil record. Communications Biologyvolume 2, Article number: 64 (2019)
- Sterling J. Nesbitt, Robert K. Denton Jr, Mark A. Loewen, Stephen L. Brusatte, Nathan D. Smith, Alan H. Turner, James I. Kirkland, Andrew T. McDonald & Douglas G. Wolfe (2019). A mid-Cretaceous tyrannosauroid and the origin of North American end-Cretaceous dinosaur assemblages. Nature Ecology & Evolution. doi:
- Stephen L. Brusatte, Alexander Averianov, Hans-Dieter Sues, Amy Muir and Ian B. Butler (2016). New tyrannosaur from the mid-Cretaceous of Uzbekistan clarifies evolution of giant body sizes and advanced senses in tyrant dinosaurs. Proceedings of the National Academy of Sciences. doi: 10.1073/pnas.1600140113
- Oliver W. M. Rauhut, Angela C. Milner, Scott Moore-Fay: Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. In: Zoological Journal of the Linnean Society. Bd. 158, Nr. 1, ISSN 0024-4082, 2009, doi:10.1111/j.1096-3642.2009.00591.x.
- Andrew T. McDonald, Douglas G. Wolfe, Alton C. Dooley Jr.: A New Tyrannosaurid (Dinosauria: Theropoda) from the Upper Cretaceous Menefee Formation of New Mexico. In: PeerJ. Bd. 6, 2018, e5749, DOI: 10.7717/peerj.5749
- Mark A. Loewen, Randall B. Irmis, Joseph J. W. Sertich, Philip J. Currie, Scott D. Sampson: Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous Oceans. In: PLoS ONE. Bd. 8, Nr. 11, 2013, e79420, doi:10.1371/journal.pone.0079420.
- Junchang Lü, Laiping Yi, Stephen L. Brusatte, Ling Yang, Hua Li, Liu Chen: A new clade of Asian Late Cretaceous long-snouted tyrannosaurids. In: Nature Communications. 5, Article Nr. 3788, 2014, ISSN 2041-1723, doi:10.1038/ncomms4788.
- Alfred Sherwood Romer: Osteology of the Reptiles. University of Chicago Press, Chicago IL 1956 (Reprint with new Preface and taxonomic Table. Krieger Publishing, Malabar FL 1997, ISBN 0-89464-985-X).
- Jacques Gauthier: Saurischian monophyly and the origin of birds. In: Kevin Padian (Hrsg.): The Origin of Birds and the Evolution of Flight (= Memoirs of the California Academy of Sciences. Bd. 8). California Academy of Sciences, San Francisco CA 1986, ISBN 0-940228-14-9, S. 1–55.
- Ralph E. Molnar, Seriozha M. Kurzanov, Zhiming Dong: Carnosauria. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. University of California Press, Berkeley CA u. a. 1990, ISBN 0-520-06727-4, S. 169–209.
- Fernando E. Novas: La evolucion de los dinosaurios carnivoros. In: José L. Sanz, Angela D. Buscalioni (Hrsg.): Los Dinosaurios y Su Entorno Biotico. Actas del Segundo Curso de Paleontologia in Cuenca (= Instituto Juan de Valdes. Serie Actas Académicas. Bd. 4, ZDB-ID 1331351-4). Instituto Juan de Valdez, Cuenca 1992, ISBN 84-86788-14-5, S. 125–163.
- William D. Matthew, Barnum Brown: The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta. In: Bulletin of the American Museum of Natural History. Bd. 46, Nr. 6, 1922, ISSN 0003-0090, S. 367–385, (PDF; 1,8 MB).
- Friedrich von Huene: Carnivorous Saurischia in Europe since the Triassic. In: Geological Society of America Bulletin. Bd. 34, Nr. 3, 1923, ISSN 0016-7606, S. 449–458, doi:10.1130/GSAB-34-449.
- Paul C. Sereno: The Evolution of Dinosaurs. In: Science. Bd. 284, Nr. 5423, 1999, S. 2137–2147, doi:10.1126/science.284.5423.2137.
- Oliver W. M. Rauhut: The interrelationships and evolution of basal theropod dinosaurs (= Special Papers in Palaeontology. Bd. 69). The Palaeontological Association, London 2003, ISBN 0-901702-79-X.
- Philip J. Currie, Jørn H. Hurum, Karol Sabath: Skull structure and evolution in tyrannosaurid dinosaurs. In: Acta Palaeontologica Polonica. Bd. 48, Nr. 2, 2003, ISSN 0567-7920, S. 227–234, (PDF; 137 kB).
- Mark A. Norell, James M. Clark, Alan H. Turner, Peter J. Makovicky, Rinchen Barsbold, Timothy Rowe: A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia). (= American Museum Novitates. Nr. 3545, ISSN 0003-0082). American Museum of Natural History, New York NY 2006, (PDF; 10,2 MB).
- Phil Senter: A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda). In: Journal of Systematic Palaeontology. Bd. 5, Nr. 4, 2007, ISSN 1477-2019, S. 429–463, doi:10.1017/S1477201907002143.
- Thomas R. Holtz Jr.: RE: Burpee Conference (LONG). In: Archives of the Dinosaur Mailing List. 20. September 2005, abgerufen am 29. Juli 2014.
- Eric Buffetaut, Varavudh Suteethorn, Haiyan Tong: The earliest known tyrannosaur from the Lower Cretaceous of Thailand. In: Nature. Bd. 381, Nr. 6584, 1996, S. 689–691, doi:10.1038/381689a0.
- Thomas R. Holtz Jr., Ralph E. Molnar, Philip J. Currie: Basal Tetanurae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2. Ausgabe. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 71–110.
- James I. Kirkland, Brooks Britt, Donald L. Burge, Kenneth Carpenter, Richard Cifelli, Frank DeCourten, Jeffrey Eaton, Steve Hasiotis, Tim Lawton: Lower to Middle Cretaceous Dinosaur faunas of the central Colorado Plateau: a key to understanding 35 million years of tectonics, sedimentology, evolution, and biogeography. In: Brigham Young University, Department of Geology. Geology Studies. Bd. 42, Nr. 2, 1997, ISSN 0068-1016, S. 69–103.
- Makoto Manabe: The early evolution of the Tyrannosauridae in Asia. In: Journal of Paleontology. Bd. 73, Nr. 6, 1999, S. 1176–1178, Zusammenfassung.
- Лев А. Несов: Динозавры Северной Евразии. новые данные о составе комплексов, экологии и палеобиогеографии. Санкт-Петербургский государственный университет, Санкт-Петербург 1995, Digitalisat, (In englischer Sprache: Dinosaurs of Northern Eurasia: new data about assemblages, ecology and paleobiogeography. online (PDF; 1,09 MB)).
- Anthony R. Fiorillo, Roland A. Gangloff: Theropod teeth from the Prince Creek Formation (Cretaceous) of northern Alaska, with speculations on Arctic dinosaur paleoecology. In: Journal of Vertebrate Paleontology. Bd. 20, Nr. 4, 2000, S. 675–682, doi:10.1671/0272-4634(2000)020[0675:TTFTPC]2.0.CO;2.
- Zhonghe Zhou, Paul M. Barrett, Jason Hilton: An exceptionally preserved Lower Cretaceous ecosystem. In: Nature. Bd. 421, Nr. 6925, 2003, S. 807–814, doi:10.1038/nature01420.
- Chen Peiji, Dong Zhiming, Zhen Shuonan: An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. In: Nature. Bd. 391, Nr. 6663, 1998, S. 147–152, doi:10.1038/34356.
- Xu Xing, Zhou Zhonghe, Richard A. Prum: Branched integumental structures in Sinornithosaurus and the origin of feathers. In: Nature. Bd. 410, Nr. 6825, 2003, S. 200–204, doi:10.1038/35065589.
- Theagarten Lingham-Soliar, Alan Feduccia, Xiaolin Wang: A new Chinese specimen indicates that „protofeathers“ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres. In: Proceedings of the Royal Society of London. Series B: Biological Sciences. Bd. 274, Nr. 1620, 2007, S. 1823–1829, doi:10.1098/rspb.2007.0352.
- Larry D. Martin, Stephan A. Czerkas: The fossil record of feather evolution in the Mesozoic. In: American Zoologist. Bd. 40, Nr. 4, 2000, S. 687–694, doi:10.1668/0003-1569(2000)040[0687:TFROFE]2.0.CO;2.
- Xing Xu, Kebai Wang, Ke Zhang, Qingyu Ma, Lida Xing, Corwin Sullivan, Dongyu Hu, Shuqing Cheng, Shuo Wang: A gigantic feathered dinosaur from the Lower Cretaceous of China. Supplementary Information. In: Nature. Bd. 484, Nr. 7392, 2012, S. 92–95, doi:10.1038/nature10906, (PDF; 961 kB).