Daspletosaurus
Daspletosaurus (griech.: „Furchtbare Echse“, von daspleto-/δασπλητο- „furchtbar“ und saurus/σαῦρος „Echse“) war ein theropoder Dinosaurier aus der Familie der Tyrannosauridae, der vor ungefähr 80 bis 72 Millionen Jahren während der Oberkreide (Campanium) im westlichen Nordamerika lebte. Die einzige bisher benannte Art (Typusart), Daspletosaurus torosus, wurde in Alberta (Kanada) entdeckt; weitere mögliche Arten aus Alberta, New Mexico und Montana warten noch auf eine Beschreibung. Einschließlich dieser noch unbenannten Arten wäre Daspletosaurus der artenreichste Tyrannosaurier.
| Daspletosaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Lebendbild von Daspletosaurus torosus | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (Mittleres bis Oberes Campanium)[1] | ||||||||||||
| 80,6 bis 72 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Daspletosaurus | ||||||||||||
| Russell, 1970 | ||||||||||||
| Art | ||||||||||||
|
Daspletosaurus war eng mit dem wesentlich größeren und geologisch jüngeren Tyrannosaurus verwandt. Wie alle Tyrannosauriden war er ein bipeder Fleischfresser mit großem Schädel und kleinen Armen, obwohl die Arme proportional länger waren als bei anderen Tyrannosauriern.
Als ein Spitzenprädator stand Daspletosaurus an der Spitze der Nahrungskette und jagte möglicherweise große Dinosaurier wie den Ceratopsia Centrosaurus und den Hadrosauriden Hypacrosaurus. In einigen Gebieten lebte er zusammen mit Gorgosaurus, einem anderen Tyrannosauriden, wobei es Hinweise darauf gibt, dass beide Gattungen unterschiedliche ökologische Nischen besetzten. Obwohl Fossilien von Daspletosaurus seltener sind als von anderen Tyrannosauriden, erlauben die Funde Rückschlüsse auf die Biologie dieser Tiere, einschließlich Sozialverhalten, Ernährung und Individualentwicklung (Ontogenese).
Merkmale
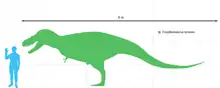
Daspletosaurus war ein mittelgroßer Tyrannosaurier, erwachsene Tiere erreichten eine Länge von acht bis neun Metern.[2] Gewichtsschätzungen reichen von 1,8 Tonnen[3] bis 3,8 Tonnen,[4] die meisten Schätzungen liegen jedoch um die 2,5 Tonnen.[2][5][6] Wie seine Verwandten hatte dieser Tyrannosauride einen kurzen, „S“-förmig gebogenen Hals und sehr kleine Arme mit nur zwei Fingern, die bei Daspletosaurus jedoch im Verhältnis länger waren als bei jedem anderen Tyrannosauriden.[2][1]
Der Schädel war massiv und konnte eine Länge von über einem Meter erreichen.[2] Die Knochen waren insgesamt schwer gebaut und einige, wie das paarige Nasenbein (Nasale), waren zur Stabilisierung miteinander verschmolzen. Große Schädelöffnungen reduzierten das Gewicht des Schädels. Daspletosaurus war mit etwa vier Dutzend sehr langen Zähnen ausgestattet, die im Querschnitt meist oval waren. Im Gegensatz dazu waren die Zähne des Zwischenkieferbeins (Prämaxillare) am vorderen Ende des Oberkiefers im Querschnitt „D“-förmig – ein Beispiel der Heterodontie, die sich bei jedem Tyrannosauriden findet. Einzigartige Schädelmerkmale schließen die raue Außenfläche des Oberkiefers (Maxilla) und die Ornamente mit ein, die um das Orbitalfenster (Augenhöhle) auf dem Tränenbein (Lacrimale), der Postorbitale und der Jugale verlaufen. Das Orbitalfenster war ein großes Oval und ein Zwischenstück zwischen der rundlichen Form bei Gorgosaurus und der „Schlüsselloch“-Form bei Tyrannosaurus.[7][8][1]
Systematik
Daspletosaurus wird innerhalb der Familie Tyrannosauridae in der Unterfamilie Tyrannosaurinae klassifiziert, zusammen mit Tarbosaurus, Tyrannosaurus, Nanotyrannus und vielleicht Alioramus. Mitglieder dieser Unterfamilie sind enger mit Tyrannosaurus als mit Albertosaurus verwandt und zeichnen sich durch proportional größere Schädel und längere Oberschenkelknochen aus als die andere Unterfamilie, die Albertosaurinae.[1][9]
Daspletosaurus gilt für gewöhnlich als enger Verwandter von Tyrannosaurus rex oder sogar als dessen direkter Vorfahr durch Anagenese.[10] Gregory Paul (1988) ordnete Daspletosaurus torosus der Gattung Tyrannosaurus zu und stellte die Kombination Tyrannosaurus torosus auf;[5] dies fand jedoch keine allgemeine Akzeptanz.[7][1] Viele Forscher halten Tarbosaurus und Tyrannosaurus für Schwestertaxa oder sogar für die gleiche Gattung, mit Daspletosaurus als ein basalerer (ursprünglicherer) Verwandter.[1][11] Auf der anderen Seite glauben Phil Currie und Kollegen, dass Daspletosaurus näher mit Tarbosaurus und anderen asiatischen Tyrannosauriden wie Alioramus verwandt ist als mit dem nordamerikanischen Tyrannosaurus.[9] Die genauen Verwandtschaftsbeziehungen von Daspletosaurus werden vielleicht klarer, wenn alle Arten beschrieben wurden.
| Tyrannosauridae |
| ||||||||||||||||||||||||||||||
Kladogramm der Tyrannosauridae nach Holtz, 2004[1]
| Tyrannosauridae |
| ||||||||||||||||||||||||||||||||||||
Alternatives Kladogramm der Tyrannosauridae nach Currie und anderen 2003[9]
Entdeckungsgeschichte und Benennung
Das Typmaterial von Daspletosaurus torosus (CMN 8506) ist ein Teilskelett inklusive des Schädels, der Schulter, eines Arms, des Beckens, eines Oberschenkelknochens (Femur) und sämtlicher Wirbel des Halses, des Rumpfs und der Hüfte sowie der ersten elf Schwanzwirbel. Es wurde bereits im Jahr 1921 von Charles Mortram Sternberg entdeckt, welcher es für eine neue Art von Gorgosaurus hielt. Erst im Jahr 1970 beschrieb Dale Russell den Fund vollständig und stellte die neue Gattung Daspletosaurus auf – der Name leitet sich von den altgriechischen Wörtern δασπλητo-/daspleto- „furchtbar“ und σαῦρος/sauros „Echse“ ab.[12] Das Artepitheth torosus ist Latein und bedeutet „muskulös“.[2] Neben dem Typfossil gibt es lediglich ein weiteres gutbekanntes Fundstück, ein im Jahr 2001 entdecktes vollständiges Skelett. Beide Skelette wurden aus den Schichten der Oldman-Formation in der Judith River-Group in Alberta geborgen, die sich während des mittleren Campaniums vor 77 bis 76 Millionen Jahren ablagerten. Ein weiteres Skelett aus der jüngeren Horseshoe-Canyon-Formation in Alberta wurde mittlerweile dem Albertosaurus sarcophagus zugeschrieben.[13]
Unbenannte Arten
Zwei oder drei weitere Arten wurden mit den Jahren der Gattung Daspletosaurus zugeschrieben, obwohl bis heute keine dieser Arten eine sachgemäße Beschreibung oder einen wissenschaftlichen Namen erhalten hat. Stattdessen wurden alle als Daspletosaurus sp. bezeichnet – was jedoch nicht bedeutet, dass sie alle zur gleichen Art gehören.[8][1]

Neben dem Holotypus bestimmte Russell einen Fund, der von Barnum Brown im Jahr 1913 ausgegraben wurde, als Paratyp von Daspletosaurus torosus. Dieses Fundstück (AMNH 5438) besteht aus Teilen der Hinterbeine, des Beckens und einigen Wirbeln und wurde im oberen Bereich der Oldman-Formation in Alberta gefunden.[8] Später wurde dieser obere Bereich in Dinosaurier-Park-Formation umbenannt; die Formation datiert auf das mittlere Campanium (vor 80 bis 76 Millionen Jahren).[13] Im Jahr 1914 sammelte Brown ein nahezu vollständiges Skelett mit Schädel; vierzig Jahre später verkaufte es sein American Museum of Natural History an das Field Museum of Natural History in Chicago. In Chicago wurde es für viele Jahre als Albertosaurus libratus ausgestellt – nachdem man aber festgestellt hat, dass verschiedene Schädelpartien einschließlich des Großteils der Zähne mit Gips modelliert waren, wurde das Skelett (FMNH PR308) dem Daspletosaurus neu zugeordnet.[7] Über die Jahre wurden insgesamt acht Skelettfunde aus der Dinosaur Park-Formation gesammelt, die meisten innerhalb des Dinosaur Provincial Parks. Phil Currie glaubt, dass die Dinosaur Park-Funde eine neue Daspletosaurus-Art repräsentieren, die aufgrund bestimmter Schädelmerkmale unterschieden werden kann. Darstellungen dieser neuen Art wurden zwar veröffentlicht, eine vollständige Beschreibung und ein wissenschaftlicher Name stehen jedoch noch aus.[8]
Ein neues Tyrannosaurier-Fossil (OMNH 10131), inklusive Schädelfragmente, Rippen und Teile der Hinterbeine, wurde im Jahr 1990 aus New Mexico gemeldet und der heute als ungültig angesehenen Gattung Aublysodon zugeschrieben.[14] Viele spätere Autoren schrieben dieses Fossil zusammen mit einigen weiteren Funden aus New Mexico einer weiteren, unbenannten Daspletosaurus-Art zu.[8][1][15] Jedenfalls lassen neue, noch unveröffentlichte Forschungsergebnisse vermuten, dass es sich bei dieser Spezies aus dem Hunter-Wash-Schichtglied der Kirtland-Formation tatsächlich um einen frühen Tyrannosauriden handelte, der vielleicht mit Appalachiosaurus verwandt war.[16] Über das Alter der Kirtland-Formation herrscht derzeit Uneinigkeit – während einige Autoren das späte Campanium als Alter angeben,[17] meinen andere, die Formation sei jünger und ins frühe Maastrichtium zu datieren.[18]

Im Jahr 1992 veröffentlichten Jack Horner und Kollegen einen sehr vorläufigen Bericht über einen Tyrannosauriden aus den oberen Abschnitten der Two-Medicine-Formation (Campanium) von Montana, der als Übergangsspezies zwischen Daspletosaurus und dem späteren Tyrannosaurus interpretiert wurde.[10] Ein anderer Bericht aus dem Jahr 2001 beschreibt ein weiteres Teilskelett aus der oberen Two-Medicine-Formation, in dessen Bauchhöhle die Überreste eines juvenilen Hadrosauriers erhalten geblieben sind. Dieser Fund wurde Daspletosaurus zugeschrieben, aber keiner bestimmten Art.[19] Die Überreste von mindestens drei weiteren Daspletosaurus fand man in einem Bonebed der Two-Medicine-Formation.[20] Diese Funde müssen noch detailliert beschrieben werden, aber Currie glaubt, dass sämtliches Material aus der Two-Medicine-Formation zu einer dritten, noch unbenannten Daspletosaurus-Art gehört.[8]
Paläobiologie
Koexistenz mit Gorgosaurus

Im späten Campanium von Nordamerika war Daspletosaurus ein Zeitgenosse des Albertosaurinen Gorgosaurus. Dies ist eines der wenigen Beispiele einer Koexistenz zweier Tyrannosauriden. In modernen Raubtier-Gilden besetzen ähnlich große Raubtiere verschiedene ökologische Nischen, durch die der Konkurrenzkampf begrenzt wird. Unterschiedliche ökologische Nischen kommen vor allem durch Unterschiede im Verhalten, in der Anatomie oder in der Geografie zustande.[21] Verschiedene Studien versuchen zu klären, welche ökologischen Nischen Daspletosaurus und Gorgosaurus besetzten.
Dale Russell stellte die Hypothese auf, dass der leichter gebaute und häufigere Gorgosaurus die zu der Zeit reichlich vorhandenen Hadrosaurier gejagt haben könnte, während der robustere und weniger häufige Daspletosaurus auf die Ceratopsier spezialisiert gewesen sein könnte, die vielleicht weniger häufig und schwerer zu jagen waren.[2] Jedenfalls sind in einem Daspletosaurus-Skelett (OTM 200) aus der Two-Medicine-Formation die verdauten Überreste eines juvenilen Hadrosauriers in der Darmregion gefunden worden.[19] Die höheren und breiteren Schnauzen von Tyrannosaurinen wie Daspletosaurus sind mechanisch stärker als die flacheren Schnauzen von Albertosaurinen wie Gorgosaurus, obwohl die Zähne bei beiden Gruppen ähnlich gut ausgeprägt sind. Dies könnte auf Unterschiede in der Ernährung oder in den Fressmechanismen hindeuten.[22]
Andere Autoren vermuteten, dass die Konkurrenz zwischen den beiden Gattungen durch geografische Separation eingeschränkt wurde. Anders als bei einigen anderen Dinosauriergruppen scheint es keinen Zusammenhang mit der Distanz zum Meer zu geben – weder Daspletosaurus noch Gorgosaurus war in höheren oder niedrigeren Höhenlagen häufiger.[21] Jedenfalls scheint Gorgosaurus in nördlicheren Breitengraden häufiger gewesen zu sein, während Daspletosaurus häufiger im Süden anzutreffen war. Dasselbe Muster zeigt sich in anderen Dinosauriergruppen: Chasmosaurine Ceratopsier und Hadrosaurine Hadrosaurier sind in der Two-Medicine-Formation und im südwestlichen Nordamerika während des Campaniums ebenfalls häufiger. Thomas Holtz vermutet daher, dass Tyrannosaurinen, Chasmosaurinen und Hadrosaurinen die gleichen Lebensräume bevorzugten. Weiter merkt er an, dass am Ende des Maastrichtiums Tyrannosaurinen wie Tyrannosaurus rex, Chasmosaurinen wie Triceratops und Hadrosaurinen im westlichen Nordamerika weit verbreitet waren, während Albertosaurinen und Centrosaurinen ausstarben und Lambeosaurinen sehr selten wurden.[1]
Sozialverhalten
Ein juveniles Exemplar der Dinosaur Park-Spezies (TMP 94.143.1) zeigt Bissspuren im Gesicht, die von einem anderen Tyrannosaurier herrührten. Die Bissspuren sind verheilt, was zeigt, dass das Tier den Biss überlebt hat. Ein erwachsener Daspletosaurus aus dem Dinosaur Park zeigt ebenfalls derartige Bissspuren, was zeigt, dass Attacken auf das Gesicht nicht auf junge Tiere beschränkt waren. Zwar könnten die Bisse auch von anderen Gattungen herrühren – Aggressionen innerhalb von Arten, einschließlich des Beißens ins Gesicht, sind jedoch unter Prädatoren sehr häufig. Gesichtsbisse können bei anderen Tyrannosauriern wie Gorgosaurus und Tyrannosaurus sowie bei anderen Theropoden wie Sinraptor und Saurornitholestes beobachtet werden. Darren Tanke und Phil Currie glauben, dass die Bisse durch intraspezifische Konkurrenz um Territorien, Ressourcen oder Dominanz innerhalb einer Gruppe entstanden.[23]
Hinweise auf ein Leben in Gruppen bei Daspletosaurus stammen von einem Bonebed aus der Two-Medicine-Formation in Montana. Dieses Bonebed beinhaltet die Überreste von drei Daspletosaurus, inklusive eines großen Erwachsenen, eines kleinen Jungtiers und eines anderen Individuums mittlerer Größe. Weiter fanden sich die Überreste von mindestens fünf Hadrosauriern. Geologische Untersuchungen zeigen, dass die Überreste nicht an diesen Ort geschwemmt wurden, sondern dass alle Tiere gleichzeitig an derselben Stelle vergraben wurden. Die Hadrosaurier-Überreste sind zerstreut und zeigen zahlreiche Tyrannosaurier-Bissspuren, was zeigt, dass die Daspletosaurus an den Hadrosaurier-Kadavern gefressen hatten, bevor sie gestorben sind. Die Todesursache ist unbekannt. Currie spekuliert, dass die Daspletosaurus in einer Gruppe lebten, obwohl dies nicht mit Sicherheit gesagt werden kann.[20] Andere Wissenschaftler stehen den Hinweisen, dass Daspletosaurus und andere große Theropoden in Gruppen lebten, skeptisch gegenüber;[24] Brian Roach und Danial Brinkman vermuten, dass die sozialen Interaktionen von Daspletosaurus denen der rezenten Komodowaranen ähneln, bei denen unkooperative Individuen über Kadaver herfallen und Artgenossen attackieren.[25]
Ontogenese und Populationsbiologie
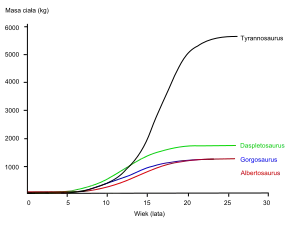
Paläontologen um Gregory Erickson haben die Individualentwicklung (Ontogenese) von Tyrannosauriden anhand von Skelettfunden studiert. So lassen Analysen des Knochengewebes (Histologie) auf das Alter des Tieres schließen, als es starb; auch können Wachstumsraten ermittelt werden, indem das Alter verschiedener Individuen mit der jeweiligen Größe verglichen wird. Erickson zeigte, dass Tyrannosauriden eine lange Zeit als Jungtiere verbrachten und dann in der Mitte ihres Lebens rapide wuchsen. Nach dieser rapiden Wachstumsperiode, die wohl mit der Geschlechtsreife endete, verlangsamte sich das Wachstum in erwachsenen Tieren beträchtlich. Zwar untersuchte Erickson Daspletosaurus lediglich aus der Dinosaur-Park-Formation, die Ergebnisse zeigen jedoch das gleiche Muster wie bei anderen Tyrannosauriden. Verglichen mit Albertosaurinen zeigte Daspletosaurus aufgrund seines höheren Erwachsenen-Gewichts ein schnelleres Wachstum während der rapiden Wachstumsperiode. Die Forscher ermittelten als größte Wachstumsgeschwindigkeit bei Daspletosaurus 180 Kilogramm pro Jahr – dabei gingen sie von einem Gewicht von 1800 Kilogramm für erwachsene Tiere aus. Zwar gaben andere Autoren an, Daspletosaurus sei schwerer; ein höheres Gewicht würde jedoch nur das Ausmaß der Wachstumsgeschwindigkeit ändern, nicht aber das generelle Muster.[3]
Indem die Forscher die Zahl der Funde jeder Altersgruppe verglichen, konnten sie Rückschlüsse auf die Populationsbiologie von Albertosaurus ziehen. Wie ihre Analysen zeigten, sind Jungtiere selten als Fossilien überliefert; Tiere innerhalb der rapiden Wachstumsphase und erwachsene Tiere finden sich wesentlich häufiger. Dies könnte daran liegen, dass die Knochen von Jungtieren seltener versteinern oder schwerer gefunden und gesammelt werden können – Erickson vermutet jedoch eine geringe Sterberate unter Jungtieren über einer bestimmten Größe, wie man es auch bei einigen modernen Großsäugern wie den Elefanten beobachten kann. Diese geringe Sterberate könnte aus dem Fehlen von Raubtieren resultieren, da Tyrannosauriden alle zeitgenössischen Raubtiere mit einem Alter von zwei Jahren in der Größe übertrafen. Zwar fanden Paläontologen bisher nicht genug Überreste für eine ähnliche Analyse bei Daspletosaurus; Erickson meint jedoch, dass das gleiche Muster zu zutreffen scheint.[26]
Paläoökologie
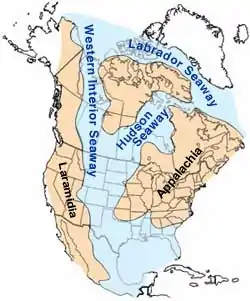
Alle bekannten Daspletosaurus-Fossilien wurden in Gesteinsformationen gefunden, die während des mittleren und späten Campaniums (vor ca. 80 bis 72 Mya) ablagerten. Seit der frühen Oberkreide wurde Nordamerika von einem Meer, dem Western Interior Seaway, in zwei Hälften geteilt, das auch den Großteil von Montana und Alberta bedeckte. Zu Zeiten von Daspletosaurus begann im Westen jedoch der Aufstieg der Rocky Mountains durch die laramische Gebirgsbildung, wodurch der Meerarm nach Osten und Süden verdrängt wurde. Flüsse strömten von den Bergen in den Western Interior Seaway und brachten Sedimente mit sich, die die Two-Medicine-Formation, die Judith River-Group und weitere Fossillagerstätten in der Region formten. Vor 73 Millionen Jahren breitete sich das Meer wieder nach Westen und Norden aus, und die ganze Region wurde von der Bearpaw-See bedeckt, deren marine Ablagerungen heute die Bearpaw-Formation im westlichen Nordamerika bilden.[27][28][29]
Daspletosaurus lebte in einem gewaltigen Überschwemmungsebene entlang der westlichen Küste der Western Interior Seaway. Große Flüsse bewässerten das Land, die gelegentlich die Region fluteten und mit neuem Sediment bedeckten. Wenn viel Wasser vorhanden war, gab es in der Region ein vielfältiges Tier- und Pflanzenleben – periodische Dürren führten aber immer wieder zu Massensterben, wie man auch an vielen Bonebeds in der Two-Medicine-Formation und der Judith-River-Formation erkennen kann, einschließlich des Daspletosaurus-Bonebed.[30] Ähnliche Umstände existieren heute in Ostafrika.[31] Periodische Vulkanausbrücke im Westen bedeckten die Region mit Aschen und führten zu weiteren Massensterben, reicherten den Boden aber mit Nährstoffen für ein zukünftiges Pflanzenwachstum an. Diese Aschen erlauben Forschern zudem eine präzise radiometrische Datierung der Gesteinsschichten. Wechselnde Meeresspiegel führten in der Judith-River-Formation zu einer Vielfalt von Lebensräumen zu verschiedenen Zeiten und Orten – neben den Überschwemmungsebenen gab es marine Habitate in Küstennähe, küstennahe Sumpfgebiete sowie Deltas und Lagunen.[28] Die Two-Medicine-Formation wurde in höhergelegenen Gebieten im Inland abgelagert als die beiden anderen Formationen.[29]
Die fossile Wirbeltierfauna der Two Medicine und der Judith-River-Formation ist sehr gut bekannt, da es zur Zeit der Ablagerungen reichliches Tierleben gab, periodische Naturkatastrophen zu hohen Sterberaten führten und große Mengen an Sediment abgelagert wurden. So sind viele Formen von Süßwasser- und Ästuar-Fischen überliefert, einschließlich Haien, Rochen, Stören, Knochenhechten und weiteren. In der Judith River-Gruppe blieben die Überreste von vielen aquatischen Amphibien und Reptilien erhalten, inklusive Frösche, Salamander, Schildkröten, Champsosaurus und Krokodile. Terrestrische Eidechsen wie Schienenechsen, Skinke, Leguane und Schleichen wurden ebenfalls entdeckt. Flugsaurier (Pterosauria) aus der Familie der Azhdarchidae und moderne Vögel wie Apatornis beherrschten den Himmel, während der Enantiornithe Vogel Avisaurus und verschiedene Säugetiere aus den Gruppen der Multituberculaten, Beutelsäuger und höheren Säugetiere zwischen den Beinen von Daspletosaurus und anderen Dinosauriern umherliefen.[28]
In der Oldman-Formation könnten verschiedene Dinosaurier zum Beutespektrum von Daspletosaurus torosus gehört haben, darunter Hadrosaurier wie Brachylophosaurus und Hypacrosaurus, kleine Ornithopoden wie Orodromeus, Ceratopsier wie Centrosaurus, Pachycephalosaurier, Ornithomimosaurier, Therizinosaurier und möglicherweise Ankylosaurier. Andere Räuber der Formation schließen Troodontiden, Oviraptorosaurier, den Dromaeosauriden Saurornitholestes und vielleicht Albertosaurine Tyrannosaurier (Gattung derzeit unbekannt) mit ein. Die Dinosaurier-Park- und Two-Medicine-Formation haben eine mit der Oldman vergleichbare Fauna, wobei insbesondere in der Dinosaurier-Park-Formation eine riesige Bandbreite an verschiedenen Dinosauriern entdeckt wurde.[28] Der Albertosaurine Gorgosaurus wurde zusammen mit Daspletosaurus in der Dinosaurier-Park-Formation und der oberen Two-Medicine-Formation gefunden.[21] Junge Tyrannosaurier haben vielleicht die Nischen zwischen adulten Tyrannosauriern und kleineren Theropoden aufgefüllt.[2][1][22][32]
Literatur
- John R. Horner, Don Lessem: The complete T. rex. Simon & Schuster, New York NY u. a. 1993, ISBN 0-671-74185-3.
Weblinks
Einzelnachweise
- Thomas R. Holtz Jr.: Tyrannosauroidea. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 111–136.
- Dale A. Russell: Tyrannosaurs from the late cretaceous of Western Canada (= Publications in Palaeontology. Bd. 1, ISSN 0068-8029). National Museum of Natural Sciences (Canada), Ottawa 1970, Digitalisat.
- Gregory M. Erickson, Peter J. Makovicky, Philip J. Currie, Mark A. Norell, Scott A. Yerby, Christopher A. Brochu: Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. In: Nature. Bd. 430, Nr. 7001, 2004, S. 772–775, doi:10.1038/nature02699.
- François Therrien, Donald M. Henderson: My theropod is bigger than yours … or not: estimating body size from skull length in theropods. In: Journal of Systematic Palaeontology. Bd. 27, Nr. 1, 2007, ISSN 1477-2019, S. 108–115, doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- Gregory S. Paul: Predatory Dinosaurs of the World. A complete and illustrated Guide. Simon & Schuster, New York NY u. a. 1988, ISBN 0-671-61946-2, S. 464.
- Per Christiansen, Richard A. Fariña: Mass prediction in theropod dinosaurs. In: Historical Biology. Bd. 16, Nr. 2/4, 2004, ISSN 0891-2963, S. 85–92, doi:10.1080/08912960412331284313.
- Thomas D. Carr: Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria). In: Journal of Vertebrate Paleontology. Bd. 19, Nr. 3, 1999, S. 497–520, doi:10.1080/02724634.1999.10011161.
- Philip J. Currie: Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta. In: Acta Palaeontologica Polonica. Bd. 48, Nr. 2, 2003, ISSN 0567-7920, S. 191–226, (PDF; 1,8 MB).
- Philip J. Currie, Jørn H. Hurum, Karol Sabath: Skull structure and evolution in tyrannosaurid dinosaurs. In: Acta Palaeontologica Polonica. Bd. 48, Nr. 2, 2003, S. 227–234, (PDF; 137 kB).
- John R. Horner, David J. Varricchio, Mark B. Goodwin: Marine transgressions and the evolution of Cretaceous dinosaurs. In: Nature. Bd. 358, Nr. 6381, 1992, S. 59–61, doi:10.1038/358059a0.
- Thomas D. Carr, Thomas E. Williamson, David R. Schwimmer: A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama. In: Journal of Vertebrate Paleontology. Bd. 25, Nr. 1, 2005, S. 119–143, doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2.
- Henry George Liddell, Robert Scott: A Lexicon abridged from Liddell and Scott's Greek-English Lexicon. 24th edition, carefully revised throughout. Ginn, Boston 1891 (Reprinted edition. Clarendon Press, Oxford 2010, ISBN 978-0-19-910207-5).
- David A. Eberth, Anthony P. Hamblin: Tectonic, stratigraphic, and sedimentologic significance of a regional discontinuity in the upper Judith River Group (Belly River wedge) of southern Alberta, Saskatchewan, and northern Montana. In: Canadian Journal of Earth Sciences. Bd. 30, Nr. 1, 1993, ISSN 0008-4077, S. 174–200, doi:10.1139/e93-016.
- Thomas M. Lehman, Kenneth Carpenter: A partial skeleton of the tyrannosaurid dinosaur Aublysodon from the Upper Cretaceous of New Mexico. In: Journal of Paleontology. Bd. 64, Nr. 6, 1990, ISSN 0022-3360, S. 1026–1032, Zusammenfassung.
- Thomas D. Carr, Thomas E. Williamson: A review of Tyrannosauridae (Dinosauria: Coelurosauria) from New Mexico. In: Spencer G. Lucas, Andrew B. Heckert (Hrsg.): Dinosaurs of New Mexico (= New Mexico Museum of Natural History and Science. Bulletin. Bd. 17, ISSN 1524-4156). New Mexico Museum of Natural History and Science, Albuquerque NM 2000, S. 113–146, Digitalisat (PDF; 4,48 MB).
- Michael Mortimer: Details on SVP 2002 Part 2. In: Dinosaur Mailing List. 16. Oktober 2002, abgerufen am 28. Juli 2014 (englisch).
- Robert M. Sullivan, Spencer G. Lucas: The Kirtlandian land-vertebrate „age“–faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of Western North America. In: Spencer G. Lucas, Robert M. Sullivan (Hrsg.): Late Cretaceous vertebrates from the Western Interior (= New Mexico Museum of Natural History and Science. Bulletin. Bd. 35). New Mexico Museum of Natural History and Science, Albuquerque NM 2006, S. 7–29.
- Michael J. Ryan: Kirtland Formation. In: Philip J. Currie, Kevin Padian (Hrsg.): Encyclopedia of dinosaurs. Academic Press, San Diego CA u. a. 1997, ISBN 0-12-226810-5, S. 390–391.
- David J. Varricchio: Gut contents from a Cretaceous tyrannosaurid: implications for theropod dinosaur digestive tracts. In: Journal of Paleontology. Bd. 75, Nr. 2, 2001, S. 401–406, doi:10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2.
- Philip J. Currie, David Trexler, Eva B. Koppelhus, Kelly Wicks, Nate Murphy: An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA). In: Kenneth Carpenter (Hrsg.): The Carnivorous Dinosaurs. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34539-1, S. 313–324.
- James O. Farlow, Eric R. Pianka: Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs. In: Historical Biology. Bd. 16, Nr. 1, 2002, S. 21–40, doi:10.1080/0891296031000154687 (zurzeit nicht erreichbar).
- Eric Snively, Donald M. Henderson, Doug S. Phillips: Fused and vaulted nasals of tyrannosaurid dinosaurs: Implications for cranial strength and feeding mechanics. In: Acta Palaeontologica Polonica. Bd. 51, Nr. 3, 2006, S. 435–454, (PDF; 682,88 kB).
- Darren H. Tanke, Philip J. Currie: Head-biting behavior in theropod dinosaurs: paleopathological evidence. In: Gaia. Revista de Geociências. Bd. 15, 1998, ISSN 0871-5424, S. 167–184, online.
- David A. Eberth, Richard T. McCrea: Were large theropods gregarious? In: Journal of Vertebrate Paleontology. Bd. 21, Supplement to Nr. 3 = Abstracts of Papers, 61st Annual Meeting of the Society of Vertebrate Paleontology, 2001, S. 46A.
- Brian T. Roach, Daniel T. Brinkman: A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. In: Bulletin of the Peabody Museum of Natural History. Bd. 48, Nr. 1, 2007, ISSN 0079-032X, S. 103–138, doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- Gregory M. Erickson, Philip J. Currie, Brian D. Inouye, Alice A. Winn: Tyrannosaur Life Tables: An Example of Nonavian Dinosaur Population Biology. In: Science. Bd. 313, Nr. 5784, 2006, S. 213–217, doi:10.1126/science.1125721, PMID 16840697.
- Joseph M. English, Stephen T. Johnston: The Laramide Orogeny: what were the driving forces?. In: International Geology Review. Bd. 46, Nr. 9, 2004, ISSN 0020-6814, S. 833–838, doi:10.2747/0020-6814.46.9.833.
- David A. Eberth: Judith River Wedge. In: Philip J. Currie, Kevin Padian (Hrsg.): Encyclopedia of dinosaurs. Academic Press, San Diego CA u. a. 1997, ISBN 0-12-226810-5, S. 379–385.
- Raymond R. Rogers: Judith River Wedge. In: Philip J. Currie, Kevin Padian (Hrsg.): Encyclopedia of dinosaurs. Academic Press, San Diego CA u. a. 1997, ISBN 0-12-226810-5, S. 760–765.
- Raymond R. Rogers: Taphonomy of three dinosaur bonebeds in the Upper Cretaceous Two Medicine Formation of northwestern Montana: evidence for drought-induced mortality. In: Palaios. Bd. 5, Nr. 5, 1990, ISSN 0883-1351, S. 394–413, doi:10.2307/3514834.
- Howard J. Falcon-Lang: Growth interruptions in silicified conifer woods from the Upper Cretaceous Two Medicine Formation, Montana, USA: implications for palaeoclimate and dinosaur palaeoecology. In: Palaeogeography, Palaeoclimatology, Palaeoecology. Bd. 199, Nr. 3/4, 2003, ISSN 0031-0182, S. 299–314, doi:10.1016/S0031-0182(03)00539-X.
- James O. Farlow: Speculations about the diet and foraging behavior of large carnivorous dinosaurs. In: The American Midland Naturalist. Bd. 95, Nr. 1, 1976, ISSN 0003-0031, S. 186–191, doi:10.2307/2424244, Zusammenfassung.