Spitzkiele
Die Spitzkiele oder Fahnenwicken (Oxytropis) sind eine Pflanzengattung in der Unterfamilie der Schmetterlingsblütler (Faboideae) innerhalb der Familie der Hülsenfrüchtler (Fabaceae). Die etwa 310 Arten sind auf der Nordhalbkugel in Nordamerika, Eurasien und Afrika weitverbreitet.[1]
| Spitzkiele | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Pracht-Spitzkiel (Oxytropis splendens) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Oxytropis | ||||||||||||
| DC. |
Beschreibung
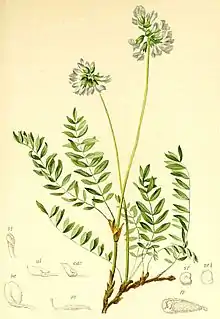



Vegetative Merkmale
Die Spitzkiel-Arten sind meist ausdauernde, krautige Pflanzen, die ein verholztes Rhizom bilden. Es gibt stängellose wie auch stängelbildende Arten.
Die Laubblätter sind in Blattstiel und Blattspreite gegliedert. Die Blattspreite ist meist unpaarig gefiedert oder die Endfieder ist zu einem Stachel reduziert; bei manchen Arten ist nur ein Fiederblatt vorhanden. Die Fiederblättchen sind ganzrandig und am Grund asymmetrisch. Die auffälligen, meist haltbaren Nebenblätter können mit dem Blattstiel verwachsen, untereinander frei oder verwachsen sein.[1]
Blütenstände und Blüten
Die seitenständigen, traubigen, ährigen oder kopfigen Blütenstände enthalten eine bis viele Blüten. Die Tragblätter sind meist dreieckig oder lanzettlich-dreieckig. Deckblätter fehlen meist oder es sind manchmal schmal-dreieckige vorhanden.[1]
Die zwittrigen Blüten sind zygomorph und fünfzählig mit doppelter Blütenhülle. Die fünf Kelchblätter sind glockenförmig bis röhrig verwachsen[1] und die Kelchzähne sind annähernd gleich. Die fünf violetten, purpurfarbenen, weißen oder blassgelben Kronblätter stehen in der typischen Form der Schmetterlingsblüte zusammen. Die Kronblätter sind lang genagelt, meistens frei und fallen nach der Anthese ab. Flügel und Schiffchen sind je nach Art unterschiedlich geformt.[1] Das Schiffchen hat vorne eine deutliche aufgesetzte Spitze. Von den zehn Staubblättern ist das oberste frei. Das sitzende oder gestielte, einzige Fruchtblatt ist kahl oder flaumig behaart und enthält meist viele Samenanlagen.[1] Der gerade oder öfter gekrümmte Griffel besitzt eine endständige Narbe.[2]
Früchte und Samen
_seed_and_embryo.jpg.webp)




Die Hülsenfrüchte sind häufig aufgedunsen, haben ein Fach, häufiger sind sie durch eine falsche Längsscheidewand zweifächrig und enthalten meist mehreren Samen. Die Oberfläche der Früchte ist kahl oder behaart. Die relativ kleinen Samen sind linsenförmig, nierenförmig oder kugelig, ohne Strophiole.[2]
Ökologie

Blütenökologisch handelt es sich um Schmetterlingsblumen mit einem einfachen Klappmechanismus. Flügel und Schiffchen sind gelenkig verbunden. Die Bestäubung erfolgt durch Hummeln oder andere langrüsselige Apoideen, in Gebirgslagen häufig durch Schmetterlinge.
Innerhalb ihres Lebensraumes sind sie wichtige Futterpflanzen für Wildtiere (mit einigen die hohe Toxizität zeigen).[3] In den nördlichen „Rockies“ sind sie Nahrung von Dickhornschafen und Schneeziegen[4]. Ebenso sind sie Nahrungsquelle der Pfeifhasen wie dem Koslow-Pfeifhase Tibets.[5] Neben dem hohen Futterwert der eiweißreichen Samen sind sie darüber hinaus auch z. T. Wintersteher und dienen Wild- wie Haustieren als wichtige Winternahrung. Da sie daneben auch in hoher Dichte aufkommen, für das Bulgarische Pirin-Gebirge wurden an zwei Beobachtungsstellen 40.000 Blüten des Alpen-Spitzkiels in einem Erfassungszeitraum angegeben, werden sie dort von Schafen dort auch gezielt gefressen.[6]
Wie andere Leguminosen fixieren Spitzkiele Stickstoff aus der Luft. Dies erfolgt durch eine Symbiose in der Leguminosenwurzel mit spezialisierten kohlenstoffheretrophen N2-Fixierern.[7] Hierbei siedeln sich Stickstoff-fixierende Mikroorganismen in Wurzelzellen der Spitzkiele an. Da sie von den Wurzelexudaten in der Rhizoplane ernährt werden, sind sie deshalb viel ergiebiger für die Bindung von Luftstickstoff als freilebende Mikroorganismen. In Spitzkielen sind Bakterien der Gattung Rhizobium mit einigen Arten und vielen physiologischen Rassen die in den Wurzelknöllchen mit den Spitzkielen zusammenleben. In den nährstoffarmen Verbreitungsgebieten der Spitzkiele in der hohen Arktis und im Hochgebirge gehören Pflanzen die symbiotisch Stickstoff fixieren zu den Schlüsselarten, da sie hierüber Boden mit Stickstoff anreichern.[8] Spitzkiele üben hierüber einen Einfluss auf Pflanzengesellschaften aus, der über die reine Fixierung von Stickstoff in den Boden hinüber geht. So verringern sie aufgrund hohen Bedarfs an Phosphor die Verfügbarkeit von Bodenphosphor, unterdrücken Konkurrenzen durch allelopathische Effekte und unterdrücken Keimung und die Etablierung von Setzlingen. Durch ihren Polsterwuchs können sie ebenfalls für andere Arten Licht- und Wasserverfügbarkeit reduzieren. Die Rolle der N-Fixierung die entweder über Rhizobien (Leguminosen) oder Mykorrhiza (Ericaceen, Dryas) erfolgt, ist über den Transfer von Stickstoff zu nicht N-fixierenden Pflanzen von Bedeutung. In der Lebenswelt der Arktis und alpiner Höhenstufen ist durch Kälte, Trockenheit und Stickstoffreglementierung die positive Interaktion und Konkurrenz von Arten für die Pflanzengesellschaften essentiell. Die Rolle der Spitzkiele wie andere Leguminosen der kontinentalen Kälteregionen sowie alpiner Habitate wird, indem sie Energie- und Nährstoffflüsse modifizieren, in der ökologischen Literatur mit dem englischen Fachbegriff Ecosystem Engineers beschrieben. Schlüsselarten wie Spitzkiel und die oft mit diesen vergesellschaftete Dryas, als „super-dominanten“ Art periglazialer Landschaften, sind insbesondere in der Pionierphase bedeutsam, indem sie Bodenentwicklung initiieren und Lockersubstrate festigen.
Für den Alpenraum gehört der Alpenspitzkiel (Oxytropis campestris) zu den Arten die als Alpenschwemmlinge regelhaft in den Schotterterrassen der Wileflüsse vorkommen. Auch hier ist neben dem Alpenspitzkiel der hohe Anteil an Fabaceen in der Artengarnitur auffallend (Astragalus alpinus, Lotus corniculatus, Trifolium repens). Fabaceen haben auf den nährstoffarmen Rohböden aufgrund Symbiose mit stickstoffbindenden Bakterien einen Konkurrenzvorteil.[9][10]
Die Samen der Spitzkiele sind ausbreitungsökologisch den Windstreuern (allg. Semachorie) zuzuordnen.[11] Sie sind allgemein kastanienbraun und nierenförmig und durch Hartschaligkeit ausgezeichnet. Durch sind sie sowohl gegen Gasaustausch als auch Wasser undurchlässig.[12] Spitzkielsamen zeigen eine physikalische Keimruhe. Eine erfolgreiche Einleitung der Samenkeimung erfolgt, nachdem diese mechanisch beschädigt, d. h. perforiert, werden. Die Oxytropis-Samen keimen damit unabhängig von klimatischen Gegebenheiten und Temperaturen; Stratifikation durch Kälteeinfluss brachte unter Laborbedingungen keine positive Korrelation zum Keimungserfolg, jedoch mechanische Skarifizierung der Samencuticula.[13] Eine Skarifizierung unter natürlichen Bedingungen erfolgt in periglazialen Klimazonen durch häufige Frostwechsel oder Windschurf; unter anderen ist der Alpen-Spitzkiel in seinem alpinen Verbreitungsgebiet auch ein typischer Vertreter der Gesellschaften an Windkanten. Erfolgt kein Aufbrechen der Schale, dann beträgt die Keimrate weniger als 10 %. In Kultur ist für eine erfolgreiche Keimung daher eine Skarifizierung der Samen notwendig. Die harte Schale wird mit feinem Sandpapier aufgeraut oder im Labor kurz (20–60 min) mit konzentrierter Schwefelsäure oder kochendem Wasser behandelt. Hierdurch können Oxytropis-Samen innerhalb kurzer Zeit im Wasser aufquellen. Dabei führen Schwefelsäure und kochendes Wasser mitunter zum Abtöten des Embryo. Diese Methoden haben einen wesentlich geringeren Keimungserfolg als mechanisches Aufrauen.[13] Die Samen beenden ihre Keimruhe hierdurch umgehend.
Die hartschaligen Samen der Spitzkiele besitzen einen einheitlichen Aufbau: der embryonale Hohlraum der Kotyledonen wird von einer äußeren Cuticula und einer dicken Makrosklereide umschlossen.[14] Auch die Oberfläche der Samencuticula ist bei den Spitzkielen charakteristisch: Bei Betrachtung unter dem Elektronenmikroskop sind spezifische Samenmantelmuster zu erkennen, die für einzelne Sektionen charakteristisch sein können. Unter dem Lichtmikroskop wirken die Samenmantel jedoch glatt. Da sich die Samenmantelmuster der Gattung Oxytropis allgemein von Astragalus unterscheiden, sind sie ein weiteres Merkmal der Gattungstrennung.
In den Ökosystemen der niederen Arktis auf der Victoria-Insel Kanadas sind Spitzkiele in Dryas-Salix-Leguminosen-Halden charakteristisch. Hierin besetzen sie immer Kuppen mit guter Dränage.[15]
Evolution
Aus evolutionärer Sicht sieht Boris Aleksandrovich Yurtsev (russ. Борис Александрович Юрцев) die Entwicklung der Gattung Oxytropis aus alpinen Hemikryptophyten zu Kryptophyten der Arktis, die sich durch Mikrophyllie, Verkleinerung der Wuchsform, Verringerung der Anzahl der Blättchen eines Blattfieders wie der Blütenzahl pro Blütenstand verändert hat und für die adaptive Radiation der Gattung wichtige taxonomische Kennzeichen geben.[16] Zytotaxonomisch vollzog sich gleichzeitig eine Entwicklung von diploiden Arten in südsibirischen Gebirgen zu polyploiden arktischen. Im Kontrast zu arktischen Arten der Gattung Astragalus finden sich bei arktischen Oxytropis-Arten keine Mesophyten. Ökologisch variieren die Spitzkiel-Arten zwischen Xeromesophyten und Cryo-Xeromesophyten zu Steppen-Xerophyten, Xero-Petrophyten oder Cryo-Xerophyten. Nur eine Unterart (Oxytropis middendorfii subsp. middendorfii) wächst in mesomorphen Kraut-Zwergstrauch-Moos-Tundren. Yurtsev nimmt an, dass die Vorfahren der heutigen Spitzkiel-Arten vielleicht auf instabilem Schutt und Geröllen siedelten, dies befähigte sie zur Adaptation an niedrige Sommertemperaturen und den Wechsel an Lebensräume in Tundren.[16]


Vorkommen und Vergesellschaftung
Verbreitung


Die Gattung Spitzkiele (Oxytropis) ist auf der Nordhalbkugel in Nordamerika, Eurasien und Afrika weitverbreitet.[1] Die Oxytropis-Arten gedeihen von gemäßigten über subarktischen bis arktischen Gebieten.[17] Spitzkiele besiedeln noch die arktischen Inseln nördlich des Polarkreises. Sie wurden noch bei 75°54' N an der Ibbett Bay in Kanada aufgesammelt.[18] Sie besiedeln dabei weder die ozeanischen Regionen der Südspitze Südgrönlands noch Island oder Spitzbergen. Erst in den kontinentalen Kältetundren reichen sie auch über die arktischen Inselgruppen wie der Wrangelinsel hinaus. Ein Zentrum der arktischen Verbreitung ist der Amphiberingische Bereich zwischen Tschukotka und Alaska. Amphiatlantisch sind arktisch verbreitete Leguminosen aufgrund des ozeanischen feuchten Klimas kaum anzutreffen oder sehr selten. Sowohl in Neuengland als auch auf den nördlichen Britischen Inseln in Schottland gehören Oxytropis-Arten hier zu den raren und auch gefährdeten Arten.[19][20] Für solche gefährdeten Populationen werden Reintroduktionskampagnen wie beispielsweise für den Seidenhaar-Spitzkiel (Oxytropis halleri) in Schottland durchgeführt.
Bei der Betrachtung der Gesamtareale fällt auf, dass die Hochgebirgselemente bei Oxytropis direkt an die Verwandtschaftskreise der Steppenflora anschließen. Die eurasisch oder zirkumpolare kontinentale arktisch + alpine (altaiisch-alpisch + arktische) Arten haben eine Entfaltung im sibirisch-ostasiatischen und im nordwestamerikanischen Raum. Diese ist viel größer als im europäischen und im nordostamerikanischen, wo beträchtliche Lücken in der Verbreitung zu verzeichnen sind. Häufung und Sippenentfaltung in den Gebirgsländern im Umkreis Zentralasiens und der Rocky Mountains betonen die Kontinentalität im Verbreitungscharakter.[21] In den süd- und mitteleuropäischen Hochgebirgen sind nur wenige der hierher gehörenden Arten weiter verbreitet (Oxytropis pyrenaica). Bei den meisten beschränkt sich die Vorkommen auf zentrale Teile der Alpen und zerstreute, meist begrenzte Gebiete der Karpaten und im Dinarisch-Pindischen Raum (Oxytropis lapponica). Neben den weitverbreiteten eurasischen und zirkumpolaren Hochgebirgsarten kommen aus denselben Verwandtschaftskreisen süd-mitteleuropäische Elemente vor, die zum Teil eine weite Verbreitung haben (Oxytropis pyrenaica), meistens aber wie Oxytropis triflora, Oxytropis foetida vorwiegend auf zentrale Teile der Alpen und mehr oder weniger kontinentale Bereiche der anderen Hochgebirge beschränkt sind. Zu erwähnen ist noch Oxytropis halleri, die eine mehr oder weniger disjunkte süd-mitteleuropäisch alpine Verbreitung hat und in Schottland wiederkehrt. Beim Alpen-Spitzkiel (Oxytropis campestris) ist in den Alpen das Vorkommen an Windkanten (Elynetum) charakteristisch. Braun-Blanquet hatte hieraufhin in der Blaugrasrasenklasse Elyno-Seslerietea den Verband Oxytropi-Elynion für die windexponierten Rasen auf flachgründigen Kalkböden aufgestellt. Eine vikariierende Variante ist in den Dinariden im pflanzensoziologischen Verband Oxytropidion dinaricae mit dem Dinarischen Spitzkiel ausgebildet. An Windkanten sind Schneearmut, Kälte und die den Gipfelwinden ausgesetzte Exposition als trocken-kalte Standorte vergleichbar den kontinentaler Klimazonen ausgeprägt. Mit der Windwirkung ist eine stärkere Evaporation als auf normalen sonnigen Standorten gegeben, die eine Dominanz des Nacktrieds (Kobresia myosuroides) bedingen. Wie Oxytropis findet auch das Nacktried seinen Verbreitungsschwerpunkt in den südsibirischen Gebirgen.[22]
Unter den Steppenpflanzen von Oxytropis erstrecken sich die Vorkommen von den submeridionalen Gebirgen (Mongolei, Altai, Kaukasus, ostsubmediterrane Bergländer) in das nördlich vorgelagerte Hügel- und Flachland (Oxytropis pilosa). Diese Arten sind Elemente der Trockenrasen. Manche der Arten sind jedoch nicht mehr in den Berg- und Hügelsteppen Europas, sondern nur in den Zentralalpen beschränkt (Oxytropis purpurea). Kein einziger Spitzkiel ist in der ozeanischen Laubwaldregion Westeuropas verbreitet, was den zutage tretenden kontinentalen Verbreitungscharakter deutlich erkennen lässt.[23]
Für den Tibetisch-Himalayischen Bereich wurden von Bernhard Dickoré und Monika Kriechbaum aufgrund der oftmals nur auf Einzelaufsammlungen basierenden Artaufstellungen realistisch nur 30–40 guter Arten mit großen Verbreitungsgebieten angegeben.[24] Hier ist es insbesondere der trockenere Nordwesten im Pamir und Karakorum sowie im NO des Tibetischen Plateaus (Qinghai), wo sich das Diversitätszentrum Zentralasiens und Mongolischen Hochlandes findet. Habitate wie bei der 2006 beschriebenen Oxytropis iridum sind Höhen um 3500 – 4150 m (bis 4660 m). Es handelt sich um semiaride Standorte innerhalb der Waldgrenze mit einer mittleren Jahrestemperatur von 5,2 °C und 258 mm Jahresniederschlag. Für Oxytropis iridum werden steile Fels- und Schutthänge mit geringer Vegetationsbedeckung (ca. 50 %) genannt.
Diversität
Von ihrem Verbreitungszentrum in Südsibirien haben sie sich voreiszeitlich zirkumpolar verbreitet. Sie stellen in den Tundren der Paläarktis eine der wenigen artenreichen Gattungen dar. Unter den Leguminosen hat nur die Gattung der Spitzkiele in einer artenreichen Radiation Lebensräume der Arktis neu besetzen können. Für die Arktis werden nach Angaben der Checklist of the Panarctic Flora (Elven et al. o. Jahr) 2043 Gefäßpflanzen angegeben. Nach Carex (137), Salix (68), Potentilla (60) ist Oxytropis mit 46 Arten vor Draba (44), Papaver (36), Poa (35), Puccinellia und Saxifraga (31) die artenreichste Gattung.[25]
Im Jahr 1948 wurden 276 Taxa für das ehemalige Territorium der UdSSR festgestellt.[26] In China gibt es 133 Arten, 74 davon nur dort (Stand 2010).[1]
In Mittel- und Südosteuropa kommen folgende 13 Arten vor: Feld- oder Alpen-Spitzkiel (Oxytropis campestris), Dinarischer Spitzkiel (Oxytropis dinarica), Drüsiger Spitzkiel (Oxytropis fetida), Seidenhaar-Spitzkiel (Oxytropis halleri), Schweizer Spitzkiel (Oxytropis helvetica), Berg-Spitzkiel (Oxytropis jacquinii), Lappland-Spitzkiel (Oxytropis lapponica), Pyrenäen-Spitzkiel (Oxytropis neglecta), Zottiger Spitzkiel (Oxytropis pilosa), Prenj-Spitzkiel (Oxytropis prenja), Dreiblüten-Spitzkiel (Oxytropis triflora), Oxytropis urumovii, Vinschgauer Seidenhaar-Spitzkiel (Oxytropis xerophila).[27][28][29][30][31]
Neben der Blütenfarbe ist bei den europäischen Spitzkiel-Arten die Pflanzengröße wichtiges Unterscheidungsmerkmal. Die Größe des Feld- oder Alpen-Spitzkiels (Oxytropis campestris) variiert zumeist je nach Höhenfundlage, er wird jedoch selten als Miniaturform aufgefunden. Der ähnliche Dinarische Spitzkiel variiert bei seinen fünf Unterarten und Varietäten sehr viel stärker, wobei die Größe nach Süden sukzessive abnimmt. Beim verwandten und ähnlichen bulgarischen Endemiten Oxytropis urumovii kommen nur sehr kleinwüchsige Pflanzenexemplare vor, die kaum ein Drittel der Größe des Alpen-Spitzkiels oder der Nominatform des Dinarischen Spitzkiels erreichen. Auch weitere Arten wie der Prenj-Spitzkiel oder der Schweizer Spitzkiel sind Miniaturformen, die in ihrem Lebensraum unverwechselbar sind, da sie nie über 10 Zentimeter hoch werden.
Lebensräume
.jpg.webp)
Die Oxytropis-Arten gedeihen in sandigen und kiesigen, generell gut drainierten Böden um Gletscherrandseen, auf Gebirgsgeröllhalden und Moränen sowie Gebirgsgrasländern.[17] Arktische Vertreter besiedeln in verschiedenen Arten sehr unterschiedliche Mikrohabitate: Presseishügel, Flussterrassen, Tundren, Hänge, offene Kämme der flachen Tundra; Fließerden auf solifludialen Hängen, mäßig bis gut dränierte Gebiete, Kies, Sand, Schluff, Geschiebemergel. Standorte mit hohen organischen Bestandteil und kalkreiche Substrate.[32] Spitzkiel-Arten fehlen in Wüsten- und Halbwüstenlandschaften der Erde und finden sich in den gemäßigten Zonen zumeist in Gebirgshabitaten.[26] Innerhalb ihres Lebensraumes stabilisieren durch die tiefe Pfahlwurzel Flussböschungen, Hänge und künstliche Dämme.[3]
Als Pionierpflanzen besiedeln sie nach Vulkanausbrüchen die Vulkane Kamtschatkas.[33] Über spezifische Eigenschaften ihres Wurzelsystems können Spitzkiel-Arten vulkanisches Lockermaterial festigen. Sie besiedeln Habitate, die noch keine Bodenbedeckung zeigen, was über diese Aktivität zur Bodenentwicklung beiträgt und einer weiteren Besiedlung von anderen Pflanzenarten führt. Die Symbiose von Spitzkiel-Arten mit Knöllchenbakterien unterstützt diese Entwicklung weiterhin.[34] Unter den Leguminosen, die typisch arktisch verbreitet sind (Oxytropis, Astragalus, Hedysarum) hat nur Oxytropis eine intensive Speziation unterlaufen.[16]
Während das Vorkommen des Alpen-Spitzkiels in seinem Teil-Verbreitungsgebieten in den Europäischen Alpen aus nacheiszeitlicher Einwanderung aus unbekannten pleistozänen Refugialräumen erklärt wird,[35] sind andere Arten wie Oxytropis urumovii aus dem Bulgarischen Pirin vermutlich Glazialrelikte.[36] Schönswetter et al. 2004 folgern aus dem Fehlen von phylogeographischen Mustern in den Genen des Alpen-Spitzkiels in den Alpen, Pyrenäen und der Tatra sowie seiner guten Anpassung an Steppenhabitate, dass diese Art während der Eiszeit möglicherweise aus Tieflandsteppen und Tundrengebieten in die Gebirge eingewandert ist.[37]
Vergesellschaftung
Pflanzensoziologische Einheiten alpiner Hochgebirge der Subtropen bis Gemäßigten Breiten haben Spitzkiel-Arten als ihre Charakter- und Kennarten: in den Südostdinariden ist es der Verband Oxytropidion dinaricae (mit Oxytropis dinarica und Oxytropis prenja) auf alpinen Kalksteinmagerrasen der Gipfel und Grate der Hochdinariden, im Iran gibt es die Klasse Oxytropidetea persica (mit Oxytropis persica) für Schneeboden-Gesellschaften auf Silikatgestein im Elburs-Gebirge, in den Westalpen sind es die pflanzensoziologische Ordnung Oxytropido-Kobresietalia mit dem Verband Oxytropido-Elynion.[38][39]
Systematik und botanische Geschichte
Die Gattung Oxytropis wurde durch Augustin Pyrame de Candolle 1802 aufgestellt. Jedoch ist aufgrund einer Übernahme der Phrase Astragalus montanus bei Adrian van Royens durch Linné, die auf einer von Clusius später nicht mehr eindeutig zuzuordnender Onobrichis montana fußte, in der nominellen Typusart Oxytropis montana (L.) DC.[40] ein Einschluss der Gruppen um Oxytropis pyrenaica oder Oxytropis jaquinii gegeben.[41] Spätere Emendationen, beispielsweise durch Bunges, stifteten noch mehr Verwirrung, so dass französische und deutsche Botaniker im Epitheton montana zwei unterschiedliche Spitzkiel-Arten einschlossen. Daher wurde das Epitheton montana von Walter Gutermann und Hermann Merxmüller 1961 als Nomen ambiguum in der weiteren taxonomische Verwendung bei Oxytropis ausgeschlossen (Oxytropis montana (L.) DC. partim et auct. partim, non sensu stricto (sec. Bunge), nomen ambiguum rejiciendum).[41] Der Gattungsname Oxytropis leitet sich von den griechischen Wörtern oxys für spitz sowie tropis für Schiffskiel ab und bezieht sich auf die am Schiffchen aufgesetzte Spitze, die die Gattungen Oxytropis und Astragalus morphologisch unterscheidet. Ein Synonym für Oxytropis DC. nom. cons. ist Spiesia Neck. ex Kuntze.[1]
Die Gattung Oxytropis gehört zur Tribus Galegeae in der Unterfamilie Faboideae innerhalb der Familie der Fabaceae.[1] Sie wurde früher als Untergattung der Gattung Astragalus, Astragalus subg. Oxytropis, geführt.
Zurzeit (Stand Mitte 2015) ermöglichen molekulargenetische Daten noch keine umfassende phylogenetische Abgrenzung innerhalb der Gattung Oxytropis und es existieren noch keine auf solchen Daten basierende Revisionen einzelner Verwandtschaftsgruppen. Allgemein sind viele Oxytropis-Arten morphologisch ähnlich, was schon in der einzigen bis dato vorliegende Monographie der Gattung Oxytropis durch Alexander von Bunge 1874 bemerkt wurde. Bunge gliederte 1874 die damals 181 Arten der Gattung Oxytropis in seinem Werk Species Generis in Oxytropis DC. in vier Untergattungen mit 19 Sektionen.[42] Die Flora der UdSSR 1948 unter Leitung der Bearbeitung der Gattung Oxytropis durch Alexander Grossheim übernahm die Gliederung von Bunge ergänzte jedoch die Bunges taxonomische Bearbeitung und Gliederung in vielerlei Hinsicht. Eine generelle Revision der Gattung steht aber nach wie vor aus.
Die Gattung Oxytropis enthält etwa 310 Arten.[1][43] Je nach taxonomischer Auffassung einzelner Autoren und Länder wird der Umfang der Gattung und die Arten kontrovers diskutiert:[44]
.jpg.webp)
.jpg.webp)




.JPG.webp)


.jpg.webp)
.jpg.webp)

.jpg.webp)



.jpg.webp)


.jpg.webp)


.jpg.webp)
- Oxytropis acanthacea Jurtzev: Sie kommt in der Mongolei nur in den Provinzen Bajan-Ölgii-Aimag sowie Chowd-Aimag und im asiatischen Teil Russlands nur in der autonomen Republik Tuwa vor.[43]
- Oxytropis aciphylla Ledeb. (Syn.: Oxytropis kossinskyi B.Fedtsch. & Basil., Oxytropis lycotriche Bunge): Sie ist in Russland, in der Mongolei, im autonomen Gebiet Innere Mongolei, im uigurischen autonomen Gebiet Xinjiang und in den chinesischen Provinzen Gansu, Ningxia sowie Qinghai verbreitet.[1]
- Oxytropis adamsiana (Trautv.) Jurtzev: Sie kommt im asiatischen Teil Russlands nur in Burjatien, im Oblast Tschita und im Autonomen Kreis Taimyr vor.[43]
- Oxytropis adenophylla Popov: Sie kommt im asiatischen Teil Russlands nur in Burjatien, Irkutsk und im Oblast Tschita vor.[43]
- Oxytropis admiranda Rech. f.: Sie kommt in Afghanistan vor.[43]
- Oxytropis adscendens Gontsch.: Sie kommt in Kasachstan nur bei Schymkent sowie Taras und in Kirgisistan nur bei Frunse vor.[43]
- Oxytropis aellenii Vassilcz.: Sie kommt im Iran vor.[43]
- Oxytropis afghanica Rech. f. & Koie: Sie kommt in Afghanistan vor.[43]
- Oxytropis ajanensis (Regel & Tiling) Bunge: Sie kommt im russischen Khabarovsk vor.[43]
- Oxytropis alajica Drobow: Dieser Endemit kommt in Kirgisistan nur in Osh vor.[43]
- Oxytropis alavae Rech. f.: Sie kommt im Iran vor.[43]
- Oxytropis alberti-regelii Vassilcz.: Sie kommt in Kasachstan nur bei Schymkent vor.[43]
- Oxytropis albiflora Bunge: Sie kommt im asiatischen Teil Russlands nur in Sacha vor.[43]
- Oxytropis albovillosa B.Fedtsch.: Sie kommt in Kasachstan nur bei Schymkent sowie Taras, in Kirgisistan nur bei Frunse sowie Osh und in Usbekistan nur in Taschkent vor.[43]
- Oxytropis almaatensis Bajtenov: Dieser Endemit kommt in Kasachstan nur bei Alma-Ata vor.[43]
- Oxytropis alpestris Schischkin: Sie kommt im asiatischen Teil Russlands nur in Gorno-Altaisk vor.[43]
- Oxytropis alpina Bunge (Syn.: Oxytropis alpicola Turcz., Oxytropis frigida Kar. & Kir.): Sie ist in Xinjiang, Kasachstan, Kirgisistan, Russland, Tadschikistan, Turkmenistan und Usbekistan verbreitet.[1]
- Oxytropis altaica (Pall.) Pers.: Sie ist in Xinjiang, Kasachstan, in der Mongolei und im asiatischen Teil Russlands verbreitet.[43]
- Oxytropis ambigua (Pall.) DC.: Sie ist in Xinjiang, Kasachstan (nur Pavlodar, Vostochno-Kazakhstanskaya), in der Mongolei (nur Khenti, Khuvsgul) und im russischen Baschkirien, Altai, Gorno-Altaisk, Tuwa, Sacha verbreitet.[43]
- Oxytropis amethystea Arv.-Touv.: Sie kommt in Spanien sowie Frankreich vor.[43]
- Oxytropis ammophila Turcz.: Sie kommt im asiatischen Teil Russlands nur in Chakassien, Krasnoyarsk und Tuva vor.[43]
- Oxytropis ampullata (Pall.) Pers.: Sie ist in Xinjiang, Kasachstan, in der Mongolei und im asiatischen Teil Russlands in Gorno-Altaisk, Chakassien, Krasnoyarsk sowie Tuva verbreitet.[43]
- Oxytropis anaulgensis Pavlov: Sie kommt in Kasachstan nur in Schymkent sowie Taras und in Usbekistan nur in Taschkent vor.[43]
- Oxytropis andersii Vassilcz.: Sie kommt in Afghanistan vor.[43]
- Oxytropis anertii Nakai: Sie kommt in der chinesischen Provinz Jilin und in Korea vor.[1]
- Oxytropis approximata Lessen: Sie kommt in Chelyabinsk, Baschkirien sowie Perm vor.[43]
- Oxytropis arassanica Gontsch.: Sie kommt in Kasachstan nur in Schymkent sowie Dzhambul und in Usbekistan nur in Taschkent vor.[43]
- Oxytropis arbaeviae Vassilcz.: Sie kommt in Kasachstan nur in Dzhambul und in Kirgisistan nur bei Frunse vor.[43]
- Oxytropis arctica R.Br.
- Oxytropis arenae-ripariae Vassilcz.
- Oxytropis argentata (Pall.) Pers. (Syn.: Astragalus argentatus Pallas, Oxytropis argyraea DC., Oxytropis argyrophylla Ledeb., Oxytropis recognita Bunge): Sie kommt in Xinjiang und Sibirien vor.[1]
- Oxytropis armeniaca Sosn. ex Mulk.
- Oxytropis arystangalievii Bajtenov
- Oxytropis aspera Gontsch.
- Oxytropis assadliensis Vassilcz.
- Oxytropis assiensis Vassilcz.: Sie ist in Tibet, Xinjiang, Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan verbreitet.[1]
- Oxytropis asterocarpa Vassilcz.
- Oxytropis astragaloides Boriss.
- Oxytropis atbaschi Saposhn.
- Oxytropis aucheri Boiss.
- Oxytropis aulieatensis Vved.
- Oxytropis auriculata C.W.Chang: Sie gedeiht nur im Grasland von Sichuan.[1]
- Oxytropis austrosachalinensis N.S.Pavlova
- Oxytropis avis Saposhn.
- Oxytropis avisoides P.C.Li: Dieser Endemit gedeiht auf Hügeln in Höhenlagen von 4600 bis 4700 Metern nur in Baxoi in Tibet.[1]
- Oxytropis babatagi Abdusal.
- Oxytropis baburi Vassilcz.
- Oxytropis baicalia (Pall.) Pers.
- Oxytropis baissunensis Vassilcz.
- Oxytropis bajtulinii Kotukhov
- Oxytropis baldshuanica B.Fedtsch.
- Oxytropis bargusinensis Peschkova
- Oxytropis barkolensis X.Y.Zhu, H.Ohashi & Y.B.Deng (Syn.: Oxytropis flavovirens H.Ohba, S.Akiyama & S.K.Wu.): Sie gedeiht auf alpinen Matten, Hügeln, an Rändern von Juniperus centrasiatica-Wäldern, exponierten Grasländern, trockenen Hängen und an Fließgewässern in Höhenlagen von 2000 bis 3400 Metern nur in Xinjiang.[1]
- Oxytropis baxoiensis P.C.Li: Sie gedeiht nur in Höhenlagen von 3900 Metern in Tibet.[1]
- Oxytropis bella B.Fedtsch. (Syn.: Oxytropis trichosphaera Freyn): Sie ist in Tibet, Xinjiang, Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan verbreitet.[1]
- Oxytropis bellii (Britton) Palib.
- Oxytropis beringensis Jurtzev
- Oxytropis besseyi (Rydb.) Blank.
- Oxytropis bicornis Vassilcz.
- Oxytropis bicolor Bunge (Syn.: Oxytropis angustifolia Ulbr.): Sie ist in der Mongolei und in den chinesischen Provinzen Gansu, Hebei, Henan, Nei Mongol, Ningxia, Shaanxi, Shandong sowie Shanxi verbreitet.[1]
- Oxytropis biflora P.C.Li: Sie gedeiht auf alpinen Matten und entlang von Fließgewässern nur in Höhenlagen von etwa 5000 Metern in Tibet.[1]
- Oxytropis biloba Saposhn.
- Oxytropis binaludensis Vassilcz.
- Oxytropis birirensis Ali: Dieser Endemit kommt nur in der pakistanischen Provinz Khyber Pakhtunkhwa vor.[2]
- Oxytropis bobrovii B.Fedtsch.
- Oxytropis bogdoschanica Jurtzev
- Oxytropis boguschi B.Fedtsch.
- Oxytropis borissoviae Polozhij
- Oxytropis bosculensis Golosk.
- Oxytropis bracteata Basil.
- Oxytropis bracteolata Vassilcz.
- Oxytropis brevicaulis Ledeb.
- Oxytropis brevipedunculata P.C.Li: Sie gedeiht auf alpinen Matten und entlang von Fließgewässern nur in Höhenlagen von 5200 bis 5400 Metern in Tibet.[1]
- Oxytropis bungei Kom.
- Oxytropis cabulica (Boiss.) Boiss.
- Oxytropis cachemiriana Cambess. (Syn.: Oxytropis proboscides Bunge): Sie kommt in der pakistanischen Provinz Khyber Pakhtunkhwa, in Kaschmir und im westlichen Tibet vor.[2]
- Oxytropis caerulea (Pall.) DC. (Syn.: Astragalus baicalensis Pall., Astragalus coeruleus Pall., Oxytropis chinensis Bunge, Oxytropis filiformis var. monticola Malyschev, Oxytropis mandshurica Bunge): Sie ist in Russland, in der Mongolei und in den chinesischen Provinzen Gansu, Hebei, Nei Mongol sowie Shanxi verbreitet.[1]
- Oxytropis caespitosa (Pall.) Pers.
- Oxytropis caespitosula Gontsch.
- Oxytropis calcareorum N.S.Pavlova
- Oxytropis callophylla Vassilcz.
- Oxytropis calva Malyschev
- Oxytropis campanulata Vassilcz.
- Feld-Spitzkiel oder Alpen-Spitzkiel (Oxytropis campestris (L.) DC.)
- Oxytropis cana Bunge
- Oxytropis candicans (Pall.) DC.
- Oxytropis canopatula Vassilcz.
- Oxytropis capusii Franch.
- Oxytropis caputoi Moraldo & la Valva
- Oxytropis caraganetorum Vassilcz.
- Oxytropis carpatica R.Uechtr.
- Oxytropis chakassiensis Polozhij
- Oxytropis chantengriensis Vassilcz.
- Oxytropis charkeviczii Vyschin
- Oxytropis chesneyoides Gontsch.
- Oxytropis chinglingensis C.W.Chang (Syn.: Oxytropis humilis C.W.Chang): Sie gedeiht in sonnigen Grasländern und Wiesen an sonnigen Hängen in Höhenlagen von 1800 bis 3900 Metern in Tibet und Shaanxi.[1]
- Oxytropis chionobia Bunge (Syn.: Oxytropis oligantha Bunge): Sie ist in Xinjiang (nur im Tian Shan) und in Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan verbreitet.[1]
- Oxytropis chionophylla Schrenk
- Oxytropis chitralensis Ali: Dieser Endemit kommt nur in der pakistanischen Provinz Khyber Pakhtunkhwa vor.[2]
- Oxytropis chorgossica Vassilcz.
- Oxytropis chrysocarpa Boiss.
- Oxytropis ciliata Turcz.: Sie ist in der Mongolei und in den chinesischen Provinzen Hebei, Nei Mongol sowie Ningxia verbreitet.[1]
- Oxytropis cinerascens Bunge: Sie kommt in Indien und Tibet vor.[1]
- Oxytropis cinerea Vassilcz.
- Oxytropis coelestis Abdusal.
- Oxytropis coerulea (Pall.) DC.
- Oxytropis collettii Fisch.
- Oxytropis columbina Vassilcz.
- Oxytropis confusa Bunge
- Oxytropis crassiuscula Boriss.: Sie kommt im pakistanischen Chitral und im Alaigebirge vor.[2]
- Oxytropis cretacea Basil.
- Oxytropis cuspidata Bunge
- Oxytropis czapandaghi B.Fedtsch.
- Oxytropis czekanowskii Jurtzev
- Oxytropis czerskii Jurtzev
- Oxytropis czukotica Jurtzev
- Oxytropis danorum Rech. f.
- Oxytropis darpirensis Jurtzev & A.P.Khokhr.
- Oxytropis dashtinavarensis Vassilcz.
- Oxytropis dasypoda Boiss.
- Oxytropis deflexa (Pall.) DC.: Sie ist in Nordamerika, Russland, in der Mongolei, Tibet und in den chinesischen Provinzen Nei Mongol, Ningxia, Qinghai sowie in Xinjiang weitverbreitet.[1]
- Oxytropis densa Bunge (Syn.: Oxytropis stipulosa Kom.): Sie ist in Kaschmir, in der pakistanischen Provinz Khyber Pakhtunkhwa, Tibet und in den chinesischen Provinzen Gansu, Qinghai sowie in Xinjiang verbreitet.[1][2]
- Oxytropis densiflora P.C.Li: Die seit 2000 zwei Varietäten gedeihen an Hängen und in Grasländern in Höhenlagen von 3200 bis 4000 Metern in Tibet und Gansu.[1]
- Oxytropis diantha Maxim.
- Oxytropis dichroantha Schrenk (Syn.: Oxytropis algida Bunge): Sie ist in Xinjiang, Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan verbreitet.[1]
- Oxytropis didymophysa Bunge
- Dinarischer Spitzkiel (Oxytropis dinarica (Murb.) Wettst.)
- Oxytropis diversifolia E.Peter: Sie kommt in der Mongolei und in der Inneren Mongolei vor.[1]
- Oxytropis dorogostajskyi Kuzen.
- Oxytropis dubia Turcz.
- Oxytropis duthieana Ali
- Oxytropis echidna Vved.
- Oxytropis erecta Kom.
- Oxytropis eriocarpa Bunge (Syn.: Oxytropis komei Saposhn.): Sie ist in Xinjiang, Russland und in der Mongolei verbreitet.[1]
- Oxytropis ervicarpa Vved. ex Filim.
- Oxytropis evenorum Jurtzev & A.P.Khokhr.
- Oxytropis exserta Jurtzev
- Oxytropis falcata Bunge (Syn.: Oxytropis hedinii Ulbr., Oxytropis holdereri Ulbr., Oxytropis popovii Vassilcz.): Sie gedeiht in Höhenlagen von 2700 bis 5200 Metern in den chinesischen Provinzen Gansu, Ningxia, Qinghai, Sichuan, Xinjiang sowie in Tibet.[1]
- Oxytropis farsi Vassilcz.
- Oxytropis fasciculiflorum Vassilcz.
- Oxytropis fedtschenkoana Vassilcz.
- Oxytropis ferganensis Vassilcz.
- Drüsiger Spitzkiel (Oxytropis fetida (Vill.) DC.)
- Oxytropis fetisowi Bunge
- Oxytropis fetisowii Bunge
- Oxytropis filiformis DC. (Syn.: Oxytropis caerulea (Pall.) Turcz. non Oxytropis caerulea (Pall.) DC.): Sie ist in Russland, in der Mongolei und in der Inneren Mongolei verbreitet.[1]
- Oxytropis floribunda (Pall.) DC.
- Oxytropis fohlenensis Vassilcz.
- Oxytropis fominii Grossh.
- Oxytropis foucaudii Gillot
- Oxytropis fragilifolia N.Ulziykh.
- Oxytropis fragiliphylla Q.Wang, ChangY.Yang, X.Y.Zhu & H.Ohashi: Sie wurde 2001 erstbeschrieben. Sie gedeiht auf subalpinen Matten in Höhenlagen von 1800 bis 2800 Metern Xinjiang.[1]
- Oxytropis fruticulosa Bunge
- Oxytropis fuscescens Vassilcz.
- Oxytropis ganningensis C.W.Chang: Sie gedeiht in Tälern und auf Grasländern an Hängen in Höhenlagen von 1100 bis 1200 Metern in den chinesischen Provinzen Gansu sowie Ningxia.[1]
- Oxytropis gebleri Bunge
- Oxytropis gebleriana Schrenk
- Oxytropis gerzeensis P.C.Li: Sie gedeiht auf Wiesen in Höhenlagen von 3400 bis 5200 Metern in den chinesischen Provinzen Qinghai, Xinjiang sowie in Tibet.[1]
- Oxytropis gilgitensis Vassilcz.
- Oxytropis giraldii Ulbr. (Syn.: Oxytropis ningxiaensis C.W.Chang.): Sie gedeiht in Höhenlagen von 2100 bis 3600 Metern in den chinesischen Provinzen Gansu, Ningxia, Qinghai, Shaanxi sowie Sichuan.[1]
- Oxytropis glabra DC. (Syn.: Oxytropis diffusa Ledeb., Oxytropis drakeana Franch., Oxytropis glareosa Vassilcz., Oxytropis salina Vassilcz., Oxytropis tenuis Palib.): Sie ist in Kasachstan, Russland, in der Mongolei, in der Inneren Mongolei, in Tibet und in den chinesischen Provinzen Gansu, Hebei, Henan, Jilin, Ningxia, Qinghai, Shaanxi, Shanxi sowie Xinjiang verbreitet.[1]
- Oxytropis glandulosa Turcz.
- Oxytropis globiflora Bunge: Sie ist in Tibet, Xinjiang, Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan verbreitet.[1]
- Oxytropis gloriosa Ali: Dieser Endemit kommt nur in der pakistanischen Provinz Chitral vor.[2]
- Oxytropis gmelinii Boriss.
- Oxytropis golengolensis Vassilcz.
- Oxytropis gorbunovii Boriss.
- Oxytropis gracillima Vassilcz. non Bunge
- Oxytropis graminetorum Vassilcz.
- Oxytropis grandiflora (Pall.) DC. (Syn.: Oxytropis collina Turcz.): Sie ist in Sibirien, in der Mongolei, in der Inneren Mongolei und in den chinesischen Provinzen Hebei, Jilin sowie Ningxia verbreitet.[1]
- Oxytropis griffithii Boiss.
- Oxytropis gubanovii Vassilcz.
- Oxytropis gueldenstaedtioides Ulbr.: Sie gedeiht in Grasländern an Hängen in den chinesischen Provinzen Gansu sowie Shaanxi.[1]
- Oxytropis guilanica Maassoumi & Moradi: Sie wurde 2015 erstbeschrieben und wurde bisher nur im westlich Elburs im nordwestlichen Iran gefunden. Sie gedeiht in Höhenlagen von 2400 bis 2700 Metern in schneereichen, Nord-Ost exponierten Lagen in alpinen Felssteppen.[45]
- Oxytropis guinanensis Y.H.Wu: Dieser Endemit gedeiht in Wüsten in Höhenlagen von etwa 3200 Metern nur im Kreis Guinan in der chinesischen Provinz Qinghai.[1]
- Oxytropis guntensis B.Fedtsch.
- Oxytropis gymnogyne Bunge
- Hallers Spitzkiel, Seidenhaar-Spitzkiel (Oxytropis halleri Koch, Syn.: Oxytropis xerophila Gutermann, Oxytropis halleri subsp. velutina (Schur) O.Schwarz, Oxytropis velutina Schur, Oxytropis halleri subsp. villoso-sericea (Shuttlew.) Bech., Oxytropis halleri subsp. korabensis (Kümmerle & Jáv.) Chrtek & Chrtková)
- Oxytropis hedgei Vassilcz.
- Oxytropis helenae N.S.Pavlova
- Schweizer Spitzkiel (Oxytropis helvetica Scheele)
- Oxytropis heratensis Bunge
- Oxytropis heterophylla Maxim.
- Oxytropis heteropoda Bunge
- Oxytropis heterotricha Turcz.
- Oxytropis hidakamontana Miyabe & Tatew.
- Oxytropis hindukushensis Vassilcz.
- Oxytropis hippolyti Boriss.
- Oxytropis hirsuta Bunge: Sie ist in Xinjiang, Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan, Usbekistan, in der Mongolei und im westlichen Sibirien verbreitet.[1]
- Oxytropis hirsutiuscula Freyn: Sie ist in Qinghai, Xinjiang, Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan, Usbekistan und im westlichen Sibirien verbreitet.[1]
- Oxytropis hirta Bunge (Syn.: Oxytropis komarovii Vassilcz.): Sie ist in Russland, in der Mongolei, in der Inneren Mongolei und in den chinesischen Provinzen Gansu, Hebei, Heilongjiang, Henan, Jilin, Liaoning, Shaanxi, Shandong sowie Shanxi verbreitet.[1]
- Oxytropis holanshanensis H.C.Fu: Sie gedeiht in Tälern und an steinigen Hängen in Höhenlagen von 2000 bis 2400 Metern in den chinesischen Provinzen Ningxia und in der südwestlichen Innere Mongolei (nur im Helan Shan).[1]
- Oxytropis humifusa Kar. & Kir. (Syn.: Oxytropis albana Steven, Oxytropis glacialis Strach. & Winterb. ex Bunge, Oxytropis melanotricha Bunge): Sie ist in Xinjiang, Tibet, Indien, Kaschmir, in der pakistanischen Provinz Khyber Pakhtunkhwa, Nepal, Afghanistan, Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan verbreitet.[1][2]
- Oxytropis hypoglottoides (Baker) Ali: Sie kommt in der pakistanischen Provinz Chitral und in Turkistan vor.[2]
- Oxytropis hypsophila Bunge
- Oxytropis hystrix Schrenk (Syn.: Oxytropis spinifer Vassilcz.): Sie kommt in Xinjiang und Kasachstan vor.[1]
- Oxytropis imbricata Kom.
- Oxytropis immersa (Baker) B.Fedtsch. (Syn.: Oxytropis incanescens Freyn, Oxytropis pamirica Danguy): Sie ist mit zwei Varietäten in Xinjiang, Tibet, im Iran, in den pakistanischen Provinzen Chitral sowie Khyber Pakhtunkhwa, Afghanistan, Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan und russischen Tian Shan sowie im Alaigebirge verbreitet.[2][1]
- Oxytropis inaria (Pall.) DC.
- Oxytropis incana Jurtzev
- Oxytropis includens Basil.
- Oxytropis indensis Vassilcz.
- Oxytropis inopinata Jurtzev
- Oxytropis inschanica H.C.Fu & S.H.Cheng: Sie gedeiht an Trockenhängen und kiesigen Gebieten in Tälern in Höhenlagen von 1800 bis 2100 Metern in der Inneren Mongolei.[1]
- Oxytropis integripetala Bunge
- Oxytropis intermedia Bunge
- Oxytropis interposita Sipliv.
- Oxytropis iranica Vassilcz.
- Oxytropis iridum Dickoré & M.Kriechb.
- Oxytropis iskanderica B.Fedtsch.
- Oxytropis itoana Tatew.
- Oxytropis jabalambrensis (Pau) Podlech
- Oxytropis japonica Maxim.
- Oxytropis jonesii Barneby
- Oxytropis jucunda Vved.
- Oxytropis jurtzevii Malyschev
- Oxytropis kamelinii Vassilcz.
- Oxytropis kamtschatica Hulten
- Oxytropis kansuensis Bunge (Syn.: Oxytropis leucocephala Ulbr., Oxytropis longipedunculata C.W.Chang, Oxytropis thionantha Ulbr.): Sie kommt in Nepal, Tibet und in den chinesischen Provinzen Gansu, Qinghai sowie Sichuan vor.[1]
- Oxytropis karataviensis Pavlov
- Oxytropis karavaevii Jurtzev
- Oxytropis karjaginii Grossh.
- Oxytropis kaspensis Krasnob. & Pshenich.
- Oxytropis katangensis Basil.
- Oxytropis kateninii Jurtzev
- Oxytropis kazidanica Vassilcz.
- Oxytropis ketmenica Saposhn.
- Oxytropis khinjahi Vassilcz.
- Oxytropis klementzii N.Ulziykh.
- Oxytropis kodarensis Jurtzev & Malyschev
- Oxytropis kokrinensis A.E.Porsild
- Oxytropis komarovii Vassilcz.
- Oxytropis kopetdagensis Gontsch.
- Oxytropis kopetdaghensis Gontsch.
- Oxytropis kotschyana Boiss. & Hohen.
- Oxytropis krylovi Shipcz.
- Oxytropis krylovii Schipcz.: Sie kommt in Xinjiang, in Kasachstan und im westlichen Sibirien vor.[1]
- Oxytropis kubanensis Leskov
- Oxytropis kuchanensis Vassilcz.
- Oxytropis kuhistanica Abdusal.
- Oxytropis kukkonenii Vassilcz.
- Oxytropis kunarensis Vassilcz.
- Oxytropis kunashiriensis Kitam.
- Oxytropis kuramensis Abdusal.
- Oxytropis kusnetzovii Krylov & Steinb.
- Oxytropis kyziltalensis Vassilcz.
- Oxytropis ladyginii Krylov: Sie kommt in Xinjiang, in der Mongolei und in Russland vor.[1]
- Oxytropis lagopus Nutt.
- Oxytropis lambertii Pursh
- Oxytropis lanata (Pall.) DC.: Sie kommt in der Inneren Mongolei und in Sibirien vor.[1]
- Oxytropis lanceatifoliola H.Ohba, S.Akiyama & S.K.Wu: Sie wurde 2000 erstbeschrieben. Dieser Endemit gedeiht an trockenen, alluvialen Hängen in einer Höhenlage von etwa 4100 Meter nur in Ruoqiang in Xinjiang.[1]
- Oxytropis langshanica H.C.Fu: Sie gedeiht in Sandwüsten in der Inneren Mongolei.[1]
- Oxytropis lanuginosa Kom.
- Lappland-Spitzkiel (Oxytropis lapponica (Wahlenb.) Gay, Syn.: Oxytropis amoena Kar. & Kir., Oxytropis carinthiaca Fisch.-Oost., Oxytropis lapponica (Wahlenb.) Gaudin Oxytropis thomasii Gaudin): Er ist im indischen Bundesstaat Punjab, in Kaschmir, Nepal, in der pakistanischen Provinz Khyber Pakhtunkhwa, in Tibet, in den chinesischen Provinzen Shaanxi sowie Xinjiang, Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan, Usbekistan, Russland, Norwegen, Schweden, in der Schweiz, in Österreich, Ungarn, Italien und in Spanien weitverbreitet.[1][2]
- Oxytropis lasiocarpa Gontsch.
- Oxytropis lasiopoda Bunge
- Oxytropis latialata P.C.Li: Dieser Endemit gedeiht an Berghängen in Höhenlagen von etwa 5100 Metern nur in Baingoin in Tibet.[1]
- Oxytropis latibracteata Jurtzev: Von den zwei Varietäten ist die eine ein Endemit in Gonghe in Qinghai und die andere gedeiht in Höhenlagen von 1700 bis 3800 Metern in Tibet und in den chinesischen Provinzen Innere Mongolei, Gansu, Hebei, Ningxia, Qinghai, Shaanxi, Sichuan sowie Xinjiang.[1]
- Oxytropis lavrenkoi N.Ulziykh.
- Oxytropis laxiracemosa Vassilcz.
- Oxytropis lazica Boiss.
- Oxytropis lehmanni Bunge (Syn.: Oxytropis aequipetala Bunge): Sie kommt in Tibet, Tadschikistan sowie Usbekistan vor.[1]
- Oxytropis leptophylla (Pall.) DC.: Die zwei Varietäten sind in Russland, in der Mongolei, Innere Mongolei und in den chinesischen Provinzen Hebei, Jilin sowie Shanxi verbreitet.[1]
- Oxytropis leptophysa Bunge
- Oxytropis leucantha (Pall.) Bunge
- Oxytropis leucocyanea Bunge
- Oxytropis leucotricha Turcz.
- Oxytropis lhasaensis X.Y.Zhu: Sie wurde 2004 erstbeschrieben. Dieser Endemit gedeiht an sandigen Ufern von Fließgewässern in Höhenlagen von etwa 3700 Metern nur in Lhasa in Tibet.[1]
- Oxytropis liliputa Vassilcz.
- Oxytropis linczevskii Gontsch.
- Oxytropis linearibracteata P.C.Li: Dieser Endemit gedeiht an Trockenhängen und Schwemmkegeln in Höhenlagen von etwa 4200 Metern nur im Sog Xian in Tibet.[1]
- Oxytropis lipskyi Gontsch.
- Oxytropis lithophila Vassilcz.
- Oxytropis litwinowii B.Fedtsch.
- Oxytropis longialata P.C.Li: Dieser Endemit gedeiht an sandigen und kiesigen Standorten in Tälern in Höhenlagen von 4000 bis 4100 Metern nur in Lhari in Tibet.[1]
- Oxytropis longibracteata Kar. & Kir.
- Oxytropis longipedunculata C.W.Chang
- Oxytropis longirostra DC.
- Oxytropis lupinoides Grossh.
- Oxytropis luteo-coerulea (Baker) Ali: Dieser Endemit kommt nur an der Grenze von Pakistan und Afghanistan im Kurram-Tal vor.[2]
- Oxytropis lydiae Vassilcz.
- Oxytropis macrobotrys Bunge
- Oxytropis macrocarpa Kar. & Kir.
- Oxytropis macrodonta Gontsch.
- Oxytropis macrosema Bunge
- Oxytropis maduoensis Y.H.Wu: Dieser Endemit gedeiht auf alpinen Matten an Hängen in Höhenlagen von 4300 bis 4600 Metern nur in Madoi in Qinghai.[1]
- Oxytropis maidantalensis B.Fedtsch.
- Oxytropis malacophylla Bunge
- Oxytropis malloryana Dunn: Sie gedeiht an steinigen Südhängen in Höhenlagen von 3800 bis 4600 Metern in Tibet.[1]
- Oxytropis maqinensis Y.H.Wu: Sie gedeiht in kiesigen Gebieten, an sonnigen Hängen und auf alpinen Matten in Höhenlagen von 3300 bis 4500 Metern in Qinghai.[1]
- Oxytropis marco-poloi Vassilcz.
- Oxytropis margacea Vassilcz.
- Oxytropis martjanovi Krylov
- Oxytropis masanderanensis Vassilcz.
- Oxytropis masarensis Vassilcz.
- Oxytropis maydelliana Trautv.
- Oxytropis megalantha H.Boissieu
- Oxytropis megalorrhyncha Nevski
- Oxytropis meinshausenii Schrenk: Sie ist in Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan und in den chinesischen Provinzen Gansu, Sichuan sowie Xinjiang verbreitet.[1]
- Oxytropis melaleuca Bunge
- Oxytropis melanocalyx Bunge: Sie gedeiht in Höhenlagen von 2200 bis 5100 Metern in den chinesischen Provinzen Gansu, Qinghai, Shaanxi, Sichuan, Yunnan, Xinjiang, Innere Mongolei sowie Tibet.[1]
- Oxytropis merkensis Bunge (Syn.: Oxytropis kanitzii N.D.Simpson): Sie ist in Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan und in den chinesischen Provinzen Gansu, Ningxia, Qinghai, Xinjiang, Innere Mongolei sowie Tibet verbreitet.[1]
- Oxytropis mertensiana Turcz.
- Oxytropis michelsonii B.Fedtsch.
- Oxytropis micrantha Bunge
- Oxytropis microcarpa Gontsch.
- Oxytropis microphylla (Pall.) DC. (Syn.: Oxytropis chiliophylla Royle ex Benth., Oxytropis grenardi Franch., Oxytropis ingrata Freyn, Oxytropis polyadenia Freyn, Oxytropis tibetica Bunge): Sie ist im nördlichen Teil des indischen Bundesstaates Punjab, Nepal, im russischen Dauria, im Alaigebirge, Kaschmir, im indischen Punjab, Afghanistan, Kirgisistan, Tadschikistan, in der Mongolei, in der inneren Mongolei, in Tibet und in den chinesischen Provinzen Gansu, Qinghai sowie Xinjiang verbreitet.[1][2]
- Oxytropis microsphaera Bunge
- Oxytropis middendorffii Trautv.
- Oxytropis minjanensis Rech. f.
- Oxytropis mixotriche Bunge
- Oxytropis moellendorffii Bunge ex Maxim. (Syn.: Oxytropis limprichtii Ulbr., Oxytropis schensiensis Kom., Oxytropis sylinchanensis Franch.): Sie gedeiht an Straßenrändern an Hügeln und kiesigen oberen Bereichen von Hügeln in Höhenlagen von 2400 bis 3400 Metern in den chinesischen Provinzen Hebei sowie Shanxi.[1]
- Oxytropis mollis Royle ex Benth. (Syn.: Oxytropis thomsonii Benth. ex Bunge, Oxytropis iridum Dickore & Kriechb.): Sie kommt im nördlichen Teil des indischen Bundesstaates Punjab, in Tibet, Nepal, Kaschmir und in der pakistanischen Provinz Khyber Pakhtunkhwa vor.[1][2]
- Oxytropis mongolica Kom.
- Berg-Spitzkiel (Oxytropis montana (L.) DC., er wurde nomenklatorisch verworfen, da nicht bekannt war, welche Art Linné ursprünglich gemeint hat und es wurde von 1961 bis 2009 das Synonym Oxytropis jacquinii Bunge verwendet.)
- Oxytropis monophylla Grubov: Sie kommt in der Mongolei, in der Inneren Mongolei sowie in Ningxia vor.[1]
- Oxytropis morenarum Vassilcz.
- Oxytropis multiceps Torr. & A.Gray
- Oxytropis multiramosa P.C.Li
- Oxytropis mumynabadensis B.Fedtsch.
- Oxytropis muricata (Pall.) DC.: Sie kommt in Sibirien, in der nördlichen Mongolei sowie in Ningxia vor.[1]
- Oxytropis myriophylla (Pall.) DC.: Sie ist in Russland, in der Mongolei, in der Inneren Mongolei und in den chinesischen Provinzen Gansu, Hebei, Heilongjiang, Jilin, Liaoning, Ningxia sowie Shanxi verbreitet.[1]
- Oxytropis nana Torr. & A.Gray
- Pyrenäen-Spitzkiel (Oxytropis neglecta Ten.)
- Oxytropis neimongolica C.W.Chang & Y.Z.Zhao: Sie gedeiht an sonnigen Hängen, Grasländern, Nordhängen, Stipa-Gesellschaften auf trockenen wüstenhaften Prairien in Höhenlagen von 1000 bis 2200 Metern in der Inneren Mongolei.[1]
- Oxytropis neo-rechingeriana Vassilcz.
- Oxytropis nepalensis Vassilcz.
- Oxytropis niedzweckiana Popov
- Oxytropis nigrescens (Pall.) DC.
- Oxytropis nikolai Filim. & Abdusal.
- Oxytropis nitens Turcz.
- Oxytropis nivea Bunge
- Oxytropis nuda Basil.
- Oxytropis nuristanica Vassilcz.
- Oxytropis nutans Bunge (Syn.: Oxytropis qiemoensis H.Ohba, S.Akiyama & S.K.Wu): Sie ist in Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan sowie Xinjiang verbreitet.[1]
- Oxytropis obnapiformis C.L.Porter
- Oxytropis ochotensis Bunge
- Oxytropis ochrantha Turcz. (Syn.: Oxytropis chrysotricha Franch., Oxytropis daqingshanica Y.Z.Zhao & ZongY.Zhu, Oxytropis turczaninovii Jurtzev): Sie ist in Russland, in der Mongolei, in der Inneren Mongolei, Tibet und in den chinesischen Provinzen Gansu, Hebei, Ningxia, Qinghai, Shanxi, Sichuan sowie Xinjiang verbreitet.[1]
- Oxytropis ochrocephala Bunge: Sie gedeiht an verunkrauteten Hängen, Grasländern und alpinen Matten in Höhenlagen von 1800 bis 4500 Metern in der Inneren Mongolei, Tibet und in den chinesischen Provinzen Gansu, Hebei, Ningxia, Qinghai, Sichuan sowie Xinjiang.[1]
- Oxytropis ochroleuca Bunge: Sie ist in Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan sowie Xinjiang verbreitet.[1]
- Oxytropis ochrolongibracteata X.Y.Zhu & H.Ohashi: Die Erstbeschreibung erfolgte 2000. Sie gedeiht an Hügeln, Straßenrändern, in trockenen Tälern, an Ufern von Fließgewässern, auf alpinen Matten und zwischen Sträuchern in Höhenlagen von 1700 bis 4300 Metern in Tibet und Gansu.[1]
- Oxytropis ocrensis F.Conti & Bartolucci: aus dem zentralen Apenninen vom Monte Ocre beschrieben.[46]
- Oxytropis oreophila A.Gray
- Oxytropis ornata Vassilcz.
- Oxytropis ovczinnikovii Abdusal.
- Oxytropis owerinii Bunge
- Oxytropis oxyphylla (Pall.) DC. (Syn.: Oxytropis andaensis P.H.Huang & L.H.Zhuo, Oxytropis arenaria Jurtzev, Oxytropis chankaensis Jurtzev, Oxytropis hailarensis Kitag., Oxytropis hulunbailensis H.C.Fu & Cheng f., Oxytropis lanata var. psilocarpa Kitag., Oxytropis selengensis Bunge, Oxytropis verticillaris Ledeb.): Sie kommt in Korea, in der Inneren Mongolei und in den chinesischen Provinzen Gansu, Heilongjiang, Jilin, Liaoning, Qinghai sowie Shaanxi vor.[1]
- Oxytropis oxyphylloides Popov
- Oxytropis pagobia Bunge: Sie gedeiht in Höhenlagen von 2100 bis 3800 Metern in Tibet.[1]
- Oxytropis pakistanica Vassilcz.
- Oxytropis pallasii Pers.
- Oxytropis pamiroalaica Abdusal.
- Oxytropis panjshinica Podlech & I.Deml
- Oxytropis parasericeopetala P.C.Li: Dieser Endemit gedeiht im Grasland in Tälern in Höhenlagen von 4500 bis 5000 Metern nur in Lhasa in Tibet.[1]
- Oxytropis parryi A.Gray
- Oxytropis parvanensis Vassilcz.
- Oxytropis pauciflora Bunge (Syn.: Oxytropis alpicola Bunge, Oxytropis friabilis H.Ohba, S.Akiyama & S.K.Wu, Oxytropis tschujae Bunge): Sie kommt in Kasachstan, Russland, Tibet und in den chinesischen Provinzen Gansu sowie Xinjiang vor.[1]
- Oxytropis pavlovii B.Fedtsch. & Basil.
- Oxytropis pellita Bunge
- Oxytropis penduliflora Gontsch.: Sie ist in Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan, Xinjiang sowie Qinghai verbreitet.[1]
- Oxytropis persica Boiss.
- Oxytropis peschkovae Popov
- Oxytropis physocarpa Ledeb.
- Oxytropis piceetorum Vassilcz.
- Zottiger Spitzkiel (Oxytropis pilosa (L.) DC.): Er ist in Europa, Kasachstan, Russland, in der Mongolei sowie in Xinjiang verbreitet.[1]
- Oxytropis pilosissima Vved.
- Oxytropis platonychia Bunge (Syn.: Oxytropis kuhazensis Vass.): Sie kommt in der pakistanischen Provinz Khyber Pakhtunkhwa und im russischen Tian Shan sowie im Alaigebirge vor.[2]
- Oxytropis platysema Schrenk: Sie ist in Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan, Tibet sowie Xinjiang verbreitet.[1]
- Oxytropis podlechii Vassilcz.
- Oxytropis podocarpa A.Gray
- Oxytropis podoloba Kar. & Kir. (Syn.: Oxytropis brachybotrys Bunge): Sie ist in Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan sowie Xinjiang verbreitet.[1]
- Oxytropis politovii Sumnev.
- Oxytropis polyphylla Ledeb.
- Oxytropis poncinsii Franch. (Syn.: Oxytropis introflexa Freyn): Sie ist in Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan, Tibet, Xinjiang sowie Gansu verbreitet.[1]
- Oxytropis popoviana Peschkova
- Oxytropis potaninii Palib.
- Prenj-Spitzkiel (Oxytropis prenja (Beck) Beck)
- Oxytropis proboscidea Bunge (Syn.: Oxytropis glacialis Benth. ex Bunge, Oxytropis nivalis Franch.): Sie gedeiht in Höhenlagen von 4100 bis 5300 Metern in Tibet und in den chinesischen Provinzen Gansu, Yunnan sowie in Xinjiang.[1]
- Oxytropis prostrata (Pall.) DC.
- Oxytropis protopopovii Kom.
- Oxytropis proxima Boriss.
- Oxytropis przewalskii Kom.: Sie gedeiht an sonnigen Hängen nur in Xinjiang.[1]
- Oxytropis pseudocoerulea P.C.Li: Sie gedeiht in Grasländern und an strachbestandenen Hängen in Höhenlagen von 2000 bis 3800 Metern in Tibet und in den chinesischen Provinzen Sichuan sowie in Xinjiang.[1]
- Oxytropis pseudofrigida Saposhn.: Sie gedeiht auf alpinen Matten zwischen Sträuchern in Höhenlagen von etwa 1400 Metern 1400 in Xinjiang.[1]
- Oxytropis pseudoglandulosa Gontscharov ex Grubov: Sie gedeiht an sandigen Standorten entlang des Qinghai-See in Höhenlagen von 3000 bis 3100 Metern in Qinghai.[1]
- Oxytropis pseudohirsuta Q.Wang & Chang Y.Yang: Die Erstbeschreibung erfolgte 2001. Sie gedeiht in Grasländern an Hängen in Höhenlagen von 700 bis 1700 Metern in Xinjiang.[1]
- Oxytropis pseudohirsutiuscula Vassilcz.
- Oxytropis pseudoleptophysa Boriss.
- Oxytropis pseudomyriophylla S.H.Cheng ex X.Y.Zhu, H.Ohashi & Y.B.Deng: Sie gedeiht an Hügeln und Straßenrändern entlang von Feldern in Höhenlagen von 1400 bis 2600 Metern in den chinesischen Provinzen Gansu, Ningxia sowie Shanxi.[1]
- Oxytropis pseudorosea Filim.
- Oxytropis puberula Boriss.
- Oxytropis pulvinoides Vassilcz.
- Oxytropis pumila Fisch. ex DC.: Sie kommt in Xinjiang vor.[1]
- Oxytropis pumilio (Pall.) Ledeb.
- Oxytropis purpurea (Bald.) Markgr.
- Oxytropis pusilla Bunge: Sie gedeiht in Höhenlagen von 3700 bis 5000 Metern in Tibet und Xinjiang.[1]
- Oxytropis pusilloides Vassilcz.
- Oxytropis putoranica M.M.Ivanova
- Oxytropis qamdoensis X.Y.Zhu, Y.F.Du & H.Ohashi: Die Erstbeschreibung erfolgte 2002. Sie gedeiht zwischen Sträuchern an trockenen und steinigen Hängen in Höhenlagen von 3200 bis 3300 Metern in Tibet.[1]
- Oxytropis qilianshanica C.W.Chang & C.L.Zhang ex X.Y.Zhu & H.Ohashi: Die Erstbeschreibung erfolgte 2000. Sie gedeiht in Höhenlagen von 2300 bis 5100 Metern in Tibet und in den chinesischen Provinzen Gansu sowie Qinghai.[1]
- Oxytropis qinghaiensis Y.H.Wu (Syn.: Oxytropis qinghaiensis Y.H.Wu): Sie gedeiht in Höhenlagen von 3400 bis 4700 Metern in Qinghai.[1]
- Oxytropis qingnanensis Y.H.Wu (Syn.: Oxytropis nangqianensis X.Y.Zhu nom. illeg. superfl.): Sie gedeiht in Wäldern in fast alpinen Gebieten in Höhenlagen von 3900 bis 4100 Metern in Qinghai.[1]
- Oxytropis qitaiensis X.Y.Zhu, H.Ohashi & Y.B.Deng: Sie gedeiht in Wiesen und an Hängen in Höhenlagen von 1900 bis 2400 Metern in Xinjiang.[1]
- Oxytropis racemosa Turcz. (Syn.: Oxytropis acutirostrata Ulbr., Oxytropis gracillima Bunge non Vassilcz., Oxytropis koreana Nakai, Oxytropis psammocharis Hance): Sie ist Korea, in der Mongolei, in der Inneren Mongolei und in den chinesischen Provinzen Gansu, Hebei, Henan, Liaoning, Ningxia, Shaanxi sowie Shanxi verbreitet.[1]
- Oxytropis ramosissima Kom.: Sie gedeiht auf aktiven sowie teilweise stabilisierten Sanddünen and sandigen Hängen in Höhenlagen von 900 bis 1400 Metern in der Inneren Mongolei und in den chinesischen Provinzen Gansu sowie Shaanxi.[1]
- Oxytropis rechingeri Vassilcz.
- Oxytropis reniformis P.C.Li: Sie gedeiht auf Grasländern an Berghängen in Höhenlagen von 4300 bis 4600 Metern in Tibet.[1]
- Oxytropis retusa Matsum.
- Oxytropis reverdattoi Jurtzev
- Oxytropis revoluta Ledeb.
- Oxytropis rhizantha Palib.
- Oxytropis rhodontha Vassilcz.
- Oxytropis rhynchophysa Schrenk
- Oxytropis riparia Litv.
- Oxytropis rosea Bunge
- Oxytropis roseiformis B.Fedtsch.
- Oxytropis rostrata Vassilcz.
- Oxytropis rubriargillosa Vassilcz.
- Oxytropis rubricaudex Hulten
- Oxytropis rudbariensis Vassilcz. Nach Massoumi (2012) "doubtful species"[47]
- Oxytropis ruebsaamenii B.Fedtsch.
- Oxytropis rupifraga Bunge: Sie kommt in Xinjiang sowie Kirgisistan vor.[1]
- Oxytropis ruthenica Vassilcz.
- Oxytropis sacciformis H.C.Fu: Dieser Endemit gedeiht an Hängen von Sanddünen nur in Ulanqab Meng in der Inneren Mongolei.[1]
- Oxytropis sachalinensis Miyabe & Tatew.
- Oxytropis sajanensis Jurtzev
- Oxytropis salangensis Podlech & I.Deml
- Oxytropis salicetorum Vassilcz.
- Oxytropis saperlebulensis Vassilcz.
- Oxytropis saposhnikovii Krylov: Sie gedeiht auf gefrorenen Standorten an steinigen alpinen Hängen in Xinjiang.[1]
- Oxytropis sarkandensis Vassilcz.
- Oxytropis sata-kandaonensis Vassilcz.
- Oxytropis satpaevii Bajtenov
- Oxytropis saurica Saposhn.
- Oxytropis savellanica Boiss. ex Boiss. (Syn.: Oxytropis carduchorum Hedge): Sie ist im Iran, Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan und Usbekistan, südlichen Transkaukasien, im russischen Tian Shan sowie im Alaigebirge, in Kaschmir, in der pakistanischen Provinz Chitral, in Tibet sowie in der chinesischen Provinz Qinghai weitverbreitet.[1][2]
- Oxytropis scabrida Gontsch.
- Oxytropis scammaniana Hulten
- Oxytropis schachimardanica Filim.
- Oxytropis scheludjakovae Karav. & Jurtzev
- Oxytropis schmorgunoviae Jurtzev
- Oxytropis schrenkii Trautv.
- Oxytropis semenowii Bunge
- Oxytropis semiglobosa Jurtzev
- Oxytropis seravschanica Gontsch.
- Oxytropis sericea Torr. & A.Gray
- Oxytropis sericopetala C.E.C.Fisch.: Sie gedeiht an sandigen Standorten, Sanddünen, auf Hügeln, in Grasländern, an überfluteten kiesigen Standorten in Höhenlagen von 2600 bis 4600 Metern in Tibet.[1]
- Oxytropis setosa (Pall.) DC.
- Oxytropis sewerzowii Bunge
- Oxytropis shanxiensis X.Y.Zhu: Die Erstbeschreibung erfolgte 2005. Sie gedeiht in Grasländern auf Hügeln in den chinesischen Provinzen Shandong sowie Shanxi.[1]
- Oxytropis shirkuhi Vassilcz.
- Oxytropis shokanbetsuensis Miyabe & Tatew.
- Oxytropis siah-sangi Vassilcz.
- Oxytropis sichuanica C.W.Chang: Sie gedeiht an Waldrändern, in Grasländern auf Bergkämmen in Höhenlagen von 3900 bis 4200 Metern in Sichuan.[1]
- Oxytropis sikaramensis (Sirjaev & Rechinger f.) Ali: Dieser Endemit kommt nur im pakistanischen Teil des Kurram-Tal vor.[2]
- Oxytropis sinkiangensis C.W.Chang: Sie gedeiht im Ödland in Höhenlagen von 500 bis 1000 Metern in den chinesischen Provinzen Gansu sowie Xinjiang.[1]
- Oxytropis siomensis Abdusal.
- Oxytropis sitaipaiensis C.W.Chang: Sie kommt seit 2000 mit zwei Varietäten auf kiesigen Talböden und an Hängen in Höhenlagen von 800 bis 2000 Metern in Shaanxi vor.[1]
- Oxytropis siziwangensis Y.Z.Zhao & Zong Y.Zhu: Dieser Endemit gedeiht auf sandigen Standorten nur in Ulanqab Meng in der Inneren Mongolei.[1]
- Oxytropis sojakii Vassilcz.
- Oxytropis songorica (Pall.) DC.: Sie gedeiht in Grasländern, steinigen Trockenhängen, alpinen bis subalpinen Matten in Höhenlagen von 1300 bis 2800 Metern in Xinjiang.[1]
- Oxytropis spicata (Pall.) O.Fedtsch. & B.Fedtsch.
- Oxytropis splendens Douglas
- Oxytropis squamulosa DC. (Syn.: Oxytropis leucopodia Ledeb.): Sie ist in Russland, in der Mongolei, in der Inneren Mongolei und in den chinesischen Provinzen Gansu, Ningxia, Qinghai, Shaanxi sowie Xinjiang verbreitet.[1]
- Oxytropis staintoniana Ali: Sie kommt nur in der pakistanischen Provinz Khyber Pakhtunkhwa vor.[2]
- Oxytropis stenofoliola Polozhij
- Oxytropis stenophylla Bunge
- Oxytropis stracheana Baker
- Oxytropis stracheyana Bunge: Sie ist Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan, Usbekistan, in der indischen Division Kumaon im Bundesstaat Uttarakhand, in der pakistanischen Provinz Khyber Pakhtunkhwa, in Tibet und im russischen Tian Shan sowie im Alaigebirge, in Tibet und in den chinesischen Provinzen Gansu, Qinghai sowie Xinjiang vor.[1][2]
- Oxytropis strobilacea Bunge
- Oxytropis stukovii Palib.
- Oxytropis suavis Boriss.
- Oxytropis subcapitata Gontsch.
- Oxytropis subfalcata Hance
- Oxytropis submutica Bunge
- Oxytropis subnutans (Jurtzev) Jurtzev
- Oxytropis subpodoloba P.C.Li: Sie gedeiht in kiesiger Ufervegetation und an Südhängen in Höhenlagen von 3500 bis 4200 Metern in Tibet.[1]
- Oxytropis subverticillaris Ledeb.
- Oxytropis sulphurea (DC.) Ledeb.
- Oxytropis sumneviczii Krylov
- Oxytropis suprajenissejensis Kuvaev & Sonnikova
- Oxytropis surculosa Rech. f.
- Oxytropis surmandehi Vassilcz.
- Oxytropis susamyrensis B.Fedtsch.
- Oxytropis susumanica Jurtzev
- Oxytropis sutaica N.Ulziykh.
- Oxytropis sverdrupii Lynge
- Oxytropis sylvatica (Pall.) DC.
- Oxytropis szovitsii Boiss. & Buhse
- Oxytropis tachtensis Franch.
- Oxytropis takhti-soleimanii Vassilcz.
- Oxytropis talassica Gontsch.
- Oxytropis talgarica Popov
- Oxytropis taochensis Kom.: Sie gedeiht auf Wiesen, an Straßenrändern, auf oberen Bereichen von Hügeln und an sandigen Standorten in Tälern in Höhenlagen von 2000 bis 3400 Metern in den chinesischen Provinzen Gansu, Ningxia, Qinghai, Shaanxi sowie Sichuan.[1]
- Oxytropis tashkurensis S.H.Cheng ex X.Y.Zhu, Y.F.Du & H.Ohashi: Die Erstbeschreibung erfolgte 2000. Sie gedeiht in Grasländern, an Südhängen und auf alpinen Matten in Höhenlagen von 1800 bis 3600 Metern in Xinjiang.[1]
- Oxytropis tatarica Baker: Sie kommt im nördlichen Teil des indischen Bundesstaates Punjab, in Kaschmir, Tibet, Nepal und vielleicht im westlichen Turkistans vor.[2]
- Oxytropis tenuirostris Boriss.
- Oxytropis tenuissima Vassilcz.
- Oxytropis terekensis B.Fedtsch.
- Oxytropis teres (Lam.) DC.
- Oxytropis tianschanica Bunge (Syn.: Oxytropis brachycarpa Vassilcz., Oxytropis pulvinata Saposhn.): Sie kommt in Kirgisistan, Tadschikistan, Tibet und Xinjiang vor.[1]
- Oxytropis tichomirovii Jurtzev
- Oxytropis tilingii Bunge
- Oxytropis todomoshiriensis Miyabe & Miyake
- Oxytropis tomentosa Gontsch.
- Oxytropis tomoriensis Kit Tan, Shuka & G. Vold[48]: Sie kommt nur auf der Südspitze des Tomorrs in S-Albanien vor.
- Oxytropis tompudae Popov
- Oxytropis torrentium Vassilcz.
- Oxytropis tragacanthoides DC. (Syn.: Oxytropis paratragacanthoides Vassilcz.): Sie kommt in Kasachstan, in der Mongolei, in der Inneren Mongolei und in den chinesischen Provinzen Gansu, Ningxia, Qinghai sowie Xinjiang vor.[1]
- Oxytropis trajectorum B.Fedtsch.
- Oxytropis transalaica Vassilcz.
- Oxytropis trichocalycina Bunge: Sie kommt in Kasachstan, Kirgisistan, Tadschikistan, Turkmenistan, Usbekistan und Xinjiang vor.[1]
- Oxytropis trichophora Franch.: Sie gedeiht auf Hügeln, an Straßenrändern und in Grasländern in Höhenlagen von 800 bis 2000 Metern in den chinesischen Provinzen Gansu, Hebei, Henan, Shaanxi sowie Shanxi.[1]
- Oxytropis trichophysa Bunge: Sie kommt im westlichen Sibirien, in der Mongolei und in den chinesischen Provinzen Gansu sowie Xinjiang vor.[1]
- Dreiblüten-Spitzkiel (Oxytropis triflora Hoppe)
- Oxytropis triphylla (Pall.) Pers.
- Oxytropis tschatkalensis L.I.Vassiljeva
- Oxytropis tschimganica Gontsch.
- Oxytropis tudanensis X.Y.Zhu, H.Ohashi & Si Feng Li: Sie gedeiht auf Hügeln in Höhenlagen von 2800 bis 4900 Metern in Tibet sowie Gansu.[1]
- Oxytropis tukemansuensis X.Y.Zhu, H.Ohashi & Y.B.Deng: Dieser Endemit kommt nur in Höhenlagen von etwa 4200 Metern in Taschkorgan in Xinjiang vor.[1]
- Oxytropis tunnellii Vassilcz.
- Oxytropis tyttantha Gontsch.
- Oxytropis ugamensis Vassilcz.
- Oxytropis ugamica Gontsch.
- Oxytropis ulzijchutagii Sanchir
- Oxytropis uniflora Jurtzev
- Oxytropis uralensis (L.) DC.
- Oxytropis urumovii Jáv.
- Oxytropis uschakovii Jurtzev
- Oxytropis vadimii Vassilcz.
- Oxytropis vakhdzhiri Vassilcz.
- Oxytropis valerii Vassilcz.: Sie gedeiht auf den oberen Bereich von Hügeln und kiesigen Standorten in Höhenlagen von 3500 bis 4000 Metern in Xinjiang.[1]
- Oxytropis varlakovii Serg.
- Oxytropis vassilczenkoi Jurtzev
- Oxytropis vassilievii Jurtzev
- Oxytropis vasskovskyi Jurtzev
- Oxytropis vavilovii Vassilcz.
- Oxytropis vermicularis Freyn: Sie gedeiht auf den oberen Bereich von Hügeln und kiesigen Standorten in Höhenlagen von 3500 bis 4000 Metern nur im südwestlichen Xinjiang.[1]
- Oxytropis viae-amicitiae Vassilcz.
- Oxytropis viridiflava Kom.
- Oxytropis viscida Torr. & A.Gray
- Oxytropis volkii Rech. f.
- Oxytropis vositensis Vassilcz.
- Oxytropis vvedenskyi Filim.
- Oxytropis williamsii Vassilcz.
- Oxytropis wrangelii Jurtzev
- Oxytropis wutaiensis Tatew. & Hurus.: Dieser Endemit gedeiht in Grasländern nur im Wutai Shan in Shanxi.[1]
- Oxytropis xinglongshanica C.W.Chang: Die Erstbeschreibung erfolgte 2004 und seit 2007 gibt es eine zweite Varietät. Sie gedeihen je nach Varietät auf Hügeln oder in Tälern in Höhenlagen von 1800 bis 2600 Metern in Gansu.[1]
- Oxytropis yanchiensis X.Y.Zhu, H.Ohashi & L.R.Xu: Dieser Endemit kommt auf Hügeln nur in Höhenlagen von etwa 2200 Metern in Yiwu in Xinjiang vor.[1]
- Oxytropis yekenensis X.Y.Zhu, H.Ohashi & Y.B.Deng: Sie gedeiht auf den oberen Bereich von Hügeln in Höhenlagen von etwa 1400 Metern in Xinjiang. vor.[1]
- Oxytropis yunnanensis Franch. (Syn.: Oxytropis konlonica H.Ohba): Sie gedeiht in Höhenlagen von 1800 bis 4900 Metern in Tibet und in den chinesischen Provinzen Gansu, Qinghai, Sichuan sowie Yunnan.[1]
- Oxytropis zangolehensis Vassilcz.: Sie kommt im Iran vor.[43]
- Oxytropis zaprjagaevae Abdusal.: Sie kommt in Tadschikistan nur in Gorno-Badakshan vor.[43]
- Oxytropis zekogensis Y.H.Wu: Sie gedeiht an Ufern von Fließgewässern und auf alpinen Matten in Höhenlagen von 2700 bis 3400 Metern in der chinesischen Provinz Qinghai.
Weidepflanzen und Weideunkräuter
Weidepflanzen
Spitzkiel-Arten sind wie viele weitere Hülsenfrüchtler relativ proteinreiche Pflanzen, die zudem atmosphärischen Stickstoff im Boden speichern können.[49] Während die europäischen Arten in der Weidewirtschaft unproblematisch sind, der Alpen-Spitzkiel als gute Futterpflanze gilt, der zum Teil als Wintersteher dem Vieh zur Verfügung steht,[50] so ist dessen Futterbedeutung da zumeist auf armen Magerrasen in populationsarmen Beständen nur sporadisch auftretend und unter kontinuierlicher Beweidung zumeist durch Ausfallen gekennzeichnet, dennoch ohne quantitativ messbaren Nutzen. Da alle Mitteleuropäischen Arten im Bestand entweder gefährdet oder zerstreut bis selten vorkommend sind, haben sie auch aus bestanderhaltenden Maßnahmen keine Bedeutung für die Beweidung.
Nach älteren Angaben aus Gustav Hegis Illustrierter Flora von Mitteleuropa enthält der Alpen-Spitzkiel 80,3 % organische Substanz (19,75 % Roheiweiss, 3,7 % Rohfett, 16,97 % Rohfaser und 39,61 % stickstofffreie Extraktstoffe u. a.)[51]
Weitere Inhaltsstoffe und Wirkung der Substanzen bei mitteleuropäischen Arten sind zum Teil bekannt. Die Samen Der Spitzkiele enthalten Blausäureglykoside und circa 4 % fettes Öl. Außerdem wurden Chinolizidinalkaloide nachgewiesen.[52]
„Weideunkräuter“
Einige insbesondere weitverbreitete nordamerikanische Arten sind durch ein toxisches Alkaloid als Giftpflanzen eine für Schafe, Ziegen, Rinder, Pferde wie für große Wildarten unerwünschte, da pathologische Nahrungsquelle. Durch die weite Verbreitung in den Weidegebieten der Plains ist ihnen ein bedeutendes Schädigungspotential zu eigen, für die eine Warnung an Weideviehzüchter durch die Agrarministerien besteht.[53] Der im angelsächsischen Raum gebräuchliche Trivialname der Oxytropis-Arten – Locoweed –, welches sich aus dem spanischen loco für verrückt und weed („Unkraut“) zurückführen lässt, weist auf diese neuropathologische Wirkung hin. Es ist der Beschreibung von Symptomen von vergifteten Tieren geschuldet, die nach längerer Aufnahme von Spitzkiel-Arten als orientierungslos, geschwächt, nervös und sich von anderen Tieren als absondernd beobachtet werden. Über das im Spitzkiel auftretende Inolizin-Alkaloid Swainsonin treten Symptome der Locoismus genannten Krankheit zwei bis drei Wochen nach Aufnahme auf was durch Abmagerung, Lethargie und depressives Verhalten wie trüben Augen und spröden Fell schnell deutlich sichtbar wird. Insbesondere ist die Prognose für Pferde ungünstig, da ihre weitere Nutzung als Trag- oder Zugtier hinfällig wird.[53] Pferde mit Locoismus sind in ihrem Beurteilung, Sehvermögen und Nerven nachhaltig irritiert: befallene Pferde springen so über kleine Spalten, treten dagegen aber in einen tiefen Graben.
Insgesamt steigt die Mortalität des Weideviehs nach Nahrungsaufnahme von Spitzkiel-Arten stark an. Nach einer Initialvergiftung reagieren Tiere zudem verstärkt auf nachfolgende Vergiftungen. Weidevieh nimmt Spitzkiel-Arten insbesondere im Frühjahr und Herbst gerne an, jedoch können die trockenen Blütenstiele auch im Winter eine Nahrungsquelle stellen. Insbesondere stellen Spitzkiel-Arten. die in den klassischen Weidegebieten des nordamerikanischen Westens in den Great Plains und Rocky Mountains vorkommen im Frühjahr, bevor sich Gras wächst eine wichtige Nahrungsquelle, das das Vieh gerne aufnimmt. Nach regelmäßiger Futteraufnahme von Oxytropis können sich bei den Weidetieren Abhängigkeiten ausbilden die danach selektiv nach den Pflanzen suchen.[49][54] Dabei sind Spitzkiel-Arten in allen ihren Teilen und Stadien giftig. Der Tod von Weidetieren tritt nach ein bis drei Monaten von großen Mengen von Oxytropis sericea auf.
Die vier prinzipiellen Effekte einer Swainsonin-Vergiftung bei Weidevieh sind: neurologische Schäden, Abmagerung, Reproduktionsstörungen und Fehlgeburten, kongestives Herzversagen der rechten Herzkammer bei Hochgebirgsweidetieren.
Bedeutendstes Weideunkraut unter den nordamerikanischen Spitzkiel-Arten ist Oxytropis sericea (white point loco), zudem sind Oxytropis lambertii (Lambert locoweed) und auch in Kanada vorkommende Oxytropis campestris agg. (auch in den Alpen als Alpen-Spitzkiel verbreitet) Arten, die in Gebieten mit Weideviehhaltung vorkommen und die toxischen Substanzen auch nach mehrjähriger Lagerung kaum verlieren.[55] Oxytropis sericea wird durch starke Beweidung gefördert, eine Ausrottung über große Weideflächen ist selten möglich.
Als Giftpflanzen sind zurzeit folgende nordamerikanischen, zentral- und ostasiatischen Arten eingestuft: Oxytropis campestris var. spicta, Oxytropis lambertii, Oxytropis ochrocephala, Oxytropis puberula und Oxytropis sericea.[56]
Nutzung
Spitzkiel-Arten werden als Heil- sowie Zierpflanzen verwendet und sind Vitaminreich.[3]
Traditionelle Medizin
So weit bekannt wurden nur einige Spitzkiel-Arten von Nordamerikanischen Indianern in der Volksmedizin genutzt. Sie wurden als rituelle oder medizinische Waschungen bei Schwitzhüttenzeremonien verwendet. In Mexiko heißt die Art Oxytropis lamberti „hierba loca“, „verrücktmachendes Kraut“.[57]
Onkologie
Die zytotoxischen, d. h. zelltötenden Eigenschaften einiger Arten könnten in der Krebstherapie genutzt werden, sind jedoch zurzeit nicht genügend erforscht.[58]
Renaturierung und Erosionsschutz
In Alaska, Britisch Kolumbia und China werden Spitzkiele für Renaturierung und Erosionsschutz empfohlen. Durch ihre Pfahlwurzel stabilisieren die Arten erosionsgefährdete Hänge langfristig. Über ihre Stickstoffautarkie können sie als Pionierpflanzen auch unwirtliche Standorte besiedeln.[59][60][61]
Zierpflanze
Spitzkiel-Arten werden in Gärten als Zierpflanzen verwendet, da sowohl die Blattrosetten mit den feinen spitzen Blättern, die oft wollig oder seidig behaart sind, als auch auf den langen Blütenstandsschäften die dichten Blütenstände und Blüten dekorativ sind.
Überwiegend werden sie im Steingarten, Hochbeet oder Alpinum angepflanzt. Einige Arten sind schwer kultivierbar, was wahrscheinlich auf das Fehlen der Knöllchenbakterien der Gattung Rhizobium zurückgeht. Alle Spitzkiel-Arten in Kultur brauchen gut drainierte, tiefe, kiesige Böden und stehen gern in voller Sonne.
Als Zierpflanzen verwendet werden: Alpen-Spitzkiel (Oxytropis campestris), Oxytropis foetida, Oxytropis halleri, Oxytropis jacquinii, Oxytropis lambertii, Oxytropis lapponica, Oxytropis lazica, Oxytropis megalantha, Oxytropis pilosa, Oxytropis pyrenaica, Oxytropis sericea, Oxytropis shokanbetsuensis, Oxytropis splendens, Oxytropis uralensis.
Belege
Literatur
- Langran Xu, Xiangyun Zhu, Bojian Bao, Mingli Zhang, Hang Sun, Dietrich Podlech, Stanley L. Welsh, Hiroyoshi Ohashi, Kai Larsen, Anthony R. Brach: Galegeae., S. 322: Xiangyun Zhu, Stanley L. Welsh, Hiroyoshi Ohashi: Oxytropis, S. 453 – textgleich online wie gedrucktes Werk, In: Wu Zheng-yi, Peter H. Raven, Deyuan Hong (Hrsg.): Flora of China, Volume 10 – Fabaceae, Science Press und Missouri Botanical Garden Press, Beijing und St. Louis, 2010. ISBN 978-1-930723-91-7. (Abschnitte Systematik und Verbreitung)
- B. A. Yurtsev: Survey of Arctic legumes with emphasis on the species concept in Oxytropis. In: Norske Vidensk. Akad. I. Mat. Naturvitensk. Kl., Skr. Ny ser. 38, 1999, S. 295–318 lokale Kopie
- T. G. Tutin, V. H. Heywood, N. A. Burges, D. M. Moore, D. H. Valentine, S. M. Walters, D. A. Webb (Hrsg.): Flora Europaea. Volume 2: Rosaceae to Umbelliferae. Cambridge University Press, Cambridge, 1968, ISBN 0-521-06662-X, S. 124–128 (Oxytropis auf S. 124–128 in der Google-Buchsuche).
- Walter Gutermann, Hermann Merxmüller: Die Europäischen Sippen von Oxytropis Sectio Oxytropis. In: Mitteilungen der Botanischen Staatssammlung München. Band 4, 1961, S. 199–276. Hier S. 231–233.
Einzelnachweise
- Xiangyun Zhu, Stanley L. Welsh, Hiroyoshi Ohashi: Oxytropis, S. 453 – textgleich online wie gedrucktes Werk, In: Wu Zheng-yi, Peter H. Raven, Deyuan Hong (Hrsg.): Flora of China, Volume 10 – Fabaceae, Science Press und Missouri Botanical Garden Press, Beijing und St. Louis, 2010, ISBN 978-1-930723-91-7.
- Syed Irtifaq Ali: Oxytropis bei Tropicos.org. In: Flora of Pakistan. Missouri Botanical Garden, St. Louis.
- Vladimir Leontyevich Komarov, B. K. Shishkin, E. G. Bobrov (Hrsg.): Flora of the U.S.S.R. Volume XIII. Leguminosae: Oxytropis, Hedysarum. Israel Program for Scientific Translations/Smithsonian Institution and the National Science Foundation, Jerusalem/Washington, D.C. 1972, ISBN 0-7065-1241-3 (engl., übersetzt von R. Lavoott; russisches Original: Botanicheskii institut, Izdatel'stvo Akademii Nauk SSSR, Moskau/Leningrad 1948) Digitalisat. S. 2.
- Norman Andrew Willey: Propagation of three native alpine Legumes Proceedings of the 7th Annual British Columbia Mine Reclamation Symposium in Victoria, BC, 1983. The Technical and Research Committee on Reclamation. doi:10.14288/1.0042077 PDF.
- Smith & Yan Xie 2009:Koslov’s Pika. In: Andrew T. Smith, Yan Xie: A Guide to the Mammals of China. Princeton University Press, 2008; S. 282–283. ISBN 978-0-691-09984-2.
- Ekaterina K. Kozuharova 2000: Reproductive Biology of Oxytropis urumovii Jav. and Oxytropis campestris (L.) DC. (FABACEAE). Annual fo Sofia University „St. Kliment Ohridski“, Faculty of Biology, Book 2, Vol. 91: 49–61
- Walter Larcher 2001: Ökophysiologie der Pflanzen: Leben, Leistung und Stre0bewältigung der Pflanzen in ihrer Umwelt. 6. neubearbeitete Auflage, Ulmer, Stuttgart. ISBN 3-8252-8074-8
- Soudzilovskaia, Nadejda, Aksenova, A. A., Makarov, Mikhail, Onipchenko, Vladimir, Logvinenko, O. A., ter Braak, Cajo and Cornelissen, Johannes 2012: Legumes affect alpine tundra community composition via multiple biotic interactions. Ecosphere 3/4, April 2012, Article 33.
- Alpenschwemmlinge am Tiroler Lech
- Alpine Flüsse mit Ufergehölzen von Myricaria germanica in den Ostalpen
- P. M. Schneider, 1964: Verbreitungsbiologie und Pflanzengesellschaften. In: Acta Botanica Croatica, Volume extraord. 1964, S. 79–87.
- Alla b. Kholina, Nina M. Voronkova 2012: Seed Cryopreservation of Some Medicinal Legumes. Hindawi Publ. Corp. Journal ov Botany. Volume 2012. Article ID 186891, doi:10.1155/2012/186891
- I. Matijevic, D. Babic, B. Radak, B. Bokic, G. Anackov 2018: Testing the Seed Germination of Oxytropis pilosa (L.) DC. 1802 (Fabaceae). Botanica Serbica, vol. 42/1: 117. (Botanica Serbica:PDF)
- Seed anatomy in Alaskan Oxytropis
- The Circumpolar Arctic Vegetation Map: AVHRR-derived base maps, environmental controls, and integrated mapping procedures D. A. WALKER, W. A. GOULD, H. A. MAIER and M. K. RAYNOLDS (PDF)
- Boris A. Yurtsev: Survey of Arctic legumes with emphasis on the species concept in Oxytropis. In: Norske Vidensk. Akad. I. Mat. Naturvitensk. Kl., Skr. Ny ser. 38, 1999, S. 295–318.
- Anthony Huxley (Hrsg.): The New Royal Horticultural Society Dictionary of Gardening, 3 L to Q. McMillan Press, London 1992, ISBN 1-56159-001-0, S. 425.
- Flora of the Canadian Arctic
- (PDF)
- An expedition to East Ross to rescue the rare Oxytropis halleri
- Hermann Meusel, Eckehart Jäger, Erich Weinert: Vergleichende Chorologie der zentraleuropäischen Flora. Bd. 1. – Gustav Fischer Verlag Stuttgart New York 1965. Hier S. 193
- Christoph Leuschner and Heinz Ellenberg 2017: Vegetation Ecology of Central Europe. Springer, Berlin. ISBN 978-3-319-50710-1 Hier S. 414–415
- Hermann Meusel, Eckehart Jäger, Erich Weinert: Vergleichende Chorologie der zentraleuropäischen Flora. S. 194
- Dickoré, W. B. & Kriechbaum, M.: Oxytropis iridum (Leguminosae), a new species from SE Tibet (Xizang, China), including phytogeographical notes. – Willdenowia 36: 857–865. – ISSN 0511-9618; © 2006 BGBM Berlin-Dahlem. (Willdenowia:PDF)
- Jörg S. Pfadenhauer; Frank A. Klötzli 2014: Vegetation der Erde – Grundlagen, Ökologie, Verbreitung. Springer Spektrum 2014, ISBN 978-3-642-41950-8. Hier S. 521
- Vladimir Leontyevich Komarov, B. K. Shishkin, E. G. Bobrov (Hrsg.): Flora of the U.S.S.R. Volume XIII. Leguminosae: Oxytropis, Hedysarum. Israel Program for Scientific Translations/Smithsonian Institution and the National Science Foundation, Jerusalem/Washington, D.C. 1972, ISBN 0-7065-1241-3, (engl., übersetzt von R. Lavoott; russisches Original: Botanicheskii institut, Izdatel'stvo Akademii Nauk SSSR, Moskau/Leningrad 1948) Digitalisat, S. 1 (Vollständiger Scan in der Biodiversity Heritage Library)
- Manfred A. Fischer, Karl Oswald, Wolfgang Adler: Exkursionsflora für Österreich, Liechtenstein und Südtirol. 3., verbesserte Auflage. Land Oberösterreich, Biologiezentrum der Oberösterreichischen Landesmuseen, Linz 2008, ISBN 978-3-85474-187-9.
- Siegmund Seybold (Hrsg.): Schmeil-Fitschen interaktiv. CD-ROM, Version 1.1, Quelle & Meyer, Wiebelsheim 2002, ISBN 3-494-01327-6.
- Jindřich Chrtek, Anna Chrtková: Bemerkungen zu einigen balkanischen Oxytropis-Arten. In: Folia Geobotanica & Phytotaxonomica. Band 18, Nr. 3, 1983, S. 309–320. JSTOR 4180441
- P. Leins, Hermann Merxmüller: Zur Gliederung der Oxytropis campestris-Gruppe. In: Mitt. d. Bot. Ges. München. Band VI, 1966, S. 19–31 (PDF)
- Čedomil Šilić: Emdemične biljke. In: Priroda Jugoslavije. 4, 1990, S. 61–62, Svjetlost Sarajevo, 3. Ausgabe, ISBN 86-01-02557-9.
- Flora of the Canadian Arctic Archipelago – S.G. Aiken, M.J. Dallwitz, L.L. Consaul, C.L. McJannet, R.L. Boles, G.W. Argus, J.M. Gillett, P.J. Scott, R. Elven, M.C. LeBlanc, L.J. Gillespie, A.K. Brysting, H. Solstad, and J.G. Harris – Oxytropis arctica R. Br.
- A. B. Kholina, O. V. Nakonechnaia, V. V. Iakubov, O. G. Koren: Genetic variation in six species of the genus Oxytropis DC. (Fabaceae) from Kamchatka Peninsula. In: Genetika. Band 49, 10, Oktober 2013, S. 1174–1182. doi:10.1134/S1022795413100049
- A. B. Kholina, O. V. Nakonechnaia, V. V. Iakubov, O. G. Koren 2013: Genetic variation in six species of the genus Oxytropis DC. (Fabaceae) from Kamchatka Peninsula., S. 1174.
- P. Schönswetter, A. Tribsch, H. Niklfeld: Amplified Fragment Length Polymorphism (AFLP) reveals no genetic divergence of the Eastern Alpine endemic Oxytropis campestris subsp. tiroliensis (Fabaceae) from widespread subsp. campestris. In: Plant Syst. Evol. Band 244, 2013, S. 245–255. Hier S. 253 (PDF)
- Ana Petrova (Hrsg.): Atlas of Bulgarian Endemic Plants. Gea-Libris, Sofia 2006, S. 146.
- P. Schönswetter, A. Tribsch, H. Niklfeld: Amplified Fragment Length Polymorphism (AFLP) reveals no genetic divergence of the Eastern Alpine endemic Oxytropis campestris subsp. tiroliensis (Fabaceae) from widespread subsp. campestris. 2004, S. 253.
- Thorsten Englisch: Multivariate Analysen zur Synsystematik und Standortsökologie der Schneebodenvegetation (Arabidetalia caereula) der Nördlichen Kalkalpen. In: Stapfia. Band 59, Linz 1999, zobodat.at [PDF]
- FloraWeb: Pflanzengesellschaften: Hochgebirgsrasen
- Oxytropis bei Tropicos.org. Missouri Botanical Garden, St. Louis Abgerufen am 1. September 2015.
- Walter Gutermann, Hermann Merxmüller: Die Europäischen Sippen von Oxytropis Sectio Oxytropis. In: Mitteilungen der Botanischen Staatssammlung München. Band 4, 1961, S. 199–276. Hier S. 231–233.
- Alexander Andrejewitsch von Bunge: Species generis Oxytropis DC. In: Mémoires de l'Academie Imperiale des Sciences de Saint Petersbourg, Ser. 7, Band 22, 1874, S. 1–166.
- Taxon in Suchmaske eingeben bei The LegumeWeb – International Legume Database and Information Service = ILDIS, Version 10 vom November 2005.
- E. V. Artyukova, M. M. Kozyrenko: Phylogenetic Relationships of Oxytropis chankaensis Jurtz. and Oxytropis oxyphylla (Pall.) DC. (Fabaceae) Inferred from the Data of Sequencing of the ITS Region of the Nuclear Ribosomal DNA Operon and Intergenic Spacers of the Chloroplast Genome. In: Russian Journal of Genetics. Band 48, Nr. 2, 2012, S. 163–169.
- Aiuob Moradi, A. A. Maassoumi, Saeid Afsharzade, B. Hamzeh׳ee, V. Mozaffarian: IRAN. J. BOT. Volume 21, 1. online.
- A new species of Oxytropis Fabaceae from Central Apennines Italy
- https://www.researchgate.net/publication/258118728_A_contribution_to_the_genus_Oxytropis_in_Iran [A contribution to the genus Oxytropis in Iran]
- (PDF)
- Lora L. Esser: Oxytropis sericea online. In: Fire Effects Information System, U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, 1993.
- Erich Oberdorfer: Pflanzensoziologische Exkursionsflora für Deutschland und angrenzende Gebiete. 7. Auflage. Verlag Eugen Ulmer, Stuttgart 2001, ISBN 3-8001-3131-5, S. 603.
- Gustav Hegi, H. Gams, H. Marzell: Illustrierte Flora von Mitteleuropa. Pteridophyta, Spermatophyta. 2. Auflage. Band IV. Teil 3: Angiospermae: Dicotyledones 2 (5) (Leguminosae – Tropaeolaceae). J.F. Lehmans, München 1924, S. 1449.
- S. G. Fleischhauer, R. Spiegelberger, J. Guthmann: Enzyklopädie Essbare Wildpflanzen. At Verlag, München 2014, ISBN 978-3-03800-752-4, S. 560.
- USDA – United States Department of Agriculture: Poisonous plant research – Locoweed (Astragalus and Oxytropis)
- Chris Allison: Livestock-Poisoning Plants of New Mexico Rangelands. In: Circular 531. College of Agricultural, Consumer and Environmental Sciences New Mexico State University (PDF)
- T. C. Jones, R. D. Hunt, N. W. King: Veterinary pathology. 6. Ausgabe. Wiley-Blackwell, 1997, ISBN 0-683-04481-8, S. 752.
- J. Wagstaff: International Poisonous Plants Checklist – an Evidence-Based Reference. CRC Press, Boca Raton 2008, ISBN 978-1-4200-6252-6, S. 284–285.
- Christian Rätsch: Enzyklopädie der psychoaktiven Pflanzen: Botanik, Ethnopharmakologie und Anwendung. At Verlag, Aarau 2007, ISBN 978-3-03800-352-6, S. 573.
- Christian Rätsch: Enzyklopädie der psychoaktiven Pflanzen: Botanik, Ethnopharmakologie und Anwendung. At Verlag, Aarau 2007, ISBN 978-3-03800-352-6, S. 548.
- Oxytropis deflexa – revegetation in Alaska (PDF)
- Propagation of three native Legumes (PDF)
- https://www.scitechnol.com/peer-review/locoweed-endophytes-a-review-XwQH.php?article_id=8302 Locoweed Endophytes: A Review
Weblinks
- Herbarbelege zu zahlreichen Oxytropis-Arten mit über 3000 Belegen im Herbarium des Museum Nationale d'Histoire Naturelle in Paris
- Oxytropis bei Tropicos.org. In: IPCN Chromosome Reports. Missouri Botanical Garden, St. Louis.
- Oxytropis im Germplasm Resources Information Network (GRIN), USDA, ARS, National Genetic Resources Program. National Germplasm Resources Laboratory, Beltsville, Maryland. Abgerufen am 4. Oktober 2017.
- Suche nach „Oxytropis“ in der Roten Liste gefährdeter Arten der IUCN.
- Arten bei FloraWeb.
- Arten bei InfoFlora dem nationalen Daten- und Informationszentrum der Schweizer Flora..
- Datenblatt bei Den virtuella floran mit Links zu Arten und dort zu den Verbreitungskarten.
- Abbildung der kräftigen Pfahlwurzel sowie von Wurzelknöllchen eines alpinen Spitzkiels aus 3.300 m Höhe
Ergänzende Literatur
- Ivan Tikhonovich Vassilczenko: Oxytropis. In: K. H. Rechinger (Hrsg.): Flora Iranica. Band 157, Akademische Druck- und Verlagsanstalt, Graz/Wien 1984, S. 101–164.
- Massoud Ranjbar, Roya Karamian, Soheila Bayat: Notes on Oxytropis sect. Mesogaea (Fabaceae) in Iran, with the description of a new species. In: Annales Botanici Fennici. Band 46, 2009, S. 235–238 (PDF-Datei).
- Seher Karaman Erkul, Zeki Aytaç: The revision of the genus Oxytropis (Leguminosae) in Turkey. In: Turkish Journal of Botany. Band 37, Nr. 1, 2013, S. 24–38 (PDF-Datei).