RNA-Welt-Hypothese
Die RNA-Welt-Hypothese besagt, dass den heutigen Lebensformen eine Welt vorausging, deren Leben auf Ribonukleinsäuren (RNA) als universellen Bausteinen zur Informationsspeicherung und zur Katalyse chemischer Reaktionen basierte. Im Rahmen dieser Hypothese wird angenommen, dass freie oder zellgebundene RNA im Rahmen der Evolution vom chemisch stabileren Informationsspeichermedium Desoxyribonukleinsäure (DNA) und von den funktionell flexibleren Proteinen abgelöst beziehungsweise um diese ergänzt wurde.
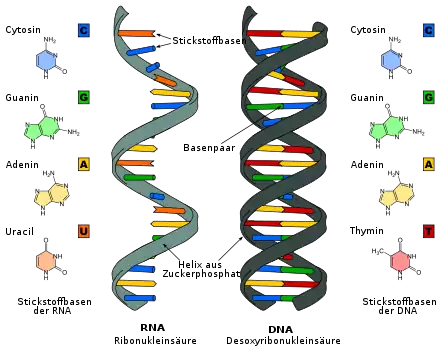
Als ein Hinweis auf die Existenz der RNA-Welt werden Ribozyme (katalytisch aktive RNA) angesehen, die als chemische (auch: molekulare) Fossilien evolutionäre Überbleibsel aus dieser Anfangszeit darstellen könnten (ähnlich Lebenden Fossilien); ein Beispiel dafür ist die ribosomale RNA (rRNA). Einen weiteren Hinweis gibt die Reihenfolge der Biosynthese-Pfade. Die Desoxyribonukleotide, aus denen die DNA zusammengebaut wird, werden in der Zelle aus Ribonukleotiden, den Bausteinen der RNA, erzeugt, indem die 2'-Hydroxygruppe entfernt wird. Die Zelle muss also erst in der Lage sein, RNA aufzubauen, bevor sie DNA herstellen kann.
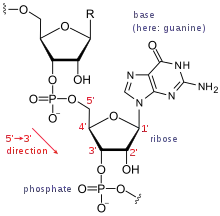
Die vier Ribonukleotide können unter bestimmten Umweltbedingungen spontan entstehen.[1] Die RNA-Welt-Hypothese ist somit ein Bindeglied zwischen den Hypothesen der chemischen Evolution, welche die Entstehung organischer Moleküle aus anorganischen Verbindungen erklären, und der Kooperation mehrerer RNAs, die eine biologische Selektion in Gang bringen,[2] sowie der heutigen biologischen Evolution auf Basis eines DNA-Genoms. Das Aufkommen zellulärer Lebensformen steht möglicherweise nicht am Ende dieses Prozesses, denn zelluläre Organismen auf RNA-Basis (Ribozyten) könnten bereits in der RNA-Welt entstanden sein.[3]
Konzept
Das Konzept einer postulierten RNA-Welt basiert auf zwei grundlegenden Eigenschaften der RNA. Zum einen dient sie wie die DNA als Medium zur Speicherung von Erbinformationen. Andererseits sind RNA wie auch Proteine in der Lage, chemische Reaktionen zu katalysieren.
Entstehung
Wenngleich RNA ein sehr komplexes Molekül ist, welches aus Ribonukleotid-Einheiten besteht, die wiederum Kondensationsprodukte der Ribose, einer Nukleinbase und Phosphat sind, gilt ihre abiotische Entstehung in der Ursuppe als möglich. Zwar ist eine direkte Kondensation katalytisch sehr aufwändig und daher unwahrscheinlich für eine abiotische Entstehung, jedoch konnte 2009 experimentell gezeigt werden, dass aus den einfachen Molekülen Cyanamid, Cyanoacetylen, Glycolaldehyd, Glyceraldehyd und Phosphat, welche mögliche Bestandteile der Ursuppe waren, in wenigen Reaktionsschritten Pyrimidinribonukleotide gebildet werden können.[4] Interessanterweise ist Phosphat in dieser Reaktion nicht nur als Ausgangsstoff (Edukt) bei der Veresterung des Nukleosids zum Nukleotid von Bedeutung. Vielmehr scheint die Anwesenheit von Phosphat selektiv gleich mehrere Teilschritte der Reaktion zu steuern, indem es z. B. als pH-Puffer und Katalysator mit nukleophilen Eigenschaften fungiert. Dadurch wird die kombinatorische Vielfalt unterdrückt, so dass kaum unerwünschte Nebenprodukte auftreten.[4]
Die komplexeren Purinnukleoside konnten 2016 mit kleinen Molekülen, wie sie heute noch auf Kometen zu finden sind, über den Formamidopyrimidin(FaPy)-Reaktionspfad hergestellt werden.[5] Zuletzt wurde gezeigt, wie die Pyrimidine und Purine auch zusammen unter den gleichen, plausiblen Bedingungen entstehen können.[6][1]
Informationsspeicherung
Auf Grund ihrer chemischen und physikalischen Eigenschaften wird die RNA für das ältere Speichermedium für Erbinformationen gehalten als die DNA. So kann beispielsweise der RNA-Baustein Ribose im Gegensatz zur Desoxyribose der DNA leicht durch eine Aldolkondensation gebildet werden. Wie die DNA ist auch die RNA in der Lage, sich zu doppelsträngigen Makromolekülen zusammenzulagern. Für eine Langzeitspeicherung von Informationen jedoch ist die RNA der DNA unterlegen, da RNA fehleranfällig und somit energieintensiv ist.
Katalyse
Mit der Entdeckung, dass die ribosomale RNA des Wimpertierchens Tetrahymena sich selbst spleißen kann, wurde 1982 erstmals der Nachweis katalytischer Eigenschaften von Ribonukleinsäuren (Ribozyme) erbracht.[7] Im Prinzip kann fast jede Nukleinsäure unter geeigneten Bedingungen in eine katalytische Sequenz evolvieren. Insbesondere für die DNA ist dies unter Laborbedingungen gelungen (so genannte Desoxyribozyme, auch DNAzyme). In der Natur wurde dies aber noch nicht beobachtet.[8][9] Dagegen konnten (neben künstlich erzeugten Ribozymen) natürliche RNA-Moleküle gefunden werden, die wichtige Bestandteile des Zellstoffwechsels, einschließlich ihrer eigenen Bausteine, produzieren[10] und sogar solche, die zur identischen Vervielfältigung (Replikation)[11][12] befähigt sind.
Im Rahmen der RNA-Welt-Hypothese wird angenommen, dass Ribozyme, die zur Übersetzung ihrer Erbinformation durch Synthese von Proteinen (Translation) befähigt waren, den Siegeszug der Proteine als Funktionsträger einleiteten. Proteine sind der RNA bezüglich ihrer katalytischen Fähigkeiten und der Katalysegeschwindigkeit überlegen.[3] Diese Annahme wurde durch die Entdeckung gestützt, dass die wichtigen katalytischen Zentren der Ribosomen von RNA und nicht, wie vorher angenommen, von Proteinen gestellt werden.[13]
Zelluläre RNA
Für die Entwicklung der RNA-Welt unter den Bedingungen der Darwinschen Evolution wird eine Kompartimentierung als wichtige Voraussetzung angesehen.[14] Es wird angenommen, dass sich diese ersten von einer Zellmembran umgebenen RNA-basierten hypothetischen Lebensformen, die Ribozyten, rasch weiterentwickelten und elementare Zellfunktionen, wie den Stofftransport durch die Zellmembran und die Zellform steuern konnten.[3]
Zwei Zeitalter der RNA-Welt
Moderne Ribosomen (die Protein-Fabriken der Zelle) sind aus ribosomaler RNA (rRNA) und Proteinen (rProtein) aufgebaut. Die Proteine scheinen hierbei nur eine Hilfsfunktion zu haben, welche die Übersetzung der Messenger-RNA (mRNA) in ein Protein schneller und/oder sicherer machen, im Prinzip reicht die rRNA als Ribozym für diesen Vorgang aus. Die RNA-Welt kann danach in zwei Zeitalter unterteilt werden: Eine frühe Phase ohne Ribosomen (Primordiale RNA-Welt, mit kurzen Peptidketten abiotischen Ursprungs aus der Ursuppe), und eine spätere mit Protein-Biosynthese durch Ribosomen (RNP-Welt, Ribonukleoprotein). Dazwischen fällt die Entstehung und Evolution des genetischen Codes.[15] Als die Organismen auf die so erzeugten Proteine angewiesen waren, musste die Evolution des genetischen Codes (weitestgehend) abgeschlossen sein, da eine Änderung des Codes eine Änderung und mögliche Funktionsunfähigkeit fast aller erzeugten Proteine bedeutet. Heute werden daher auch nur noch sehr geringe natürliche Abweichungen des genetischen Codes beobachtet.[16][17][18]
Alanin-Welt-Hypothese
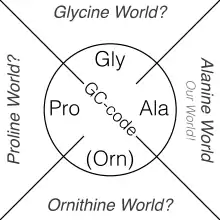
Es gibt mehrere Szenarien, wie die kanonischen Aminosäuren für die Proteinbiosynthese in der RNA-Welt gewählt wurden. Die „Alanin-Welt“-Hypothese[19] stellt die kanonische Aminosäure Alanin in den Mittelpunkt der sogenannten Proteinwelt. Als Sekundärstrukturen von Proteinen herrschen die α-Helix und das β-Faltblatt vor. Die am häufigsten als Monomere ausgewählten Aminosäuren für die ribosomale Synthese der Polypeptidketten können als chemische Derivate des Alanins aufgefasst werden. Sie eignen sich am besten für den Aufbau von α-Helices und β-Faltblätter in Proteinen.[20] Die Alanin-Welt-Hypothese wird praktisch bestätigt durch Alaninscanning, da bei dieser Methode Aminosäuren durch Punktmutationen zu Ala ausgetauscht werden, während die Sekundärstruktur noch intakt bleibt. Auch in der klassischen Röntgenkristallographie kommt dieses Prinzip zur Geltung, da häufig das Polyalanin-Grundgerüst-Modell verwendet wird um dreidimensionale Strukturen von Proteinen zu bestimmen.[21]
In Transmembranproteinen bestehen die membrandurchspannenden Anteile, die als Transmembrandomäne das Protein in der Lipid-Doppelschicht einer Biomembran verankern, soweit bekannt ganz überwiegend aus diesen Sekundärstrukturen.
Prä-RNA-Welt
Da die RNA ein bereits sehr komplexes Molekül ist, wurden sowohl alternative als auch ergänzende Hypothesen zur Entwicklung des Lebens auf der Erde entwickelt. Dazu gehört insbesondere eine mögliche Rolle von Proteinoiden, Peptid-Nukleinsäuren (PNA), Threose-Nukleinsäuren (TNA)[22] und Glycerol-Nukleinsäuren (GNA)[23]. Da sie chemisch einfacher gebaut sind, werden diese entweder als mögliche Vorgänger der RNA angesehen, oder es wird eine chemische Koevolution angenommen. Für Peptid-Nukleinsäuren konnte nachgewiesen werden, dass sie sich selbst replizieren und als Vorlage der RNA dienen können.[24] Eine Entstehung von Peptid-Nukleinsäuren in der Ursuppe gilt als möglich.[25] Auch die in der Ursuppe vermuteten polycyclischen aromatischen Kohlenwasserstoffe (PAHs) werden als mögliche Vorgänger der RNA angesehen.[26]
Folgende Hypothesen wurden diskutiert, die andere Nukeinsäure-Welten (Xeno-Nukleinsäuren, XNA) als Vorgänger einer RNA-Welt vorschlagen:
- XNA allgemein:
- Robert T. Gonzalez: XNA is synthetic DNA that's stronger than the real thing, in: io9, 19. April 2012
- M. Schmidt: Xenobiology: a new form of life as the ultimate biosafety tool. In: BioEssays : news and reviews in molecular, cellular and developmental biology. Band 32, Nummer 4, April 2010, S. 322–331, doi:10.1002/bies.200900147, PMID 20217844, PMC 2909387 (freier Volltext) (Review).
- M. P. Robertson, G. F. Joyce: The origins of the RNA world. In: Cold Spring Harbor perspectives in biology. Band 4, Nummer 5, Mai 2012, S. , doi:10.1101/cshperspect.a003608, PMID 20739415, PMC 3331698 (freier Volltext) (Review).
- TNA:
- David Bradley: The TNA world that came before the RNA one, in: Chemistry World, 8. Januar 2012
- Am Anfang war die TNA?, in: spektrum.de vom 20. November 2000
- PNA:
- Peter E. Nielsen: Ein neues Molekül des Lebens?, in: Spektrum der Wissenschaft, Oktober 2009, S. 42–49, Abschnitt Ursprung des Lebens
- GNA:
- Orgel L: Origin of life. A simpler nucleic acid. In: Science. 290, Nr. 5495, Nov 2000, S. 1306–7. doi:10.1126/science.290.5495.1306. PMID 11185405.
- Nelson KE, Levy M, Miller SL: Peptide nucleic acids rather than RNA may have been the first genetic molecule. In: Proceedings of the National Academy of Sciences. 97, Nr. 8, Apr 2000, S. 3868–71. bibcode:2000PNAS...97.3868N. doi:10.1073/pnas.97.8.3868. PMID 10760258. PMC 18108 (freier Volltext).
Alternativhypothesen
Nicht alle der vorgeschlagenen alternativen Hypothesen schließen sich gegenseitig und die RNA-Welt-Hypothese aus, vielmehr könnten diese Szenarien meist auch gemeinsam bei der Entstehung moderner Zellen beteiligt gewesen sein. Eine Auswahl:
Eisen-Schwefel-Welt
Die von Günter Wächtershäuser vorgeschlagene Eisen-Schwefel-Welt-Theorie nimmt an, dass sich einfache Stoffwechselprozesse bereits vor den Nukleinsäuren als genetischem Material entwickelten, und dass diese Energie erzeugenden Zyklen die Produktion der genetischen Information, also von Genen, katalysierten.
Proteinhypothese
Sidney W. Fox schlug vor, dass Proteinoide, durch Kondensationsreaktionen abiotisch entstandene, proteinähnliche Aminosäureketten, eine Vorstufe von Lebewesen im Sinne einer Abiogenese darstellen könnten, in dem sie ebenfalls Mikrosphären und später eine Speicherstruktur der genetischen Information erzeugten.
RNA-Peptid-Koevolution
Die RNA-Welt-Hypothese im strengen Sinn geht davon aus, dass zunächst nur Ribozyme katalytische Fähigkeiten entfalteten, und Peptide oder gar die längerkettigen Proteine keine oder nur eine marginale Rolle gespielt haben. Erst in dem Umfang, in dem die Biosynthese von Aminosäureketten (an den Ribosomen) aufkam, nahmen auch die so erzeugten Peptide ihre enzymatische Funktion auf und aus der RNA-Welt wurde eine RNP-Welt (Ribonukleoprotein-Welt).
Im Gegensatz dazu geht die Hypothese der RNA-Peptid-Koevolution[27] davon aus, dass es ausgehend von abiotisch entstandenen primordialen Oligopeptiden (kurze Aminosäureketten) zu einer chemischen Koevolution der Peptide und der RNA und ihrer katalytischen Fähigkeiten kam. Allerdings zeigen die verschiedenen Ursuppen-Experimente, dass kurze Peptidketten (Oligopeptide) bereits in der Anfangszeit der Chemischen Evolution entstehen konnten. Die Theorie setzt also die gleichzeitige Entstehung und Entwicklung zweier komplexer Moleküle (Peptid als Enzym und RNA als Träger der Erbinformation) voraus. Sie beschreibt somit eine 'von Anfang an' existierende RNP-Welt: Das heutige duale System, in dem Nukleinsäuren benötigt werden, um Proteine herzustellen, und Moleküle auf Proteinbasis umgekehrt die Nukleinsäuren verketten, hätte es also immer schon gegeben, lediglich eine Effizienzsteigerung und Zunahme der jeweiligen Kettenlängen wäre zu verzeichnen.[28][29] Die Annahme wird unter anderem dadurch unterstützt, dass es offenbar eine ganze Reihe evolutionär sehr alter Protein-RNA-Komplexe gibt, unter anderem die Ribosomen und die Telomerasen. Die Hypothese könnte die schnelle Evolution der Genauigkeit der RNA-Replikation erklären, da die Proteine wie in rezenten Ribosomen als Katalysatoren wirken.[30][31][32]
Lipid-Hypothese
Ein im März 2015 von der John-Sutherland-Gruppe abgeschlossenes Forschungsprojekt stellte fest, dass ein Netzwerk von Reaktionen beginnend mit Cyanwasserstoff und Schwefelwasserstoff in fließendem Wasser, das durch UV-Licht bestrahlt wird, die chemischen Komponenten von Proteinen und Lipiden zusammen mit RNA produzieren könnte.[33][34] In diesem Szenario entstand zuerst (mit Hilfe der Lipide) eine Protozelle, die dann durch Aufnahme geeigneter Moleküle einen Stoffwechsel entwickelte.
Geschichte
In seinem Buch „The Genetic Code“ stellte 1967 Carl Woese die Hypothese eines auf RNA basierenden einfachen Lebens vor.[38] Auch Leslie Orgel beschrieb 1968 die RNA als eine wichtige Stufe in der Evolution.[39] Der Begriff der „RNA-Welt“ wurde erstmals 1986 von Walter Gilbert gebraucht[40] und hat sich seitdem etabliert.
Literatur
- A. Wochner, J. Attwater u. a.: Ribozyme-catalyzed transcription of an active ribozyme. In: Science. Band 332, Nummer 6026, April 2011, S. 209–212, ISSN 1095-9203. doi:10.1126/science.1200752. PMID 21474753.
- Michael Yarus: Life from an RNA World: The Ancestor Within. Harvard University Press, Cambridge 2010, ISBN 0-674-05075-4.
- Martin Neukamm: Die Entdeckung eines neuen Synthesewegs: Ribonukleotide in der RNA-Welt. In: Laborjournal. 15, Nr. 12, 2. Juni 2009, S. 22–23.
- Martina Preiner: Schöne, alte RNA Welt. In: Spektrum.de. 16. Juni 2016.
Weblinks
- Patrick Cramer: Vom Ursprung der biologischen Evolution - Blick in die Zelle zeigt frühes Protein aus der „RNA-Welt“, LMU München vom 14. November 2017
- Alexandra Kühnlein et al.: tRNA sequences can assemble into a replicator. In: eLife 10 vom 2. März 2021, e63431, doi:10.7554/eLife.63431. Dazu:
- Origin of life: The chicken-and-egg problem. Auf: ScienceDaily vom 2. März 2021.
- Yana Bromberg et al.: Quantifying structural relationships of metal-binding sites suggests origins of biological electron transfer. In: ScienceAdvances, Band 8, Nr. 2, 14. Januar 2022, doi:10.1126/sciadv.abj3984. Dazu:
- David Nield: Study Finds Protein Structures That Could Be Responsible For The Origins of Life. Auf: sciencealert vom 19. Januar 2022.
Einzelnachweise
- Sidney Becker et al.: Unified prebiotically plausible synthesis of pyrimidine and purine RNA ribonucleotides. Science 366, 2019, doi:10.1126/science.aax2747 (Pressemitteilung, Besprechung in Nature News).
- Nilesh Vaidya, Michael L. Manapat, Irene A. Chen, Ramon Xulvi-Brunet, Eric J. Hayden: Spontaneous network formation among cooperative RNA replicators. In: Nature. Band 491, Nr. 7422, 2012, S. 72–77, doi:10.1038/nature11549.
- Yarus M: Primordial genetics: phenotype of the ribocyte. In: Annu. Rev. Genet.. 36, 2002, S. 125–51. doi:10.1146/annurev.genet.36.031902.105056. PMID 12429689.
- Powner MW, Gerland B, Sutherland JD: Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. In: Nature. 459, Nr. 7244, Mai 2009, S. 239–42. doi:10.1038/nature08013. PMID 19444213.
- Sidney Becker et al.: A high-yielding, strictly regioselective prebiotic purine nucleoside formation pathway. In: Science. Band 352, 2016, doi:10.1126/science.aad2808.
- Robert F. Service: Chemists find a recipe that may have jump-started life on Earth. Science News, 2018, doi:10.1126/science.aav7778.
- Kruger K, Grabowski PJ, Zaug AJ, Sands J, Gottschling DE, Cech TR: Self-splicing RNA: autoexcision and autocyclization of the ribosomal RNA intervening sequence of Tetrahymena. In: Cell. 31, Nr. 1, November 1982, S. 147–57. PMID 6297745.
- Gysbers R, Tram K, Gu J, Li Y: Evolution of an Enzyme from a Noncatalytic Nucleic Acid Sequence. In: Scientific Reports. 5, 2015, S. 11405. doi:10.1038/srep11405. PMID 26091540. PMC 4473686 (freier Volltext).
- DNAzymes – How Active DNA Biocatalysts That Destroy Unwanted RNA Molecules Work, auf: SciTechDaily vom 29. Dezember 2021. Quelle: Heinrich-Heine University Duesseldorf.
- Unrau PJ, Bartel DP: RNA-catalysed nucleotide synthesis. In: Nature. 395, Nr. 6699, September 1998, S. 260–3. doi:10.1038/26193. PMID 9751052.
- Johnston WK, Unrau PJ, Lawrence MS, Glasner ME, Bartel DP: RNA-catalyzed RNA polymerization: accurate and general RNA-templated primer extension. In: Science. 292, Nr. 5520, Mai 2001, S. 1319–25. doi:10.1126/science.1060786. PMID 11358999.
- Lincoln TA, Joyce GF: Self-sustained replication of an RNA enzyme. In: Science. 323, Nr. 5918, Februar 2009, S. 1229–32. doi:10.1126/science.1167856. PMID 19131595. PMC 2652413 (freier Volltext).
- Yusupov MM, Yusupova GZ, Baucom A, et al.: Crystal structure of the ribosome at 5.5 A resolution. In: Science. 292, Nr. 5518, Mai 2001, S. 883–96. doi:10.1126/science.1060089. PMID 11283358.
- Szostak JW, Bartel DP, Luisi PL: Synthesizing life. In: Nature. 409, Nr. 6818, Januar 2001, S. 387–90. doi:10.1038/35053176. PMID 11201752.
- Patrick Forterre: The two ages of the RNA world, and the transition to the DNA world: a story of viruses and cells. In: Biochimie. Band 87, Nummer 9–10, 2005 Sep-Oct, S. 793–803, doi:10.1016/j.biochi.2005.03.015, PMID 16164990.
- N. A. Kovacs, A. S. Petrov, K. A. Lanier, L. D. Williams: Frozen in Time: The History of Proteins. In: Molecular biology and evolution. Band 34, Nummer 5, Mai 2017, S. 1252–1260, doi:10.1093/molbev/msx086, PMID 28201543, PMC 5400399 (freier Volltext).
- T. R. Cech: The RNA worlds in context. In: Cold Spring Harbor perspectives in biology. Band 4, Nummer 7, Juli 2012, S. a006742, doi:10.1101/cshperspect.a006742, PMID 21441585, PMC 3385955 (freier Volltext).
- P. T. van der Gulik, D. Speijer: How amino acids and peptides shaped the RNA world. In: Life. Band 5, Nummer 1, Januar 2015, S. 230–246, doi:10.3390/life5010230, PMID 25607813, PMC 4390850 (freier Volltext) (Review).
- Vladimir Kubyshkin, Nediljko Budisa: Anticipating alien cells with alternative genetic codes: away from the alanine world!. In: Curr. Op. Biotechnol.. 60, 3. Juli 2019, S. 242–249. doi:10.1016/j.copbio.2019.05.006.
- Vladimir Kubyshkin, Nediljko Budisa: The Alanine World Model for the Development of the Amino Acid Repertoire in Protein Biosynthesis. In: Int. J. Mol. Sci.. 20, Nr. 21, 24. September 2019, S. 5507. doi:10.3390/ijms20215507.
- A. M. Karmali, T. L. Blundell, N. Furnham: Model-building strategies for low-resolution X-ray crystallographic data. In: Acta Cryst.. 65, 2009, S. 121–127. doi:10.1107/S0907444908040006.
- Schöning K, Scholz P, Guntha S, Wu X, Krishnamurthy R, Eschenmoser A: Chemical etiology of nucleic acid structure: the alpha-threofuranosyl-(3'→2') oligonucleotide system. In: Science. 290, Nr. 5495, November 2000, S. 1347–51. PMID 11082060.
- Zhang L, Peritz A, Meggers E: A simple glycol nucleic acid. In: J. Am. Chem. Soc.. 127, Nr. 12, März 2005, S. 4174–5. doi:10.1021/ja042564z. PMID 15783191.
- Böhler C, Nielsen PE, Orgel LE: Template switching between PNA and RNA oligonucleotides. In: Nature. 376, Nr. 6541, August 1995, S. 578–81. doi:10.1038/376578a0. PMID 7543656.
- Nelson KE, Levy M, Miller SL: Peptide nucleic acids rather than RNA may have been the first genetic molecule. In: Proc. Natl. Acad. Sci. USA. 97, Nr. 8, April 2000, S. 3868–71. PMID 10760258. PMC 18108 (freier Volltext).
- Ehrenfreund P, Rasmussen S, Cleaves J, Chen L: Experimentally tracing the key steps in the origin of life: The aromatic world. In: Astrobiology. 6, Nr. 3, Juni 2006, S. 490–520. doi:10.1089/ast.2006.6.490. PMID 16805704.
- Pascal, Robert: A scenario starting from the first chemical building blocks, in: Muriel Gargaud, Jaques Reisse et al.: From Suns to Life: A Chronological Approach to the History of Life on Earth, Springer Science & Business Media 2007, Seiten 163–166, ISBN 0-387-45083-1
- Kunin V: A system of two polymerases - a model for the origin of life. In: Origins of Life and Evolution of the Biosphere. 30, Nr. 5, Oktober 2000, S. 459–466. bibcode:2000OLEB...30..459K. doi:10.1023/A:1006672126867. PMID 11002892.
- L. Li, C. Francklyn, C. W. Carter. Aminoacylating Urzymes Challenge the RNA World Hypothesis. Journal of Biological Chemistry, 2013; 288 (37): 26856 DOI: 10.1074/jbc.M113.496125
- C. W. Carter: What RNA World? Why a Peptide/RNA Partnership Merits Renewed Experimental Attention. In: Life. Band 5, Nummer 1, Januar 2015, S. 294–320, doi:10.3390/life5010294, PMID 25625599, PMC 4390853 (freier Volltext) (Review).
- Peter T.S. van der Gulik et al.: How Amino Acids and Peptides Shaped the RNA World, in: Life, Januar 2015, 5, Seiten 230–246; doi:10.3390/life5010230
- Joseph Flannery Sutherland: B.C of Prehistory, Palaeontology and the Past. Der Autor ist Verfechter der RNA-Peptid-Koevolution, sein Blog enthält zu diesem Thema u. a. folgende Beiträge:
- Updating The RNA World vom 24. Juni 2015 4:03
- Lost In Translation vom 24. Juni 2015 3:18
- A Brilliant New Update For The RNA World Hypothesis vom 17. September 2013 11:28
- Bhavesh H. Patel, Claudia Percivalle, Dougal J. Ritson, Colm D. Duffy, John D. Sutherland: Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. In: Nature Chemistry. 7, Nr. 4, April 2015, S. 301–307. bibcode:2015NatCh...7..301P. doi:10.1038/nchem.2202. PMID 25803468. PMC 4568310 (freier Volltext).
- Robert F. Service: Researchers may have solved origin-of-life conundrum. In: Science. 16. März 2015, abgerufen am 27. September 2017 (doi:10.1126/science.aab0325).
- Jianfeng Xu, Václav Chmela, Nicholas J. Green, David A. Russell, Mikołaj J. Janicki, Robert W. Góra, Rafał Szabla, Andrew D. Bond, John D. Sutherland: Selective prebiotic formation of RNA pyrimidine and DNA purine nucleosides, in: Nature Band 582, 3. Juni 2020, S. 60–66, doi:10.1038/s41586-020-2330-9
- Michael Marshall: First life on Earth may actually have been built from both RNA and DNA, auf: NewScientist vom 3. Juni 2020
- Did Life Emerge in the “Primordial Soup” via DNA or RNA? Surprising Answer From New Research, auf: SciTechDaily vom 7. Juni 2020, Quelle: UK Research and Innovation
- Woese C: The basic nature of the genetic code. In: The Genetic Code. Harper & Row, Januar 1967, ISBN 978-0-06-047176-7, S. 150–178.
- Orgel LE: Evolution of the genetic apparatus. In: J. Mol. Biol.. 38, Nr. 3, Dezember 1968, S. 381–393. PMID 5718557.
- Gilbert W: The RNA World. In: Nature. 319, Februar 1986, S. 618. doi:10.1038/319618a0.