Inoviridae
Inoviridae, informell auch genannt Filamentöse Bakteriophagen (englisch Filamentous bacteriophage, Singular oder Plural)[Anmerkung 1] sind eine Familie von Viren, die Bakterien infizieren (Bakteriophagen). Die Phagen sind nach ihrer meist filamentösen Form benannt, ihre Virionen (Virusteilchen) sind meist lang, dünn und biegsam und sehen aus wie eine wurmartige Kette (was auch ein Stück gekochter Spaghetti erinnert). Sie haben einem Durchmesser von ca. 6 nm bei einer Länge von bis zu ca. 1000–2000 nm.[3][4][5][6][7]
| Inoviridae | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

Virionen des Filamentösen Bakteriophagen fd, | ||||||||||||||
| Systematik | ||||||||||||||
| ||||||||||||||
| Taxonomische Merkmale | ||||||||||||||
| ||||||||||||||
| Wissenschaftlicher Name | ||||||||||||||
| Inoviridae | ||||||||||||||
| Links | ||||||||||||||
|
Die Inoviridae gehören zu den einfachsten Viren, die bekannt sind. Sie haben weit weniger Gene als die klassischen Bakteriophagen der Ordnung Caudovirales (mit Kopf-Schwanz-Struktur, sog. Schwanzbakteriophagen), wie sie von der American Phage Group um Max Delbrück untersucht wurden. Die Einfachheit dieser Familie macht sie zu einem attraktiven Modellsystem, um grundlegende Aspekte der Molekularbiologie zu untersuchen und zu einem bewährten Werkzeug in der Immunologie und Nanotechnologie.
Die Familie enthält 29 offiziell vom International Committee on Taxonomy of Viruses (ICTV) bestätigte Arten (Spezies), die sich auf 23 offizielle Gattungen verteilen.[8][1] Die Auswertung von genomischen und metagenomischen Datensätzen mit Hilfe eines maschinellen Lernansatzes führte jedoch zur Entdeckung von 10.295 Inovirus-ähnlichen Sequenzen mit Treffern in fast allen bakteriellen Phyla (als Wirte) und in praktisch jedem Ökosystem. Dies ist ein starker Hinweis darauf, dass Gruppe von Viren wesentlich vielfältiger und weiter verbreitet ist als ursprünglich angenommen.[7]
Beschreibung
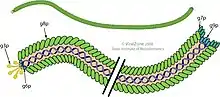
_assembled_major_coat_protein%252C_exploded_view.tif.jpg.webp)
Filamentöse Phagen werden im Gegensatz zu den meisten anderen Phagen kontinuierlich durch die Bakterienmembran extrudiert (ausgeschieden), ohne den bakteriellen Wirt zu töten.[9]
Die Hülle eines Virions besteht aus fünf Arten viraler Proteine, die sich während des Zusammenbaus (Assemblierung) der Phagen-Virionen in der inneren Membran des Wirtsbakteriums befinden und dem heranwachsenden Virion hinzugefügt werden, während es durch die Membran extrudiert wird.
Die von der Gruppe um David Pratt eingeführte einfache Nummerierung der Gene mit den arabischen Ziffern 1,2,3,4, … wird manchmal durch die Praxis der Verwendung der römischen Ziffern I, II, III, IV, … verdrängt, aber die von den beiden Systemen definierten Gennummern sind die gleichen. Für die Akronyme der kodierten Proteine existieren also zwei einander äquivalente Bezeichnungsweisen mit den gleichen Nummern in römischen bzw. in arabischen Ziffern: Protein 1 (pI = g1p), …, Protein 8 (pVIII = g8p), …; die Bezeichnungen der entsprechenden Gene sind analog, jedoch kursiv gesetzt.
Drei filamentöse Bakteriophagen, fd, f1 und M13, wurden in den frühen 1960er Jahren von drei verschiedenen Forschergruppen isoliert und charakterisiert. Sie sind jedoch so ähnlich, dass sie zu einer Gruppe mit dem Namen "Ff-Phagen" zusammengefasst werden, die vom ICTV als Gattung Inovirus offiziell bestätigt wurde.[10][11] Die molekulare Struktur der filamentösen Ff-Phagen wurde mit einer Reihe von physikalischen Techniken, insbesondere der Röntgenfaserbeugung,[4][12] bestimmt und mit Festkörper-NMR und Kryo-Elektronenmikroskopie weiter verfeinert.[4][2] Die einzelsträngige Ff-Phagen-DNA verläuft entlang des zentralen Kerns (englisch cor) des Phagen und wird durch eine zylindrische Proteinhülle geschützt und verdichtet, die aus Tausenden von identischen α-helikalen Untereinheiten des Haupt-Hüllproteins (en. major coat protein) aufgebaut ist, und die vom Phagen-Gen pVIII ([en]) kodiert werden. Das pVIII-Protein wird in die Plasmamembran als ein früher Schritt der Phagen-Assemblierung eingefügt.[4]
Einige Phagenstämme haben eine Leader-Sequenz auf dem pVIII-Protein, um die Einfügung in die Membran zu fördern, aber andere scheinen die Leader-Sequenz nicht zu benötigen. An den beiden Enden des Phagen befinden sich einige Kopien von Proteinen, die für die Infektion des Wirtsbakteriums und auch für den Zusammenbau der wachsenden Phagenpartikel wichtig sind. Diese Proteine sind die Produkte der Phagengene 3 und 6 (pIII und pVI) an einem Ende des Phagen, und der Phagengene 7 und 9 (pVII und pIX) am anderen Ende. Die Faserbeugungsstudien identifizierten zwei strukturelle Klassen von Phagen, die sich in den Details der Anordnung des Gen-8-Proteins (pVIII) unterscheiden. Klasse I, zu der die Stämme fd, f1, M13 der Gattung Inovirus sowie If1 (Spezies Escherichia-Virus If1, Gattung Infulavirus)[13] und IKe (Spezies Salmonella-Virus IKe, Gattung Lineavirus) gehören,[14] hat eine Rotationsachse, die die Gen-8-Hüllproteine (pVIII) miteinander verbindet, während Klasse II, einschließlich der Stämme Pf1 (Spezies Pseudomonas-Virus Pf1, Gattung Primolicivirus),[15] Pf3 (Spezies Pseudomonas-Virus Pf3, Gattung Tertilicivirus),[16] Pf4 (ohne zugewiesene Spezies oder Gattung)[17][18][2] und PH75 (vorgeschlagene Spezies „Thermus-Phage PH75“, incertae sedis innerhalb der Inoviridae),[19] diese Rotationsachse durch eine Helixachse ersetzt ist. Dieser technische Unterschied hat kaum merkliche Auswirkungen auf die Gesamtstruktur des Phagen, aber der Umfang der unabhängigen Beugungsdaten ist für die Symmetrieklasse II größer als für die Klasse I. Dies half bei der Bestimmung der Struktur des Klasse-II-Phagen Pf1,[20] und damit auch der Klasse-I-Struktur.[12]
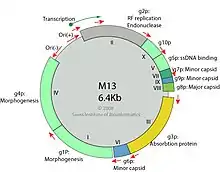
Die aus den fd-Phagen der Gattung Inovirus isolierte DNA ist einzelsträngig und topologisch ein Kreis (zirkulär). Das heißt, der DNA-Einzelstrang erstreckt sich von einem Ende des Phagenpartikels (Virions) zum anderen und dann wieder zurück, um den Kreis zu schließen, obwohl die beiden Stränge nicht basengepaart sind. Es wurde angenommen, dass sich diese Topologie auf alle anderen filamentösen Phagen erstreckt, aber dies ist nicht der Fall für den Phagen Pf4,[17][18][2] bei dem die DNA im Phagen zwar einzelsträngig, aber topologisch linear und nicht kreisförmig ist..[2]
Während des Zusammenbaus (Assemblierung) von fd-Phagen wird die ihre DNA zunächst in einen linearen intrazellulären Nukleoprotein-Komplex mit vielen Kopien des Phagen-Gens 5 (pV, das Replikations-/Assemblierungsprotein) verpackt. Das Gen 5-Protein (pV) wird dann durch das Gen 8-Hüllprotein (pVIII) verdrängt, wenn der heranwachsende Phage durch die bakterielle Plasmamembran extrudiert wird, ohne dabei den bakteriellen Wirt abzutöten.[21][22][4][9] Dieses Protein bindet auch mit hoher Affinität an G-Quadruplex-Strukturen (G4-DNA, „Vier-Strang-DNA“), obwohl diese in der Phagen-DNA nicht vorhanden sind; sowie an ähnliche Haarnadelstrukturen (englisch hairpins) in der Phagen-DNA.[23]
Gen 1 (pI) kodiert für eine ATPase.[24] Das Protein 1 (pI) der Ff-Phagen (d. h. in der Gattung Inovirus) ist für die Phagen-Assemblierung an der Membran erforderlich. Gen 1 (pI) ist daher ein konserviertes Markergen, der (zusammen mit drei weiteren genetischen Merkmalen) zur automatischen Erkennung von Inovirus-Sequenzen verwendet wird.[7] Protein 1 (pI) ist hat eine membranüberspannende hydrophobe Domäne am N-terminalen Teil im Zytoplasma und dem C-terminalen Teil im Periplasma (vgl. ATP-Synthase). Die Monomere bleiben dank der hydrophoben Wechselwirkungen zusammen. Der Assemblierungsmechanismus macht diese Phagen zu einem wertvollen System, um Transmembranproteine zu untersuchen.[4][25][6]
Replikationszyklus
Die Virus-Replikation der Inoviridae erfolgt im Zytoplasma. Die Replikation folgt dem Modell der Rolling-Circle-Replikation für ssDNA.
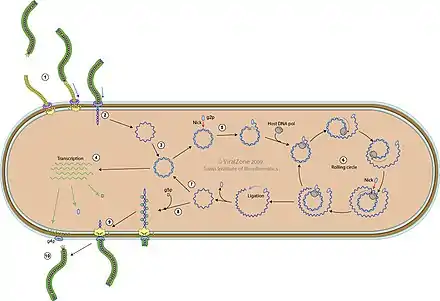
Der Replikationszyklus besteht im Detail aus folgenden Schritten (siehe Schemazeichnung):[26]
- Das virale g3p-Protein (alias pIII, minor coat protein, mCP) vermittelt Anheftung (Adsorption) des Virions an einen Pilus der Wirtszelle. Die Pilus-Retraktion zieht das Virion an die innere Membran des Wirts.
- Die Proteine des Virus-Kapsids vermitteln die Injektion der Virus-DNA durch die bakteriellen Membranen hindurch in das Zellzytoplasma.
- Die Polymerase des Wirtsbakteriums wandelt das (+)ssDNA-Virusgenom (Einzelstrang) in eine kovalent zyklisch geschlossene dsDNA (Doppelstrang) um, die als replikative Form (RF) der DNA (replicative form DNA) bezeichnet wird.
- Durch Transkription der dsDNA mit Hilfe der RNA-Polymerase des Wirts entstehen virale mRNAs.
- Das virale g2p-Protein (alias pII) kappt den RF-DNA-Strang am Replikationsursprung.
- Die Replikation des (+)-Strangs erfolgt nach dem Rolling-Circle-Verfahren.
- Auch neue genomische (+)ssDNA wird in weitere RF-dsDNA umgewandelt, an der ebenfalls Transkription stattfindet.
- Wenn genügend g5p-Protein (alias pIV) synthetisiert wurde, wird die Umwandlung in RF-dsDNA gehemmt, da das neusynthetisierte genomische ssDNA mit g5p bedeckt wird.
- Das g5p wird durch Protein g8p (alias pVIII) ersetzt, um die Assemblierung des viralen Kapsids zu starten.
- Neue Virionen werden aus der Wirtszelle ausgeschieden.
- (nicht dargestellt) Die infizierten Zellen teilen sich weiter und produzieren unbegrenzt viele Virionen.
Die virale Assemblierung erfolgt an der inneren Membran (bei Gram-negativen Bakterien), vermittelt durch einen in die Membran eingebetteten Motorproteinkomplex.[27] Dieser multimere Assemblierungskomplex wird von Gen 1 (pI) kodiert und oft als ZOT (Zonula Occludens Toxin) bezeichnet. Es handelt sich dabei um eine ATPase, die funktionelle und essentielle Walker A- und Walker B-Motife ([en]) enthält,[24] von denen man annimmt, dass sie die Hydrolyse von ATP (Adenosintriphosphat) vermitteln, wodurch die Energie für die Assemblierung des Phagenfilaments geliefert wird. Die Virionen verlassen die Wirtszelle durch virale Extrusion, ohne diese abzutöten.[27] Dies ist ein für Viren der Ordnung Tubulavirales typisch, anders als bei lytischen Bakteriophagen, ihre Wirtszellen am Schluss zum Platzen bringen und abtöten. Auch der lysogene Zyklus, bei dem sich die Viren mit der Wirtszelle vermehren und sich ggf. auch in ihr Genom integrieren, wurde bei den Inoviridae zunächst nicht beobachtet. Vom filamentösen Phage Cf1t der Bakterienspezies Xanthomonas campestris (vorgeschlagene Phagenspezies „Xanthomonas-Phage Cf1t“, incertae sedis innerhalb der Familie Inoviridae, gelegentlich als Cflt verschrieben)[28] wurde allerdings 1987 gezeigt, dass er sich in das bakterielle Wirtsgenom integrieren kann. Seitdem wurden weitere solche temperenten filamentösen Phagen gefunden, von denen viele in die Pathogenese involviert sind, indem sie an sich ungefährlichen Bakterien Toxingene vermitteln, wodurch diese Krankheiten auslösen können.[3] Ein Beispiel für einen solchen Phagen ist der Vibrio-Phage CTXphi
Systematik
Die Taxonomie der filamentösen Bakteriophagen wurde von Andre Lwoff und Paul Tournier als Familie Inophagoviridae, Gattung Inophagovirus, Spezies Inophagovirus bacterii (nach griechisch ἰνός Inos, deutsch ‚Faser oder Faden (Genitiv)‘) definiert, mit dem Phagen fd (Hoffmann-Berling) als Typusart.[29][30] Die Bezeichnung „Phagovirus“ ist tautologisch (alle Phagen sind Viren), und der Name der Familie wurde in Inoviridae und die Typusgattung in Inovirus geändert. Diese Nomenklatur blieb für viele Jahrzehnte bestehen, obwohl die Definition von fd als Typusart ersetzt wurde, als M13 für genetische Manipulationen und für Studien von pVIII breiter eingesetzt wurde.[4][31][32]
Die Phagen fd, f1, M13 (und andere verwandte Phagen) werden oft als Ff-Phagen bezeichnet, mit dem ersten F für F-Plasmid (auch Fertilitätsfaktor genannt, englisch F-episome – sie infizieren Colibakterien, die das F-Episom tragen), und dem zweiten f für filamentös (fadenartig) für F-spezifische (sie infizieren Escherichia coli, die das F-Episom tragen) filamentöse Phagen, unter Verwendung des Konzepts der umgangssprachlichen Namen.[33]
Zu Untersuchungen der Taxonomie der Inoviridae werden zunehmend Phylogenetische Bäume und Kladen verwendet.[3][5][34][7][35] Die Anzahl der bekannten filamentösen Bakteriophagen hat sich durch die Verwendung eines maschinellen Lernansatzes (KI) um ein Vielfaches vermehrt. Daher wurde vorgeschlagen, die Familie Inoviridae neu zu ordnen und in mehrere Familien einer neu geschaffenen Ordnung (den Tubulavirales) aufzuspalten. Vorgeschlagen sind vorläufig 6 Kandidatenfamilien mit 212 Kandidatenunterfamilien.[7]
In die folgende Gliederung der Familie Inoviridae nach ICTV (Master Species List #35)[1] ist diese vorgeschlagene Neuordnung der Familien in der Ordnung Tubulavirales nach Roux et al. (2019) eingearbeitet, jedoch ohne die vorgeschlagenen Unterfamilien (die bisher nur Nummern haben);[7] zusätzlich mit einer Auswahl vorgeschlagener Spezies in Anführungszeichen (nach NCBI mit Stand 15. Februar 2021):
Familie: Inoviridae (veraltet: Inophagoviridae)
- Familie „Amplinoviridae“ (Vorschlag, neu)
- Familie „Densinoviridae“ (Vorschlag, neu)
- Familie „Paulinoviridae“ (Vorschlag, neu)
- Familie „Photinoviridae“ (Vorschlag, neu)
- Familie „Protoinoviridae“ (Vorschlag, bisheriger Name Inoviridae):
- Gattung: Fibrovirus
- Spezies: Vibrio-Virus fs1 (englisch Vibrio virus fs1, Typus)
- Spezies: Vibrio-Virus VGJ (en. Vibrio virus VGJ)
- Spezies „Vibrio-Phage ND1-fs1“ (en. „Vibrio phage ND1-fs1“)
- Spezies „Vibrio-Phage VEJphi“ (en. „Vibrio phage VEJphi“)
- Spezies „Vibrio-Phage VP24-2_Ke“ (en. „Vibrio phage VP24-2_Ke“)
- Gattung: Habenivirus
- Spezies: Ralstonia-Virus RS551 (en. Ralstonia virus RS551)
- Spezies: Ralstonia-Virus RS603 (en. Ralstonia virus RS603)
- Spezies: Ralstonia-Virus RSM1 (en. Ralstonia virus RSM1, Typus)
- Spezies: Ralstonia-Virus RSM3 (en. Ralstonia virus RSM3)
- Spezies „Ralstonia-Virus RSIBR1“ (en. „Ralstonia virus RSIBR1“)
- Gattung: Inovirus (veraltet: Inophagovirus, Ff-Phagen, en. Ff phages)
- Gattung: Lineavirus[14]
- Spezies: Escherichia-Virus I22 (en. Escherichia virus I22)
- Spezies: Salmonella-Virus IKe (en. Salmonella virus IKe, Monotypus)
- Phage IKe
- Gattung: Saetivirus
- Spezies: Vibrio-Virus fs2 (en. Vibrio virus fs2, Typus)
- Spezies: Vibrio-Virus VFJ (en. Vibrio virus VFJ)
- Gattung: Fibrovirus
- Familie „Vespertilinoviridae“ (Vorschlag, bisher Plectroviridae):
- Gattung: Plectrovirus
- Spezies: Acholeplasma-Virus L51 (en. Acholeplasma virus L51, Monotypus)
- Gattung: Vespertiliovirus
- Spezies: Spiroplasma-Virus C74 (en. Spiroplasma virus C74)
- Spezies: Spiroplasma-Virus R8A2B (en. Spiroplasma virus R8A2B, Typus)
- Spezies: Spiroplasma-Virus SkV1CR23x (en. Spiroplasma virus SkV1CR23x)
- Gattung: Plectrovirus
- ohne Vorschlag für eine neue Familienzuordnung, bisher zu Inoviridae:
- Gattung: Affertcholeramvirus[37]
- Spezies: Vibrio-Virus CTXphi (englisch Vibrio virus CTXphi, Monotypus)
- Vibrio-Phage CTXphi (en. CTXφ bacteriophage, nicht zu verwechseln mit Pseudomonas-Phage ΦCTX, Spezies Pseudomonas-Virus phiCTX, Myoviridae)[38] nicht zu verwechseln mit Pseudomonas-Phage ΦCTX, Spezies Pseudomonas-Virus phiCTX, Myoviridae[38]
- Spezies: Vibrio-Virus CTXphi (englisch Vibrio virus CTXphi, Monotypus)
- Gattung: Bifilivirus
- Spezies: Propionibacterium-Virus B5 (en. Propionibacterium virus B5, Monotypus)
- Gattung: Capistrivirus
- Spezies: Vibrio-Virus KSF1 (en. Vibrio virus KSF1, Monotypus)
- Gattung: Coriovirus
- Spezies: Xanthomonas-Virus Cf1c (en. Xanthomonas virus Cf1c, mit Ziffer 1 in der Mitte von Vf1c, Monotypus)
- Phage XacF1[39]
- Spezies: Xanthomonas-Virus Cf1c (en. Xanthomonas virus Cf1c, mit Ziffer 1 in der Mitte von Vf1c, Monotypus)
- Gattung: Infulavirus[13]
- Gattung: Parhipatevirus
- Spezies: Ralstonia-Virus PE226 (en. Ralstonia virus PE226, Monotypus)
- Gattung: Primolicivirus[15]
- Spezies: Pseudomonas-Virus Pf1 (en. Pseudomonas virus Pf1, Monotypus, infiziert Pseudomonas aeruginosa)
- Pseudomonas-Phage Pf1 alias Phage Pf1
- Spezies: Pseudomonas-Virus Pf1 (en. Pseudomonas virus Pf1, Monotypus, infiziert Pseudomonas aeruginosa)
- Gattung: Psecadovirus
- Spezies: Stenotrophomonas-Virus PSH1 (en. Stenotrophomonas virus PSH1, Monotypus)
- Gattung: Restivirus
- Spezies: Ralstonia-Virus RSS1 (en. Ralstonia virus RSS1, Monotypus)
- Gattung: Scuticavirus
- Spezies: Stenotrophomonas-Virus SMA6 (en. Stenotrophomonas virus SMA6, Monotypus)
- Gattung: Staminivirus
- Spezies: Stenotrophomonas-Virus SMA9 (en. Stenotrophomonas virus SMA9, Monotypus)
- Gattung: Subteminivirus
- Spezies: Stenotrophomonas-Virus SMA7 (en. Stenotrophomonas virus SMA7, Monotypus)
- Gattung: Tertilicivirus[16]
- Spezies: Pseudomonas-Virus Pf3 (en. Pseudomonas virus Pf3, Monotypus, infiziert Pseudomonas aeruginosa)
- Gattung: Thomixvirus
- Spezies: Thermus-Virus OH3 (en. Thermus virus OH3, Monotypus)
- Gattung: Versovirus
- Spezies: Vibrio-Virus VfO3K6 (en. Vibrio virus VfO3K6, Monotypus)
- Gattung: Vicialiavirus
- Spezies: Vibrio-Virus VCY (en. Vibrio virus VCY, Monotypus)
- Gattung: Villovirus
- Spezies: Vibrio-Virus Vf33 (en. Vibrio virus Vf33, Monotypus)
- Gattung: Xylivirus
- Spezies: Xanthomonas-Virus Xf109 (en. Xanthomonas virus Xf109, Monotypus)
- ohne Gattungszuweisung, bisher vorgeschlagen für Inoviridae:[40]
- Spezies: „Enterobacteria-Phage ZJ/2“ (en. „Enterobacteria phage ZJ/2“)[41][42]
- Spezies: „Erwinia-Phage PEar1“ (en. „Erwinia phage PEar1“)
- Spezies: „Erwinia-Phage PEar2“ (en. „Erwinia phage PEar2“)
- Spezies: „Erwinia-Phage PEar4“ (en. „Erwinia phage PEar4“)
- Spezies: „Erwinia-Phage PEar6“ (en. „Erwinia phage PEar6“)
- Spezies: „Inovirus C2“
- Spezies: „Phage M13mp18“ (en. „Phage M13mp18“)
- Spezies: „Phage M13mp7“ (en. „Phage M13mp7“)
- Spezies: „Phage M13mp8“ (en. „Phage M13mp8“)
- Spezies: „Pseudomonas-Phage pf8_ST274-AUS411“ (en. „Pseudomonas phage pf8_ST274-AUS411“)
- Spezies: „Pseudomonas-Phage PfLES“ (en. „Pseudomonas phage PfLES“)
- Spezies: „Ralstonia-Phage 1 NP-2014“ (en. „Ralstonia phage 1 NP-2014“)
- Spezies: „Thermus phage PH75“ (en. „Thermus phage PH75“)[19]
- Phage PH75
- Spezies: „Ralstonia-Phage p12J“ (en. „Ralstonia phage p12J“)
- Spezies: „Ralstonia-Phage RS611“ (en. „Ralstonia phage RS611“)
- Spezies: „Ralstonia-Phage RSBg“ (en. „Ralstonia phage RSBg“)
- Spezies: „Ralstonia-Phage RSS-TH1“ (en. „Ralstonia phage RSS-TH1“)
- Spezies: „Ralstonia-Phage RSS0“ (en. „Ralstonia phage RSS0“)
- Spezies: „Ralstonia-Phage RSS20“ (en. „Ralstonia phage RSS20“)
- Spezies: „Ralstonia-Phage RSS30“ (en. „Ralstonia phage RSS30“)
- Spezies: „Spiroplasma-Phage SVGII3“ (en. „Spiroplasma phage SVGII3“)
- Spezies: „Stenotrophomonas-Phage phiSHP2“ (en. „Stenotrophomonas phage phiSHP2“)
- Spezies: „Thermus-Phage PH75“ (en. „Thermus phage PH75“)
- Spezies: „Thermus-Phage phiOH16“ (en. „Thermus phage phiOH16“)
- Spezies: „Vibrio-Phage f237“ (en. „Vibrio phage f237“)
- Spezies: „Vibrio-Phage K04M1_VK04M1“ (en. „Vibrio phage K04M1_VK04M1“)
- Spezies: „Vibrio-Phage K05K4_VK05K4_1“ (en. „Vibrio phage K05K4_VK05K4_1“)
- Spezies: „Vibrio-Phage K05K4_VK05K4_2“ (en. „Vibrio phage K05K4_VK05K4_2“)
- Spezies: „Vibrio-Phage pre-CTX“ (en. „Vibrio phage pre-CTX“)
- Spezies: „Vibrio-Phage RS1“ (en. „Vibrio phage RS1“)
- Spezies: „Vibrio-Phage VAI1“ (en. „Vibrio phage VAI1“)
- Spezies: „Vibrio-Phage VAI2“ (en. „Vibrio phage VAI2“)
- Spezies: „Vibrio-Phage VALG_phi6“ (en. „Vibrio phage VALG_phi6“)
- Spezies: „Vibrio-Phage VALG_phi8“ (en. „Vibrio phage VALG_phi8“)
- Spezies: „Vibrio-Phage VfO4K68“ (en. „Vibrio phage VfO4K68“)
- Spezies: „Xanthomonas-Phage Cf“ (en. „Xanthomonas phage Cf“)
- Spezies: „Xanthomonas-Phage Cf16“ (en. „Xanthomonas phage Cf16“)
- Spezies: „Xanthomonas-Phage Cf1t“ (en. „Xanthomonas phage Cf1t“)[28]
- Phage Cf1t (mit Ziffer 1, gernbe verschrieben als Cflt)
- Spezies: „Xanthomonas-Phage Cf2“ (en. „Xanthomonas phage Cf2“)
- Spezies: „Xanthomonas-Phage Lf“ (en. „Xanthomonas phage Lf“)
- Spezies: „Xanthomonas-Phage phiLF“ (en. „Xanthomonas phage phiLF“, φLf)[43][44]
- Spezies: „Xanthomonas-Phage phiLf2“ (en. „Xanthomonas phage phiLf2“)
- Spezies: „Xanthomonas-Phage phiXo“ (en. „Xanthomonas phage phiXo“)
- Spezies: „Xanthomonas-Phage phiXv“ (en. „Xanthomonas phage phiXv“)
- Spezies: „Xanthomonas-Phage phiXv2“ (en. „Xanthomonas phage phiXv2“)
- Spezies: „Xanthomonas-Phage XaF13“ (en. „Xanthomonas phage XaF13“)
- Spezies: „Xanthomonas-Phage Xf“ (en. „Xanthomonas phage Xf“)
- Spezies: „Xanthomonas-Phage Xf409“ (en. „Xanthomonas phage Xf409“)
- Spezies: „Inoviridae-Spezies ctBZ32“ (en. „Inoviridae sp. ctBZ32“)
- Gattung: Affertcholeramvirus[37]
- ohne Vorschlag für eine neue Familienzuordnung, bisher zu Plectroviridae:
- Gattung: Suturavirus
- Spezies: Spiroplasma-Virus SVTS2 (en. Spiroplasma virus SVTS2, Monotypus)
- Gattung: Suturavirus
- anderweitig vorgeschlagene Stämme ohne Familienzuordnung:
Forschungsgeschichte
Die Erforschung der heute zu den Inoviridae gestellten Bakteriophagen begann in den frühen 1960er Jahren. Das in elektronenmikroskopischen Aufnahmen zu sehende fadenförmige (filamentöse) Teil wurde zunächst irrtümlich als ein bakterieller Pilus interpretiert. Der Abbau der beobachteten Strukturen mit Hilfe on Ultraschall, wodurch die flexiblen Filamente ungefähr in der Mitte auseinandergebrochen wurden, inaktivierte die Infektiosität, wie für eine fadenförmige Bakteriophagenmorphologie vorhergesagt.[45][46][47] Die ersten drei fadenförmigen Bakteriophagen, fd, f1 und M13, wurden von drei verschiedenen Forschungsgruppen isoliert und charakterisiert. Da sich diese drei Phagen in ihren DNA-Sequenzen um weniger als 2 Prozent unterscheiden, was Änderungen in nur wenigen Dutzend Codons im gesamten Genom entspricht, können sie für viele Zwecke als identisch angesehen werden[48] und werden oft mit dem Oberbegriff Ff-Phagen bezeichnet.[33] und heute der gemeinsamen Gattung Inovirus zugeordnet.[10] Die weitere unabhängige Charakterisierung in den folgenden 50 Jahren war von den Interessen dieser Forschungsgruppen und ihrer Anhänger geprägt.[4]
Genetische Studien an M13 unter Verwendung von bedingt letalen Mutanten (Varianten), die von David Pratt und Kollegen initiiert wurden, entschlüsselten die Funktionen der Phagengene.[49][50] So zeigte sich beispielsweise, dass das Proteinprodukt von Gen 5 (g5p bzw. pV), das für die Synthese der einzelsträngigen DNA (ssDNA) der neuen Virionen verantwortlich ist, in großen Mengen in den infizierten Bakterien gebildet wird.[51][52][53] Es bindet an die naszierende (neu gebildete) DNA, um einen linearen intrazellulären Komplex zu bilden.[21]
Das Genom des Phagen fd war eines der ersten vollständigen Genome, die sequenziert wurden.[54]
Anwendung
Filamentöse Bakteriophagen, die so verändert wurden, dass sie immunogene Peptide anzeigen, sind nicht nur in der Immunologie, sondern auch darüber hinaus für weitere biologische Anwendungen nützlich.[55][56][57][58]
Da während der Assemblierung je nach Bedarf mehr (oder weniger) Proteinuntereinheiten hinzugefügt werden können um die DNA zu schützen, kann in fd-Phagen längere (oder kürzere) DNA enthalten sein.[59] Dies macht diese Phagen für genetische Studien besonders geeignet.[60][61]
George Smith und Greg Winter verwendeten f1 und fd für ihre Arbeiten zum Phagen-Display, für die sie einen Teil des Nobelpreises für Chemie 2018 erhielten. Von Angela Belcher und Kollegen stammt die Schaffung und Nutzung vieler Mutanten von M13 für eine breite Palette von Zwecken, insbesondere in der Materialwissenschaft.[58][62][63][64]
Filamentöse Bakteriophagen können die Antibiotikatoleranz ([en]) fördern, indem sie flüssigkristalline Domänen um Bakterienzellen bilden.[65][66][2]
Anmerkungen
- Die Bezeichnung Filamentous phage (bzw. analog Filamentous bacteriophage, deutsch Filamentöser Phage) ist nach ICTV eine veraltete Bezeichnung für die heutige Bezeichnung der Typusgattung Inovirus (ab 1971); entspricht aber heute nach Entdeckung einer großen Zahl weiterer fadenförmiger Bakteriophagen (ggf. mit Plural-s) der ganzen Familie um diese Gattung, also aktuell den Inoviridae.
Einzelnachweise
- ICTV: ICTV Master Species List 2019.v1, New MSL including all taxa updates since the 2018b release, March 2020 (MSL #35)
- Adul K. Tarafder, Andriko von Kügelgen, Adam J. Mellul, Ulrike Schulze, Dirk G. A. L. Aarts, Tanmay A. M. Bharat: Phage liquid crystalline droplets form occlusive sheaths that encapsulate and protect infectious rod-shaped bacteria. In: Proceedings of the National Academy of Sciences of the United States of America (PNAS). 117, Nr. 9, 3. März 2020, S. 4724–4731. doi:10.1073/pnas.1917726117. PMID 32071243. PMC 7060675 (freier Volltext). ePub: 18. Februar 2020. Dazu:
- Yi-Wei Chang: Bacteria suit up with virus armor, in: PNAS 17 (12), 24. März 2020, S.&mnsp;16297-6299, ePub: 9. März 2020, doi:10.1073/pnas.2001931117
- I. D.Hay, T. Lithgow: Filamentous phages: masters of a microbial sharing economy. In: EMBO Reports. 20, Nr. 6, Juni 2019, S. e47427. doi:10.15252/embr.201847427. PMID 30952693. PMC 6549030 (freier Volltext).
- Suzana K. Straus, Htet E. Bo; J. R. Bhella, D. Harris (Hrsg.): Filamentous Bacteriophage Proteins and Assembly. In: Springer Singapore (Hrsg.): Sub-Cellular Biochemistry. 88, 2018, S. 261–279. doi:10.1007/978-981-10-8456-0_12. PMID 29900501. ISBN 978-981-10-8455-3
- Anne Mai-Prochnow, Janice Gee Kay Hui, Staffan Kjelleberg, Jasna Rakonjac, Diane McDougald, Scott A. Rice: Big things in small packages: the genetics of filamentous phage and effects on fitness of their host. In: FEMS Microbiology Reviews. 39, Nr. 4, Juli 2015, S. 465–487. doi:10.1093/femsre/fuu007. PMID 25670735.
- J. Rakonjac, M. Russel, S. Khanum, S. J. Brooke, M. Rajič; T. S. Lim (Hrsg.): Filamentous Phage: Structure and Biology. In: Springer International Publishing (Hrsg.): Advances in Experimental Medicine and Biology. 1053, 2017, S. 1–20. doi:10.1007/978-3-319-72077-7_1. PMID 29549632. ISBN 978-3-319-72076-0
- Simon Roux, Mart Krupovic, Rebecca A. Daly, Adair L. Borges, Stephen Nayfach, Frederik Schulz, Emiley A. Eloe-Fadrosh et al.: Cryptic inoviruses revealed as pervasive in bacteria and archaea across Earth's biomes. In: Nature Microbiology. 4, Nr. 11, November 2019, S. 1895–1906. doi:10.1038/s41564-019-0510-x. PMID 31332386. PMC 6813254 (freier Volltext). ePub 22. Juli 2019, insbes. Supplementary Table 3
- SIB: Viral Zone: Inoviridae. ExPASy. Abgerufen am 15. Februar 2021.
- H. Hoffmann Berling, R. Maze: Release of male-specific bacteriophages from surviving host bacteria. In: Virology. 22, Nr. 3, März 1964, S. 305–313. doi:10.1016/0042-6822(64)90021-2. PMID 14127828.
- NCBI: Inovirus (genus)
- ICTV: ICTV Taxonomy history: Inovirus. 2019 EC 51, Berlin, Germany, July 2019; Email ratification March 2020 (MSL #35).
- NCBI: Infulavirus (genus)
- NCBI: Lineavirus (genus)
- NCBI: Primolicivirus (genus)
- NCBI: Tertilicivirus (genus)
- Jack C. Penner, Jose A. G. Ferreira, Patrick R. Secor, Johanna M. Sweere et al.: Pf4 bacteriophage produced by Pseudomonas aeruginosa inhibits Aspergillus fumigatus metabolism via iron sequestration, in: Microbiology 162(9), September 2016, S. 1583–1594, Epub 29. Juli 2016, doi: 10.1099/mic.0.000344, PMID 27473221
- Patrick R. Secor, Johanna M. Sweere, Lia A. Michaels, Andrey V. Malkovskiy et al.: Filamentous Bacteriophage Promote Biofilm Assembly And Function, in: Cell Host & Microbe, Band 18, Nr. 5, S. 549–559, 11. November 2015, doi:10.1016/j.chom.2015.10.013
- NCBI: Thermus phage PH75 (species)
- Pratt D, Laws P, Griffith J: Complex of bacteriophage M13 single-stranded DNA and gene 5 protein. In: Journal of Molecular Biology. 82, Nr. 4, Februar 1974, S. 425–439. doi:10.1016/0022-2836(74)90239-3. PMID 4594145.
- C. W. Gray: Three-dimensional structure of complexes of single-stranded DNA-binding proteins with DNA. IKe and fd gene 5 proteins form left-handed helices with single-stranded DNA. In: Journal of Molecular Biology. 208, Nr. 1, Juli 1989, S. 57–64. doi:10.1016/0022-2836(89)90087-9. PMID 2671388.
- J. D. Wen, D. M. Gray: Ff gene 5 single-stranded DNA-binding protein assembles on nucleotides constrained by a DNA hairpin. In: Biochemistry. 43, Nr. 9, März 2004, S. 2622–2634. doi:10.1021/bi030177g. PMID 14992600.
- Belinda Loh, Maximilian Haase, Lukas Mueller, Andreas Kuhn, Sebastian Leptihn: The Transmembrane Morphogenesis Protein gp1 of Filamentous Phages Contains Walker A and Walker B Motifs Essential for Phage Assembly. In: Viruses. 9, Nr. 4, 9. April 2017, S. 73. doi:10.3390/v9040073.
- M. P. Rapoza, R. L. Webster: The Products of Gene I and the Overlapping in-Frame Gene XI are Required for Filamentous Phage Assembly. In: J. Molec Biol.. 248, 1995, S. 627–638.
- SIB: Inoviridae — replication cycle, ViralZone
- Belinda Loh, Andreas Kuhn, Sebastian Leptihn: The fascinating biology behind phage display: filamentous phage assembly. In: Molecular Microbiology. 111, Nr. 5, Mai 2019, S. 1132–1138. doi:10.1111/mmi.14187.
- NCBI: Xanthomonas phage Cf1t (species)
- A. Lwoff, P. Tournier: The classification of viruses. In: Annual Review of Microbiology. 20, Nr. 1, 1966, S. 45–74. doi:10.1146/annurev.mi.20.100166.000401. PMID 5330240.
- R. E. Matthews: Classification and nomenclature of viruses. Fourth report of the International Committee on Taxonomy of Viruses. In: Intervirology. 17, Nr. 1–3, 1982, S. 1–199. doi:10.1159/000149278. PMID 6811498.
- Joachim Messing: Cloning in M13 phage or how to use biology at its best. In: Gene. 100, 1991, ISSN 0378-1119, S. 3–12. doi:10.1016/0378-1119(91)90344-B.
- Joachim Messing: Cloning Single-Stranded DNA. In: Molecular Biotechnology. 5, 1996, S. 39–47.
- A. J. Gibbs, B. D. Harrison, D. H. Watson, P. Wildy: What's in a virus name?. In: Nature. 209, Nr. 5022, Januar 1966, S. 450–454. bibcode:1966Natur.209..450G. doi:10.1038/209450a0. PMID 5919575.
- International Committee on Taxonomy of Viruses Executive Committee: The new scope of virus taxonomy: partitioning the virosphere into 15 hierarchical ranks. In: Nature Microbiology. 5, Nr. 5, Mai 2020, S. 668–674. doi:10.1038/s41564-020-0709-x. PMID 32341570. PMC 7186216 (freier Volltext).
- D. Kazlauskas, A. Varsani, E. V. Koonin, M. Krupovic: Multiple origins of prokaryotic and eukaryotic single-stranded DNA viruses from bacterial and archaeal plasmids. In: Nature Communications. 10, Nr. 1, Juli 2019, S. 3425. doi:10.1038/s41467-019-11433-0. PMID 31366885. PMC 6668415 (freier Volltext).
- William O. Salivar. Helen Tzagoloff, David Pratt: Some physical-chemical and biological properties of the rod-shaped coliphage M13, in: Virology, Band 24, Nr. 3, November 1964, S. 359–371, doi:10.1016/0042-6822(64)90173-4
- NCBI: Affertcholeramvirus (genus)
- SIB: Viral exotoxin
- Rakonjac J, Bas B, Derda R (Hrsg.): Filamentous Bacteriophage in Bio/Nano/Technology, Bacterial Pathogenesis and Ecology (= Frontiers Research Topics). Frontiers Media SA, 2017, ISBN 978-2-88945-095-4, doi:10.3389/978-2-88945-095-4.
- NCBI: unclassified Inoviridae
- NCBI: Enterobacteria phage ZJ/2 (species)
- Mai-Prochnow A, Hui JG, Kjelleberg S, Rakonjac J, McDougald D, Rice SA: Big things in small packages: the genetics of filamentous phage and effects on fitness of their host. In: FEMS Microbiology Reviews. 39, Nr. 4, Juli 2015, S. 465–487. doi:10.1093/femsre/fuu007. PMID 25670735.
- NCBI: Xanthomonas phage phiLF (species)
- Nien-Tsung Lin, Tzu-Jun Liu, Tze-Ching Lee, Bih-Yuh You, Ming-Haw Yang, Fu-Shyan Wen, Yi-Hsiung Tseng: The Adsorption Protein Genes of Xanthomonas campestris Filamentous Phages Determining Host Specificity, in: J Bacteriol. Band 181, Nr. 8, April 1999 S. 2465–2471, doi:10.1128/JB.181.8.2465-2471.1999, PMC 93672 (freier Volltext), PMID 10198010
- Johanna M. Sweere et al.: Bacteriophage trigger antiviral immunity and prevent clearance of bacterial infection, Science Band 363, Nr. 6434, 29. März 2019, eaat9691, doi:10.1126/science.aat9691 – über Pseudomonas aeruginosa filamentous phages (Pf-Phagen), Inoviridae. Dazu:
- UM researchers publish new discoveries on bacterial viruses, auf: EurekAlert!, 1. April 2019, Quelle: University of Montana
- David Freifelder, Peter F. Davison: Studies on the sonic degradation of deoxyribonucleic acid. In: Biophysical Journal. 2, Nr. 3, Mai 1962, S. 235–247. bibcode:1962BpJ.....2..235F. doi:10.1016/S0006-3495(62)86852-0. PMID 13894963. PMC 1366369 (freier Volltext).
- D. A. Marvin, H. Hoffmann-Berling: Physical and Chemical Properties of Two New Small Bacteriophages. In: Nature. 197, Nr. 4866, 1963, S. 517–518. bibcode:1963Natur.197..517M. doi:10.1038/197517b0.
- O. Morag, G. Abramov, A. Goldbourt: Similarities and differences within members of the Ff family of filamentous bacteriophage viruses. In: The Journal of Physical Chemistry B. 115, Nr. 51, Dezember 2011, S. 15370–15379. doi:10.1021/jp2079742. PMID 22085310.
- D. Pratt, H. Tzagoloff, W. S. Erdahl: Conditional lethal mutants of the small filamentous coliphage M13. I. Isolation, complementation, cell killing, time of cistron action. In: Virology. 30, Nr. 3, November 1966, S. 397–410. doi:10.1016/0042-6822(66)90118-8. PMID 5921643.
- D. Pratt, H. Tzagoloff, J. Beaudoin: Conditional lethal mutants of the small filamentous coliphage M13. II. Two genes for coat proteins. In: Virology. 39, Nr. 1, September 1969, S. 42–53. doi:10.1016/0042-6822(69)90346-8. PMID 5807970.
- D. Pratt, W. S. Erdahl: Genetic control of bacteriophage M13 DNA synthesis. In: Journal of Molecular Biology. 37, Nr. 1, Oktober 1968, S. 181–200. doi:10.1016/0022-2836(68)90082-X. PMID 4939035.
- T. J. Henry, D. Pratt: The proteins of bacteriophage M13. In: Proceedings of the National Academy of Sciences of the United States of America. 62, Nr. 3, März 1969, S. 800–807. doi:10.1073/pnas.62.3.800. PMID 5257006. PMC 223669 (freier Volltext).
- B. Alberts, L. Frey, H. Delius: Isolation and characterization of gene 5 protein of filamentous bacterial viruses. In: Journal of Molecular Biology. 68, Nr. 1, Juli 1972, S. 139–152. doi:10.1016/0022-2836(72)90269-0. PMID 4115107.
- Beck E, Sommer R, Auerswald EA, Kurz C, Zink B, Osterburg G, Schaller H, Sugimoto K, Sugisaki H, Okamoto T, Takanami M: Nucleotide sequence of bacteriophage fd DNA. In: Nucleic Acids Research. 5, Nr. 12, Dezember 1978, S. 4495–4503. doi:10.1093/nar/5.12.4495. PMID 745987. PMC 342768 (freier Volltext).
- G. P. Smith: Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. In: Science. 228, Nr. 4705, Juni 1985, S. 1315–1317. doi:10.1126/science.4001944. PMID 4001944.
- A. Prisco, F. De Berardinis: Filamentous bacteriophage fd as an antigen delivery system in vaccination. In: International Journal of Molecular Sciences. 13, Nr. 4, 24. April 2012, S. 5179–5194. doi:10.3390/ijms13045179. PMID 22606037. PMC 3344273 (freier Volltext).
- M. Sioud: Phage Display Libraries: From Binders to Targeted Drug Delivery and Human Therapeutics. In: Molecular Biotechnology. 61, Nr. 4, April 2019, S. 286–303. doi:10.1007/s12033-019-00156-8. PMID 30729435.
- K. A. Henry, M. Arbabi-Ghahroudi, J. K. Scott: Beyond phage display: non-traditional applications of the filamentous bacteriophage as a vaccine carrier, therapeutic biologic, and bioconjugation scaffold. In: Frontiers in Microbiology. 6, 2015, S. 755. doi:10.3389/fmicb.2015.00755. PMID 26300850. PMC 4523942 (freier Volltext).
- J. Greenwood, G. J. Hunter, R. N. Perham: Regulation of filamentous bacteriophage length by modification of electrostatic interactions between coat protein and DNA. In: Journal of Molecular Biology. 217, Nr. 2, Januar 1991, S. 223–227. doi:10.1016/0022-2836(91)90534-d. PMID 1992159.
- R. Herrmann, K. Neugebauer, H. Zentgraf, H. Schaller: Transposition of a DNA sequence determining kanamycin resistance into the single-stranded genome of bacteriophage fd. In: Molecular & General Genetics. 159, Nr. 2, Februar 1978, S. 171–178. doi:10.1007/bf00270890. PMID 345091.
- Sadia Sattar, Nicholas J. Bennett, Wesley X. Wen, Jenness M. Guthrie, Len F . Blackwell, James F. Conway, Jasna Rakonjac: Ff-nano, short functionalized nanorods derived from Ff (f1, fd, or M13) filamentous bacteriophage. In: Frontiers in Microbiology. 6, 2015, ISSN 1664-302X. doi:10.3389/fmicb.2015.00316.
- J. P. Casey, R. J. Barbero, N. Heldman, A. M. Belcher: Versatile de novo enzyme activity in capsid proteins from an engineered M13 bacteriophage library. In: Journal of the American Chemical Society. 136, Nr. 47, November 2014, S. 16508–16514. doi:10.1021/ja506346f. PMID 25343220.
- D. Oh, J. Qi, B. Han, G. Zhang, T. J. Carney, J. Ohmura, Y. Zhang, Y. Shao-Horn, A. M. Belcher: M13 virus-directed synthesis of nanostructured metal oxides for lithium-oxygen batteries. In: Nano Letters. 14, Nr. 8, August 2014, S. 4837–4845. doi:10.1021/nl502078m. PMID 25058851.
- Noémie-Manuelle Dorval Courchesne, Matthew T. Klug, Kevin J. Huang, Mark C. Weidman, Victor J. Cantú, Po-Yen Chen, Steven E. Kooi, Dong Soo Yun, William A. Tisdale, Nicholas X. Fang, Angela M. Belcher, Paula T. Hammond: Constructing Multifunctional Virus-Templated Nanoporous Composites for Thin Film Solar Cells: Contributions of Morphology and Optics to Photocurrent Generation. In: The Journal of Physical Chemistry C. 119, Nr. 25, Juni 2015, ISSN 1932-7447, S. 13987–14000. doi:10.1021/acs.jpcc.5b00295.
- Z. Dogic: Filamentous Phages As a Model System in Soft Matter Physics. In: Frontiers in Microbiology. 7, 30. Juni 2016, S. 1013. doi:10.3389/fmicb.2016.01013. PMID 27446051. PMC 4927585 (freier Volltext).
- Patrick R. Secor, Laura K. Jennings, Lia A. Michaels, Johanna M. Sweere, Pradeep K. Singh, William C. Parks, Paul L. Bollyky: Biofilm assembly becomes crystal clear – filamentous bacteriophage organize the Pseudomonas aeruginosa biofilm matrix into a liquid crystal. In: Microbial Cell. 3, Nr. 1, Dezember 2015, S. 49–52. doi:10.15698/mic2016.01.475. PMID 28357315. PMC 5354590 (freier Volltext).