Bewegungskontrolle
Bewegungskontrolle (englisch motor control[1]) ist ein Begriff aus der Bewegungswissenschaft.
Die Bewegungskontrolle ist ein körperinterner Vorgang, bei dem bei Menschen und Tieren ihr Bewegungssystem so koordiniert wird, dass geplante und ungeplante Bewegungen so ablaufen, dass deren beabsichtigtes Ziel sicher erreicht wird. Keine körperliche Bewegung wird ohne Bewegungskontrolle ausgeführt. Sie garantiert dem Ausführenden die physiologische Sicherheit des Ablaufs und spielt eine wichtige Rolle bei der Ausführung und beim Lernen von Bewegungsabläufen. Dabei ist sie dem Ausführenden in der Regel nicht bewusst. Sie kann als Überwachung (Monitoring) der Bewegungsabläufe bezeichnet werden. Einbezogen in die Bewegungskontrolle werden Informationen sowohl aus der Umwelt als auch aus dem Körper selbst, die dann durch kooperative Interaktion zwischen dem peripheren und dem zentralen Nervensystem (ZNS) sowie dem Muskelsystem zielgerichtet verarbeitet werden.
In den vergangenen zehn Jahren haben sich die Vorstellungen über die Bewegungskontrolle aufgrund neuerer Erkenntnisse über die Hirnstrukturen stark geändert. Lange Zeit ging man davon aus, dass für Bewegungen Kommandos vom Gehirn ausgehen, und nur dann, wenn etwas den Ablauf stört oder behindert, eine Korrektur vorgenommen wird. Heute weiß man, dass durch diese Kommandos, die eine Bewegung auslösen, gleichzeitig ein ganzes Netzwerk der Bewegungskontrolle aktiviert wird. Das bedeutet, dass die ständig eintreffenden Informationen über die Umgebung und den inneren Zustand der Organismus auf die erfolgreiche Ausführung dieses Bewegungsablaufs ausgerichtet wird.[2] Das bedeutet, dass ein Bewegungsablauf während seiner Ausführung unter einer ständigen online Kontrolle steht.
Bei der Erforschung der Bewegungskontrolle arbeiten zahlreiche Disziplinen zusammen: von der Biomechanik, Mechanik, Physik und den Ingenieurwissenschaften bis zur Psychologie, den Verhaltens- und den Neurowissenschaften.
Begriffsgeschichte
In Deutschland wurde Bewegungskontrolle lange Zeit so verstanden, dass der Lehrende die Ausführung des Lernenden kontrolliert in dem Sinne, dass Fehler, nicht gewünschte Komponenten oder Qualitäten einer Bewegungsausführung erkannt und genannt werden, die bei folgenden Ausführungen vermieden werden oder zusätzlich beachtet werden sollen. Das heißt, eine Kontrolle wurde als von außen gegeben verstanden.
Mit Bewegungskontrolle ist heute in der Bewegungswissenschaft die interne, vom Organismus selbst durchgeführte Kontrolle des Ablaufs gemeint. Sie entspricht dem in der Technik eingesetzten Verfahren der Regelung eines Prozesses – Das ist jedoch im menschlichen Organismus kein technischer, sondern ein biologischer Vorgang. Der Begriff ist unmittelbar mit dem der Rückmeldung beziehungsweise Rückkoppelung eines (Teil-)Ergebnisses des Ablaufs verbunden, denn durch die Rückmeldung kann eine Differenz zwischen der erfolgten und der beabsichtigten Durchführung festgestellt werden, die es dann zu verringern gilt.
Der Kontrollprozess
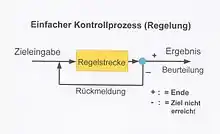
Bei einer Regelung (Kontrollprozess) wird ein Ziel für einen Prozess vorgegeben (input). Auf der sogenannten Regelstrecke soll durch vorgegebene oder konstruierte Abläufe dieses Ziel erarbeitet werden. Sind die Abläufe abgeschlossen, wird das Ergebnis (output) beurteilt (gemessen). Ist das Ziel erreicht, ist der Prozess abgeschlossen. Ist es nicht erreicht, kann durch Korrekturvorgaben und eine neue Eingabe an die Regelstrecke (veränderte Abläufe) der Versuch erfolgen, das Ziel in erneuten Ablauf zu erreichen. Dieser Kreislauf kann so lange wiederholt werden, bis das Ziel erreicht ist. Je früher in ihrem Ablauf eine Abweichung des geplanten Ablaufs entdeckt wird, desto größer ist die Chance, durch gezielte Korrekturmaßnahmen das Ziel dennoch sicher und in der geplanten Zeit zu erreichen. Für all diese Abläufe ist es wichtig, dass die notwendigen Informationen – über Ziel, aktuelle Zustände, Teilergebnisse – bereitgestellt und zur richtigen Zeit an den richtigen Orten zur Verfügung stehen. Insofern hat Bewegungskontrolle sehr viel mit Informationsaufnahme und Informationsverarbeitung im menschlichen Organismus zu tun. Diese Aufgaben werden durch das Nervensystem erfüllt.
Bei der Bewegungsregelung im menschlichen Organismus – wie sie lange Zeit verstanden wurde – wird das Ziel vom Ausführenden selbst oder von einem Lehrer (Trainer, Therapeuten) bestimmt. Die Regelstrecke ist das Bewegungssystem des Menschen. Die Abläufe sind die Ausführungen von motorischen (Teil-)Handlungen. Die Beurteilung, ob das Ziel erreicht ist oder nicht, erfolgt für den bewusst wahrnehmbaren Teil vom Ausführenden, Lehrer oder Trainer, für die internen Kontrollmechanismen wird auch dies intern vom Organismus geleistet (heutiges Wissen).
Warum ist Bewegungskontrolle notwendig?
Bei den ersten Annahmen über die Ausführung von Bewegungsabläufen – noch bis etwa Mitte des 20. Jahrhunderts – fand man heraus, dass die vom Großhirn erteilten die Befehle unmittelbar an die Motoneurone im Rückenmark und von dort an die entsprechenden Muskeln geleitet (Pyramidalsystem[3]) und man nahm an, dass sie und dort genauso, wie geplant ausgeführt werden – wie bei einem früheren (wie damals üblich: starren) Computerprogramm. Dahinter stand auch die Beobachtung, dass ein Bewegungsablauf, wenn er beherrscht wird, die visuelle Wahrnehmung eines stereotypen, invarianten Ablaufs vermittelt (automatisierter Bewegungsablauf, der vermeintlich ohne Kontrolle abläuft). Dem Pyramidalsystem gegenüber stand das Extrapyramidalsystem, dem alle neuronalen Verbindungen zugeordnet wurden, die nicht zum Pyramidalsystem gehörten[4]. Ihre Funktionsweise erfolgt „automatisch“. Das bedeutet, ohne bewusste Kontrolle. Es war aber klar, dass sie dazu beitragen, dass das Gesamtsystem der Bewegung sicher funktioniert. Heute sind viele im so genannten Extrapyramidalsystem vorhandenen Regelkreise erforscht und man weiß, welchen Anteil sie an der Bewegungskontrolle der Wirbeltiere (damit auch des Menschen) haben. Es handelt sich hauptsächlich um neuronale Netzwerke für grundlegende Bewegungen, die sich im Laufe der Evolution entwickelt haben. Sie werden als Zentrale Mustergeneratoren (Central Pattern Generator, CPGs) bezeichnet. Sie spielen eine wichtige Rolle bei der Bewegungskontrolle. Ihre Funktionen und ihr Zusammenwirken mit der Willkürmotorik sind Gegenstand[5] intensiver Forschung.
Wenn man jedoch bedenkt, dass auch die Signal- beziehungsweise Informationsvermittlung im Organismus, ganz zu schweigen vom Auftreten und Erkennen der Signale aus der Umwelt mit Ungenauigkeiten, ja sogar mit Fehlern behaftet ist[6], wird deutlich, dass für eine korrekte, vor allem aber eine sichere Ausführung Korrekturmaßnahmen notwendig sind. Die Bewegungskontrolle dient dazu, solche Fehler – vielleicht sogar nur möglicherweise auftretende Ungenauigkeiten und Fehler – rechtzeitig zu erkennen, ihre Korrektur einzuleiten und zu überwachen.
Gleichgewichtskontrolle
Der Gleichgewichtssinn und vor allem eine Störung des Gleichgewichts gehört zu sehr frühen Interessen nicht nur der Neurophysiologie, sondern der Medizin generell. Ein Problem in der Medizin ist es deswegen, weil es die Gesundheit und damit die Lebensqualität (Körperliche Sicherheit) eines Menschen betrifft. Es wurde deswegen auch nicht unter dem Gesichtspunkt einer technisch/biologischen Kontrolle betrachtet und untersucht, sondern wegen des gesundheitlichen Risikos.
Das Problem der Gleichgewichtskontrolle ist sehr komplex. Untersuchungen zu diesem Komplex sind lange Zeit nicht im Sinne der Bewegungsforschung durchgeführt worden, sondern aus dem medizinischen Aspekt, herauszufinden, warum Menschen Gleichgewichtsstörungen haben und wie man ihnen helfen kann, die Folgen möglichst zu vermeiden (zum Beispiel Stürze) und mit den Folgen umzugehen. Die Erkenntnisse aus dieser Forschung haben inzwischen jedoch eine zentrale Bedeutung bei der Gesamtbetrachtung der Bewegungskontrolle, weil ihre Mechanismen grundlegend auch für die Kontrolle anderer Bewegungsabläufe ist.
Warum ist Bewegungskontrolle nötig?
Bei den ersten Annahmen über die Ausführung von Bewegungsabläufen – noch bis etwa Mitte des 20. Jahrhunderts – stellte man sich vor, dass die vom Großhirn erteilten Befehle unmittelbar an die Motoneurone im Rückenmark, an die entsprechenden Muskeln geleitet und dort genauso, wie geplant ausgeführt werden – wie bei einem früheren (wie damals üblich: starren) Computerprogramm. Dahinter stand auch die Beobachtung, dass ein Bewegungsablauf, wenn er beherrscht wird, die optische Wahrnehmung eines stereotypen, invarianten Ablaufs vermittelt (automatisierter Bewegungsablauf, der vermeintlich ohne Kontrolle abläuft). Wenn man jedoch bedenkt, dass auch die Signal- beziehungsweise Informationsvermittlung im Organismus, ganz zu schweigen vom Auftreten und Erkennen der Signale aus der Umwelt mit Ungenauigkeiten, ja sogar mit Fehlern behaftet ist, wird deutlich, dass für eine korrekte, vor allem aber eine sichere Ausführung der Korrekturmaßnahmen notwendig sind. Die Bewegungskontrolle dient dazu, solche Fehler – vielleicht sogar nur möglicherweise auftretende Ungenauigkeiten und Fehler – rechtzeitig zu erkennen, ihre Korrektur einzuleiten und zu überwachen.
Die beteiligten Forschungsdisziplinen
Neurophysiologie, Neurowissenschaften
Die Neurophysiologie beschäftigt sich mit der Anatomie und der Physiologie des zentralen und des peripheren Nervensystems. Das Zusammenwirken von Nerventätigkeit (Elektrizität) und Bewegung wurde bereits im 18. Jahrhundert von Luigi Galvani entdeckt.[7] Eine kontrollierte Erforschung dieser Phänomene fand damals noch nicht statt. Erst zu Beginn des 19. Jahrhunderts begann man mit Experimenten über gezielte elektrische Reizung und Reflexantworten auf Reize einzelner Sinnesorgane.[8]
Durch neue Erkenntnisse zum Beispiel über die Färbung von bestimmten Nervenfasern wurde es zu Beginn des 20. Jahrhunderts möglich, den Verlauf von Nerven zu verfolgen. Damit gelang es Charles Scott Sherrington einzelne Nervenfasern zu verfolgen und auf diese Weise die spinalen Reflexe und auch ihre Bedeutung für eine Regelung von Bewegungen auf spinaler Ebene zu erkennen und zu beschreiben. Heute liefert die Technik zahlreiche Verfahren, mit deren Hilf nicht nur der Verlauf einzelner Nervenfasern verfolgt werden kann, sondern auch der Verlauf einer aktuellen Aktion.
Verhaltenswissenschaften
In der Verhaltensforschung wird das Verhalten von Tieren erforscht, um von den Erkenntnissen bei Tieren auf das Verhalten von Menschen zu schließen. Sie wurde in Deutschland und den USA von unterschiedlichen Mutterdisziplinen aus und mit unterschiedlichen Methoden betrieben.
In Deutschland ist die Verhaltenswissenschaft ein Teilbereich der Tierphysiologie, die Wissenschaftler Biologen (Zoologen) oder Mediziner (Physiologen). Als Begründer gilt Konrad Lorenz. Dieser arbeitete vor allem mit dem Verfahren der reinen Beobachtung von Tieren – ähnlich dem Behaviorismus. Andere Forscher wie Erich von Holst und Bernard Hassenstein untersuchten die Nervensysteme der Tiere, um zum Beispiel herauszufinden, wie diese ihre Bewegungen koordinieren.
Der Begriff der Bewegungskontrolle war in der ersten Hälfte des 20. Jahrhunderts nicht gebräuchlich. Man sprach von Bewegungskoordination oder Bewegungsregulation.
In den USA beschäftigten sich Psychologen mit den Verhaltenswissenschaften. Sie entwickelten die Methoden des Behaviorismus. Das bedeutet, es wurde nur das objektiv beobachtbare Verhalten zur Bildung von Theorien herangezogen. Man versuchte auf diese Weise, Lernprozesse zu verstehen und übertrug die Erkenntnisse auf das Verhalten und das Lernen von Menschen. Man interessierte sich nicht für die Physiologie, die sich innerhalb der Tiere abspielte und zum Zustandekommen der Bewegungen führte, lehnte dies sogar als unwissenschaftlich ab, weil zu wenig darüber bekannt war. Erst als sich die Bewegungswissenschaft (Psycho Motor Behavior) als eigene Disziplin etablierte, wurde die Neurophysiologie in die Erklärung von Verhaltensweisen mit einbezogen.
Ingenieurwissenschaften
In den Ingenieurwissenschaften beschäftigte man sich – mit Ausnahme der Prothesenkonstruktion nach dem Ersten Weltkrieg – zu Beginn des 20. Jahrhunderts nicht mit der Bewegung des Menschen. Die Regelung von technischen Prozessen war bekannt, wurde aber erst in den 1930er-Jahren zu einem Routineverfahren. Heute spielen die Ingenieurwissenschaften mit zahlreichen ihrer Teildisziplinen (zum Beispiel Biomechanik, Messtechnik und Regelungstechnik) eine wichtige Rolle bei der Erforschung der Bewegungskontrolle.
| Bewegungs- wissenschaft | |||||||||||||||||||||||||||||||||||||||||||||||
| Energie- verarbeitung | Informations- verarbeitung | ||||||||||||||||||||||||||||||||||||||||||||||
| Funktionelle Anatomie | Arbeits- physiologie | Biomechanik | Bewegungs- kontrolle | Psychomotorisches Verhalten | Bewegungs- soziologie | ||||||||||||||||||||||||||||||||||||||||||
| Knochen | Atmung | Ergonomie | Bewegungslernen | genetische Vorgaben | Gruppeneinfluss | ||||||||||||||||||||||||||||||||||||||||||
| Gelenke | Kreislauf | Orthopädie | Informations- verarbeitung | Erfahrung | Traditionen | ||||||||||||||||||||||||||||||||||||||||||
| Bänder | Muskel- arbeit | Rehabilitation | Kontrollmechanismen | Wissen | Meinungen | ||||||||||||||||||||||||||||||||||||||||||
| Sehnen | Fitness | Gewebemechanik | Neurologische Strukturen | ||||||||||||||||||||||||||||||||||||||||||||
| Muskeln | Arbeit in Wasser/Hitze | Sport | Aufbau und Funktion des Motoneurons | ||||||||||||||||||||||||||||||||||||||||||||
| Zahnmedizin | Aufbau des Nervensystems aus motorischer Sicht | ||||||||||||||||||||||||||||||||||||||||||||||
| Forensische Biomechanik | Kontrollaufgabe der einzelnen Hirnabschnitte | ||||||||||||||||||||||||||||||||||||||||||||||
Von getrennten Anfängen zur Zusammenarbeit
Zu Beginn des 20. Jahrhunderts entwickelten sich die genannten Disziplinen bezüglich der Bewegungskontrolle getrennt voneinander. Um die Mitte des vorigen Jahrhunderts kam es zu den ersten Zusammenarbeiten zwischen Forschern dieser Disziplinen. Um diese Zeit entstand auch das Bewusstsein der Notwendigkeit einer Zusammenarbeit zwischen Ingenieuren und Physiologen, als in der Technik die Regelungstechnik eine immer bedeutendere Rolle zu spielen begann und Ingenieure und Biologen feststellten, dass es sich in beiden Bereichen um vergleichbare Prozesse handelt. Diese Zusammenarbeit zwischen Ingenieuren, Biologen, Physiologen und Psychologen wird auch in den damals entstandenen integrativen Disziplinen wie der Kybernetik, Synergetik schließlich den Neuronalen Netzen deutlich.
In den USA trugen zu dieser Entwicklung in hohem Maß die Untersuchungen an Piloten für ihren Einsatz im Zweiten Weltkrieg bei.[9] In Deutschland sei hier stellvertretend der Professor für Elektrotechnik Karl Küpfmüller genannt,[10] der die Physiologen unterstützte, die technischen Elemente zu verstehen.
Einen großen Anteil an der Entwicklung der Bewegungskontrolle hat der Verhaltenswissenschaftler Erich von Holst (Physiologe). Im Gegensatz zu Konrad Lorenz, der mit Beobachtungen und zum Teil behavioristischen Methoden arbeitete, konzentrierte sich von Holst auf die (physiologischen) Abläufe im Nervensystem bei den Bewegungen der Tiere, um die Funktionen des Zentralnervensystems dabei zu untersuchen. Er promovierte über die Funktionen des Zentralnervensystems beim Regenwurm.[11] Zukunftsweisend waren seine Untersuchungen zum Reafferenzprinzip[12]. Er bediente sich nicht der Sprache der Ingenieure, obwohl es sich deutlich um Regelungsprozesse handelte, die er beschrieb. Das ist auch ein Grund dafür, dass die Bedeutung dieser Arbeit für die Bewegungskontrolle erst in den 1980er Jahren erkannt wurde. Ein anderer Grund ist, dass seine Arbeiten nicht oder nur unvollständig ins Englische übersetzt wurden und daher dort, wo man sich intensiver um das Verständnis der Bewegungskontrolle bemühte, lange nicht bekannt waren.
Die Abläufe der Bewegungskontrolle wurden vor allen Dingen in den USA untersucht und beschrieben. Ihr Studium begann aber auch dort, bedingt durch die lange Zeit der Vorherrschaft der behavioristischen Lern- und Verhaltensforschung in der Psychologie, erst nach 1960 in der Motorikforschung eine Rolle zu spielen. In Deutschland ist das Bewusstsein der Bedeutung der Bewegungskontrolle noch immer gering und führt gelegentlich zu Missverständnissen. Auch in den USA bestand zunächst scheinbar lediglich ein allgemeines Interesse daran, zu wissen, wie Bewegung funktioniert – allgemeine Bewegung und Sport spielen in den USA traditionell eine große Rolle. Behavioristisch geprägt sind die Arbeiten von Richard A. Schmidt.[13] Er entwickelte die Schematheorie, durch die der notwendige Speicherbedarf im Hirn geringer ist als bei der Programmtheorie. bei der für jede gelernte Bewegung ein einzelnes Programm gespeichert werden muss. Der Speicherbedarf ist deswegen geringer, weil sozusagen für eine Bewegungsfamilie ein Ausführungsschema vorliegt, das in der aktuellen Situation nicht nur an die notwendige Größe und Ausführungsgeschwindigkeit angepasst werden kann, sondern auch an die Ausführung mit verschiedenen Gliedmaßen. So kann man ein Wort, wenn man es schreiben kann, auch mit der nicht bevorzugten Hand, mit dem Fuß oder mit einem Griffel im Mund schreiben (generalisiertes Schema).
Es wurde dann auch immer mehr versucht, die Experimente so aufzubauen, dass man daraus Rückschlüsse dahingehend ziehen konnte, wie die Bewegungen möglicherweise im Organismus zustande kommen – zunächst im Muskelsystem, dann aber auch im Nervensystem, da dies die Muskelarbeit auslöst und bestimmt. Die Neurophysiologie begann, eine Rolle in der Bewegungsforschung zu spielen. Mit dem Einzug der Informationsverarbeitung in das psychologische Denken durch Lindsay und Norman[14] und der entsprechenden Erweiterung auf den Sport durch Ronald Marteniuk[15] spielten dann auch Überlegungen darüber, wie die Informationen im Organismus bei Bewegungen übermittelt werden, eine immer größere Rolle. Dabei tauchte zum ersten Mal eine interne Rückmeldung auf, die eine Voraussetzung für die Kontrolle einer Bewegung ist. Neu an diesen Ansätzen ist auch, dass man nicht wie im Behaviorismus von der Bewegung als von einer Reaktion auf ein Signal hin ausgeht, sondern dass eine Bewegung willentlich von einer handelnden Person ausgelöst und auf ein Ziel gerichtet ausgeführt werden kann.
Die 1980er Jahre lassen sich als die Jahre bezeichnen, in denen die Motor Control Theory sich als eine eigenständige Bewegungstheorie etablierte, die vorher benutzte Bezeichnungen und Auffassungen ordnete und zusammenfasste. Die Forscher der oben beteiligten Disziplinen traten in einen direkten Austausch miteinander. Es fanden internationale Kongresse statt, in denen die Theorien diskutiert und zukünftige Richtungen erarbeitet wurden. Seit der Zeit wird auch der Begriff der Motor Control einheitlicher für diesen Forschungsbereich verwendet, der andere häufig unklare (wie Bewegungsregulation) oder vieldeutige Begriffe (wie Bewegungskoordination) eine eindeutige Bedeutung zuwies.
Eine der neuen Richtungen der Bewegungstheorie – mit eigenen Ansätzen zur Bewegungskontrolle war z. B. die sogenannte ökologischen Theorie (oder action theory), die von James Gibson[16] und Edward Reed[17] vertreten wurde, und die zum Teil auf Überlegungen und Veröffentlichungen von Nikolai Bernstein (siehe unten) zurückgingen. Diese Theorie besagt, dass nicht das Lebewesen, also auch der Mensch, eine Bewegung initiiert und durchführt, sondern dass die Umgebung, in der es sich befindet, Herausforderungen (affordances) enthält, die ihn anregen bzw. zwingen, bestimmte Aktionen auszuführen, dass also die Umgebung unsere Bewegungen bestimmt.
Es gab verschiedene internationale Kongresse, auf denen diese Ansätze und Überlegungen diskutiert wurden. Einer dieser Kongresse – oder workshop – fand 1986 in Münster statt. Er hatte den Untertitel The Motor-Action Controversy.[18] Hier berichteten hauptsächlich die alten Bewegungsforscher, die noch in der Zeit des Behaviorismus ausgebildet waren, und sich vorsichtig mit informationstheoretischen und rückgekoppelten Ansätzen sowie mit der neuen Action Theory auseinandersetzten (zum Beispiel Richard A. Schmidt). Mehr naturwissenschaftlich ausgerichtet (Neurophysiologie sowie Modellansätze aus der Regelungstechnik) war der 1985 in Varna/Bulgarien abgehaltene Kongress, der bereits Motor Control als seinen Namen hatte.[19]
Bedeutend für die weitere Entwicklung in den englischsprachigen Ländern war außer der genannten action theory die etwa gleichzeitig stattfindende Diskussion über die Arbeiten zur Bewegungsforschung des russischen Physiologen Nikolai Bernstein. Sie gingen zurück auf dessen Buch: The Coordination and Regulation of Movements, das 1967 in England erschienen war.[20] Da sich alle amerikanischen Bewegungswissenschaftler nach dieser Zeit – bis heute – auf dieses Buch berufen, es aber irgendwann vergriffen war und nicht wieder aufgelegt wurde, wurden seine Arbeiten von ausgewählten Bewegungsforschern besprochen und kommentiert und in einem neuen Band Human Motor Actions, Bernstein reassessed von H.T.A. Whiting zusammengefasst und herausgegeben.[21]
Bernstein war im Gegensatz zu den amerikanischen Bewegungsforschern wie fast alle russischen Bewegungsforscher von seiner Ausbildung her Physiologe und Mathematiker/Ingenieur. Er wollte „sein Verständnis für die Funktion des Gehirns durch das Studium der Bewegung fördern“ (He wanted to further his understanding of the brain through the study of movement[22]). Er verfasste mehr als 140 Publikationen, von denen ein Teil ins Deutsche oder Englische übersetzt wurde. Seine bis heute wichtigsten Beiträge zur Bewegungskontrolle sind seine Aussagen über die Redundanz der durch die Anatomie des Körpers gegebenen Bewegungsmöglichkeiten, die einerseits eine hohe Variabilität der möglichen Bewegungen zulässt, die aber andererseits den Organismus zwingen, die hohe Zahl der Freiheitsgrade einer Bewegung so zu reduzieren und zu kontrollieren, dass eine koordinierte Bewegung möglich wird. Seine Ausdrucksweise war entsprechend seiner Ausbildung stark mathematisch geprägt, so dass in der Folge auch Ingenieure in den USA und England sich für die Kontrolle der menschlichen Bewegung interessierten, und versuchten, ihre Ansichten und Erkenntnisse durch spezielle Experimente zu zeigen und zu vertiefen. Allmählich verlagerte sich der Schwerpunkt der Forschung, neben der Neurophysiologie, in der man vor allem versuchte, die für die Kontrolle notwendigen schnellen Informationspfade im Organismus zu finden, in den Forschungsbereich der Ingenieure. Diese versuchen, die Erkenntnisse durch mathematische Modelle darzustellen.
Die Erforschung der Bewegungskontrolle nimmt zu, weil ihre Bedeutung eine immer größere Rolle spielt, zum Beispiel für die Konstruktion und Steuerung künstlicher Gliedmaßen (Rehabilitation), vor allem aber für die Konstruktion lernfähiger Roboter. Die Robotikkonstrukteure interessieren sich dafür, wie die Bewegungskontrolle von Lebewesen organisiert ist, weil die Natur mit Ihrer Millionenjahre langen Evolution die Bewegungen lebender Organismen so organisiert hat, dass sie optimal und damit in vorbildlicher Weise anpassungs- und lernfähig sind und sich durch eine hohe Effizienz und Eleganz auszeichnen.
Theorien über die Bewegungskontrolle
Die Psychologen (vor allem in den USA), die sich zu Beginn des 20. Jahrhunderts nicht mit neurophysiologischen Fragestellungen beschäftigten, gingen bei der Bewegungskontrolle hauptsächlich von einer sogenannten open loop control (offene Kontrollschleife) aus. Bei dieser Vorstellung wird ein Bewegungsablauf geplant, die notwendigen Befehle zu ihrer Ausführung von einem Zentrum (dem Primären Motorischen Kortex, MI) ausgegeben und die Bewegung ausgeführt. Klassisch erfolgt bei dem gesamten Ablauf keinerlei Kontrolle – open loop. Allerdings besteht immer die Möglichkeit, nach der Ausführung diese zu beurteilen – und entsprechend dem Erfolg beziehungsweise Nichterfolg des Ablaufs Korrekturen für eine folgende Ausführung vorzuschlagen beziehungsweise zu planen – vom Ausführenden selber oder von einem Außenstehenden. Im eigentlichen Sinn wird dadurch dann die Kontrollschleife geschlossen.
Man glaubte lange Zeit, dass diese Art der Kontrolle, vor allem bei schnellen (zum Beispiel sogenannten ballistischen Bewegungen) die einzig mögliche ist, weil man davon ausging, dass die gesamte Ausführung zu schnell erfolgt, als dass sie durch interne Kontrollmechanismen beeinflusst werden könnte.[23]
Der Vorteil der open loop control besteht darin, dass im Laufe des Lernprozesses eine Art (wahrnehmbarer) Optimierung des Ablaufs stattfinden kann und dass das Ergebnis, wenn sein Ablauf nicht gestört wird, exakt vorausgesagt werden kann – solange keine Störungen auftreten. Ihr Nachteil liegt aber darin, dass der Ablauf starr und nicht variabel ist und er sich deshalb nicht an geänderte Umgebungs- oder innerkörperliche Bedingungen anpassen kann[24].
Der open loop control steht die closed loop control (geschlossene Kontrollschleife) gegenüber.[25] Man könnte, wie bereits erwähnt, auch eine Korrektur nach der Bewegungsausführung als ein Schließen der Kontrollschleife bei Bewegungen ansehen. Jedoch wird die closed loop control verstanden als die interne Kontrolle des Organismus, die sich während einer Bewegungsausführung abspielt. Um zu verstehen, wie es zu einer solchen Kontrolle kommen kann, ist es notwendig, dass man in den Organismus hineinsieht und die physiologischen Strukturen, die dies leisten können, sucht und beschreibt. Es zeigt sich, dass das Wissen über die Bewegungskontrolle abhängig ist von Kenntnissen über das zentrale (ZNS) und periphere Nervensystem (PNS). Dabei ist die Suche nach den Wegen der Signale, die die Laufzeit und damit die Schnelligkeit einer Rückkoppelung und Korrektur eines Bewegungsablaufs bestimmen, ein wichtiger Forschungsgegenstand.
Bei der closed loop control liegt der Vorteil darin, dass bei allen Störungen, die im Laufe der Bewegungsausführung eintreten können, wie Verzögerungen von Informationsleitungen, von Rauschen (noise) in der Umgebung, in den Sinnesorganen oder den Leitungen, diese schnell kompensiert werden können. Auch ist durch die hohe Variabilität der Ausführungsmöglichkeiten eine Anpassung an aktuelle Anforderungen der Umgebung oder den Zustand des ausführenden Organismus gegeben.[26]
Das Hauptproblem der Bewegungskontrolle
Als Hauptproblem der Bewegungskontrolle wird deswegen – zum Teil noch immer – ihr Zeitbedarf angesehen. Die Abläufe der Kontrolle benötigen nämlich eine bestimmte Zeitspanne, um wirksam werden zu können. Die Rückkoppelungsschleifen müssen, wie man meinte, nämlich tatsächlich durchlaufen werden und dabei verstreicht eine gewisse Zeit, die, wie man glaubt(e), nicht ausreicht – vor allem bei schnellen Bewegungen nicht – um in den Ablauf eingreifen zu können. Die vor 1980 bekannten Informationswege im Organismus gaben die dazu notwendigen schnellen Informationsflüsse nicht her.
Dieses Zeitproblem spielt bis heute eine große Rolle bei der Diskussion über Möglichkeiten und Formen der Bewegungskontrolle. Es wird zum Beispiel auch heute noch als Begründung dafür verwendet, dass am Beginn des Lernens einer neuen Bewegung die Ausführung langsam erfolgen soll, weil dann die Kontrollschleifen erst aufgebaut werden müssen, damit sie später schnell und reibungslos erfolgen können.[27]
Um dieses Zeitproblem zu lösen, werden im Organismus die Strukturen im Zentralen und Peripheren Nervensystem gesucht, die eine Informationsübertragung gewährleisten, die schnell genug ist, eine online Kontrolle zu gewährleisten, weil das den Beobachtungen und Notwendigkeiten entspricht.
Struktur des Kontrollsystems
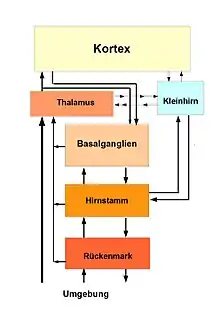
Das Zentralnervensystem lässt eine hierarchische Struktur erkennen, die unterschiedliche Ebenen besitzt, auf denen man die Bewegungskontrolle untersuchen und beschreiben kann. Das ist lange bekannt. Eine Beschreibung der Bewegungskontrolle innerhalb dieses hierarchischen Systems hat Erich von Holst in seinem Aufsatz über das Reafferenzprinzip[28] gegeben. (Nach seinen Beobachtungen und seiner Vorstellung werden die vom Kortex ausgegebenen motorischen Befehle im Nervensystem in einer Ablage (Efferenzkopie) im ZNS gespeichert. Die dann erfolgenden motorischen Befehle (Efferenzen) werden ausgeführt und die daraus resultierenden (Rück-)Meldungen (Reafferenzen) auf der untersten zentralen Ebene mit der Efferenzkopie verglichen. Sind diese Meldungen mit den Erwartungen aus der Efferenzkopie identisch, wird die Efferenzkopie gelöscht und der Bewegungsablauf ist vollendet. Bleiben Reste der Efferenzkopie bestehen, wird versucht, diese durch Maßnahmen auf der nächsthöheren zentralen Ebene zu lösen und zu löschen. Das wird fortgesetzt, bis die Efferenzkopie ganz gelöscht ist und kann bis zu einer bewussten Kontrolle (Veränderung) der Befehle führen). Der Begriff der Kontrolle war zu von Holsts Zeit noch nicht geläufig. Er hat seine Erkenntnisse über sorgfältige Beobachtungen des Verhaltens und des Nervensystems zahlreicher einfacher (Würmer, Fische, Fliegen) Lebewesen gewonnen. Die Lektüre des Artikels verhilft zu einem grundlegenden Verständnis der Bewegungskontrolle.
Überblick
Das Netzwerk der Bewegungskontrolle besteht aus gleichberechtigten – auch ineinander geschachtelten – Netzen (Elementen), die aber dennoch eine hierarchische Struktur haben.
Die höchste Ebene der neuralen Kontrolle dieser hierarchischen Struktur innerhalb des Großhirns, (Zentralnervensystem), ist der Kortex die Hirnrinde, die graue Substanz. Im Kortex liegt gleichsam das Management aller Funktionen. Es wird dort zum Beispiel darüber entschieden, welche Produkte (Bewegungen, Denken etc.) und wie sie hergestellt werden sollen. Weiterhin wird der gesamte (Entstehung und Durchführung) Produktionsprozess überwacht. Es ist die oberste Instanz für alle Aktionen (bewusste und unbewusste) des Organismus, von der Planung über Ausführung und Überwachung (Kontrolle). Schließlich ist es verantwortlich für die Beurteilung des Ergebnisses sowie deren Aufnahme in den Erfahrungsschatz.
Als die mittlere Ebene lassen sich die subkortikalen Zentren darstellen. Ihre wichtigsten für die Bewegungskontrolle sind die Basalganglien zur Auswahl von und als Initiator und Starter von Bewegungsabläufen[29], das Kleinhirn als Recheneinheit des Hirns sowie der Hirnstamm als eine Art Werkzeugkiste (toolbox) der neuronalen Netzwerke – beteiligt sind auch andere subkortikale Zentren zum Beispiel der Thalamus, der Hypothalamus und der Hippocampus. Sie sind aber für die Bewegungskontrolle nicht hauptsächlich zuständig.
Die unterste Ebene der Bewegungskontrolle ist das Rückenmark als eine Art Werkstatt, in der die resultierenden Informationen, die in elektrischer Form (Energie) vorliegen, zur mechanischen Umsetzung in den Muskeln zu den Bewegungen (Energie) weitergeleitet werden. Hier wird dafür Sorge getragen, dass das Produkt (der Bewegungsablauf) der Absicht und der Planung präzise entspricht. Dazu muss die Entwicklung des Produkts ständig beobachtet und kontrolliert und bei auch nur kleinen Abweichungen sofort korrigiert und angepasst werden. Dazu sind kleinere Netze vorhanden, die einfache Abfolgen schnell und variabler Form auslösen und durchführen können.
Das Großhirn
Aufbau und Funktion
Das Großhirn besteht aus der außen liegenden Großhirnrinde (Kortex, graue Substanz), und den darunter liegenden subkortikalen Strukturen, die die Nervenzellen enthalten, sowie aus den Leitungsbahnen (weiße Substanz), die all diese Strukturen sowohl untereinander als auch mit den nachgeordneten Hirnstrukturen und letztlich auch mit den Muskeln verbinden. Es ist das zentrale Verarbeitungs- und Integrationsorgan für Informationen aus dem Körper und aus der Umwelt.
Das Großhirn besteht aus zwei gleich aufgebauten Hemisphären (Hälften), die durch den Balken (Leitungsbahnen) miteinander verbunden sind. Die Oberfläche des Kortex ist stark gegliedert mit vielen Furchen (lat.: sulci, Singular: sulcus) und Spalten (Fissuren, fissura). Dadurch wird sie erheblich vergrößert. Größere Bereiche werden als Lappen (lobuli) bezeichnet, die entsprechend ihrer Lage (frontal, parietal, occipital) bezeichnet werden.
Der Kortex ist in verschiedene Regionen (Areale) unterteilt. Sie werden nach Arbeiten von Korbinian Brodman entsprechend den von ihm ihnen zugewiesenen Funktionen, nummeriert. Sie haben unterschiedliche Aufgaben zu erfüllen, arbeiten aber durch jeweils rekursive Informationsflüsse intensiv zusammen. Dadurch können sie auch aktuell ablaufende Handlungen (zum Beispiel Bewegungen) modulieren. Zu diesem Zweck bilden sie mit den untergeordneten Strukturen wie Thalamus, Basalganglien aber auch mit dem Kleinhirn und dem Hirnstamm Netzwerke, von denen ihnen ständig die Informationen über das aktuelle Geschehen zufließen. Insofern stellt der Kortex die übergeordnete Kontrollinstanz dar, die alle Abläufe kontrolliert. Die meisten dieser Informationsflüsse sind uns nicht bewusst.
Es lässt sich eine grobe Einteilung dieser Regionen (Areale) und ihrer Aufgaben in sensorische, motorische und assoziative Areale vornehmen. Die meisten Areale sind, wie man heute weiß, nicht homogen, sondern in kleinere Bereiche unterteilt, die auch jeweils unterschiedliche Aufgaben haben, indem sie zum Beispiel jeweils die Bewegungen unterschiedlicher Körperteile kontrollieren. Alle Bereiche des Großhirns sind aber in sich und mit allen anderen Bereichen als Netzwerke organisiert – auch über weit auseinander liegende Regionen.
In den sensorischen Arealen erfolgt die Aufnahme der Sinneswahrnehmungen. Ihre Verarbeitung geschieht auf unterschiedlich tiefen Stufen (primäre Areale, sekundäre Areale usw.). Dabei haben die verschiedenen Sinnesreize in den primären Arealen (sehen, hören, fühlen (Propriozeption, Hautafferenzen)) jeweils ihre eigenen Gebiete. In den höheren Arealen werden sie mehr und mehr ineinander integriert.
Die primären Areale (primary areas) erhalten ihre Informationen über den Thalamus. Sie sind topografisch aufgebaut. Das bedeutet, dass Informationen von benachbarten Wahrnehmungs-Feldern (der Signalaufnahme) auch an benachbarte Stellen gesendet und dort gespeichert werden (maps, Landkarten). Der primäre motorische Kortex erhält dadurch die Gestalt eines kleinen Menschen (Homunkulus). Dabei zeigt die relative Größe der Repräsentationszonen die Dichte der Innervation in den dazu gehörenden Wahrnehmungsfeldern an, zum Beispiel eine hohe Dichte für die Fingerspitzen. Die beiden Hemisphären erhalten im Allgemeinen die Sinneswahrnehmungen von der kontralateralen Körperseite, also rechter motorischer Kortex von der linken Körperseite.
In den motorischen Arealen werden die Bewegungen vorbereitet und ihre Ausführung veranlasst. Klassisch wird dieses motorische Areal (Brodmann-Areal 6) in 3 Unterbereiche geteilt. Auch hier kennt man heute eine viel feinere Gliederung. Die einzelnen Bereiche unterscheiden sich in ihren Neuronentypen, aber auch durch ihre mehr differenzierten Aufgaben.
Lange Zeit nahm man an, dass die eine Bewegung auslösenden Kommandos nur vom primären motorischen Kortex (primary motor cortex, M1), gelegen im vorderen (anterioren) parazentralen Lobus an der mittleren, internen Oberfläche, ausgehen. Diese Kommandos werden aber in den subkortikalen Strukturen ergänzt, aktualisiert, schließlich ins Rückenmark geleitet und dort in Bewegung umgesetzt. Aber auch die anderen motorischen Areale spielen eine wichtige Rolle bei der Auslösung von Bewegungen. Direkte Verbindungen vom primär motorischen Kortex zu Motoneuronen im Rückenmark mit einer schnellen Umsetzung in Bewegung bestehen nicht für alle Muskeln, aber für fein koordinierte Bewegungen der Hände und Finger. Letzteres ist die Voraussetzung für eine schnelle und feine Kontrolle von Hand- und Fingerbewegungen.
Die Signale vom primär motorischen Kortex enthalten hauptsächlich Kommandos für die kinetischen Parameter der Bewegungen, die bei Änderung der Umgebungsbedingungen entsprechend angepasst werden. Die Erforschung der komplexen Aufgaben und Fähigkeiten der Neurone des primär motorischen Kortex liefern noch immer neue Erkenntnisse.[30]
Bewegungen höherer Komplexität scheinen mehr durch das praemotorische Areal, das vor dem primär motorischen Kortex lokalisiert ist, kontrolliert zu werden, zum Beispiel für die direkte Kontrolle der proximalen Rumpfmuskeln. Hier werden auch Vorbereitungen von Bewegungen geleistet, wie die räumliche Führung für das Reichen und Zeigen. Hierzu werden entsprechende Informationen benötigt, die aus den Sinnesmeldungen im parietalen Kortex erarbeitet werden.
Der supplement motorische Komplex (SMC)[31], früher als supplement motorisches Areal (SMA) bezeichnet, liegt vor dem primär motorischen Areal, etwa an der Mittellinie des Kortex. Er hat viele unterstützende Funktionen, unter anderem die der internen Planung von Bewegungsabläufen, vor allem von Bewegungssequenzen, sowie die der Koordination der Bewegungen beider Körperseiten, besonders der bimanualen Koordination sowie für die Kontrolle und Anpassung der Körperhaltung an eine Bewegung.
Eine wichtige Rolle für die Bewegungsplanung und Ausführung spielen auch Teile des posterioren parietalen Kortex, der zu den Assoziationsarealen gehört, der aber auch hauptsächlich für Planung und Transformation von multisensorischen Informationen in motorische Kommandos zuständig ist. Ebenfalls an der Entwicklung von Bewegungen ist auch der primäre somatosensorische Kortex beteiligt, weil er nahe am primärmotorischen Kortex liegend, eng mit diesem kooperieren kann.
Viele weitere Areale – über große Teile des Kortex verteilt – werden als Assoziationsareale bezeichnet. Die Assoziationsareale nehmen den größten Teil der Oberfläche des Kortex ein. Sie sind verantwortlich für komplexe Verarbeitung der Informationen zwischen den primären sensorischen Gebieten und der Entstehung von Verhalten (motorischem und kognitivem). Sie sind damit für eine bedeutungsvolle Interpretation der Wahrnehmungen verantwortlich und ermöglichen es uns damit, erfolgreich mit der Welt zu interagieren. Sie befinden sich in den parietalen, temporalen und occipitalen Loben, die alle im hinteren (posterioren) Teil des Kortex lokalisiert sind. Sie sind, wie das ganze Hirn auch in sich als ein aktives Netzwerk organisiert. Das bedeutet, sie kommunizieren intern in allen ihren Funktionen untereinander und mit allen anderen Teilen des gesamten Hirns unter anderem mit den Basalganglien, dem Kleinhirn und der Pons im Mittelhirn.
Willkürliche Bewegungen werden nach Vorarbeit zahlreicher, wenn nicht aller Hirnstrukturen (wegen der ständig stattfindenden aktualisierten Analyse der äußeren (Umgebung) und inneren (Organismus) Situation) hauptsächlich initiiert von der motorischen Hirnrinde (M1). Eine wichtige Rolle für deren Entwicklung und Kontrolle spielt auch der praefrontale Kortex, der eine Art Filterfunktion hat und dadurch zielgerichtete Aktionen fördern und irrelevante abblocken kann. Die meisten Bewegungsabläufe, vor allem die des täglichen Lebens wie Atmen, Kauen, Schlucken, aber auch die Augenbewegungen, die Gleichgewichtskontrolle sowie die Fortbewegung können ohne den Kortex und Teile des Großhirns initiiert und auch kontrolliert werden.
Eine wichtige Rolle für die Bewegungskontrolle spielen die visuellen Informationen, die über sehr schnelle Verbindungen zum Kortex verfügen. Dabei hat die präzise Kontrolle von Arm-, Hand- und Fingermuskeln für feinkoordinierte Manipulation von Abläufen eine besondere Bedeutung. Das ist eine evolutionäre Weiterentwicklung der Motorik bei Primaten und Menschen. Die Regelkreise für diese Abläufe sind aus den phylogenetisch vorhandenen Netzwerken für die Aufgaben der oberen Extremitäten entstanden und sind für die Fortbewegung mit diesen Urformen auf das Engste verbunden.[32]
Die Bewegungskontrolle des Kortex unterscheidet sich von der der anderen Kontrollinstanzen auch dadurch, dass zum Einen vom Kortex auch die willkürlichen Bewegungen ausgewählt, zusammengestellt und initiiert werden, so dass sie über rekurrente Verbindungen auch während des Ablaufs geändert oder abgebrochen werden können (point of no return). Auch können im Kortex auf gleiche sensorischen Eingaben unterschiedliche Aktionen ausgelöst werden, je nach der auch im Kortex entwickelten Absicht oder Notwendigkeit.[33]
Die Erforschung der komplexen Aufgaben und Fähigkeiten der Neurone des primär motorischen Kortex liefern noch immer neue Erkenntnisse.[30]
Die Untersuchung der Zeiten für die Rückmeldung über ablaufende Bewegungen ist von großem wissenschaftlichem Interesse. Informationen über ausgeführte Bewegungen (Reafferenzen) erreichen zuerst (nach 25 ms) den parietalen Kortex, (Scheitellappen), etwas später (zwischen 40 ms und 60 ms) mehrere Teilen des dem somatosensorischen sowie den praemotorischen Kortex[34].
Die Zusammenarbeit aller an einer Bewegung oder Handlung beteiligten Hirnstrukturen kann durch Wiederholung verbessert werden. Das wird als Lernen bezeichnet.[35]
Die Basalganglien
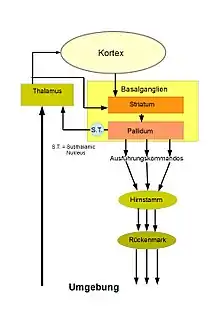
Aufbau
Die Basalganglien spielen eine wichtige Rolle bei der Auswahl der Bewegungsabläufe und deren Initiierung. Sie befinden sich im Inneren des Großhirns (subkortikal) und bestehen aus mehreren Teilen: Dem Nucleus caudatus (geschweifter Kern) und dem Nucleus lentiformis (linsenförmiger Kern) der wiederum aus dem Putamen und dem Globus pallidus (dem internen und dem externen) besteht. Nucleus caudatus und Putamen werden erst in der späteren Embryonalphase voneinander getrennt, bleiben aber lose miteinander verbunden und werden dann zusammen als Striatum bezeichnet. Im weiteren Sinne (funktionell) wird zu den Basalganglien auch der Nucleus subthalamicus und die Substantia nigra (schwarze Substanz), gerechnet. Letztere bildet ein Kerngebiet im Mittelhirn. Sie besteht aus einer pars compacta und einer pars reticulata.
Alle diese Teile sind netzwerkartig miteinander verbunden. Sie erhalten ihre wichtigsten Informationen von allen Teilen des Großhirns, zu denen sie auch wieder Signale zurücksenden, auch von anderen Teilen des Gehirns, zum Beispiel dem Thalamus und der Formatio reticularis, die den Hirnstamm durchzieht.
Funktion
Die Hauptfunktionen der Basalganglien sind die Auslösung und Auswahl der Bewegungen sowie nach neueren Theorien und Modellen (siehe unten) das bestärkende Lernen (reinforcement learning).[36] Dafür, wie die Auswahl der Bewegungen erfolgt, gibt es unterschiedliche Modelle.[37]
Die Auslösung der Bewegungen geschieht durch die Informationen vom Pallidum (globus pallidus internus), der Ausgabeformation der Basalganglien. Unter Ruhebedingungen werden diese Auslösekommandos durch starke inhibitorische (tonische Hemmung) Kontrollbefehle verhindert. Um die Bewegungsfolgen auszulösen, muss diese Hemmung (durch Disinhibition) aufgehoben werden. Diese erfolgt durch Neurone in der Eingangsformation der Basalganglien, dem Striatum.
Das Striatum erhält seine Informationen von verschiedenen Gebieten des Großhirns und direkt vom Thalamus, über den die Sinneswahrnehmungen zum Großhirn gelangen. Die Informationen aus diesen Quellen werden in den Netzwerken des Striatums verarbeitet und – falls es zur Auslösung der Bewegungen kommen soll – werden durch die exzitatorische Verbindungen zum Pallidum die Hemmungen dort aufgehoben. Der Globus pallidus internus sowie die Substantia nigra (pars reticulata) senden beide Rückmeldungen über ihre Aktionen zum Thalamus und zum Kortex (thalamocortical loop). Durch das Dopamin, das von der Pars compacta der Substantia nigra an alle Bereiche der Basalganglien wie auch den Nucleus subthalamicus ausgesendet wird, können die Prozesse die zu der Bewegungsauslösung führen, moduliert werden[38].
Es wird davon ausgegangen, dass die Schleifen zwischen den Basalganglien, dem Thalamus und dem Kortex an der Planung und Kontrolle von willkürlichen Bewegungen beteiligt sind, während die zwischen den Basalganglien und dem Hirnstamm für die korrekte und sichere Ausführung der unwillkürlichen Bewegungen, wie Schlucken oder Husten aber auch zum Beispiel der Durchführung der Fortbewegung (gehen) verantwortlich sind. Die Tätigkeit beider Schleifen wirken intensiv zusammen. So sind zum Beispiel die thalamokortikalen Schleifen für den geplanten Beginn oder das Ende des eines Gehvorgangs verantwortlich, oder wenn eine Bodenunebenheit eine (bewusste) Anpassung des Ablaufs notwendig macht. Dagegen sorgen die Verbindungen zum Hirnstamm für den sicheren Ablaufs der reinen Gehbewegung. Diese beiden Kontrollvorgänge müssen ineinander integriert werden.[39]
Die Basalganglien stellen also den entscheidenden Ort dar, von dem aus die Bewegungsabläufe aktiviert und dann im Hirnstamm koordiniert werden,[40]
Das Kleinhirn (Cerebellum)
Aufbau
Das Kleinhirn liegt an der Basis des Schädels unter dem Hinterhauptlappen, hinter dem Hirnstamm. Es besteht wie der Kortex aus einer außen liegenden Hülle aus Neuronen (graue Substanz) die zur Vergrößerung seiner Oberfläche sehr viel stärker durch Furchen gegliedert ist als der Kortex. Innerhalb dieser Hülle liegen die Leitungsbahnen, die weiße Substanz (Marklager). In sie eingelagert sind Gruppen (Kerne) von grauer Substanz (Neurone), die tiefen Kleinhirnkerne: Der Nucleus fastigius, der Nucleus globosus der Nucleus emboliformis sowie der größte, der Nucleus dentatus. Drei Kleinhirnteile (Pedunculi) verbinden das Kleinhirn mit den anderen Teilen des Nervensystems.
Sichtbar gliedert sich sie Oberfläche des Kleinhirns in horizontaler Richtung in die zwei Hemisphären mit den außen liegenden Teilen des Cerebrocerebellums und dem mehr innen liegenden Teilen des Spinocerebellums dessen mittlerer Teil auch als Vermis (Wurm) bezeichnet wird. Zusätzlich befindet sich vorne unten, an der dem Hirnstamm zugewandten Seite, vom Wurm ausgehend, ein Ausläufer, der Flocculus, der zusammen mit dem angrenzenden Wurmteil, dem Nodulus, zum Lobus flocculonodularis (auch Vestibulocerebellum) zusammengefasst wird. Diese einzelnen Bereiche haben unterschiedliche Funktionen und sind mit den ihren Funktionen entsprechenden Hirnteilen verbunden. Sie erhalten ihre Informationen von jeweils anderen Teilen des Hirns, verarbeiten sie – durchaus auch mit Informationen aus den anderen Kleinhirnbereichen – und geben die Ergebnisse – wieder über die tiefen Kerne direkt oder indirekt an unterschiedliche motorische (und andere, nicht motorische) Teilsysteme ab.
Die Zuflüsse zum Kleinhirn (Afferenzen) kommen aus fast allen Teilen des Nervensystems: Von vielen Teilen des Kortex, von Kernen im Hirnstamm (Umschaltstationen) und vom Rückenmark. Über die tiefen Kleinhirnkerne gelangen sie in das Kleinhirn und von dort über 2 Arten von afferenten Fasern zur Kleinhirnhülle.
Funktion
Die Hülle des Kleinhirns (Cerebellum) enthält fast die Hälfte aller Neurone, obwohl es nur 10 % des Hirnvolumens einnimmt. Jeder Abschnitt seiner Teile besteht aus den gleichen Einheiten von neuralen Elementen mit einer sehr stereotypen Geometrie. Diese Strukturen lassen sich in zahlreiche separate Module teilen, die auch als Mikroschaltkreise bezeichnet werden.
Diese Mikroschaltkreise mit ihren Neuronen und deren Verbindungen befinden sich in den drei Schichten der grauen Substanz. In der äußeren, der molekularen Schicht, befinden sich die Korb- und die (äußeren) Sternzellen. In der darunter liegenden Purkinjezell-Schicht die Purkinjezellen und in der untersten, der Körnerschicht, die Körnerzellen und die Golgi- oder (inneren) Sternzellen.
Es gibt zwei Arten von afferenten Fasern, die Moos- und die Kletterfasern, die zu den Neuronen führen. Die Moosfasern kommen aus verschiedenen Kernen außerhalb des Kleinhirns, die Kletterfasern nur vom unteren Olivenkern der kontralateralen Seite im Hirnstamm. Die einzigen Efferenzen führen von den Purkinjezellen zu den tiefen Kleinhirnkernen, der Ausgabestruktur des Kleinhirns.
Die Kletterfasern bilden mit den Dendriten der Purkinjezellen erregende Synapsen. Die Moosfasern bilden zunächst in der Körnerschicht erregende Synapsen mit vielen Körnerzellen, deren Axone sich zur Molekularschicht ziehen und dort parallel zur Kleinhirnrinde verlaufen. Dort bilden sie zahlreiche Kontakte mit Dendriten der Purkinjezellen. Kollateralen der Moos- und Kletterfasern ziehen auch zu den tiefen Kleinhirnkernen. Von den Kleinhirnkernen verlaufen dann zum Beispiel hemmende Verbindungen zum Olivenkomplex und erregende Verbindungen zu verschiedenen anderen Nervenkernen, zum Thalamus und zum Hirnstamm.
Man nahm lange Zeit an, dass die Hauptverarbeitungsleistung des Kleinhirns innerhalb dieser Mikroschaltkreise erfolgt, dass einkommende und ausgehende sowie erregende und hemmende Signale miteinander verglichen und miteinander Weise verrechnet werden. Auch nimmt man an, dass sie zur Koordination von Bewegungen und deren Teilen dienen. Inzwischen weiß man jedoch, dass diese Leistung der Verarbeitung hauptsächlich in den Kleinhirnkernen erfolgt. Die Verarbeitungsschritte in diesen tiefen Kleinhirnkernen sind noch wenig erforscht[41].
Die Funktionen der einzelnen Kleinhirnbereiche lassen sich wie folgt beschreiben:
Das Vestibulocerebellum (Lobus flocculonodularis) ist der entwicklungsgeschichtlich älteste Teil des Kleinhirns (daher auch Archizerebellum genannt). Es ist für das Gleichgewicht und die Balance des Körpers zuständig. Es erhält seine Informationen (Afferenzen) daher von den Bogengängen und dem Otholitenorgan im Innenohr (diese melden zum Beispiel die Position des Kopfes bezüglich der Schwerkraft). Zusätzlich erhält es über pontine und prätektale Kerne visuelle Informationen vom 1. und 2. visuellen Kortex. Das zeigt die Bedeutung der visuellen Informationen – vor allem auch unbewussten Informationen – für Balance und Gleichgewicht. Die Efferenzen des Vestibulocerebellums erfolgen zu den vestibulären Kernen. Durch sie werden die Augenmuskeln und die Koordination von Augenmuskeln und Kopf kontrolliert.
Das Spinocerebellum lässt sich in den mehr mittig liegenden Vermis und einen mehr außen liegenden Bereich teilen. Der Vermis kontrolliert die Stützmotorik. Er erhält daher seine Informationen (Afferenzen) von den somatosensorischen Rezeptoren der Rumpfmuskeln und den äußeren Augenmuskeln sowie von den vestibulären Kernen. Alle diese Informationen sind für Gleichgewicht und Balance von Bedeutung. Seine Efferenzen erfolgen über die tiefen Kerne (vor allem den Nucleus fastigius) zu weiteren Kernen im Hirnstamm und von dort über den retikulären und den vestibulospinalen Trakten zu Inter- und Motoneuronen im Rückenmark.
Der äußere Teil des Spinocerebellums erhält seine Informationen von den somatosensorischen Rezeptoren der Gliedmaßen (Berührung, Druck, Position der Gliedmaßen). Er erhält sie sowohl auf direktem als auch auf indirektem Weg. Der indirekte Weg verläuft über verschiedene Hirnstammkerne, meist denen der Formatio reticularis. Es lassen sich dabei zwei Übertragungslinien unterscheiden. Auf der dorsalen Linie werden Muskel- und Gelenkinformationen von erfolgten Bewegungen, die in die Planung mit übergehen, übertragen. Sie bilden die Efferenzkopie (feedforward). Die Informationen über eine aktuell ablaufende Bewegung erfolgen über die ventrale Linie. Die Informationen dieser beiden Linien werden miteinander verglichen (feedback), wodurch notwendige Modulationen des Bewegungsablaufs ausgelöst und durchgeführt werden können.
Das Cerebrocerebellum ist mit dem Assoziationskortex verbunden. Es erhält Informationen vom prämotorischen und supplementmotorischen Kortex über die pontinen Kerne und den mittleren Kleinhirnstiel. Nach der Verarbeitung gibt es die efferenten Informationen über den oberen Kleinhirnstiel und den Nucleus dentatus ab. Von dort erreicht ein Teil der Informationen über den Thalamus wieder die motorischen Kortizes, ein zweiter Teil bildet eine Schleife über den roten Kern (Nucleus ruber) und den Olivenkern wieder zum Kleinhirn. Man nimmt an, dass die Schleife über den roten Kern (nucleus ruber) und den prämotorischen Kortex die Einübung (Training) von Bewegungsabläufen und daher das Bewegungslernen unterstützt.
Die Aufgabe des Cerebrocerebellums ist die Planung von Bewegungen und die Regelung der vom Kortex ausgehenden motorischen Programme. Es ist aber auch an der Planung und Regulation reiner kognitiver Funktionen beteiligt.
Aufgaben und Eigenschaften des Kleinhirns lassen sich insgesamt wie folgt beschreiben:
Das Kleinhirn sorgt auf allen Ebenen der Planung und Durchführung dafür, dass willkürliche wie auch unwillkürliche Bewegungen sicher und erfolgreich durchgeführt werden können. Dazu vergleicht es aufgrund der Planung erwartete Signale mit ankommenden (afferenten wie reafferenten), die den aktuellen Zustand sowohl des Organismus als auch der Umgebung darstellen aber auch mit gespeicherten Daten von früheren Durchführungen von gleichen oder ähnlichen Abläufen. Auf diese Weise werden Bewegungserfahrungen eingearbeitet und neuen Situationen angepasst. Dadurch können die Muskelkontraktionen so koordiniert werden, dass die Bewegungen harmonisch ablaufen. Alle diese Aktionen des Kleinhirns erfolgen online.
Als besondere Hilfen können dabei folgende spezifische Mechanismen dienen:
- Die Purkinjezellen und die tiefen Kleinhirnkerne feuern nahezu gleichzeitig heftig bei willkürlichen Bewegungen. Das Ergebnis steht in enger Beziehung zu Richtung und Geschwindigkeit der Bewegung. Auch lässt sich die somatotopische Struktur des primärmotorischen Kortex innerhalb dieser Kerne finden. Das weist auf die rekurrenten Schleifen zwischen diesen beiden Strukturen hin.
- Die Muskelkontraktionen und damit das Timing (= zeitliche Abstimmung) werden durch Feedforwardstrukturen reguliert. Diese sorgen auch dafür, dass der Kraftverlauf nicht wie bei Feedbackstrukturen ruckartig verläuft, sondern sanft zum Endpunkt geführt wird.
- Man geht heute davon aus, dass das Kleinhirn über interne dynamische und kinematische Modelle verfügt (vor allem für Arme und Augen), die dafür sorgen, dass zum Beispiel bei Greifbewegungen der Endpunkt durch eine Folge von zeitlich abgestimmten Kommandos für die Muskelkontraktionen erfolgt. Gleichzeitig beschreiben präzise kinematische Modelle die Beziehung zwischen den Gelenkwinkeln und der Position der Finger die notwendige Position des gewünschten Endpunkts.[42][43]
Der Hirnstamm
Der Hirnstamm ist im Gegensatz zum Großhirn für die unbewussten Zustände verantwortlich, für die Vorbereitung von Aktionen und die Kommunikation mit anderen Individuen. Er sorgt für die Hintergrundzustände wie Wachheit und bewusste Aufmerksamkeit ebenso wie für den Schlaf. Er hat weitreichende steuernde Aufgaben in den Bereichen der Motorik, der vegetativen Zustände aber auch der kognitiven Funktionen. Zusammen mit dem Rückenmark lässt sich der Hirnstamm als eine Art Werkzeugkiste (toolbox) der neuronalen Netzwerke betrachten, denn er enthält das grundlegende Repertoire für die konkrete Vorbereitung, Ausführung und Kontrolle aller motorischen Aktionen: Man kann sich das vielleicht wie eine Art Fließband (im Englischen trifft das Wort assemblyline diese Funktion ziemlich gut) vorstellen: Wenn von den Basalganglien die Anforderung für einen bestimmten Bewegungsablauf ankommt, werden im Hirnstamm alle für diesen Ablauf benötigten Informationen von den Orten, an denen sie von ihren Quellen abgeliefert wurden und aufbereitet sind, auf das Fließband gesendet, zusammengestellt und für die Aufgabe angemessen – unter ständiger Kontrolle und Aktualisierung – ineinander integriert, so dass das jeweils aktuelle Produkt zur Ausführung an das Rückenmark übergeben werden kann. Er sorgt für die Abläufe, die zum Überleben notwendig sind wie Atmung, Schlucken, Gestik und Mimik, aber auch für andere notwendige wie Augenbewegungen, Körperhaltung und Fortbewegung ebenso wie für die willkürlichen Bewegungen. Er sorgt auch zum Beispiel nach Erhalt der Auswahl der Aktion durch das Striatum in den Basalganglien für die Auslösung der Zentralen Mustergeneratoren (CPGs), deren Ausführung vom Rückenmark aus erfolgt.
Möglich ist das dadurch, dass alle Informationsstränge – absteigend (efferent) und aufsteigend (afferent) – zwischen Großhirn und Rückenmark durch den Hirnstamm verlaufen, und andere wichtige Informationen dazukommen. Dort treffen die vom Großhirn ausgehenden und die aus dem Rückenmark rückgekoppelten Meldungen, die vom Kleinhirn koordinierten sowie die Informationen der Hirnnerven aus den Sinnesorganen des Kopfes und den vitalen Prozessen im Organismus aufeinander und werden ineinander integriert. Zu dieser Integrationsarbeit dienen die zahlreichen Nervenkerne (das sind Zusammenballungen zahlreicher Neurone, die zur Erfüllung bestimmter Aufgaben zusammenarbeiten und durch weitreichende und reziproke Verzweigungen und Verknüpfungen miteinander verbunden sind).
Aufbau und Funktionen
Der Hirnstamm verbindet die funktionell unterschiedlichen Strukturen des Großhirns und des Rückenmarks miteinander. Er liegt hinter und unterhalb (kaudal) des Großhirns und oberhalb (rostral) des Rückenmarks. Er besteht aus dem Mittelhirn (Mesencephalon), der Brücke (Pons) und dem verlängerten Mark (Medulla oblongata) und enthält hauptsächlich Nervenverbindungen und Nervenstränge (Trakte) zum Beispiel den cortikospinalen Trakt (Motorik), den medialen Lemniskustrakt (Sensorik) oder den spinothalamischen Trakt (für Schmerz, Berührung und Temperaturempfindung) sowie Nervenkerne.
Im Bereich des Mittelhirns befindet sich im obersten Teil, dem Mittelhirndach, die Vierhügelplatte mit dem Colliculus superior (oberer Hügel), der wie auch der laterale Kniehöcker (geniculate lateralis), mit dem er verbunden ist, an Vorgängen des Sehens beteiligt ist. Der ebenfalls dort befindliche Colliculus inferior (unterer Hügel) ist der wichtigsten Kern des Mittelhirns für die Gehörbahn. Er erhält Informationen vom auditorischen Kortex und einigen weiteren Hirnstammkernen. Im Tegmentum, dem unter dem Tectum liegenden Teil, befinden sich außer einer Reihe von Kernen der Hirnnerven weitere Nervenkerne und Nerventrakte die Formatio reticularis, ein Netz von Nervenzellen, das sich über das gesamte Tegmentum und durch das ganze Rückenmark hindurch zieht. Dort liegen die Kerne für die Regulierung der lebenswichtigen Funktionen wie Kreislauf, Atmung, sowie für das aufsteigende retikuläre Aktivierungssystem (ARAS), das Aufmerksamkeit und Wachheit, aber auch Schläfrigkeit und geistige Abwesenheit reguliert.
Im Mittelhirn befinden sich ebenfalls Kerne einiger Hirnnerven, der Nucleus ruber (roter Kern), ein Umschaltkern der motorischen Bahnen zwischen Großhirn und Kleinhirn und die Substantia nigra, der Ursprungskern des dopaminergen Systems, das zum Beispiel die Intensität von Aktionen steuert, positiv aufgrund von Belohnung, negativ aufgrund von Enttäuschung. Scheitelwärts (cranial) schließt sich an das Mittelhirn das Zwischenhirn (Diencephalon) an.
Die Pons (Brücke) ist ein Gebiet durch das hauptsächlich Verbindungsfasern zwischen vorderen und hinteren Gehirnbereichen verlaufen. Auch die Kleinhirnstiele, die die Nervenbahnen zwischen dem Kleinhirn und dem Großhirn enthalten, befinden sich hier.
Die Medulla oblongata ist die craniale (zum Kopf hin) Fortsetzung des Rückenmarks. Sie erstreckt sich vom sakralen Abschnitt des Rückenmarks bis zum Thalamus und besteht aus diffus angeordneten Neuronen.
Wie erwähnt, werden im Hirnstamm Bewegungen vorbereitet, die dann von Netzwerken der Zentralen Mustergeneratoren (CPGs) im Rückenmark ausgeführt werden.
Zum besseren Verständnis sollen hier zwei Beispiele kurz beschrieben werden.
1. Für die Organisation (Aktivierung – über die retikulospinale Verbindung) der Fortbewegung (Gehen) und deren Kontrolle existieren im Hirnstamm zwei Zentren, die mesencephalische motorische Region (MLR) und die diencephalisch motorische Region (DLR). (evolutionär bei allen Wirbeltieren – auch Menschen vorhanden – intensiv untersucht bei Tieren bis hin zu monkeys). Bei milder Reizung dieser Gebiete beginnen die Tiere zu gehen. Erhöht man den Reiz, beschleunigen sie ihre Fortbewegung bis hin zu einem Galopp – Vögel beginnen zu fliegen. Das Gehirn kontrolliert dabei das gesamte motorische Muster mit den zahlreichen dazu notwendigen unterschiedlichen Muskeln mit Hilfe eines einfachen, abgestuften Signals. – Die muskulären Details des Bewegungsablaufs werden dann von den Mustergeneratoren im Rückenmark und durch sensorische Kontrolle an die akute externe Situation angepasst[44].
2. Das Netzwerk, das die Sakkaden der Augen und die begleitenden langsamen Kopfbewegungen organisiert, befindet sich im oberen (superioren) Colliculus. Es gibt ein schnelles zentrales Kommando (das Richtung und Amplitude bestimmt), das die Augen (die Fovea centralis) schnell auf den Punkt des visuellen Interesses bringt, dann eine langsamere Gegenbewegung der Augen, wenn sie bei der langsamen Kopfbewegung das Objekt erreichen. Begleitet wird dieser Ablauf durch den vestibulo-ocularen Reflex. Dieser Vorgang wird durch die visuellen Signale aus dem vorderen Sehfeld der Augen kontrolliert.
Ein wichtiges Zusammenwirken beider Systeme findet statt, wenn die Fortbewegung über einen unsicheren Untergrund erfolgt, wenn also eine genaue Positionierung der Füße erforderlich ist. Bei diesen Bedingungen überlagert die visuomotorische Kontrolle das im Rückenmark erzeugte Muster der Fortbewegung. Das erfolgt über die Projektion von den visuellen Bereichen der Großhirnrinde zu den motorischen Bereichen über den dorsalen Strom für Bewegungswahrnehmung[45].
Das Rückenmark
Aufbau
Das Rückenmark durchzieht die gesamte Wirbelsäule. Es hat eine segmentale (Wirbelsäulensegmente) Struktur und besteht aus grauer und weißer Substanz. Die graue Substanz der Nervenzellen liegt im Innern und hat etwa die Form eines Schmetterlings. Bei den Nervenzellen handelt es sich um Interneurone, die zahlreiche Informationen zusammenführen, verarbeiten und weiterleiten und dabei Netzwerke organisieren, sowie um Motoneurone, von denen die Muskeln des entsprechenden Segments innerviert werden. Die Motoneurone sind topografisch präzise angeordnet entsprechend der Lage der von ihnen innervierten Muskeln. Sie sind jeweils in sogenannten pools (großen Gruppen, die denselben Muskel innervieren) zusammengefasst.
Die weiße Substanz enthält die Nervenstränge, die sowohl die Informationen von den übergeordneten Gehirnstrukturen zu den Nervenzellen der Segmente leiten (absteigende, efferente Bahnen) als auch umgekehrt die Informationen aus den einzelnen Segmenten zu den übergeordneten Strukturen (afferente Bahnen). Die Nervenstränge haben eine festgelegte Anordnung. Die afferenten Bahnen liegen mehr am äußeren Umfang des Rückenmarks, die efferenten mehr zur Mitte hin. Weitere afferente Bahnen befinden sich in der hinteren (dorsalen) Lücke zwischen den Schmetterlingsflügeln. Efferente Bahnen, zum Beispiel Teile der Pyramidenbahn befinden sich im vorderen (ventralen) Teil. Zwischen den Wirbeln treten die für das jeweilige Segment notwendigen neuralen Verbindungen aus beziehungsweise ein.
Funktion
Im Rückenmark werden die Informationen aus den höheren Zentren (efferent) sowie aus den Muskeln, den Sehnen, den Gelenken und der Haut (afferent) integriert und für das Umsetzen in das die Bewegungen auslösende mechanische System der Muskeln umgesetzt.
Am längsten bekannt und auch am sorgfältigsten untersucht und beschrieben sind die vom Rückenmark ausgehenden spinalen Reflexe. Sie bilden eine sehr schnelle Antwort (Kontraktion) z. B. auf die Dehnung eines Muskels – zum Beispiel der Kniesehnenreflex. Diese Reflexe wurden bereits zu Beginn des 20. Jahrhunderts von dem Physiologen Charles Sherington[46] beschrieben. Man nahm lange Zeit an, dass sie starre Verhaltensmuster darstellen und nicht modifizierbar sind. Insofern wurden sie nur wenig in die Überlegungen zur Bewegungskontrolle miteinbezogen. Man nahm lange Zeit an, dass diese „Reflexe“ starre Verhaltensmuster darstellen und nicht modifizierbar sind. Insofern wurden sie nicht in die Überlegungen zur Bewegungskontrolle miteinbezogen.
Später entdeckte man sie als Kontrollschleifen, die auf der Rückenmarksebene arbeiten. Sie sind schnell genug (ca. 20–45 ms),[47] um dort während eines Bewegungsablaufs wirksam werden zu können. Ihre Schleifen verlaufen von den Motoneuronen im Rückenmark zu den diesen zugeordneten Muskeln. In den betreffenden Muskeln beziehungsweise Sehnen befinden sich Messfühler (die Muskelspindeln bzw. Golgi-Sehnenrezeptoren), die den Erfolg der Innervation an die Motoneurone zurückmelden. Die Muskelspindeln, die innerhalb des Muskels liegen, messen die Länge und die Geschwindigkeit der Längenänderung der Muskelfasern. Die Golgi-Sehnenrezeptoren, die in den Sehnen liegen, messen die Spannung der Muskeln. Die Messergebnisse dieser Sensoren werden zu verschiedenen Neuronen in dem Rückenmarksegment zurück gemeldet, aus dem die Innervation der Muskelfasern kam – die Muskelspindeln melden direkt (monosynaptisch) erregende Signale an das auslösende Motoneuron zurück, die Sehnenrezeptoren ihre hemmenden Signale an Interneurone. Diese Verschaltungen werden den aktuellen Anforderungen entsprechend modifiziert. Insofern lässt sich von Regel- oder Kontrollkreisen sprechen. Außer diesen rückgekoppelten Meldungen aus den Muskeln gelangen auch die Signale anderer Rezeptoren (zum Beispiel Hautafferenzen und Gelenkinformationen) zu den Motoneuronen. Dadurch ist eine schnelle Reaktion auf eine komplexe Umgebungssituation möglich und sie bieten eine schnelle erste Antwort auf die Störung eines Bewegungsablaufs.
Heute weiß man, dass diese Kontrollschleifen Teile größer Netzwerke sind, die ganze Bewegungsabläufe organisieren und kontrollieren. Diese Netzwerke, die auch als Zentrale Mustergeneratoren (central pattern generator, CPG) bezeichnet werden, haben sich im Laufe der Evolution entwickelt.
Die Mustergeneratoren sind Segment übergreifend angeordnet, so dass durch sie auch Bewegungen Segment übergreifender Muskelgruppen (Beine zum Gehen, Arme für Manipulationen) organisiert und kontrolliert werden. Diese Netzwerke gehören für elementare Bewegungsabläufe (zum Beispiel Gehen, Greifen, aber auch Schlucken, Husten) zur Ausstattung der Lebewesen von Geburt an. Sie haben sich von den einfachsten Bewegungsformen evolutionär für die höheren Lebensformen weiterentwickelt und werden im Laufe des Lebens durch Anpassungs- und Lernprozesse modifiziert und erweitert.[48]
Diese Muster werden von lokalen Interneuronen im Rückenmark zu geordneten Netzwerken zusammengestellt. Sie koordinieren die Befehle an die Muskeln und kontrollieren deren Aktionen. Bei der Ausführung sorgen sensible Rückkoppelungsmechanismen (Afferenzen) für eine Verfeinerung der Ausführung und eine Anpassung an die aktuelle Umgebungssituation. Diese Netzwerke sind dadurch in höchstem Grad flexibel[49]
Die Auslösung, Koordination und Kontrolle dieser Netzwerke erfolgt im Hirnstamm, denn hier werden die Signale (Informationen) vom Großhirn, den Hirnnerven (Sinnesorgane, vegetative und emotionale Einflüsse) sowie die Informationen über den Zustand der Skelettmuskulatur und den aktuellen Zustand des Körpers zusammengeführt und dann an die ausführenden Systeme (Muskeln) weitergeleitet.
Die Komplexität, die auf dem Zusammenwirken dieses Kontrollkreise mit den höheren Zentren erfolgt, wird zurzeit intensiv erforscht.
Untersuchungsverfahren und Ergebnisse
Das klassische Paradigma bei der Forschung im Bereich der Motor Control ist es, eine vorgeschriebene Körperhaltung oder Bewegung zu stören (perturbation) und zu beobachten und zu messen, ob und wie die beabsichtigte Bewegung oder Haltung wieder hergestellt (kompensiert) wird. Zu diesem Zweck wird der Versuchsperson die Aufgabe gestellt, entweder eine bestimmte Position beizubehalten oder eine vorgeschriebene Bewegung auszuführen. Die Ausführung dieser Aufgabe wird dann gestört, meist durch eine mechanische Einwirkung auf die Körperhaltung oder die ausführenden Gliedmaßen. Es werden dann sowohl die Art als auch der Beginn und die Dauer der Reaktion auf diese Störung gemessen. Als Messwerte dienen die kinematischen Werte für Ort, Winkel, Geschwindigkeit und Beschleunigung, die kinetischen von einwirkenden Kräften sowie die Aktionen der Muskeln, die bei den Bewegungsabläufen auftreten.
Aktuell ist in erster Linie das Studium der Kontrolle von Arm-/Handpositionen und deren Bewegungen. Diese sind von Bedeutung für die Steuerung von künstlichen Gliedmaßen zum Beispiel nach dem Verlust des natürlichen Körperteils (zum Beispiel einer Hand) aber auch zur Konstruktion von lernfähigen Robotern, die in der Industrie benötigt werden. Aus diesem Grund ist es wichtig, die Untersuchungsergebnisse in mathematische Modelle umzusetzen, das bedeutet, die dazugehörenden Gleichungen aufzustellen.
Da die Komplexität der mathematischen Gleichungen für die Modelle mit der Anzahl der beteiligten bewegten Glieder zunimmt, beschränkt man sich zurzeit hauptsächlich auf die Untersuchung von Armbewegungen, weil sich diese mit wenigen kontrollierbaren Körperteilen (Hand, Unterarm, Oberarm) und wenigen Gelenken (Handgelenk, Ellenbogen- und Schultergelenk – meist werden nur jeweils nur eins oder maximal zwei Gelenke betrachtet) ausführen lassen und nur eine geringe Anzahl von Muskeln beteiligt sind.
Bei einem Teil der Untersuchungen wird so vorgegangen, dass die Versuchsperson in einer definierten Umgebung aufgefordert wird, den Arm oder die Hand in einer bestimmten Position zu halten, auch wenn durch eine plötzlich einwirkende (in Stärke und Richtung definierte) Kraft versucht wird, den Arm (Hand) aus dieser Position zu bewegen. Gemessen werden dann: Die Reaktionszeit der Versuchsperson, die Richtung, Geschwindigkeit und Beschleunigung der ausgelösten Bewegung sowie die Intensität und der Zeitverlauf der Aktivität der beteiligten Muskeln (elektromyografische Aufzeichnungen – Elektromyografie). Schließlich wird die Dauer der gesamten Aktion – bis der Arm (Hand) wieder zur Ruhe kommt, bis also die Störung kompensiert ist – gemessen. Aus der Dauer der Ausgleichsbewegung wird versucht, auf die möglichen Wege der Signalverarbeitung im Organismus von der Wahrnehmung der Störung bis zu deren Kompensation zu schließen.
Bei einem anderen Teil der Untersuchungen ist es die Aufgabe des Probanden, aus einer definierten Position heraus eine vorgeschriebene zielgerichtete Bewegung auszuführen, die dann durch einwirkende Kräfte (oder durch eine Manipulation der Wahrnehmung der Umgebung) gestört wird. Es werden dabei analog die gleichen Messwerte erhoben, wie im vorigen Absatz beschrieben.
Auch bei der Auswertung der Ergebnisse dieser Untersuchungen spielen Werte wie die Zeit, die vergeht, bis der Organismus überhaupt eine regulierende Reaktion zeigt (Reaktionszeit), sowie der Beendigung der Ausgleichsbewegung eine Rolle. Die Ausgleichsbewegung besteht darin, den Arm aus der ausgelenkten Position wieder in die beabsichtigte Bewegung zu überführen. Dabei werden Gelenkwinkel und Muskelaktionen gemessen und aufgezeichnet. Es werden dann die Unterschiede der Reaktionen auf unterschiedliche, aber stets kontrollierte Störungen ausgewertet. Von den Zeitverläufen der Muskelaktionen und deren Zuordnung zu den unterschiedlichen Störungen hofft man auf die Kontrollwege schließen zu können.
Die Ergebnisse bestätigen bereits länger bestehende Annahmen über einen möglichen Kontrollaufbau. Für die Armbewegungen zeigen sich 3 bis 4 unterschiedliche Zeitabschnitte (Epochen) der Reaktion. Es sind die: Short-latency response (20–45 ms), die long-latency response (50–105 ms) und die volitional response (120–180 ms). Dazu wird gelegentlich ein Zeitraum für eine Vorbereitungszeit (baseline epoch (−100–0 ms)) gerechnet.[50] Das bedeutet: Bereits nach weniger als 40 ms der Störung lässt sich im EMG (Elektromyografie) eine erhöhte Aktivität erkennen. Dieser Anteil wird als 1. Epoche bezeichnet.[51] Sie ist eine Folge des kurzen Kontrollkreises über das Rückenmark. Danach erhöht sich die elektrische Aktivität. Es beginnt die „long latency response“, die in die willkürliche Phase (100–180 ms) der Bewegungskontrolle übergeht. Hieraus ergibt sich die früher angenommene Korrekturzeit (bei Sichtkontrolle) bei einer open loop control. von ca. 200 ms.
Die Angaben über die Laufzeiten – auch für die 1. Epoche – sind so unterschiedlich, weil es jeweils unterschiedliche Wege der Signale gibt. Bei der 1. Epoche der Reaktion ist die kürzeste Verbindung die monosynaptische (nur die Synapsen am Motoneuron) Verbindung von der Muskelspindel zum Motoneuron des homonymen Muskels (er innerviert die Muskelfasern, die kontrahiert wurden). Die Signale von den Muskelspindeln können aber auch über Kollateralen zu Interneuronen und dadurch zu anderen Motoneuronen – das bedeutet längere Schaltwege – gelangen. Kollateralen werden aber auch an aufsteigende Nervenbahnen gesendet, die die übergeordneten Kontrollkreise und möglicherweise weitere Aktivierungen der Muskeln auslösen.
Zu beobachten ist auch, dass in der ersten sehr kurzen Reaktionszeit keine großen Variationen der Muskeltätigkeit zu beobachten sind – bedingt durch die Beschränkung der Rückmeldung auf den Motoneuronenpool nur weniger Muskeln im direkten Wirkungsbereich des Rückenmarks. Diese Neurone können sehr schnell reagieren. Die Variabilität der Muskeltätigkeit nimmt dann mit zunehmender Dauer der Reaktion zu, weil immer mehr Systemteile einbezogen werden können.
In dem Zeitbereich der short latency und dem 1. Teil der long latency response können sehr schnelle Antworten auf die Störung eines Bewegungsablaufs erfolgen, die häufig als automatisierte Antworten bezeichnet werden. Aber auch sie sind spezifisch für jeden Bewegungsablauf und werden zusammen mit diesem gelernt.
Anders als über die short-latency response über deren Ursachen und Quellen es in der Bewegungswissenschaft kaum unterschiedliche Ansichten gibt, existieren über die Quellen und die Zusammensetzung der long-latency response auseinandergehende Ansichten, die seit über 60 Jahren diskutiert werden.
Ein Anteil wird noch immer von einigen Forschern als eine Art automatische – also unabhängig von der spezifischen Aufgabe oder Bewegung – Antwort gesehen, die lediglich von höheren Zentren der Bewegungskontrolle, also Anteilen im Hirnstamm ausgehen. Ein anderer Teil wird der spezifischen Aufgabenstellung zugeordnet. Wie groß diese jeweiligen Anteile sind und ob sie zeitlich voneinander getrennt werden können oder sich überlappen, ist Gegenstand aktueller Untersuchungen.[50]
Literatur
- J. A. Adams: A closed loop theory of motor learning. In: Journal of Motor Behavior. 3 (1971), S. 111–150.
- Tyler Cluff, Fréderic Crevecoeur, Stephen H Scott: A perspective on multisensory integration an rapid perturbation. In: Vision Research. 2014, doi:10.1016/j.visres.2014.06.011
- F. Crevecoeur, I. Kurtzer, S. H. Scott: Fast corrective responses are evoked by perturbations approaching the natural variability of posture and movement tasks. In: Journal of Neurophysiology. 107 (2012), S. 2821–2832.
- F. Crevecoeur, I. Kurtzer, T. Bourke, S. H. Scott: Feedback responses rapidly scale with the urgency to correct for external perturbations. In: Journal of Neurophysiology. 110 (2013) S. 1323–1332.
- James Gibson: Wahrnehmung und Umwelt. Urban und Schwarzenberg, München 1982.
- Sten Grillner: Biological Pattern generation: The Cellular and Computational Logic of Networks in Motion in: Neuron 52 (2006). S. 751–766.
- Sten Grillner, Thomas Jessel; Measured Motion: Searching for Simplicity in Spinal Locomotor Networks. In: Current Opinion in Neurobiology. 19 (2009) S. 572–586. doi. 10.1016/jconb.2009.10.011.
- Sten Grillner, Peter Wallén: Innate versus learned movements – a false dichotomy? in: Progress in Brain Research. 143 (2004). doi:10.1016/S0079-6123j43001-X
- Sten Grillner, Jeannette Hellgren, Ariane Ménard, Kazuya Saitoh, Martin A. Wikström: Mechanisms for selection of basic motor programs – Roles for the striatum and Pallidum. In: Trends in Neuroscience 28 (2005). S. 363–370.
- Joachim Haase, Hans Dieter Henatsch, Richard Jung, Piergiorgio Brescia, Uwe Toden; Sensomotorik in O.H.Gauer/K.Kramer/R. Jung (Hrsg.); Physiologie des Menschen. Urban und Schwarzenberg München 1976. Bd. 14.
- Erich von Holst: Untersuchungen über die Funktionen des Zentralnervensystems beim Regenwurm. Dissertation 1932. Abgedruckt in: International Journal of Zoological Sciences. 51 (1932), S. 547–588.
- Erich von Holst: Zur Verhaltensphysiologie bei Tieren und Menschen. gesammelte Abhandlungen Band I und II. Piper & Co Verlag, München 1969.
- Erich von Holst, Horst Mittelstaedt: Das Reafferenzprinzip. In: Naturwissenschaften. 37 (1950) S. 464–476.
- Jim C. Houk, S. P. Wise: Distributed modular architectures linking basal ganglia, cerebellum, and cerebral cortex: Their role in planning and controlling action. In: Cerebral Cortex. Band 5, 1995, S. 95–110 (englisch).
- Eric R. Kandel, James H. Schwartz, Thomas M. Jessel, Steven A. Siegelbaum, A.J. Hudson; Principles of Neural Science McGraw Hill, New York. 5. Auflage 2013, ISBN 978-0-07-139011-8
- Karl Küpfmüller: Grundlagen der Informationstheorie und Kybernetik. In: O. H. Gauer, K. Kramer, R. Jung: Physiologie des Menschen. Band 10, Urban & Schwarzenberg, München 1974, S. 209–248.
- Mark Latash: Progress in Motor Control. Volume I, Movement Kinetics Publishers. Champaign, Illinois 1996.
- Peter H. Lindsay, Donald A. Norman: Human Information Processing. 2. Auflage. Academic Press, New York 1977.
- Ronald G. Marteniuk: Information Processing in Motor Skills. Holt Rinehart & Winston, New York 1976.
- J. Andrew Pruszinski, Isaac Kurtzer, Stephen H. Scott: The long latency reflex is composed of at least two functionally independent processes. In: Journal of Neurophysiology. 106 (2011), S. 451.
- Edward S. Reed: An Outline of the Theory of Action Systems. In: Journal of Motor Behavior. 14, 1982, S. 98–134.
- Richard A. Schmidt: Motor control and Learning. Human Kinetics Publishers, Champaign, Illinois 1982, ISBN 0-931250-21-8.
- Stephen Scott: Optimal feedback control and the neural basis of volitional motor control. in: Nature Reviews Neuroscience 5 (2004) S. 532–546. doi:10.1038/nrn1427
- Charles C. Sherington: Flexion-reflex of the limb, crossed extension-reflex, and reflex stepping and standing. In: Journal of Physiology. 40 (1910), S. 28–121.
- Lior Shmuelof, John W. Krakauer, Pietro Mazzoni: How is a motor skill learned? Change and invariance at the levels of task success and trajectory control. In: Journal of Neurophysiology. 108 (2012) S. 578–594.
- Paul S.G. Stein, Sten Grillner, Allen I. Selverston, Douglas G. Stuart; Neurons, Networks, and Motor Behavior. MIT Press, Cambridge Massachusetts. 1997, ISBN 0-262-19390-6
- K. Takakusaki, K.Saitoh, H. Harada, M.Kashiwayanagi: Role of basal ganglia – brainstem pathways in the control of motor behavior. Neurosci. Res. 50(2) (2004). S. 137–51. doi:10.1016/j.neures.2004.06.015
- Emanuel Todorov: Optimality principles in sensorimotor control. In: Nature Neuroscience. 7 (2004), S. 907–715.
- Norbert Wiener: Mensch und Menschmaschine. Kybernetik und Gesellschaft. Alfred Metzner Verlag, Frankfurt am Main 1952.
- Howard N. Zelaznik (Hrsg.): Advances in Motor Learning and Control. Human Kinetics Publishers, Champaign, Ill. 1996, ISBN 0-87322-947-9.
Einzelnachweise
- Mark Latash: Progress in Motor Control I. Movement. Kinetics Publishers, Champaign, Illinois 1996. Einleitung
- Zum Beispiel: Paul S.G. Stein, Sten Grillner, Allen I. Selverston, Douglas G. Stuart; Neurons, Networks, and Motor Behavior. MIT Press, Cambridge Massachusetts. 1997, ISBN 0-262-19390-6.
- Joachim Haase, Hans Dieter Henatsch, Richard Jung, Piergiorgio Brescia, Uwe Toden; Sensomotorik in O.H.Gauer/K.Kramer/R. Jung (Hrsg.); Physiologie des Menschen. Urban und Schwarzenberg, München 1976. Bd. 14. S. 292 f.
- Joachim Haase, Hans Dieter Henatsch, Richard Jung, Piergeorgio Brescia, Uwe Toden; Sensomotorik in O.H.Gauer/K.Kramer/R. Jung (Hrsg.); Physiologie des Menschen. Urban und Schwarzenberg München 1976. Bd. 14. S. 318 f.
- siehe zum Beispiel: Sten Grillner. Biological Pattern generation: The Cellular and Computational Logic of Networks in Motion in: Neuron 52 (2006) S. 751–766.
- zum Beispiel: Tyler Cluff, Fréderic Crevecoeur, Stephen H Scott: A perspective on multisensory integration an rapid perturbation. In: Vision Research. (2014), doi:10.1016/j.visres.2014.06.011
- Luigi Galvani: De viribus electricitatis in motu musculari. Verlag Deutsch, Frankfurt am Main 1996, ISBN 3-8171-3052-X.
- Richard Jung: Einführung in die Bewegungsphysiologie. In: J. Haase, H.-D. Henatsch, R. Jung, P. Strata, U. Thoden: Sensomotorik. In: O. H. Gauer, K. Kramer, R. Jung: Physiologie des Menschen. Band 14. Urban und Schwarzenberg, München 197. S. 2.
- siehe zum Beispiel: Edwin A. Fleishman, Walter Hempel: Factorial Analysis of Complex Psychomotor Performance and Related Skills. In: The Journal of Applied Psychology. 40 (1956) S. 96–104.
- Karl Küpfmüller: Grundlagen der Informationstheorie und Kybernetik. In: O. H. Gauer, K. Kramer, R. Jung: Physiologie des Menschen. Band 10, Urban & Schwarzenberg, München 1974, S. 209–248.
- Erich von Holst: Untersuchungen über die Funktionen des Zentralnervensystems beim Regenwurm. Dissertation. 1932. Abgedruckt in: International Journal of Zoological Sciences. 51 (1932) S. 547–588.
- Erich von Holst, Horst Mittelstaedt: Das Reafferenzprinzip. In: Naturwissenschaften. 37 (1950) S. 464–476.
- Richard A. Schmidt: Motor control and Learning. Human Kinetics Publishers, Champaign, Illinois 1982.
- Lindsay/Norman
- Ron Marteniuk
- zum Beispiel: Wahrnehmung und Umwelt. Urban und Schwarzenberg, München 1982.
- Edward S. Reed: An Outline of the Theory of Action Systems. In: Journal of Motor Behavior. 14, 1982, S. 98–134.
- O. G. Meijer, K. Roth (Hrsg.): Complex Motor Behavior, The Motor-Action Controversy. North Holland Publishers, Amsterdam.
- G. N. Gantchev, B. Dimitrov, P. Gatev (Hrsg.): Motor Control, proceedings of the fifth International Symposium on Motor Control 1985 in Varna, Bulgarien. Plenum Press, New York 1987.
- Nicolas Bernstein: The Coordination and Regulation of Movements. Pergamon Press, Oxford 1967.
- H. T. A. Whiting (Hrsg.): Human Motor Actions, Bernstein reassessed. 2. Auflage. North Holland. 1986.
- Rob Bongaard, Onno G Meijer: Bernstein’s Theory of Movement Behavior: Historical Development and Contemporary Relevance. In: Journal of Motor Behavior. 2000, S. 59.
- siehe: Richard A. Schmidt: Motor control and Learning. Human Kinetics Publishers, Champaign, Illinois 1982, S. 190f.
- Emanuel Todorov: Optimality principles in sensorimotor control. In: Nature Neuroscience. 7 (2004), S. 907/908.
- J. A. Adams: A closed loop theory of motor learning. In: Journal of Motor Behavior. 3 (1971), S. 111–150.
- Emanuel Todorov: Optimality principles in sensorimotor control. In: Nature Neuroscience. 7 (2004), S. 908.
- zum Beispiel: Lior Shmuelof, John W. Krakauer, Pietro Mazzoni: How is a motor skill learned? Change and invariance at the levels of task success and trajectory control. In: Journal of Neurophysiology. 108 (2012), S. 578–594.
- Erich von Holst, Horst Mittelstaedt: Das Reafferenzprinzip In: Naturwissenschaften. 37 (1950) S. 464–476
- Sten Grillner, Jeannette Hellgren, Ariane Ménard, Kazuya Saitoh, Martin A. Wikström; Mechanisms for selection of basic motor programs – Roles for the striatum and Pallidum. In Trends in Neuroscience 28 2005. S. 363–370.
- Eric R. Kandel, James H. Schwartz, Thomas M. Jessel, Steven A. Siegelbaum, A.J. Hudson; Principles of Neural Science. McGraw Hill, New York. 5. Auflage 2013, S. 843
- Eric R. Kandel, James H. Schwartz, Thomas M. Jessel, Steven A. Siegelbaum, A.J. Hudson; Principles of Neural Science. McGraw Hill, New York. 5. Auflage 2013, S. 841
- Eric R. Kandel, James H. Schwartz, Thomas M. Jessel, Steven A. Siegelbaum, A.J. Hudson; Principles of Neural Science. McGraw Hill, New York. 5. Auflage 2013. S. 845, ISBN 978-0-07-139011-8
- Sten Grillner, Peter Wallén: Innate versus learned movements – a false dichotomy? in: Progress in Brain Research. 143 (2004). doi:10.1016/S0079-6123j43001-X
- unter anderem: Mohsen Omrani, Chantelle D. Murnaghan, J. Andrew Pruszynski, Stephen H. Scott; Distributed task-specific processing of somatosensory feedback for voluntary motor control. In: eLive 2016; 5: e13141. doi:10.7554/eLive.13141
- Eric R. Kandel, James H. Schwartz, Thomas M. Jessel, Steven A. Siegelbaum, A.J. Hudson; Principles of Neural Science McGraw Hill, New York. 5. Auflage 2013, ISBN 978-0-07-139011-8
- J. C. Houk, S. P. Wise: Distributed modular architectures linking basal ganglia, cerebellum, and cerebral cortex: Their role in planning and controlling action. In: Cerebral Cortex. Band 5, 1995, S. 95–110 (englisch).
- Jim Houk, Models of the basal ganglia. in. Scholarpedia (2007) 2(10):1633. doi:10.4249/scholarpedia.1633
- Jim Houk, Models of the basal ganglia. in. Scholarpedia (2007) 2(10):1633. doi:10.4249/scholarpedia.1633. S. 2
- K. Takakusaki, K. Saitoh u. a.: Role of basal ganglia–brainstem pathways in the control of motor behaviors. In: Neuroscience Research. 50, 2004, S. 137, doi:10.1016/j.neures.2004.06.015.
- Sten Grillner; Biological pattern generation: The cellular and computational logic of networks in motion. In: Neuron 52 (2006). S. 751–766.
- M Uusisaari, E. De Schutter: The mysterious microcircuitry of the cerebellar nuclei. In: Journal of Physiology 15 (2011). S. 3441–3457. doi:10.1113/jphysiol.2010.201582
- Eric R. Kandel, James H. Schwartz, Thomas M. Jessel, Steven A. Siegelbaum, A.J. Hudson; Principles of Neural Science. McGraw Hill, New York. 5. Auflage 2013, S. 973
- Eric R. Kandel, James H. Schwartz, Thomas M. Jessel, Steven A. Siegelbaum, A.J. Hudson; Principles of Neural Science McGraw Hill, New York. 5. Auflage 2013, ISBN 978-0-07-139011-8, S. 960 ff.
- Sten Grillner, Peter Wallén; Innate versus learned movements – a false dichotomy? In: Progress in Brain Research 143 (2004). doi:10.1016/S0079-6123(03)43001-X. S. 4
- Sten Grillner, Peter Wallén; Innate versus learned movements – a false dichotomy? In: Progress in Brain Research 143 (2004). doi:10.1016/S0079-6123(03)43001-X. S. 7
- C. S. Sherrington: Flexion-reflex of the limb, crossed extension-reflex, and reflex stepping and standing. In: Journal of Physiology. 40 (1910), S. 28–121
- zum Beispiel: F. Crevecoeur, I. Kurtzer, S.H. Scott: Fast corrective responses are evoked by perturbations approaching the natural variability of posture and movement tasks. In: Journal of Neurophysiology. 107 (2012), S. 2823.
- Sten Grillner, Peter Wallén: Innate versus learned movements – a false dichotomy? in: Progress in Brain Research. 143 (2004). doi:10.1016/S0079-6123j43001-X.
- Sten Grillner, Thomas Jessel; Measured Motion: Searching for Simplicity in Spinal Locomotor Networks. In:Current Opinion in Neurobiology. 19 (2009) S. 572–586. doi. 10.1016/jconb.2009.10.011.
- J. Andrew Pruszinski, Isaac Kurtzer, Stephen H. Scott: The long latency reflex is composed of at least two functionally independent processes. In: Journal of Neurophysiology. 106 (2011), S. 451.
- F. Crevecoeur, I. Kurtzer, T.Bourke, S.H.Scott: Feedback responses rapidly scale with the urgency to correct for external perturbations. In: Journal of Neurophysiology. 110 (2013), S. 1323–1332.