Visueller Cortex
Der visuelle Cortex (auch Sehrinde) ist der Teil der Großhirnrinde, der zum visuellen System zählt, welches wiederum die visuelle Wahrnehmung ermöglicht. Der visuelle Cortex nimmt den Großteil des Occipitallappens des Gehirns ein. Nach der Hirnkarte von Korbinian Brodmann entsprechen ihm die Areale 17, 18 und 19. Er wird in die primäre Sehrinde (V1) und sekundäre bzw. tertiäre (assoziative, V2–V5) Bereiche unterteilt. Histologisch zeichnet ihn bei Primaten die hohe Zelldichte und die vergleichsweise geringe Dicke (beim Menschen 1,5–2 mm) aus. Das Areal 17 (auch Area striata) repräsentiert unmittelbar die gegenseitige Hälfte des Gesichtsfeldes und ist „retinotop“ aufgebaut, was bedeutet, dass auf der Netzhaut (Retina) nebeneinander abgebildete Punkte auch hier nebeneinander liegen. Die Zellen, die die Fovea repräsentieren, sind dabei deutlich überrepräsentiert und nehmen etwa die Hälfte der Area striata ein.
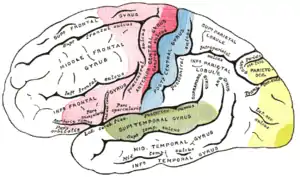
| Primär-motorisches Areal | |
| Prä/Supplementär-motorische Areale | |
| Primär-sensible Areale | |
| Sensible Assoziationsareale | |
| Sehfelder | |
| Hörfelder |
Primäre visuelle Rinde (V1)
Histologie
Der primäre visuelle Cortex (Brodmann-Areal 17, V1; Area striata) ist wie der gesamte Neocortex (unterschieden von Archi- und Paläocortex) in sechs Schichten eingeteilt:
- Schicht I: Molekularschicht
- Schicht II: äußere Körnerschicht
- Schicht III: äußere Pyramidenschicht
- Schicht IV: innere Körnerschicht
- Schicht V: innere Pyramidenzellschicht
- Schicht VI: multiforme Schicht.
Neben der Schichtenorganisation ergibt sich in V1, wie im übrigen Cortex auch, eine kolumnäre (säulenartige) Struktur, die hier jeweils eine Orientierung wiedergibt.
Afferenzen und innere Organisation
Das primäre visuelle Cortexareal V1 verfügt über eine besonders dicke Schicht IV. Hier enden viele afferente Fasern der Neurone des Corpus geniculatum laterale (CGL). Die Schicht IV wird weiter unterteilt in A, B und C, C wiederum in Cα und Cβ. In Schicht IVCα enden die Axone der magnozellulären Schichten 1 und 2 des CGL. Diese Neurone wiederum leiten sich von Ganglienzellen der Netzhaut mit großen rezeptiven Feldern ab, die eine hohe zeitliche aber geringe räumliche Auflösung aufweisen. Die Arbeiten von David Hubel und Torsten Wiesel erbrachten die Erkenntnis, dass Neurone der Schicht IVCα orientierungsselektiv sind. Einzelne Gruppen dieser Zellen antworten jeweils auf visuelle Reize entlang des Zentrums ihrer länglichen rezeptiven Felder bis zu einer Abweichung von 10°. Diese Zellen werden simple cells genannt. Sie projizieren in die Schicht IVB, wo sie binokular verschaltet werden und Richtungsselektivität aufweisen (M-Kanal).
Parvozelluläre Neurone der Schichten III–VI des CGL enden in Schicht IVCβ sowie zum geringen Teil in der Schicht IVA und I. Sie erhalten Afferenzen der Ganglienzellen mit kleinen rezeptiven Feldern (Fovea), zeigen Farbsensitivität und besitzen eine hohe räumliche Auflösung.
Aus den Eingängen der Schicht IVCα erhalten weitere Cortexareale die Informationen über Bewegungen im visuellen Feld. Aus den Eingängen der Schicht IVCβ erhalten andere Cortexareale die Informationen über Form und teilweise Farbe von Objekten im visuellen Feld.
Läsionen von magnozellulären Schichten bei Affen hatten zur Folge, dass diese die Fähigkeit der Bewegungswahrnehmung verloren, wohingegen eine Schädigung der parvozellulären Schichten die Wahrnehmung von Farbe, feinen Texturen sowie Formen und räumlicher Tiefe verhinderte.[1]
Die Schicht IV weist okulare Dominanzbänder auf, auch als Okuläre Dominanzsäulen bezeichnet, die sich durch retrograden axonalen Transport von radioaktivem Prolin (oder anderen Aminosäuren) darstellen lassen.
Sekundäre und tertiäre visuelle Rinde (V2–V5)
Der V2 wird nach den Arealen von Korbinian Brodmann in Areal 18 verortet und hat eine Streifenstruktur, die sich durch histologische Färbung der Cytochromoxidaseaktivität sichtbar machen lässt. Diese Streifen werden nach Livingstone und Hubel (1988) für die Verarbeitung von einzelnen Aspekten der Wahrnehmung, Form, Wahrnehmung und Stereosehen in Verbindung gebracht und haben dann Efferenzen in die höheren Hirnareale.
Nach der Verschaltung in V1 und V2 teilt sich die Verarbeitung in einen dorsalen Pfad durch den Parietallappen, in die motorischen Areale entlang der Zentralfurche und einen ventralen Pfad, der in die Areale des Temporallappens führt, denen man den Anfang der Objekterkennung zuschreibt.[2] Dem ventralen Pfad werden unter anderem V4 und der inferotemporale Cortex zugerechnet. Der dorsale Pfad läuft durch V5 (auch mediotemporaler Cortex genannt). Die Assoziationscortices sind allerdings insgesamt nicht so genau kartiert wie z. B. V1, da sich interindividuelle Unterschiede ergeben.
Siehe auch
Literatur
- Otto Detlev Creutzfeldt: Cortex cerebri. Springer (1983). ISBN 3-540-12193-5
- Karl Zilles, G. Rehkämper: Funktionelle Neuroanatomie. Springer, Berlin 1993. ISBN 3-540-54690-1
- D. Drenckhahn, W. Zenker: Benninghoff. Anatomie. Urban & Schwarzenberg, München 1994 ISBN 3-541-00255-7
- Eric Kandel, James Schwartz, Thomas Jessell: Neurowissenschaften. Spektrum Akademischer Verlag, Heidelberg 1996 ISBN 3-86025-391-3
- Edwin Clarke, C. D. O'Mally: The Human Brain and Spinal Cord: A Historical Study Illustrated by Writings from Antiquity to the 20th Century, University of California Press, 1968
- Mitchell Glickstein, Giacomo Rizzolattic: Francesco Gennari and the structure of the cerebral cortex, Trends in Neurosciences, Band 7, Heft 12, S. 464–467, 1. Dezember 1984, doi:10.1016/S0166-2236(84)80255-6
- Mitchell Glickstein, David Whitteridge: Tatsuji Inouye and the mapping of the visual fields on the human cerebral cortex, Trends in Neurosciences, Band 10, Heft 9, S. 350–353, 1. Januar 1987, doi:10.1016/0166-2236(87)90066-X
Weblinks
Einzelnachweise
- Jochen Müsseler, Allgemeine Psychologie, 2. Auflage (Spektrum Akademischer Verlag (Springer), Heidelberg 2008), S. 18
- E.Bruce Goldstein Wahrnehmungspsychologie: Der Grundkurs, 7. Auflage (Spektrum Akademischer Verlag (Springer), Heidelberg 2007)