Rückenmark
Das Rückenmark (lateinisch Medulla spinalis[1] oder Medulla dorsalis,[2] altgriechisch ῥαχίτης μύελος rhachítes mýelos[3] oder νωτιαῖος μύελος nōtiaios mýelos[3]) wird gemeinsam mit dem Gehirn von meningealen Hüllen umgeben und bildet zusammen mit diesem das zentrale Nervensystem der Wirbeltiere. Es ist dessen einfacher gebauter Teil, der im Wirbelkanal innerhalb der Wirbelsäule liegt. Zum Schädel hin geht das Rückenmark ohne scharfe Grenze in das Verlängerte Mark (Medulla oblongata) des Rautenhirns über. Dem Rückenmark sind über die Spinalnerven wesentliche Anteile des peripheren Nervensystems des Körpers mit Ausnahme des Kopfes angeschlossen. Damit sind den Segmenten des Rückenmarks neben der gesamten (somatischen) Innervation von Rumpfwand und Extremitäten auch bedeutende Anteile der (viszeralen) Innervation von Eingeweiden von Brust-, Bauch- und Beckenhöhle zugeordnet.

Lagebeziehungen
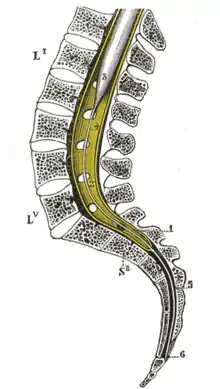
Das Rückenmark entwickelt sich aus dem hinteren Neuralrohr und liegt als zylindrischer Strang von bindegewebigen Häuten (Meningen) umhüllt, von Gehirn-Rückenmarks-Flüssigkeit (Liquor cerebrospinalis) umspült und von der Wirbelsäule umschlossen im Rückenbereich der Wirbeltiere. Beim Menschen geht es kranial etwa auf Höhe des großen Hinterhauptlochs ohne scharfe Grenze in das verlängerte Mark des Hirnstamms über; am kaudalen Ende verjüngt es sich kegelförmig zum Conus medullaris (Markkegel), dessen Spitze fadenförmig ausläuft mit dem Filum terminale (Endfaden).
Während das Rückenmark vor dem 3. menschlichen Embryonalmonat den Wirbelkanal noch in ganzer Länge ausfüllt, verschiebt sich wegen des relativ stärkeren Längenwachstums der Wirbelsäule schon pränatal in der fetalen Entwicklung regelmäßig das Conusende nach kranial,[4] was auch als Aufstieg oder Aszensus bezeichnet wird. Beim Neugeborenen liegt die Conusspitze normalerweise etwa in Höhe des dritten Lendenwirbelkörpers (L3),[5] beim ausgewachsenen Menschen endet das Rückenmark in 95 % bei L2 oder höher.[6]
Infolgedessen legen die aus dem Rückenmark beidseits austretenden Wurzeln der Spinalnerven einen nach unten hin zunehmend längeren Weg innerhalb des Wirbelkanals zurück, bevor sie ihn durch ihr Zwischenwirbelloch verlassen. Unterhalb der Conusspitze verlaufen so neben dem Filum terminale nur noch Wurzelfasern der paarigen Spinalnerven als Cauda equina (Pferdeschweif). Beim Erwachsenen kann daher bei einer Lumbalpunktion, normalerweise nach Einstich zwischen den Dornfortsätzen des 3. und 4. Lendenwirbels, Liquor entnommen oder ein Medikament injiziert werden, ohne Gefahr, das Rückenmark unmittelbar zu verletzen.
Ein Aszensus tritt auch bei anderen Säugetieren auf – und endet hier schon auf Höhe der hinteren Lendenwirbel –, nicht aber bei sonstigen Wirbeltieren. Beim Menschen können Störungen des Aszensus nicht nur zu einem Konustiefstand führen, sondern bei abnorm vermehrtem Zug an Rückenmark und Nervenfasern auch zu dem klinischen Bild eines Tethered cord.[7]
Rückenmarkshäute
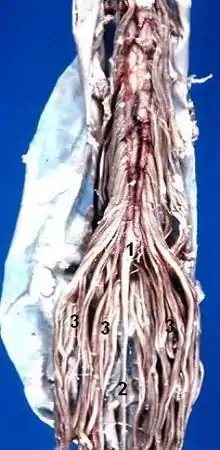
(1) Conus medullaris,
(2) Filum terminale,
(3) Cauda equina.
Die Rückenmarkshäute (Meninges medullae spinalis) gehen in die Hirnhäute über und sind wie diese unterschieden in die äußere harte Hirnhaut, Dura mater, und die beiden weichen Hirnhäute, die mittlere Arachnoidea (Spinnengewebshaut) und die direkt dem Rückenmark anliegende Pia mater. Verglichen mit der häutigen Umhüllung im Bereich des Gehirns fallen einige bauliche Besonderheiten auf. So ist die Dura mater nicht mit dem Periost der knöchernen Elemente des Wirbelkanals verwachsen. Daher ist zwischen Knochen und Dura mater ein Epiduralraum bzw. Periduralraum ausgebildet. Er enthält neben einem dichten Venenplexus auch Binde- und Fettgewebe. In den Epiduralraum lassen sich beispielsweise Lokalanästhetika einbringen, um die Erregungsleitung von Nervenfasern aufzuheben (Periduralanästhesie).
Des Weiteren bestehen zwischen den Rückenmarkshäuten segmental ausgebildete Brücken, die das sogenannte Ligamentum denticulatum als „gezahntes Band“ bilden. Diese in Aufsicht dreieckigen Strukturen dienen der stabilisierenden Aufhängung des Rückenmarks in dem mit Liquor cerebrospinalis gefüllten Subarachnoidalraum zwischen Arachnoidea und Pia mater.
Äußere Gestalt
Das Rückenmark ist ein weißlicher, runder Strangzylinder, der in der Intumescentia cervicalis bzw. der Intumescentia lumbosacralis verdickt ist, da diese Bereiche neben dem Rumpf auch die oberen bzw. unteren Extremitäten innervieren. Nach kaudal hin wird das Rückenmark ansonsten wegen der geringeren auf- und absteigenden Verbindungen mit Hirnregionen etwas schmaler und läuft am unteren Ende als Conus medullaris (Markkegel) spitz aus. Dessen Spitze setzt das etwa 1 mm dicke Filum terminale (Endfaden) fort, das neben dem Bindegewebe der Pia mater auch Gliazellen und Fett enthält. Es verbindet die Conusspitze mit dem unteren Ende des Duralsacks bzw. darüber beim Menschen mit der Hinterfläche des Steißbeins.
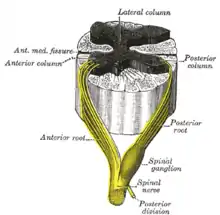
Auf der Vorderseite des Rückenmarks verläuft von oben nach unten eine feine Furche, die bauchseitig (ventral) als Fissura mediana ventralis linke von rechter Seite scheidet. Rückseitig entspricht dem hinten ein tieferer Graben zwischen beiden Seiten, der Sulcus medianus posterior. Im seitlichen Umfang lassen sich jederseits drei Stränge (lat. funiculi) voneinander abgrenzen: paramedian der Fissura mediana der Vorderstrang (Funiculus anterior, in der Tieranatomie Funiculus ventralis), hinten der Hinterstrang (Funiculus posterior, in der Tieranatomie Funiculus dorsalis) und zwischen den beiden der Seitenstrang (Funiculus lateralis).
In der Einfurchung einer Seite zwischen Hinter- und Seitenstrang, dem Sulcus lateralis posterior, treten jeweils die Hinterwurzeln der Spinalnerven ein, und zwischen dem Seiten- und dem Vorderstrang, im Sulcus lateralis anterior, treten jeweils deren Vorderwurzeln aus dem Rückenmark. Als Fila radicularia bilden die Vorderwurzeln und die Hinterwurzeln zwei kontinuierliche Reihen auf jeder Seite. Sie vereinen sich erst in Höhe der Zwischenwirbellöcher (Foramina intervertebralia), beim Durchtritt durch die Wirbelsäule (Columna vertebralis), zu den einzelnen Spinalnerven jeder Seite.
Vögel sind die einzigen rezenten Wirbeltiere, bei denen im Bereich der Intumescentia lumbosacralis zwischen die Hinterstränge beider Seiten der sogenannte Glykogenkörper eingelagert ist (auch Corpus gelatinosum genannt). Es handelt sich hierbei um eine gallertige, eiförmige Bildung mit glykogenreichen Gliazellen, deren funktionelle Bedeutung bislang nicht geklärt ist.[8] Beim Haushuhn ist sie etwa 1 cm lang und 4 mm breit.
Gliederung
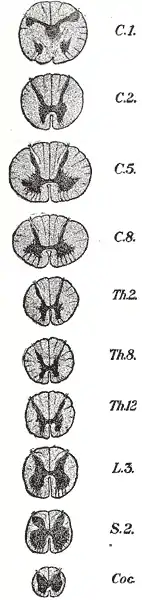
Das Rückenmark lässt sich äußerlich je nach den gemeinsamen Wurzeln eines Spinalnervenpaares in segmentale Abschnitte gliedern, die fünf aufeinander folgenden Bereichen zugeordnet sind:
- Hals- oder Zervikalmark (Pars cervicalis), bei Säugetieren mit 8 Segmenten (C1–C8)
- Brust- oder Thorakalmark (Pars thoracica), beim Menschen mit 12 Segmenten (Th1–Th12)
- Lenden- oder Lumbalmark (Pars lumbalis), beim Menschen mit 5 Segmenten (L1–L5)
- Kreuz- oder Sakralmark (Pars sacralis), beim Menschen mit 5 Segmenten (S1–S5)
- Schwanz- oder Kokzygealmark (Pars coccygis), beim Menschen meist 1 Segment (Co1)
Eines der beim Menschen meist 31 Segmente entspricht dabei jeweils einem Rückenmarksabschnitt, dem die Wurzelfasern für je einen linken und einen rechten Spinalnerven zugehören. Das zugehörige Spinalnervenpaar verlässt den Wirbelkanal oberhalb bzw. unterhalb eines Wirbels und tritt beidseits durch Zwischenwirbellöcher aus, die meist nicht mehr genau auf Höhe seines Rückenmarksegments liegen. Nach diesen Austrittstellen wird es einem Wirbelsäulenbereich zugeordnet und benannt.
Im Thorakalbereich sowie weiter kaudal heißt ein Spinalnerv samt seinem Segment nach dem darüber liegenden Wirbel und die Anzahl der Rückenarksegmente stimmt mit der Anzahl der Wirbel überein. Dem Zervikalbereich sind dagegen bei sieben Halswirbeln acht Segmente des Halsmarks zugeordnet, da der erste Spinalnerv (C1) nicht unter, sondern über dem ersten Halswirbel, Atlas, austritt; das unterhalb des siebten Halswirbels austretende Spinalnervenpaar (C8) ist noch dem Halsbereich zugeordnet. Was oberflächlich einem Zaunpfahlproblem gleicht, findet seinen tieferen Grund darin, dass Teile des Hinterhauptbeines (Os occipitale) entwicklungsgeschichtlich als verschmolzene Wirbelanlagen betrachtet werden können.
Feinbau
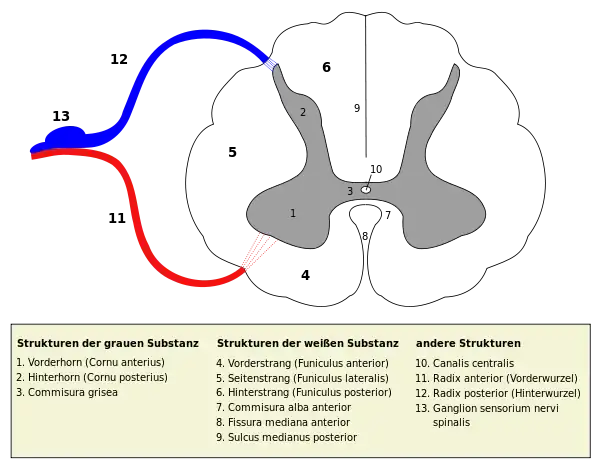
Das Rückenmark besteht aus Grauer Substanz (Substantia grisea) und Weißer Substanz (Substantia alba). Die Graue Substanz besteht überwiegend aus Nervenzellkörpern (Perikaryen). Sie liegt im Rückenmark innen, umgibt den Zentralkanal und bildet Kerngebiete in Kernsäulen (Columnae griseae). Beiderseits liegen in der Vordersäule (Columna ventralis) Perikaryen von somatomotorischen Nervenzellen, in der Seitensäule (Columna lateralis bzw. intermedia) von viszeromotorischen, während die Hintersäule (Columna dorsalis) somatosensible und viszerosensible Nervenzellkörper enthält. Die Weiße Substanz besteht dagegen vornehmlich aus Nervenfasern. Sie umschließt die Graue Substanz des Rückenmarks vollständig, ist ihr in Form von Strängen (Funiculi) außen angelagert und enthält in der Commissura alba von einer zur anderen Seite wechselnde Nervenfasern.
Graue Substanz
Die graue Substanz hat im Rückenmarksquerschnitt die Form eines Schmetterlings. Den vorderen, breiteren Flügelteil nennt man Vorderhorn (Cornu anterius, bei Tieren Cornu ventrale), den hinteren, schmaleren Teil Hinterhorn (Cornu posterius bzw. dorsale). Im Bereich des Thorakal- und des Lumbalmarks ist zwischen Vorder- und Hinterhorn das kleinere Seitenhorn (Cornu laterale) ausgeprägt. Hierbei wird eine quergeschnittene Kernsäule als Horn bezeichnet.
Der zwischen den beiden schmetterlingsflügelähnlichen Seiten gelegene Bereich (Substantia intermedia centralis) enthält innerhalb der grauen Substanz Querverbindungen, als Commissura grisea. Sie verlaufen in der Zentralregion vor und hinter dem in der Mitte liegenden Zentralkanal (Canalis centralis). Dieser röhrenförmige Hohlraum ist mit Liquor gefüllt und stellt den inneren Liquorraum des Rückenmarks dar.
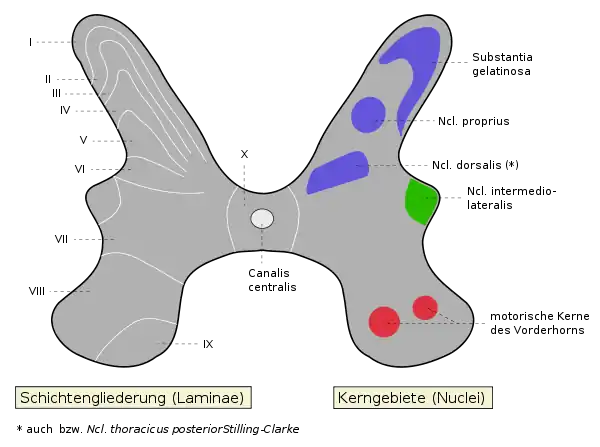
Die graue Substanz kann in zehn Schichten (Laminae) eingeteilt werden, die zum Teil auch unter anderen Namen bekannt sind. So wird die Lamina II auch als Substantia gelatinosa [Rolandi] (nach Luigi Rolando) bezeichnet. Die Laminae I–VII befinden sich im Hinterhorn, die Laminae VIII und IX im Vorderhorn. Die Lamina X enthält die Commissura grisea anterior bzw. posterior. Funktionell können verschiedene einzelne Kerne innerhalb der Kernsäulen (Columnae) und der Zwischenzone unterschieden werden (siehe Abbildung).
Nervenzelltypen
In der Zytoarchitektur der spinalen grauen Substanz lassen sich verschiedene Zelltypen hervorheben:
- Wurzelzellen, deren Axone die Vorderwurzel bilden, können unterschiedlich groß sein. Besonders große liegen im Vorderhorn und leiten als Motoneuronen ihre Signale zu Muskelfasern von Skelettmuskeln.
- Binnenzellen, deren Axone das Rückenmark nicht verlassen. Sie stellen den sogenannten Eigenapparat des Rückenmarks dar, auch propriospinales System genannt. Die neuronalen Netze dieses rückenmarkeigenen Binnensystems realisieren die spinalen Integrationsleistungen. Sie können beispielsweise auf einfachen (monosynaptischen) Reflexbögen aufbauend Motoneuronen untereinander verknüpfen und so im Muskelspiel einzelne Muskeln in Muskelgruppen als Mitspieler oder Gegenspieler zueinander in Beziehung setzen und ihre Aktivität aufeinander abstimmen. Zum Eigenapparat gehören Schaltzellen, Assoziationszellen und Kommissurenzellen.
- Schaltzellen sind Interneuronen, deren Fortsätze die graue Substanz eines Rückenmarkssegments nicht verlassen. Sie stellen auf der gleichen Seite (ipsilateral) Verknüpfungen zwischen Zellen des gleichen Segments her (intrasegmental) und vermitteln z. B. Eigenreflexe. Zu diesen Interneuronen gehören auch die Renshaw-Zellen, die rückkoppelnd einen hemmenden Einfluss auf Motoneuronen haben.
- Assoziationszellen sind propriospinale Neuronen, die verschiedene Rückenmarkssegmente untereinander verbinden. Ihre Fortsätze verlaufen ebenfalls ipsilateral, doch längs zwischen verschiedenen Segmentebenen (intersegmental), in der grauen Substanz und den ihr anliegenden Bahnen der Grundbündel (Fasciculi proprii).
- Kommissurenzellen sind Neuronen des Eigenapparates, deren Neuriten die Medianebene kreuzen und (als Commissura) auf die andere Seite ziehen. Sie verknüpfen innerhalb eines Rückenmarkssegments mit der Gegenseite (kontralateral) und sorgen dafür, dass Erregungsimpulse auch die andere Seite erreichen. Derart vermitteln sie auch gekreuzte Reflexe, bei denen die andere Körperseite reagiert, wie sie für verschiedene tetrapodische Bewegungsmuster typisch sind.
- Strangzellen sind Projektionsneuronen, deren Axone die graue Substanz verlassen und in der weißen Substanz weiterziehen. Hier steigen sie in Strängen ab oder, häufiger, auf und leiten Signale an andere Rückenmarkssegmente und an Abschnitte des Gehirns. Sie gehören somit nur teilweise zum Eigenapparat des Rückenmarks.
An der Auslösung von Fremdreflexen sind oft Assoziations-, Kommissuren- und Strangzellen beteiligt.
Hinterhorn
Das Hinterhorn erhält über die rückenseitige (dorsale), sensible Wurzel des Spinalnervs sensible Informationen aus der Peripherie. Diese Informationen werden in Richtung Gehirn weitergeleitet, aber teils auch bereits im Rückenmark verarbeitet.
Die Hinterwurzel wird von Axonen pseudounipolarer Nervenzellen gebildet, deren Zellkörper außerhalb des Rückenmarks im Spinalganglion liegen. Ihre Axone treten über die dorsale (sensible) Wurzel in das Rückenmark, wo ein Teil im Hinterhorn auf ein zweites Neuron umgeschaltet wird. Andere Hinterwurzelaxone verlaufen ohne Umschaltung bis in die Medulla oblongata.
Das Hinterhorn, welches embryologisch aus der Flügelplatte hervorgeht, besteht von dorsal nach ventral (vom Rücken zum Bauch hin) aus einer dünnen Zona marginalis, an die sich die Substantia gelatinosa anschließt, in der Schmerzafferenzen der Hautrezeptoren auf das 2. Neuron umgeschaltet werden, bevor sie im Tractus spinothalamicus lateralis zum Thalamus ziehen. Hier hemmen Interneurone, die durch den Transmitter Serotonin aktiviert werden, die Schmerzweiterleitung durch die Freisetzung von Endorphinen. Hierdurch wird die Substantia gelatinosa klinisch äußerst relevant und bietet Ansätze für die Erklärung von Mechanismen der umstrittenen Akupunktur oder der schmerzhemmenden Wirkung einiger Antidepressiva.
In den Laminae 3 und 4 liegt der Nucleus proprius, der in gesamter Höhe des Rückenmarks zu finden ist. Er ist Ziel sensibler Afferenzen der Tiefensensibilität und bildet in seinem kranialen (zum Kopf führenden) Verlauf den Tractus spinocerebellaris anterior.
Laminae 5 und 6 beinhalten schließlich den Nucleus dorsalis (Stilling-Clarke), der auch Nucleus thoracicus posterior genannt wird. Ebenso wie der Nucleus proprius erhält der Nucleus dorsalis Afferenzen der Tiefensensibilität (Muskelspindeln, Gelenk- und Sehnenrezeptoren), welche über den Tractus spinocerebellaris posterior zum Kleinhirn geleitet werden.
Vorderhorn
Das Vorderhorn entsteht aus der Grundplatte und enthält Perikarya, deren Axone als Radix anterior das Rückenmark verlassen, um die Skelettmuskulatur zu innervieren. In der Schichtengliederung bestehen die Vorderhörner aus den Laminae 8 und 9. Hier kommen die großen multipolaren α-Motoneurone und auch die etwas kleineren ɣ-Motoneurone sowie Renshaw-Zellen vor. Hier enden die Impulse der aus dem Gehirn kommenden Bahnen (Pyramidenbahn und extrapyramidale Bahnen) sowie die Impulse der Muskelreflexe und werden als „gemeinsame motorische Endstrecke“ zum Erfolgsorgan – der Skelettmuskulatur – weitergeleitet.
Seitenhorn
Im Bereich des Brustmarks und, etwas weniger deutlich, im Lendenmark befindet sich, zwischen dem Vorder- und dem Hinterhorn liegend, das sogenannte Seitenhorn, das von den Zellkörpern vegetativer Neurone gebildet wird und zum Sympathikus gehört. Ihre efferenten Fasern verlassen das Rückenmark über die Vorderwurzel, ziehen zum Grenzstrang und werden zum Teil dort auf das zweite Neuron verschaltet. Der Rest zieht zu den prävertebralen Ganglien, um dort verschaltet zu werden. Sympathisch-afferente Fasern gelangen aus der Peripherie mit der Hinterwurzel zum Seitenhorn. An äquivalenter Stelle liegen im Sakralmark Zellkörper parasympathischer Neurone, allerdings ohne ein erkennbares Seitenhorn zu bilden. Die Axone ziehen als Nervi splanchnici pelvici zum Ganglion pelvinum sowie zu den intramuralen Ganglien.
Weiße Substanz
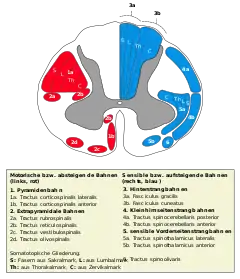
In der weißen Substanz des Rückenmarks verlaufen aufsteigende (meist sensible) Bahnen, die zum Gehirn ziehen, und absteigende (meist motorische) Bahnen, die vom Gehirn kommen. Ursache für die weiße Farbe ist die Myelinisierung der Axone.
Aufsteigende Bahnen
Die Hinterstrangbahn leitet Information von Mechanorezeptoren für epikritische Sensibilität und Tiefensensibilität jeweils der gleichen Körperseite zunächst bis in das Verlängerte Mark (Medulla oblongata). Die Zellkörper der im Hinterstrang aufsteigenden Axone liegen in den Spinalganglien und sind jeweils verschiedenen Segmenten des Rückenmarks zugeordnet. Ohne Umschaltung im Hinterhorn ziehen die Fasern ins Markhirn, und endigen in den dortigen Kerngebieten, dem Nucleus gracilis für Informationen aus der unteren Körperhälfte und dem Nucleus cuneatus für solche aus der oberen Körperhälfte.
Der Hinterstrang zeigt im oberen Rückenmark jederseits eine Unterteilung in den Fasciculus cuneatus und den Fasciculus gracilis, deren primärafferente Fasern jeweils in den gleichnamigen schon genannten Kernen auf sekundär-afferente Neuronen umgeschaltet werden. Die weiter aufsteigenden Fasern dieser (2.) Neuronen in den Hinterstrangkernen kreuzen in der Medulla oblongata auf die Gegenseite (Decussatio lemniscorum medialium) und verlaufen im Lemnicus medialis zu kontralateralen Thalamuskernen (VPL). Dieser Bahnabschnitt ist namensgebend für das lemniskale System.
Dagegen leiten die Tractus spinothalamicus lateralis und ventralis [anterior] als Vorderseitenstrangbahn Informationen für die protopathische Sensibilität (Schmerz-, Temperatur- sowie grobe Druckempfindung) zu verschiedenen Kerngebieten im Thalamus (u. a. MD, VPL, VPM). Die Fasern des Tractus spinothalmicus gehen von Nervenzellkörpern im Rückenmark aus, deren Axone bereits auf Höhe des Ursprungssegmentes zur Gegenseite kreuzen. Diese Strangzellen im Hinterhorn sind sekundäre (2.) afferente Neuronen, sie erhalten Primärafferenzen der Spinalganglienzellen mit Information von Mechanorezeptoren sowie von Nozizeptoren und Thermorezeptoren. Diese spinothalamische Bahn im Vorder- und Seitenstrang ist Teil des sogenannten anterolateralen Systems.
Der Tractus spinocerebellaris anterior (Kleinhirnseitenstrangbahn oder Gowerssches Bündel) entspringt vor allem aus dem Nucleus dorsalis des Hinterhorns und endet im Kleinhirn. Er verläuft entweder ungekreuzt oder aber kreuzt die Seite zweimal und leitet propriozeptive (= Informationen über Lage und Stellung von Gelenken, Muskeln, Sehnen) Informationen.
Der Tractus spinocerebellaris posterior (Flechsig-Bündel) geht von den Zellen des Nucleus thoracicus aus und erreicht ungekreuzt über den Vorderseitenstrang das Kleinhirn. Er dient der Tiefensensibilität.
Absteigende Bahnen
Die Pyramidenbahn (Tractus corticospinalis) reicht vom motorischen Cortex der Großhirnrinde bis ins Vorderhorn.
Als extrapyramidale Bahnen bezeichnet man alle motorischen Bahnen, die nicht zur Pyramidenbahn gehören. Zu ihnen gehören der Tractus rubrospinalis, der Tractus vestibulospinalis, der Tractus reticulospinalis und der Tractus tectospinalis. Sie alle enden ebenfalls im Vorderhorn.
Neurotransmitter
Der wichtigste inhibitorische Neurotransmitter im Rückenmark ist Glycin. Interneurone und z. B. die Renshaw-Zellen sind glycinerg, d. h., sie setzen Glycin aus ihren synaptischen Endigungen frei.
Embryologie
Das Rückenmark entsteht beim Wirbeltier-Embryo aus den Neuralfalten des Ektoderms, unmittelbar über der Chorda dorsalis. Die Neuralfalten verschließen sich zum Neuralrohr, einem mit Flüssigkeit gefüllten Kanal, der von einem Epithel umgeben ist. Bei Störungen dieses Verschlusses treten verschiedene Formen der Dysraphie auf. Es kann eine Spina bifida aperta („offener Rücken“) entstehen, bei der die Wirbelbögen rückenseitig unvollständig bleiben. Dabei können sich nur die Rückenmarkshäute (Meningozele) oder zusätzlich auch das Rückenmark (Meningomyelozele) durch diesen Defekt nach außen vorwölben. Dort, wo sich die Neuralrinne selbst nicht verschließt, entsteht eine Myeloschisis (Rachischisis). Erfolgt dieser Defekt am rostralen (vorderen) Ende der Neuralrinne, kommt es sogar zu einem Ausbleiben der Bildung des Gehirns (Anenzephalie).
Aus dem Neuralepithel differenzieren sich die Nerven- und Gliazellen. Die Spinalganglien und sympathischen Neurone entstehen aus der sogenannten Neuralleiste. Um das Neuralrohr entstehen aus dem Mesenchym die Urwirbel, aus denen unter anderem die Wirbelsäule entsteht.
Zunächst verlassen Proneurone (die Vorläufer der Neurone) das Neuroepithel und bilden eine Mantelschicht (Mantelzone), aus der die graue Substanz entsteht. Die meisten Axone dieser Proneurone wachsen in Richtung Oberfläche und bilden so die Randschicht (Marginalzone), die später zur weißen Substanz wird.
Innerhalb der Mantelschicht entwickeln sich durch Zellteilungen und Zellmigration zwei Verdickungen, die Flügelplatte (Alarplatte) und die Grundplatte (Basalplatte), die nach und nach das spätere Erscheinungsbild eines Schmetterlings aufweisen. Die Randschicht wird durch das weitere Einwachsen auf- und absteigender Axone, deren Myelinisierung und die Vergrößerung der Flügel- und Grundplatten in 3 Stränge gegliedert: Hinterstrang, Seitenstrang und Vorderstrang.
Beim Tethered cord unterbleibt die relative Verkürzung des Rückenmarks.
Blutversorgung
Drei Arterien versorgen das Rückenmark: Vorn die Arteria spinalis anterior, hinten zwei Arteriae spinales posteriores. Bei Tieren mit horizontaler Körperstellung wird stattdessen von Arteria spinalis ventralis und dorsalis gesprochen. Diese drei Arterien entspringen im Halsbereich beidseits aus den seitlich in der Wirbelsäule verlaufenden Arteriae vertebrales. Im Thorax- und Lendenbereich erhalten sie segmentale Zuflüsse aus den Interkostal- (Arteriae intercostales posteriores) und den Lendenarterien (Arteriae lumbales). Die größte und wichtigste Segmentarterie ist die Adamkiewicz-Arterie (Arteria radicularis magna), die in der Regel zwischen dem 9. und 12. Brustsegment liegt. Ein Verschluss dieses Gefäßes ist besonders folgenreich für die Blutversorgung des thorakalen Rückenmarks.
Im Bereich zwischen dem 4. und dem 6. Brustwirbel ist die Ausbildung von solchen Zuflüssen gering, sodass bei einem Gefäßverschluss dieser Abschnitt besonders gefährdet ist, einen Rückenmarksinfarkt zu erleiden. Das venöse Blut wird über zwei große Venen abtransportiert, die teilweise parallel zu den Arterien verlaufen: Vena spinalis anterior und Vena spinalis posterior.
Phylogenese
In der hier beschriebenen Form ist das Rückenmark bei allen Vertretern der Schädeltiere (Gnathostomata) aufgebaut, wobei es in den unterschiedlichen Taxa jeweils geringe Unterschiede im Detail gibt.
Bei den basalen Chordatieren, zu denen die Schädellosen (Acrania) sowie die Manteltiere (Tunicata) gehören, gibt es kein differenziertes Rückenmark. Hier existiert stattdessen nur das Neuralrohr, welches dem späteren Rückenmark homolog ist. An seinem Vorderende besitzt es eine Verdickung, die als Hirnbläschen bezeichnet wird, mit dem Gehirn der Wirbeltiere allerdings nicht viel gemein hat. Das Neuralrohr selbst ist aus teilweise hoch differenzierten Neuronen aufgebaut, über Verschaltungen und Funktionen derselben ist allerdings wenig bekannt. Die Hauptaxone gehen von den so genannten Kolossalzellen aus, die große Zwischenneurone darstellen und deren Axone ventromediane Riesenfasern bilden. Segmental befinden sich dorsale Nervenwurzeln, die den späteren Spinalganglien entsprechen, aufgrund der versetzten Myomere allerdings versetzt zueinander sind. Während bei den Acraniern dieser Aufbau bis zum adulten Tier besteht, findet er sich bei den Tunicata nur im Larvenzustand.
Bei den Schleimaalen (Myxinoida) und den Neunaugen (Petromyzontida), die als Kieferlose (Agnatha) die ursprünglichsten Taxa der Schädeltiere (Craniota) darstellen, existiert bereits ein Rückenmark, bei dem das Neuralrohr wie bei den späteren Wirbeltieren nur das Lumen darstellt. Bei den Schleimaalen ist es bandförmig aufgebaut und besitzt verschiedene Motoneurone und Interneurone ohne eine segmentale Anordnung, die Wurzeln der Spinalnerven vereinigen sich erst in der somatischen Muskulatur, und eine Reihe von Organen werden direkt von Spinalnerven innerviert. Das Rückenmark der Neunaugen ähnelt dagegen bereits deutlicher dem der späteren Wirbeltiere. Es weist eine Trennung in weiße und graue Substanz auf, und die Zellkörper der Riesenaxone liegen vornehmlich im Hirnstamm. Andererseits ist die Verschaltung der Neurone in der Wirbelsäule so komplex, dass die Tiere auch nach einer Durchtrennung derselben vor dem Hirn, etwa durch Entfernung des Kopfes, noch koordiniert schwimmen können. Die dorsalen und ventralen Wurzeln der Spinalnerven vereinigen sich bei den Tieren nahe dem Rückenmark, treten jedoch nicht segmental in gleicher Höhe aus dem Rückenmark hervor.
Die Knorpelfische (Chondrichthyes), zu denen die Haie und Rochen gehören, besitzen ein Rückenmark mit segmentalen Spinalnerven, wie es für die Schädeltiere typisch ist. Bei ihnen sind jedoch einige Besonderheiten vorhanden. Auch bei ihnen ist das Rückenmark sehr stark autonom, und ein Hai kann auch dann noch koordiniert schwimmen, wenn das Rückenmark am Hals durchtrennt wird. Anders als bei allen anderen Wirbeltieren nimmt die Anzahl der sensorischen und motorischen Wurzelganglien während des Wachstums lebenslang zu. Neben weiteren Unterschieden im Feinbau, wie etwa einem separaten absteigenden Strang für die Kerne des Hirnnerven VIII, besitzen die Knorpelfische ein kaudales neurosekretorisches System, welches eine wichtige Rolle bei der Osmoregulation sowie bei der Fortpflanzung spielt.
Bei allen folgenden Taxa, von den Strahlenflossern bis zu den Amphibien, den verschiedenen Reptilientaxa sowie den Vögeln und den Säugetieren entspricht das Rückenmark in Aufbau und Funktion dem dargestellten Grundmuster und unterscheidet sich nur in sehr speziellen Details voneinander.
Myelopathien
Myelopathien (Schädigungen des Rückenmarks) werden verursacht durch:
- Degeneration (Degenerative Myelopathien der Hunde), Syringomyelie
- Druck (Tumor, Bandscheibenvorfall, Schleudertrauma)
- Durchblutungsstörungen
- Durchtrennung (Querschnittlähmung)
- Entzündungen (Multiple Sklerose, Poliomyelitis)
- Infektionen wie Neurolues, Sepsis
- Spinale Abszesse
- Missbildungen wie die Chiari-Malformation
Untersuchung des Rückenmarks
Das Rückenmark selbst ist der klinischen Untersuchung durch seine Lage im knöchernen Wirbelkanal nicht direkt zugänglich. Reflexuntersuchungen und das Verteilungsbild (→ Kennmuskeln) von Lähmungen (Paresen) oder Gefühlsstörungen geben dem Neurologen jedoch wichtige Hinweise auf die Art und Lokalisation der Schädigung. Neurophysiologische Untersuchungsmethoden wie motorisch und somatosensibel evozierte Potentiale erlauben eine funktionelle Überprüfung einzelner Bahnsysteme des Rückenmarks wie den Tractus corticospinalis und die Hinterstrangbahnen. Untersuchungen von Reflexbahnen über Motoneurone im Rückenmark sind möglich mit Hilfe des H-Reflexes und der F-Welle.
Strukturelle Veränderungen lassen sich dagegen mit den bildgebenden Verfahren vor allem der Magnetresonanztomografie, der Computertomografie und der Myelografie erkennen. Fehlbildungen im Blutgefäßsystem des Rückenmarks lassen sich auch mit der Angiografie darstellen. Hinweise auf Entzündungen kann auch die Lumbalpunktion und Untersuchung der Rückenmarksflüssigkeit liefern.
Literatur
- Franz-Viktor Salomon, Hans Geyer, Uwe Gille: Anatomie für die Tiermedizin. Lehrbuch. Enke/ MVS Medizinverlage Stuttgart, Stuttgart 2004, ISBN 3-8304-1007-7.
- Theodor Heinrich Schiebler (Hrsg.): Anatomie. Histologie, Entwicklungsgeschichte, makroskopische und mikroskopische Anatomie, Topographie. 9., vollständig überarbeitete Auflage, Springer-Verlag, Berlin 2005, ISBN 3-540-21966-8.
- W. Westheide, R. Rieger: Spezielle Zoologe; Teil 2: Wirbel- und Schädeltiere. Fischer, Stuttgart/ Jena, 2004, ISBN 3-8274-0307-3.
- Emil Villinger: Leitfaden für das Studium der Morphologie und des Faserverlaufs. Engelmann, Leipzig 1905, S. 7, 8, 95, 151 (Online).
Weblinks
Einzelnachweise
- Federative Committee on Anatomical Terminology: Terminologia Anatomica. Medienpaket, Thieme, Stuttgart 1998, ISBN 3-13-115251-6.
- F. J. Siebenhaar: Terminologisches Wörterbuch der medicinischen Wissenschaften. 2. Auflage, Arnoldische Buchhandlung, Leipzig 1850.
- H. G. Liddell, R. Scott: A Greek-English Lexicon. revised and augmented throughout by Sir Henry Stuart Jones. with the assistance of. Roderick McKenzie. Clarendon Press, Oxford 1940.
- auch laut kernspintomographischen Untersuchungen toter Feten, siehe O. J. Arthurs, S. Thayyil, A. Wade, W. K. Chong, N. J. Sebire, A. M. Taylor: Normal ascent of the conus medullaris: a post-mortem foetal MRI study. In: The journal of maternal-fetal & neonatal medicine : the official journal of the European Association of Perinatal Medicine, the Federation of Asia and Oceania Perinatal Societies, the International Society of Perinatal Obstetricians. Band 26, Nummer 7, Mai 2013, S. 697–702, ISSN 1476-4954, doi:10.3109/14767058.2012.746307, PMID 23211127.
- Marjorie A. England: Color Atlas of Life Before Birth. Normal Fetal Development. Wolfe Medical Publications, London 1983, ISBN 0-8151-3119-4.
- A. F. Reimann, B. J. Anson: Vertebral level of termination of the spinal cord with report of a case of sacral cord. In: Anatomical Record-advances in Integrative Anatomy and Evolutionary Biology. Band 88, S. 27–138, 1944.
- D. A. Hertzler, J. J. DePowell, C. B. Stevenson, F. T. Mangano: Tethered cord syndrome: a review of the literature from embryology to adult presentation. In: Neurosurgical Focus. Band 29, Nummer 1, Juli 2010, S. E1, ISSN 1092-0684, doi:10.3171/2010.3.FOCUS1079, PMID 20593997 (Review).
- W. Möller, W. Kummer: The blood-brain barrier of the chick glycogen body (corpus gelatinosum) and its functional implications. In: Cell Tissue Res.. Band 313, Nr. 1, Juli 2003, S. 71–80. PMID 12768407.