Bewegungslernen
Als Bewegungslernen (englisch motor learning) bezeichnet man die relativ dauerhafte Veränderung eines Bewegungsablaufs (Koordinationsmuster der Muskeln) eines Lebewesens, wenn dies durch die Absicht erfolgt, ein bestimmtes Ziel, das man bis dahin noch nicht erreichen konnte, durch diesen Bewegungsablauf zu erreichen (z. B. ein Hindernis überspringen, einen Ball fangen, eine Fremdsprache sprechen lernen oder nach einem Schlaganfall das Gehen wieder erlernen). Dieser Vorgang muss nicht bewusst sein. Auch die Verbesserung eines Bewegungsablaufs (Ökonomisierung, schnellere, flüssigere Ausführung) ist ein Bewegungslernen.
Bewegungslernen findet ständig statt, da es auch zur Anpassung an neue Umweltsituationen dient. Insofern ist das Bewegungslernen ein wesentlicher Bestandteil der evolutionären Entwicklung der Lebewesen. Da das Bewegungslernen durch die Sinnesorgane gesteuert (Suchen und Einschätzen des Ziels) und geregelt wird (ständige online Überprüfung, ob man auf dem richtigen Weg zum Ziel ist), spricht man auch von sensomotorischem Lernen. Berufe, zu deren Ausübung gute Kenntnisse des Bewegungslernens notwendig sind, sind die des Sportlehrers, des Trainers, des Ergo- und des Physiotherapeuten.
Gegenstand des Bewegungslernens
Im Allgemeinen verbinden wir mit dem Begriff des Bewegungslernens die Vorstellung des Lernens von Bewegungsabläufen aus dem Bereich des Sports. Auch das Erlernen oder Verbessern von Bewegungsabläufen in der Physiotherapie ist uns geläufig. Dazu gehört z. B. das Lernen (bzw. Wiedererlernen) des Gehens nach einer Verletzung des Bewegungsapparats (Knochenbruch, Gelenk- oder Sehnen-, Bänderverletzung, Amputation) oder des Nervensystems (Schlaganfall, Querschnittlähmung).
Etwas anders verhält es sich mit dem Bewegungslernen in der Ergotherapie, weil es hier weniger darauf ankommt, den physiologischen Ablauf zu üben, als vielmehr darauf, durch die Bewegung eine Handlung zu vollziehen, die immer einen Zweck, eine bestimmte Bedeutung hat.
Das Gehen ist ein Beispiel für eine Alltagsbewegung, wie Sprechen, Schreiben, Lasten Tragen etc., die zunächst gelernt werden, dann aber ständig unseren aktuellen Umgebungsbedingungen und Veränderungen unseres Körpers (z. B. Wachstum, Altern) angepasst werden müssen. Ein weiterer Bereich des Bewegungslernens betrifft die beruflichen Tätigkeiten. Dazu gehören z. B. Bewegungen beim Handwerken, beim Spielen eines Musikinstruments, beim Malen, Bild-Hauen oder bei der Akrobatik. Besondere Anforderungen an das Bewegungslernen stellen das Singen (Mund-, Rachen- und Körperbewegungen) oder das Erlernen (Motorik der Aussprache) einer fremden Sprache dar.
In der heutigen Zeit der Technisierung der Welt spielen Roboter eine zunehmende Rolle. Deren Bewegungen müssen programmiert werden. Das Wissen dazu wird aus der Kenntnis natürlicher Bewegungen abgeleitet. Die aktuelle Entwicklung geht dahin, dass die Roboter dazu in die Lage versetzt werden sollen, auch Lernprozesse und Optimierungen von Bewegungsabläufen selber zu initiieren und durchzuführen. Daran arbeitet vor allem die Forschergruppe um Daniel M. Wolpert in Cambridge (UK).[1]
Funktion und biologische Grundlagen
Bewegungslernen erfolgt durch Imitation, Ausprobieren (ähnlich wie Trial and Error) oder Instruktion. Meist sind alle diese Vorgehensweisen beteiligt.
Die Fähigkeit lebender Organismen, neue Bewegungen zu erlernen und diese neuen Situationen anzupassen, beruht auf der Plastizität des Nervensystems und den biochemischen Eigenschaften der Nervenzellen. Von den Nervenzellen aus werden die Muskeln aktiviert, das bedeutet, es werden Anfang und Ende sowie die Stärke (Intensität) ihrer Anspannung bestimmt.
Um das Verständnis des Zusammenspiels zwischen Nervenzellen und Muskeln zu verstehen, bedarf es der Kenntnis von Aufbau und Funktionsweise von Muskeln und Nerven.
Aufbau
Es gibt drei verschiedene Muskeltypen (siehe Muskulatur) im menschlichen Organismus: Die glatten Muskeln, den Herzmuskel und die Skelettmuskeln. Sie unterscheiden sich leicht in ihrem Aufbau. Der Skelettmuskel ist die Funktionseinheit im menschlichen Organismus, der für die Bewegungen des Körpers verantwortlich ist. Dafür sind zwei seiner Merkmale verantwortlich:
- Er ist kontraktil, d. h. er kann sich verkürzen (Kontraktion, aktiv) und erschlaffen (passiv).
- Er ist mit mehr als einem Knochen verbunden. Deswegen ist er in der Lage, die Knochen, mit denen er verbunden ist, zu bewegen.
Der Skelettmuskel wird auch als quer gestreifter Muskel bezeichnet, weil seine Feinstruktur im Mikroskop eine Querstreifung zeigt. Der Aufbau, der zu dieser Querstreifung führt, ist dafür verantwortlich, dass sich der Muskel kontrahieren kann.
Ein quer gestreifter Muskel besteht aus mehreren (die Anzahl ist von seiner Größe abhängig) parallelen Muskelfaserbündeln. Er ist von einer Bindegewebshülle, der Muskelfaszie, umgeben. Die Muskelfaserbündel bestehen ihrerseits wiederum aus Muskelfasern, das sind die Muskelzellen. Auch diese wiederum bestehen aus einer größeren Anzahl von parallelen Strukturen, den Myofibrillen, die für die Kontraktion des Muskels verantwortlich sind. Auch diese sind mit einem Bindegewebe eng umgeben und bilden mit diesem eine Funktionseinheit.
Funktion
Für jede Bewegung des Körpers sind mindestens 2 Muskeln notwendig. Einer, der Agonist (griech.: αγω = tun, in Bewegung setzen), der sich kontrahiert und dadurch aktiv die Bewegung in einem Gelenk hervorruft, ein anderer Muskel in der Regel an der „Gegenseite“ des Gelenks gelegen, muss diese Bewegung zulassen, indem er sich dehnen lässt. Dieser Muskel wird als Antagonist (griech. αντι ‚entgegen‘) bezeichnet. Beim Lernen eines neuen Bewegungsablaufs muss die Arbeit dieser Muskeln so aufeinander abgestimmt werden, dass eine erfolgreiche und geschmeidige Bewegung entsteht. Wird eine Bewegung zum ersten Mal ausgeführt, werden meistens Agonist und Antagonist zusammen aktiviert und kontrahieren sich. Das ist nicht sehr ökonomisch. Im Laufe des Lernprozesses, also der Übung bzw. der Wiederholungen muss der Antagonist „lernen“ die Aktivierung „zurückzufahren“ und der Agonist, nur so viel an Kontraktion aufzubringen, dass die Bewegung erfolgreich und ökonomisch wird. Das muss für alle an der Bewegung beteiligten Muskeln geleistet werden und erfolgt durch interne Rückkoppelungs-(Regelungs-)prozesse.
Die Aktivierung bzw. Nichtaktivierung der einzelnen Muskeln erfolgt (als die letzte gemeinsame Endstrecke der Motorik) über die motorischen Nervenzellen, die Motoneurone im Rückenmark. Von dort ziehen Nervenleitungen zu den Muskelfasern in den Muskeln. Dort enden sie an speziellen Strukturen, den Motorischen Endplatten. Wird ein elektrisches Signal (Aktionspotential) vom Motoneuron über diesen Nerv bis zu den motorischen Endplatten am Muskel gesendet, wird dieses Signal unter Umwandlung in einen chemischen Prozess auf den Muskel übertragen. Dort kommt es durch spezifische chemische, elektrische und mechanische Prozesse zur Kontraktion der Muskelfasern. Es werden jeweils mehrere, aber unterschiedlich viele, Muskelfasern von einem Motoneuron gleichzeitig innerviert. Diese Gruppe zusammen mit dem betreffenden Motorneuron wird als motorische Einheit bezeichnet.
Ein Muskel enthält viele – häufig über 1000 – Muskelfasern, die zu verschiedenen Motorischen Einheiten zusammengefasst sind. Es werden bei der Ausführung eines Bewegungsablaufs nie alle Motorischen Einheiten eines Muskels innerviert – außer in absoluten Notfällen. Die Auswahl der aktivierten Motorischen Einheiten folgt zwar einer bestimmten Regel (dem size principle: Das bedeutet, es werden zuerst kleine motorische Einheiten innerviert, dann immer größere), aber es müssen nicht immer bei jedem Ablauf dieselben sein.
Veränderung durch Lernen
Beim Lernen neuer bzw. der Modifikation von Bewegungsabläufen werden am Beginn des Lernprozesses immer sehr viel mehr Motorische Einheiten für den Ablauf aktiviert, als unbedingt notwendig ist. Damit soll die Sicherheit der Ausführung gewährleistet werden – zur Absicherung der Ausführung. Es führt aber häufig auch dazu, dass der Bewegungsablauf etwas grob und eckig aussieht. Im Laufe der Verbesserung (durch Wiederholung ≈ Übung) wird die Anzahl der aktivierten Motorischen Einheiten immer mehr reduziert (das bedeutet, dass immer weniger Nervenzellen aus der Menge – dem pool – der Motoneurone, die für den betreffenden Muskel zuständig sind, ein Aktionspotential aussenden) bis ein Optimum erreicht ist, mit dem das Bewegungsziel noch sicher erreicht wird. Der Bewegungsablauf sieht in diesem Stadium leicht und geschmeidig aus. Auch die Dauer der Muskelkontraktion wird im Laufe des Lern-(Übungs-)Prozesses für den betreffenden Bewegungsablauf angepasst. Diese Vorgänge sind Prozesse der Ökonomisierung.
Die Auswahl der aktivierten Motorischen Einheiten erfolgt durch Neurone auf einer höheren Ebene, die die Motoneurone aktivieren. Diese Auswahl kann durch Lernprozesse einer neuen Aufgabe (Zielsetzung) angepasst werden. Aber auch die Motoneurone selbst können durch höhere Einflüsse so auf neue Aufgaben eingestellt werden, dass sie mehr oder weniger Transmitter (hier das Acetylcholin) ausschütten. Das führt zu einer Veränderung der Kontraktionsstärke. Das bedeutet allerdings nicht, dass die Kontraktionsstärke nicht bei jeder einzelnen Bewegungsausführung an die aktuellen Notwendigkeiten (Regelung jedes einzelnen Ablaufs) angepasst wird.
Da auch die Bewältigung größerer Lasten eine Anpassung (also ein Lernen) darstellt, gehört auch die Vermehrung von Mitochondrien sowie die Verdickung von Muskelzellen, die durch Ausdauertraining und Krafttraining erfolgt, zu den Veränderungen des Muskels durch Lernen.
Aufbau
Das Nervensystem besteht aus dem zentralen Nervensystem: Großhirn, Kleinhirn, Hirnstamm und Rückenmark und dem peripheren Nervensystem (Nervenleitungen zu den Erfolgsorganen). Durch das Lernen von Bewegungen kommt es zu Veränderungen in allen Teilen dieses Systems.
Der zentrale Baustein des Nervensystems ist die Nervenzelle. Die Form der Nervenzellen kann, je nach ihrer Aufgabe, sehr unterschiedlich sein, ihr Aufbau ist aber immer gleich. Sie besteht aus dem Zellkern (Nucleus), der im Körper (Soma) der Nervenzelle liegt. Dieser ist mit dem Zellplasma gefüllt und wird von einer Membran umgeben. Im Plasma befinden sich zahlreiche Organellen, die zum Teil zum Transport von Substanzen, zum Beispiel Proteinen, zwischen Kern und Membran notwendig sind, der Erneuerung der Nervenzelle oder zur Unterstützung der Informationsverarbeitung dienen.
Die einzelnen Nervenzellen tauschen über ihre Fortsätze (Dendriten und Axone) Informationen aus. Jede Nervenzelle verfügt über eine große Anzahl von Dendriten, aber nur jeweils über ein Axon. Beide Arten der Fortsätze sind an ihren Enden sehr stark verzweigt. Die Axone enden an den Dendriten, am Körper anderer Nervenzellen oder an Axonen, meist anderer Nervenzellen. An den Endpunkten haben sie spezifische Verdickungen, die Synapsen.
Im Rückenmark befinden sich Motoneurone und Interneurone. Die Zielgebiete der Axone der Motoneurone sind keine anderen Nervenzellen, sondern die Fasern der Muskeln. Die Axone der Interneurone enden an Motoneuronen oder anderen Interneuronen und können auf diese Weise dazu dienen, Netzwerke zu bilden.
Funktion
Die Aufgabe der Nervenzellen ist es, Informationen aufzunehmen, diese zu verarbeiten und die Ergebnisse weiterzuleiten. Ihre Informationen erhalten sie über die Synapsen der Dendriten. Das Ergebnis der Informationsverarbeitung geben sie über das Axon weiter. Der Träger der Information ist das Aktionspotential. Da ein Aktionspotential immer die gleiche Form hat, liegt die Information in der Frequenz des Aktionspotentials. Diese kann zum Beispiel gleichmäßig sein oder in spezifischen Gruppierungen auftreten.
Die Informationsverarbeitung erfolgt an der Plasmamembran. Nachdem an einer Synapse durch den Transmitter der aussendenden Nervenzelle an der Empfängerzelle (postsynaptische Membran) spezifische Rezeptoren dafür sorgen, dass Ionenkanäle geöffnet werden, verändert der Einstrom dieser Ionen das Transmembranpotential. Das gilt für ionotrope Rezeptoren, die für schnelle Informationsverarbeitung sorgen, wie sie für Bewegungen notwendig sind.
Neuronale Steuerung
Bevor ein Bewegungsablauf zur Ausführung kommt, waren bereits viele Teile des Gehirns an der Vorbereitung und Planung einer korrekten Ausführung beteiligt. Es gibt so genannte Bereitschaftspotentiale, die jeder Aktion vorausgehen und eine allgemeine Disposition und Aktivierung des Organismus herstellen. Soll ein bestimmtes Ziel mit der Bewegung erreicht werden, muss ein Plan erstellt und die Einzelteile aus bekannten Bewegungsabläufen zusammengestellt werden. Das geschieht in verschiedenen Teilen des Großhirns. Dabei werden ständig die erfolgten Teilschritte mit dem Ziel verglichen und an die Bedingungen der augenblicklichen Situation (z. B. körperliche und emotionale Verfassung) des Organismus angepasst. Insofern ist kaum ein Ablauf vollkommen identisch mit einem anderen, obwohl das häufig so aussieht.
Der „Ausgabebereich“ einer Bewegung beginnt im motorischen Cortex. Von dort werden einerseits die Motorischen Nervenzellen (Motoneurone) im Rückenmark (direkt) innerviert, zum anderen werden Impulse zu Kontrollzentren, wie z. B. dem Kleinhirn gesendet, die die korrekte Ausführung – in die nicht bewusst eingegriffen werden kann – überwachen. Für das Lernen neuer oder die Modifizierung von bereits gekonnter Bewegungen ist es deswegen wichtig, dass auch diese Kontrollstrukturen den neuen Anforderungen angepasst werden. Das geschieht über Rückkoppelungen und folgende Modifikationen in Nervenzellen.
Nervenzellen im Zentralen Nervensystem (siehe: Zentralnervensystem) sind für die Innervation der Motoneurone im Rückenmark zuständig. Wenn also mehr oder weniger Motoneurone im Rückenmark für die Ausführung benötigt werden, geschieht auf höherer Ebene die „Entscheidung“ dazu. Das bedeutet in den Nervenzellen der höheren Ebenen kommt es zu Veränderungen (Modifikationen), die für das Lernen notwendig sind. Sollen Funktionen des Organismus auf Dauer verändert werden, wie das beim Lernen der Fall ist, müssen diese Veränderungen bis hin zur Modifikation der DNA und der Gene erfolgen. Solche Modifikationen erfolgen in den Zellkernen. Für das Bewegungslernen müssen diese Veränderungen in den Nervenzellen, die für den Ablauf und die Kontrolle von Bewegungen zuständig sind, erfolgen. Wenn diese Änderung nicht zufällig sein soll, sondern gezielt und wiederholt, muss eine entsprechende Information dazu in den Zellkern gelangen. Für die Signalverarbeitung innerhalb der Nervenzelle – von der Plasmamembran zu den ausführenden Strukturen – existieren verschiedene Wege. Alle werden dadurch eingeleitet, dass von der extrazellulären Seite spezifische Ionen, durch spezifische Kanäle (Ionenkanäle) in die Zelle einfließen. Dazu müssen sich diese Kanäle öffnen. Das ist wiederum abhängig von deren Rezeptoren, die an der Außenseite der Zellmembran auf spezifische Substanzen, entweder ein von einem anderen Neuron ausgeschütteten Transmitter oder einem Hormon, reagieren. Es werden verschiedene Rezeptortypen unterschieden – die ionotropen und die metabotropen.[2] Die ionotropen reagieren in der Regel auf Transmitter. Die Kanäle öffnen sich dann sehr schnell – es fließen z. B. Natriumionen in die Zelle, die das Membranpotential verändern und dadurch bei ausreichender Menge zu einem Aktionspotential führen, durch das das Signal weitergeleitet wird. Diese Kanäle sorgen also für eine direkte schnelle Weiterleitung von Signalen, wie sie für die schnelle Ausführung von Handlungen wie z. B. Bewegungen notwendig sind. Für Vorgänge, die nicht akut so schnell ablaufen müssen, wie das Lernen, sind die Rezeptoren vielfältiger und die Mechanismen zur Öffnung der Kanäle komplexer und sie erfolgen meist über so genannte G-Proteine. Sie reagieren z. B. auf Hormone (metabotrop). Anschließend erfolgt die Informationsverarbeitung innerhalb der Nervenzelle.[3] Dabei spielen so genannte second messenger eine wichtige Rolle. Auf diesem Weg können notwendige Informationen an alle Bestandteile der Nervenzelle gelangen, auch in den Nervenzellkern. Dort können sie dann, wenn sie sich in ähnlicher Weise häufig wiederholen, Veränderungen (Modifikationen) z. B. bis hin zur DNA- und der Genmodifikation bewirken.
Veränderungen durch Lernen
Veränderungen der Funktion der Nervenzellen durch Lernen treten in erster Linie an zwei Stellen auf: an den Synapsen und im Zellkern. Sollen diese Änderungen nicht zufällig, sondern dauerhaft sein, wie das beim Lernen der Fall ist, muss eine entsprechende Information dazu in den Zellkern gelangen, und die DNA muss entsprechend modifiziert werden.
Veränderungen lassen sich auch im motorischen Kortex (MI = primärer motorischer Kortex – dort befinden sich die Nervenzellen, von denen die Muskeln direkt innerviert werden können) zeigen. Sobald Bewegungen wiederholt ausgeführt werden, vergrößern sich dort die Areale der Neurone, die diese Bewegungen auslösen können (Neural plasticity). Das geschieht bereits nach nur wenigen Wiederholungen.[4] Dabei lassen sich 2 Stufen unterscheiden. Eine frühe Lernstufe, bei der es zu einer sehr schnellen Verbesserung der Bewegungsausführung bereits in einer einzigen Übungseinheit kommt. Es folgt eine spätere langsamere Lernstufe, bei der ein weiterer, aber langsamerer Lernfortschritt über mehrere Übungseinheiten, die durchaus einige Wochen dauern können, beobachtet werden kann.
Forschungsgeschichte
Das Interesse daran, wie eine neue Bewegung gelernt werden kann, ist sehr alt, weil vor allem in den Zeiten, in denen noch keine oder nur wenige Maschinen den Menschen die körperliche Arbeit abnahmen, diese Arbeiten von den Menschen und das möglichst gut geleistet werden mussten. Bei den Hinweisen zu deren Ausführung ging es meist um Anweisungen und Beschreibungen aus der sichtbaren Veränderung der Bewegungen des Lernenden. In den Künsten hatten sich seit alters her Techniken zur Vermittlung durch Erfahrung und Überlieferung stetig optimiert. Auch die allgemeinen Theorien über das Lernen beruhten auf Erfahrungen von Lehrenden und deren Aufzeichnungen. Daraus ergaben sich die methodischen Hinweise. In der wissenschaftlichen Forschung verlaufen die Theorien des Lernens – auch des Bewegungslernens – parallel und etwa zeitgleich mit der Erforschung des Nervensystems und seines Aufbaus und seiner Funktionen.
Reflexe und Reflexketten
Erst seit Beginn des 20. Jahrhunderts lässt sich eine systematische Erforschung der Lernvorgänge von Bewegungen beobachten. Sie wurden jedoch lange nicht in die allgemeinen Lerntheorien integriert, weil sie als nicht ausreichend angesehen wurden (siehe Behaviorismus), um vor allem kognitives Lernen zu beschreiben und zu erklären. Dabei bot die Erforschung des Nervensystems seit ihrem Beginn ausreichende Hinweise für das Zustandekommen von Bewegungen, da die physiologischen Ursachen den Untersuchungen leichter zugänglich sind. Sie wurden lange Zeit nur durch Untersuchungen von Bewegungen von Tieren gewonnen.
Die herausragenden Entdecker und Erforscher dieses Nervensystems waren Charles Scott Sherrington (1857–1952) und Iwan Petrowitsch Pawlow (1849–1936). Sherrington zum Beispiel entdeckte, dass das Nervensystem aus einzelnen Nervenzellen besteht, die physiologisch voneinander getrennt sind, aber an bestimmten Stellen Signale untereinander austauschen können und dieser Austausch von Signalen die Ursache der Bewegungen ist. Beide beschäftigten sich mit den Reflexen. Während aber Sherrington hauptsächlich die Anatomie der einzelnen Nervenzellen und deren Funktion untersuchte, entwickelte Pawlow wichtige Modelle über die Reflexe Reiz-Reaktions-Theorie (siehe Reiz-Reaktions-Modell). Pawlow erarbeitete unter anderem die Theorie des bedingten Reflexes (siehe Hunde-Experiment von Pawlow), die auch zur Grundlage des Behaviorismus wurde.
Beide kamen zu dem Schluss, dass die Bewegung eines Lebewesens durch eine Kette von Reflexen entsteht. Dabei dient die Kontraktion eine Muskels als Reaktion auf einen Reiz. Diese Kontraktion dient wiederum als Reiz (Stimulus) für die Kontraktion des nächsten Muskels und so weiter. Ein solcher Ablauf wurde dann als Reflexkette (siehe Reflexkettentheorie) bezeichnet. Die Vorstellung von diesen Reflexketten war es, dass sie in sehr stereotyper Weise ablaufen, ähnlich einem Münzautomat. Diese Vorstellung wurde dadurch unterstützt, dass die beobachteten Bewegungen von Tieren und Menschen, die meist vollkommen beherrschte Ausführungen – wie Alltagsbewegungen – waren, immer gleichförmig aussahen.
Entdeckung der Regelkreise in lebenden Organismen
Bei der genaueren Untersuchung des Verhaltens und der Bewegung von Tieren, die z. B. in Seewiesen (siehe Konrad Lorenz) etwa von 1930 an durchgeführt wurden, stellte man fest, dass die Bewegungen der Tiere Störungen ihres Bewegungsablaufs zum Beispiel durch Hindernisse ohne Schwierigkeiten ausgleichen konnten – die Bewegungsabläufe also nicht vollkommen starr sind.
In besonderer Weise hat sich Erich von Holst (1908–1962) mit der Frage beschäftigt, auf welche Weise Tiere solche Störungen durch Modifikationen ihrer Bewegungsabläufe ausgleichen können. Im Zuge dieser Untersuchungen entwickelte er das Reafferenzprinzip,[5] das diese Abläufe erklärt. Es dauerte aber noch einige Jahre, bis die Erkenntnis auch die Motorikforschung erreichte, dass das Reafferenzprinzip ein wichtiger Baustein zum Verständnis der Bewegung von lebenden Organismen und des Lernens von Bewegungen ist.
Im Prinzip handelt es sich bei dem Reafferenzprinzip um einen verschachtelten Regelkreis. Es passte damit in die Entdeckungen der Zeit, in der Biologen und Ingenieure herausfanden, dass der Gegenstand ihrer jeweiligen Forschung viele Gemeinsamkeiten hat, nur auf jeweils anderer Materie stattfand und meistens anders beschrieben wurde. Es hat sich seit der Zeit durchgesetzt, dass auch lebende Systeme mit technischen Mitteln beschrieben werden.
Aus dieser Zusammenarbeit entwickelt sich auch die Vorstellung und die Beschreibung menschlicher Handlungen als Prozess der Informationsverarbeitung.[6] Da hierbei der gesamte Handlungs- und Lernprozess in unterschiedliche überschaubare Teilprozesse gegliedert wird, wurde erkennbar, an welchen Stellen im Lern- bzw. Lehrprozess, Eingriffe in den Lernprozess sinnvoll sind und wie sie gestaltet sein müssen, um wirksam sein zu können. Diese Erkenntnisse flossen auch in die Motorikforschung ein.[7][8]
Als in den 1980er Jahren das Reafferenzprinzip als ein wichtiger Baustein zum Verständnis von Bewegung und Bewegungslernen erkannt wurde, setzte ein neues, verstärktes Interesse an der Erforschung der Bewegung und seiner Kontrolle – bei der Bewegungsausführung sowie beim Lernen neuer Bewegungen – ein.
Bewegungslernen und Neurowissenschaften
Einen großen Fortschritt brachten die neuen Erkenntnisse aus den Neurowissenschaften, die auch durch die technischen Fortschritte bei der Untersuchung der Hirnstrukturen möglich geworden waren.
Eine Richtung der Forschung beschäftigt sich damit, festzustellen, in welchen Arealen des Gehirns bei den Tätigkeiten des Menschen die einzelnen Aktionen stattfinden – den Fluss der Informationen im Hirn zu verfolgen. Es wurden Modelle aufgestellt die diesen Fluss nachbilden und auf diese Weise das Verständnis vertiefen können[9] in einer anderen Richtung untersucht man die Aktivitäten der Nervenzellen und versucht in diese Zellen hineinzuschauen, um herauszufinden, auf welche Weise sie ihre Leistungen erbringen und Einfluss auf die Modifikationen von Bewegungsabläufen haben können.[10]
Durch die Entwicklung neuer technischer Verfahren (EEG, PET), mit deren Hilfe sich Vorgänge im Gehirn noch genauer darstellen lassen, war es möglich, die Aktivierung von Hirnbereichen bei der Ausführung und beim Lernen von Bewegungsabläufen zu beobachten. Dadurch ließen sich die Vorstellungen überprüfen. Man stellte zum Beispiel fest, dass sehr viel mehr kortikale Areale am Zustandekommen einer Bewegung beteiligt sind als nur die motorischen Kortices und einige subkortikale Regionen wie Basalganglien und Kleinhirn.
Mit der funktionellen Magnetresonanztomographie (fMRT) lassen sich die Aktivitätsflüsse im Gehirn noch genauer beobachten, so dass es heute möglich ist, die Stationen von Regelkreisen bei der Ausführung von Bewegungen zu lokalisieren und zu verfolgen, wie sie beim Erlernen von Bewegungen aufgebaut werden. Man hofft, dass sich durch diese Untersuchungen Maßnahmen entwickeln lassen, mit denen man Lernprozesse beeinflussen kann.[11] Das Problem dabei ist allerdings, dass in dem Tomografen keine großräumigen Bewegungen durchgeführt werden können. Auch stellte man fest, dass bei Lernprozessen Veränderungen auf Ebene der Synapsen beobachtet werden können[12], was aktuell intensiv erforscht wird.
Bewegungslernen und Robotik
Für den Einsatz von Robotern in der Industrie (Konstruktion von Industriegütern), in Medizin, Landwirtschaft und im Militärbereich aber auch in den neueren Anwendungen zur Hilfe in vielen Lebensbereichen hat es sich als wichtig erwiesen, dass Roboter nicht nur in einer vorgeschriebenen notwendigen Weise ihnen einprogrammierte Funktionen erfüllen können. Es ist vielmehr auch notwendig, dass sie ihre Fähigkeiten selbst weiterentwickeln an neue Situationen anpassen (adaptieren) – also lernen – können.
Bereits für die Kontrolle der Bewegungen von Robotern hatten sich Ingenieure intensiv mit Bewegungen von Lebewesen auseinandergesetzt und die Erkenntnisse auch für die Neurowissenschaftler vorangebracht.[13] Vom Studium der Kontrollvorgänge zur Untersuchung der Lernvorgänge war es ein logischer Schritt, denn die Kontrolle von Bewegungen muss gelernt werden, führt aber auch darüber hinaus.
Da sich die Kontrolle und damit das Lernen an komplexen Bewegungen aber nur sehr schwer untersuchen lässt, suchte man dafür einfache Bewegungen, bei denen nur wenige (1 oder 2) Gelenke und nur eine begrenzte Anzahl von Muskeln beteiligt sind. Dies ist zum Beispiel die Beugung eines Unterarms der Fall, wenn man sie isoliert betrachtet. Diese Bewegung ist vielfältig und unter sehr unterschiedlichen Bedingungen der Störung des Ablaufs (Kontrolle heißt auch, eine Störung auszugleichen) untersucht worden.[14]
Als derartige Teilbewegungen lassen sich auch Augenbewegungen zu bestimmten vorgegebenen Zielen hin betrachten – das wurde zum Beispiel an Affen untersucht.[15] Diese können dann als Bewegungsmodule betrachtet werden, die dann auch als „primitives“ bezeichnet werden. (Primitives in der Softwaretechnik sind kleinste Elemente eines größeren (komplexen) Computerprogramms).
An diesen „primitives“ lässt sich sehr präzise untersuchen, wie bestimmte – mit neuen Aufgaben versehene – Bewegungen gelernt werden können. Dabei gefundene Lerngesetze stimmen mit denen überein, die auch von den Behavioristen beschrieben wurden – man nannte sie allerdings anders. Zum Beispiel: Eine neue Verhaltensweise wird durch „rewards“ (Belohnung) gelernt (= Adaptation, Behaviorismus: Acquisition, Erwerb). Wird sie nicht mehr belohnt, kommt es zum „washout“ (Behaviorismus: extinction, Löschung), wird sie dann nach einer Weile „wieder erlernt“ erfolgt das Lernen durch „savings“ schneller (Behaviorismus: „spontaneous recovery“ – spontane Erholung). Bei diesen Forschern heute werden aber im Unterschied zu den Behavioristen, die Vorgänge bei den Lernprozessen im Innern des Organismus ebenfalls untersucht. Man fand, dass sich diese bezüglich der Lernvorgänge hauptsächlich im Kleinhirn (Cerebellum) abspielen.
Die Forscher dieser neuen Studien machen aber keine Aussagen darüber, wie später nach dem Erlernen der einzelnen „primitives“ (Teilbewegungen), diese zu einem „flüssigen, ganzen Bewegungsablauf“ zusammengesetzt werden können. Dies ist aber immer das Ziel des Bewegungslernens beim Menschen (Die Bewegungen sind als Einzelübungen alle bekannt. Die Schwierigkeit liegt lediglich in ihrer Koppelung.[16])
Bewegungslernen in den Lerntheorien
Es existieren verschiedene Modelle, die das Bewegungslernen erklären sollen. Diese sind meist eng verbunden mit einer allgemeinen Lerntheorie. Sie werden verändert, wenn neue Erkenntnisse oder Ansichten über das (Bewegungs-)Lernen dies notwendig erscheinen lassen.
Die Theorie des Bewegungslernens dient, anders als die allgemeine Lerntheorie meist dazu, eine Vorgehensweise, die häufig auf Überlieferung und/oder Erfahrung beruht und umstritten ist, experimentell zu prüfen und zu bestätigen oder zu widerlegen.
Die Theorien für das Bewegungslernen wurden und werden im Wesentlichen bestimmt durch die in den USA vorherrschenden Theorien – es gibt auch eine Lerntheorie für das Bewegungslernen aus dem sowjetischen Wissenschaftsbereich, die jedoch in den westlichen Staaten nur lückenhaft rezipiert wird. Während die Forschung in der DDR stark auf diesen Forschungsstrang Bezug nahm, verhinderte die Auflösung zahlreicher Institute im Zuge der deutschen Wiedervereinigung eine Übernahme dieser Konzepte in die bundesdeutsche Forschung.
Von der Gestalt- oder Ganzheitstheorie, (≈ Kognitivismus in den USA), der ersten wissenschaftlichen Lerntheorie, wird Bewegungslernen nicht behandelt, weil Bewegungslernen nichts Kognitives sei. In der Mitte des 20. Jahrhunderts brachte es jedoch ein Ableger des Kognitivismus, in Form des Österreichischen Schulturnens zu einer zumindest begrenzten Bedeutung auch in Deutschland (Es wurde von der Sporthochschule Köln in die Didaktik des Sportunterrichts eingegliedert und gelangte in die ersten Nachkriegsrichtlinien des Sportunterrichts für das Land Nordrhein-Westfalen). Karl Gaulhofer (1885–1941) und Margarete Streicher (1891–1985)[17] hatten dem physiologischen Bewegungsaufbau von Pehr Henrik Ling das natürliche Turnen entgegengestellt, bei dem immer von der Ganzheit eines Bewegungsablaufs ausgegangen wird, ein Lernender also zunächst den gesamten Ablauf – häufig vereinfacht – ausführt, damit er das Bewegungsziel erreicht – also den ganzen Ablauf versteht. Danach können einzelne Teile der Bewegung separat geübt werden. Dieses Konzept war hauptsächlich für das Grundschulturnen gedacht und hat dort seine Berechtigung.
Behaviorismus
Die bis heute einflussreichste Lerntheorie ist der Behaviorismus (behavior, auch: behaviour = Verhalten). Sie entstand zu Beginn des 20. Jahrhunderts in den USA und geht auf Vorstellungen von John Broadus Watson[18] zurück, obwohl meist Edward Lee Thorndike (1874–1949) oder Burrhus Frederic Skinner (1904–1990) als ihre Begründer gelten. Watson ging davon aus, dass sich Lernen wissenschaftlich verantwortungsvoll nur dadurch analysieren lässt, dass das, was objektiv beobachtbar ist, nämlich das Verhalten des Lernenden, präzise beschrieben und ausgewertet wird. Introspektion – das Mittel der kognitiven Lerntheorie – wurde von ihm als unwissenschaftlich abgelehnt.
Die Behavioristen waren die ersten, die die Vorgänge des Lernens systematisch untersucht und beschrieben haben. Es wird ihnen häufig vorgeworfen, dass sie ihre Untersuchungen hauptsächlich an Tieren durchgeführt haben (zum Beispiel Tauben und Katzen), und sie diese deswegen nicht, wie es geschehen ist, einfach auf Menschen hätten übertragen werden dürfen. Es wurde argumentiert, dass ihre Theorie Bewusstsein und Gefühlszustände übergehe, vernachlässige, dass alles Verhalten während und aufgrund der individuellen Lebensgeschichte erworben werde, Leistungen in den Künsten (zum Beispiel in Musik, Literatur und auch in den exakten Wissenschaften) nicht erklären könne und anderes mehr.[19] Es handelt sich aber bei den beschriebenen Verhaltensweisen um elementare Vorgänge, die bei jedem Lebewesen auslösbar sind. Bei den Tieren mögen es die einzigen sein, Menschen können auf vielfältigere Weise lernen.
Bei den einfachen Lernmechanismen, die die Behavioristen beschrieben haben, handelt es sich im Wesentlichen um die Wirkung von Belohnung (Verstärkung; reward) und Bestrafung (punishment), den Erwerb (acquisition), die Löschung (extinction), die spontane Erholung (spontanious recovery) von Verhaltensweisen, um die wichtigsten zu nennen.
Erkenntnisse für das Bewegungslernen
Das Bewegungslernen gilt im Behaviorismus als mehr oder weniger komplexe Verkettung einfacher S-R-Verbindungen, bei dem ein bereits erlerntes S-R-Element als Ausgangsreiz für eine nachfolgende S-R-Verbindung dient – das kann jeweils auf klassischem oder operantem Konditionieren beruhen.
Eine Tür aufzuschließen besteht z. B. aus den Einzelverbindungen[20]
- Schlüssel in die Hand nehmen (S: Schlüssel sehen – R: Schlüssel in die Hand nehmen)
- Schlüssel richtig fassen (S: Schlüssel in der Hand halten – R. Schlüssel in die richtige Richtung drehen)
- Schlüssel in das Schlüsselloch einführen
- Schlüssel drehen
- Tür öffnen
Alle diese Einzelverbindungen mussten gekonnt sein, damit sie dann, wenn die gesamte Aufgabe angeboten würde, als Ganzes in einem einzigen Prozess (d. h. spontan, ohne zu üben) ausgeführt werden konnte.
Bei komplexeren Abläufen muss das nicht immer gelingen. Zum Beispiel: Mit dem Auto rechts abbiegen, wenn die Ampel auf „Grün“ schaltet.
Kette:
- Grünes Licht sehen → Gang einlegen und/oder Gas geben
- Nach rechts umschauen (wegen Radfahrer)
- Lenkrad drehen → Abbiegevorgang kontrollieren
- Lenkrad zurückdrehen
- Fahrt kontrollieren: (Fahrspur einhalten, Richtungsanzeiger zurückgesetzt?, Tempo)
Alle diese Elemente beherrscht der Fahrschüler. Werden sie zum ersten Mal zusammen gefordert, gelingen sie häufig nicht.
Bedeutend sind die Verstärkerpläne von Burrhus Frederic Skinner.[21] Sie werden z. B. in der Physiotherapie (PT) meist unbewusst angewendet: In der PT Praxis wird durch kontinuierliche Verstärkung, d. h. jede Ausführung wird verstärkt (Richtigkeit bestätigt)der Ablauf am sichersten gefestigt. Später, zu Hause wird nicht mehr jede Ausführung verstärkt, es kommt zu einer Verstärkung in unregelmäßigen Intervallen (intermittent, variable ratio; wobei die Intervalle immer länger werden können). Das führt zu der sichersten Behaltensform.
Skinner entwickelte ebenfalls das Verfahren einer höheren Form des operanten Konditionierens: Das Shaping, bei dem das Zielverhalten systematisch erarbeitet wird. Es findet Anwendung in der Tierdressur. Es wird aus dem jeweiligen Verhaltensrepertoire, das das Tier in der Versuchssituation zeigt, die Verhaltensweise verstärkt, die dem Zielverhalten am nächsten kommt. Dadurch wird das Repertoire des Antwortverhaltens des Tieres verändert und dem Zielverhalten angenähert. Auf diese Weise lassen sich – in manchmal langen Prozessen – sehr präzise und auch artfremde Verhaltensweisen vermitteln – nicht nur bei Tieren.
Vor allem in den Arbeitswissenschaften wurde der Zusammenhang zwischen der räumlichen Reizkonstellation und dem Lernerfolg untersucht. Man stellte fest: Bei ungünstiger Anordnung von Elementen, die zu bedienen sind, stellt sich ein Lernerfolg nur verzögert ein und es kommt auch bei Beherrschen des Ablaufs leicht zu Fehlern, z. B. Anordnung der Schalter für die Platten eines Herdes oder wenn ein Element, das mit der rechten Hand manipuliert werden soll, auf der linken Seite des Körpers angebracht ist (Inkompatibilität von Reiz und Reaktion). Diese Kompatibilitätsregeln spielen eine Rolle für das Design von Gegenständen, vor allem für das Industriedesign zur Optimierung von Arbeitsabläufen.
Kontrolltheorie (Motor Control)
Kontrolle bedeutet ständige Überwachung eines Ablaufs, damit dieser erfolgreich abgeschlossen werden kann (von: control engineering = Regelungstechnik). Die Bedeutung der Kontrolle für das Bewegungslernen wurde durch Zusammenarbeit von Bewegungswissenschaftlern mit Ingenieuren um 1940 erkannt (siehe auch Kybernetik), in Deutschland in der Verhaltensphysiologie,[22] in den USA bei der Ausbildung von Piloten für den Zweiten Weltkrieg.
Es war bereits den Behavioristen bekannt, dass ohne die Bestätigung der Richtigkeit (Verstärkung, reward) einer Bewegung, kein Lernen erfolgen kann. Für Bewegungsabläufe galt für sie, dass beim Aufbau einer Bewegungskette, jedes Kettenglied bei seinem Erwerb verstärkt und somit kontrolliert worden war[23] und der Gesamtablauf dann sozusagen automatisch – ohne Einzelkontrolle – ablaufen kann. Es ergibt sich eine Art Bewegungsprogramms, das zunächst in Abschnitten gelernt wird, dann aber unbewusst abläuft.
Wird eine Bewegung nur von außen betrachtet (wie im Behaviorismus), bereitet – trotz des bereits vorhandenen Wissens um physiologische Rückkoppelungsmechanismen, z. B. die Gamma-Schleife der Muskelkontrolle (siehe: Motorik) – das Verständnis für eine ständige Überwachung von Bewegungsabläufen Schwierigkeiten. Eine Kontrolle schien nicht nützlich, da Korrekturen einer Bewegung nur nach deren Abschluss mit Wirksamkeit für den nächsten Ablauf für möglich gehalten wurden, denn die Zeit für eine Korrektur des Ablaufs dauere zu lange (man ging von ungefähr 200 ms aus), als dass sie während eines Bewegungsablaufs wirksam werden könne. Für langsam ablaufende Bewegungen galt das jedoch nicht. Dort konnte zum Beispiel auch der Ausführende selbst Korrekturen durchführen.
Es wurde deswegen zwischen „open loop“ ablaufenden schnellen Bewegungen, die lediglich nach ihrer Ausführung beurteilt (kontrolliert) und für den nächsten Ablauf korrigiert werden können (beispielsweise der Wurf eines Balles) und „closed loop“ ablaufenden langsamen Bewegungen, bei denen während des Ablaufs Korrekturen möglich sind (beispielsweise das Zeichnen eines Kreises) unterschieden.
Für den langsamen Ablauf einer „closed loop“ Bewegungsausführung bediente man sich zunehmend der ingenieurmäßigen Darstellung durch Fluss- und Strukturdiagramme, die die Informationsverarbeitung während des Ablaufs darstellen sollen. Obwohl diese Strukturdiagramme eine Aufforderung darstellen, über die Vorgänge im Organismus nachzudenken, weil sie Elemente im Innern der „Organismusbox“ enthalten, die mit physiologischen Erkenntnissen zu füllen wären, taten sich die Behavioristen auch hier schwer, ihre Vorstellung von der Black-Box zu revidieren. Bewegungsprogramme boten eine Lösung dieses Problems an.
Bewegungsprogramme
Zu der damaligen Zeit galten aber Programme (z. B. Computerprogramme) als starr (Vorstellung: Automat: Oben wird Münze (= der Bewegungswunsch) eingesteckt. Dann kommt immer exakt derselbe Bewegungsablauf heraus). Dies entsprach der Vorstellung des „unbewusst“ ablaufenden Bewegungsablaufs und konnte auch so beobachtet werden. Das war aber unerwünscht, denn Bewegungsabläufe müssen flexibel sein, d. h. sich an unterschiedliche Bedingungen (im Organismus, z. B. Müdigkeit, oder in der Umgebung, z. B. rauer Untergrund) anpassen können.
Das Aufkommen von verzweigten Programmen in der Informatik schien dieses Problem der Starrheit der Bewegungsprogramme auf einfache Weise zu lösen.
Die Analogie mit den verzweigten Computerprogrammen führte aber zu dem Problem, wie der Organismus solch ein verzweigtes Programm lernen kann, d. h. sich programmiert: wie weiß er, wo die Abzweigstellen platziert werden müssen und nach welchen Kriterien der eine oder andere Weg eingeschlagen werden muss. Auch die Kontrolle eines solchen Programms muss gelernt werden.
Als ein weiteres Problem ergab sich, dass der Mensch im Laufe seines Lebens sehr viele unterschiedliche Bewegungsprogramme lernen und behalten muss. Diese müssen in einem Langzeitgedächtnis gespeichert werden. Eine gewisse Lösung dieses Problems ließ sich durch die so genannte Schema Theorie erzielen. Diese besagt, dass beim Lernen eines neuen Bewegungsablaufs ebenfalls ein gröberes Schema des Gesamtablaufs ausgebildet wird, so dass die gelernte Bewegung unter ähnlichen Bedingungen nicht völlig neu gelernt, sondern nur angepasst werden und als solche gespeichert werden muss. Beispiel: Wir lernen das Schreiben mit der rechten Hand. Wir können es dann aber auch nicht nur an der Tafel korrekt ausführen, sondern sogar mit der linken Hand oder mit dem Fuß einigermaßen leserlich, obwohl wir dazu ganz andere Muskeln aktivieren müssen, als die, mit denen wir das Schreiben gelernt haben. Intensiv mit dieser Schema-Theorie hat sich Richard A. Schmidt[24] auseinandergesetzt.
Einen wichtigen Beitrag zum Verständnis der Kontrolle für menschliches Handeln lieferten die Psychologen G. Miller, E. Galanter K. Pribram[25] mit ihrem TOTE (Test-Operate-Test-Exit) Modell.
Einen Ansatz, auch die physiologischen Grundlagen der Bewegungskontrolle zu erklären und zu verstehen, bietet Erich v. Holst mit dem Reafferenzprinzip.[26]
Theorie der Informationsverarbeitung
Von Psychologen war die Kontrolltheorie in den 1960er und 1970er Jahren zur Theorie der Informationsverarbeitung im menschlichen Organismus weiterentwickelt worden. Diese half einige Probleme aus der Kontrolltheorie zu lösen.
In der Theorie der Informationsverarbeitung wird wie in der Kontrolltheorie das menschliche Handeln als Strukturdiagramm dargestellt, die einzelnen Elemente aber mit den entsprechenden physiologischen und neurophysiologischen Inhalten gefüllt. Diese geben Auskunft darüber, in welcher Weise die Informationen, die aus der Umwelt auf den Organismus treffen, verarbeitet werden bevor das Ergebnis auf die Umgebung einwirkt. Die Psychologen beschränken sich dabei hauptsächlich auf Wahrnehmung und Gedächtnisses.[27]
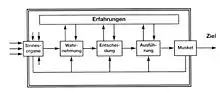
Für die Bewegungswissenschaftler ist aber besonders die Ausführung der Handlung/Bewegung mit der Vorbereitung und Kontrolle der Muskeltätigkeit von Bedeutung. Der Kanadier Ronald G. Marteniuk[28] hat 1976 auf Vorarbeiten von H.T.A. Whiting[29] diesen theoretischen Ansatz für das Bewegungslernen umgesetzt. Dieser wurde von Heidrun Schewe in Deutschland bekannt gemacht.
Das Modell
Es ergibt sich eine Struktur mit fünf Hauptelementen:
- Über die Sinnesorgane werden die Signale (Informationen) aus der Umwelt in das Nervensystem aufgenommen. Das ist ein rein biologischer Vorgang.
- Im nächsten Schritt – Beginn der kognitiven Verarbeitung – werden die für die spezifische Bewegung wichtigen Informationen herausgefiltert und so gruppiert, dass die Situation, in der sich der Organismus befindet, identifiziert und ihre Bedeutung für die augenblickliche Situation bzw. für eine geplante Handlung erkannt wird. Am Ende dieses Schrittes, den man als Wahrnehmungsbereich (Wahrnehmung, perception mechanism) bezeichnet, steht die Klassifizierung der Gesamtsituation.
- Kann die Situation eindeutig einer bestimmten bereits bekannten Klasse von Situationen zugeordnet werden, und es liegt nur eine einzige Handlungsalternative vor, kann diese abgerufen und ausgeführt werden. Ist eine eindeutige Klassifizierung nicht möglich oder existieren mehrere Alternativen (wie z. B. in einem Sportspiel, oder für eine Autofahrt verschiedene Fahrtrouten), muss eine adäquate oder erfolgreiche Lösungsstrategie ausgewählt oder gesucht werden. Diese muss für eine Entscheidung (decision) dafür (geistig) durchgespielt und auf ihre Konsequenzen hin überprüft werden. Dies kostet Zeit. Ist diese nicht vorhanden oder der Ausführende ungeduldig, kann es zu Fehlentscheidungen kommen, die zum Scheitern der Bewegung führen. Ist eine Lösungsstrategie gefunden, deren Konsequenzen akzeptiert werden können, wird diese Bewegungsabsicht an den „Ausführungsbereich“ übergeben.
- Im Ausführungsbereich (effector mechanism) werden die Ablaufmuster der Bewegung zusammengestellt. Ist die beabsichtigte Bewegung schon öfter ausgeführt worden – das ist meistens der Fall, da wir vom Kindesalter an Bewegungserfahrungen sammeln, und für alle unsere Muskeln geeignete Innervierungsmuster vorliegen, so dass im Erwachsenenalter völlig neue Bewegungen kaum mehr vorkommen –, müssen die dazu notwendigen Bewegungsmuster lediglich bereitgestellt und aktiviert werden. Ist die beabsichtigte Bewegung neu oder relativ neu, müssen für sie geeignete Muster (Teilmuster) gesammelt und miteinander koordiniert werden.
- Der letzte Teilabschnitt, das „Muskelsystem“, ist für die Umsetzung der kognitiven Ergebnisse in den mechanischen Bewegungsablauf zuständig. Dieser läuft unter ständiger Kontrolle ab.
In jedem Verarbeitungsschritt ist ein Rückgriff auf alle zurückliegenden Verarbeitungsschritte möglich. Sie werden ständig mit der Umgebungssituation und dem beabsichtigten Ziel abgeglichen.
Den Abschluss des Gesamtablaufs bildet die Feststellung, ob das beabsichtigte Ziel erreicht wurde oder nicht. Entsprechend wird der Ablauf als positiv oder negativ gespeichert (knowledge of result) = Bewertung. Dieses Modell lässt auch eine Detailanalyse des Gesamtablaufs sowohl für den Lehrenden als auch für den Lernenden zu (knowledge of performance) zu. Die Bedeutung dieser Rückkopplungsmechanismen wird eingehend von A. Gentile[30] diskutiert, ebenfalls von H. Schewe.[31]
Die Vorteile dieser Theorie für Ausführende und Lehrende besteht in der Möglichkeit, die Entwicklung und Durchführung eines Bewegungsablaufs in ihren einzelnen Schritten verfolgen und analysieren zu können. Das ist für Planung, Analyse, Beurteilung und Korrektur, besonders beim Lernen der Bewegung, wichtig. Durch Einbeziehung zusätzlicher psychologischer Erkenntnisse (z. B. Aufmerksamkeit für die Wahrnehmung) lassen sich Leistungsunterschiede bei der Ausführung erklären und Einfluss auf sie nehmen.
Das Problem aus der Kontrolltheorie, wie neue Programme und deren Kontrolle gelernt werden können, wird durch den Rückgriff auf die Bewegungserfahrungen des Einzelnen gelöst.
Die Theorie der Informationsverarbeitung stellt das heute gültige Wissen über das Zustandekommen der Verhaltensweisen von lebenden Systemen, also auch des Menschen, dar. Diese Grundlage hat zu weiteren Fragestellungen geführt. Die erste und einfache Frage zu diesem Modell ist die danach, in welchen Strukturen des Organismus diese Verarbeitungsschritte erfolgen. Dafür ist selbstverständlich das Nervensystem als das Informationssystem verantwortlich, aber für die einzelnen Teilvorgänge mussten die Orte gesucht und vor allem die Übergangsstrukturen untersucht werden. Das ist heute zu einem großen Teil geschehen. Es sind aufgrund der aktuellen Untersuchungsmethoden, von vielen der beteiligten Vorgänge bekannt, wo sie erfolgen.
Eine weitere Fragestellung ist die nach dem Zeitbedarf für die Durchführung der einzelnen Verarbeitungsschritte. Das führt zu 2 weiteren Problemen. Zum einen der Frage danach, wie viel Zeit man benötigt, um eine Bewegung vorzubereiten – wie verhält sich das bei spontanen Bewegungen? Die zweite, besonders wichtige ist: Kann unter diesen Bedingungen eine Bewegung während ihres Ablaufs korrigiert werden? Das ist die Frage nach der Kontrolle der Bewegung (open loop – closed loop). In dem grafischen Modell sind bei jedem der Bearbeitungsschritte Rückkoppelungsschleifen eingetragen – das bedeutet, man geht von einer internen Kontrolle der einzelnen Schritte aus. Diese Problematik ist gegenwärtig noch immer ein Hauptgegenstand der Bewegungsforschung.[32]
Als eine Art Zwischenergebnis lässt sich mit einer alten Vorstellung über die Ausführung von Bewegungsabläufen beschreiben. Diese besagt, dass die Kontrolle von Bewegungsabläufen eine hierarchische Struktur hat, sie also auf verschiedenen Ebenen erfolgt. Dabei gelten die Reflexe über die kurzen Rückenmarksbögen als die unterste Ebene und die bewusste Kontrolle eines Ablaufs als die oberste Ebene. Dies lässt sich auch gut mit den Vorstellungen des Reafferenzprinzips darstellen und wird heute allgemein akzeptiert.
Mit dieser Darstellung der Hierarchie der Bewegungskontrolle verbindet sich auch die Vorstellung der Bewusstheit der Ausführung von Bewegungsabläufen. Diese Fragestellung ob und unter welche Bedingungen die Ausführung dem Bewusstsein zugänglich ist, und ob sich eine solche erlernen lässt muss Gegenstand philosophischer Betrachtung und Forschung sein. Ergebnisse darüber sind heute eher spekulativer Natur.
Neural Group Selection
Waren die Thesen der bisher beschriebenen Theorien aus experimentellen Untersuchungen des Verhaltens abgeleitet, zieht die Theorie der Neural Group Selection (TNGS) (siehe Evolution) ihre Schlüsse unmittelbar aus den Untersuchungen des Gehirns und dessen Tätigkeit.
Neue Fragen und Zweifel – z. B. wie die Klassen von Handlungen/ Bewegungen, nach denen man sich für eine bestimmte Ausführung entscheidet, entstanden sein könnten, da dazu eine entsprechende Vorstrukturierung vorhanden sein muss; dass Varianten von Bewegungen – zur Anpassung an neue Umgebungsbedingungen – nur in einer Breite von natürlichem Rauschen vorstellbar waren; und dass sich auf neuronaler Ebene (siehe Nervenzelle) nicht erklären ließ, warum große Mengen an neuronalen Verbindungen bei Bewegungen brachzuliegen scheinen – machten neue Überlegungen notwendig.
Lernen von Bewegungen
Nach der Theorie der Neural Group Selection (TNGS) (siehe Evolution)[33] folgt, dass alle Prozesse (mentale und motorische) – jeder Denkprozess, jede Handlung und jede Bewegung – nicht nach erlernten, im Gehirn in repräsentativer Form vorliegenden Programmen, sondern nach dem evolutionären Selektionsprinzip geplant, entwickelt, koordiniert und ausgeführt werden. Es wird also jeder Ablauf völlig neu aufgebaut und zusammengestellt. Dabei werden vorhandene neurale Verbindungen genutzt und gleichzeitig verändert. Das gilt auch für das Bewegungslernen.
Das Prinzip ist, jeweils eine geeignete Auswahl aus einer sehr großen (mehrere Milliarden Elemente) Population (im Organismus: Populationen von Synapsen an Neuronen (siehe Nervenzelle, Neuronennetzwerken und Gruppen von Neuronennetzwerken, zu Gruppierungen, Repertoires und Karten)) zu treffen. Diese jeweiligen Gruppierungen innerhalb eines Repertoires sind ähnlich aber nicht gleich (Degeneracy), so dass auch bei unterschiedlicher Auswahl die gleiche Leistung zustande kommen kann. Durch ständigen rekursiven Austausch zeitgleich aktiver Neuronengruppen in den reziprok verschalteten Regionen, die über den gesamten Cortex (siehe Großhirnrinde) verteilt sind (Reentry), werden die sensorischen und motorischen Ereignisse räumlich und zeitlich koordiniert.
Die jeweils getroffene Auswahl ist bei jeder Ausführung eine andere und sie ist in der Regel keine exakte, sondern eine ausreichend gute Lösung, die durch die aktuelle ständige Überprüfung und Veränderung der Synapsen optimiert wird und auf höchster Könnensstufe nahezu optimal sein kann.
Bei diesem Verfahren können auch völlig neue Abläufe erfolgreich ausgeführt werden. Diese benötigen mehr Zeit für ihre Planung und Vorbereitung (Auswahl und Koordination durch Reentry). Sie sind auch weniger präzise. Aber durch Wiederholung (Üben) wird die Auswahl verbessert, und die Koordination kann sparsamer und schneller ablaufen.
Soll ein neuer Bewegungsablauf gelernt werden, ist es deswegen sinnvoll, von vornherein Variationen der Ausführung vom Lernenden zu fordern, damit mehr „geeignete“ Verbindungen für die Auswahl der jeweiligen optimalen Lösung zur Verfügung stehen.
Gelöste Probleme
Durch diese Theorie ist das Problem des Gedächtnisses als repräsentativer Speicher aller gelernten Bewegungsabläufe gelöst, da jede Bewegung aus den am besten geeigneten Verknüpfungen neu zusammengestellt und optimiert wird.
Es wird ebenfalls die Beobachtung bestätigt und erklärt, dass am Beginn des Lernens von Bewegungen das Bewusstsein in stärkerem Maß beteiligt ist als auf höheren Stufen des Könnens. Der Bewegungsablauf wird im Zuge häufiger Wiederverwendung (Übung) automatisiert, wie man sagt. Es wird dann weniger Aufmerksamkeit d. h. auch Energie für den Ablauf benötigt wie z. B. für Alltagsbewegungen.[34] Das bedeutet aber nicht, dass er ohne Kontrolle abläuft.
Diese Theorie eignet sich ebenfalls, viele der aus der behavioristischen Lernforschung widersprüchlichen Untersuchungsergebnisse zu erklären (z. B. massed oder distributed practice).
Vorstellungshilfe: Auf der letzten Stufe der Ausführung werden Muskeln aktiviert. Ein Muskel besteht aus einer sehr großen Anzahl motorischer Einheiten (siehe Motorik). Diese motorischen Einheiten werden von Motoneuronen im Rückenmark innerviert. Es wird aber für jede Bewegung nur eine begrenzte Zahl von motorischen Einheiten benötigt. Man überlege sich, ob für eine Kontraktion des Muskels, bei der eine vorgegebene definierte Kraft aufgebracht werden soll, immer dieselben motorischen Einheiten aktiviert werden (Programmvorstellung) oder ob es durch Auswahl (siehe Evolution) geeigneter, aber jeweils anderer motorischer Einheiten nicht ebenfalls zu dem geforderten präzisen Output (Leistung) kommt.
Kritischer Rationalismus
Der Kritische Rationalismus von Karl Popper wählt eine philosophische, metaphysische und erkenntnistheoretische Herangehensweise an das Problem des Bewegungslernens, die jedoch die wissenschaftlichen Erkenntnisse kritisch einbezieht: Die evolutionäre Erkenntnistheorie.[35][36][37][38] Der Kritische Rationalismus betont den wesentlichen Zusammenhang zwischen einer Bewegung des Organismus und dem Wissen des Organismus über Bewegungsabläufe und somit die Relevanz der Erkenntnistheorie in dieser Frage. Er verwirft alle behavioristischen wie biologistischen Positionen zum Bewegungslernen als reduktionistisch und szientistisch. Er nimmt an, dass Bewegungen Handlungen sind und damit Problemlösungsversuche, also auf die Verwirklichung eines Zwecks hin abzielen, auch wenn Zwecke als solche metaphysisch sind.
Der Kritische Rationalismus betont eine fundamentale Zweiteilung des Lernvorgangs. Der erste Teil des Bewegungslernens ist demnach das Lernen eines adäquaten Bewegungsablaufs selbst. Denn nur wenige Bewegungsabläufe sind adäquat im Hinblick auf einen gegebenen Zweck, und ein Organismus wird viele Bewegungsversuche und Fehlschläge durchlaufen, bis die Bewegung zum gewünschten Zweck führt, oder der Zweck selbst als unerreichbar aufgegeben wird. Diese Bewegungsversuche wählt er letztendlich mithilfe des Zufalls, jedoch nicht rein zufällig, da er schon Hintergrundwissen über viele Sachverhalte haben wird und somit viele Bewegungsversuche von vornherein schon als nicht zweckdienlich ausschließen kann (z. B. dass, um an die Früchte am Baum zu gelangen, ein sprungartiges Bewegen der Beine vielversprechender ist als das Wackeln der Ohren). Bei niederen Lebewesen sind diese Zwecke sowie das Wissen über Bewegungsabläufe selbst mehr oder weniger fest im genetischen Material eingebaut, und können nur durch Mutation und natürliche Selektion verändert werden. Höhere Lebewesen hingegen haben höhere Kontrollorgane (z. B. Gehirn), durch das sie sich diese Zwecke bewusst setzen und sie im Laufe des Lebens auch ändern können, bis hin zum Menschen, der seine Zwecke sogar in sich selbst bewusster und weitsichtiger Weise wählen kann. Höhere Lebewesen können demnach Fehlschläge eines Bewegungsablaufs bewusst als Falsifikation der Annahme erkennen, er verwirkliche den Zweck. Sie können den Bewegungsablauf dann bewusst abändern und es erneut versuchen.
Hat ein höherer Organismus irgendwann einen adäquaten Bewegungsablauf auf diese Weise gelernt, so folgt die zweite Stufe des Lernprozesses, nämlich die stetige Wiederholung des Bewegungsablaufs. Durch diese Wiederholung wird er eingeübt und verlässt dadurch das Bewusstsein, wird in das Unterbewusstsein eingeprägt und kann dann von dem Organismus nunmehr unbewusst und sozusagen „automatisch“ abgerufen und ausgeführt werden. Ein im Unterbewusstsein befindlicher Bewegungsablauf kann jedoch zu jeder Zeit wieder auf die Ebene des Bewusstseins gelangen, wenn er unter anderen Umständen sich nicht mehr als adäquat zum Erreichen des Zwecks erweisen sollte. Und er kann bei Menschen sogar ohne konkreten Fehlschlag ganz bewusst wieder ins Bewusstsein gerückt und aufs schärfste kritisch geprüft werden, auch mit Voraussicht im Hinblick auf Umstände, die bisher noch gar nicht eingetreten sind. Popper sieht darin das Wesen der menschlichen Rationalität und letztendlich der Wissenschaft. Zudem kann der Mensch Bewegungsabläufe nicht nur auf Adäquatheit für einen unmittelbaren Zweck hin prüfen, sondern gleichzeitig auch für mehrere andere, gegebenenfalls höhere Zwecke, wie die Moral. Er kann so Bewegungen bewusst unterlassen, die zwar adäquat in Hinblick auf einen unmittelbaren Zweck sind (z. B. Vorwärtsbewegung auf einen Brunnen zu, um zu trinken, obwohl ein anderer Mensch im Weg steht), die aber einem höheren moralischen Zweck zuwiderlaufen (dass man keine Mitmenschen anrempeln soll).
Der wesentliche, erste Teil des Lernvorgangs ist damit asymmetrisch vom negativen Element des Fehlschlags beherrscht, d. h. von der Inadäquatheit in Bezug auf einen oder mehrere Zwecke. Bestätigung hingegen spielt überhaupt keine Rolle, selbst beim zweiten Teil des Lernvorgangs nicht, jedenfalls soweit Bestätigung mehr sein soll als die bloße (vorläufige) Abwesenheit des Fehlschlags. Das zeigt sich dadurch, dass auch ein inadäquater, nicht oder nicht mehr als zweckmäßig bestätigter Bewegungsvorgang durch Wiederholung in das Unterbewusstsein verlagert werden kann (Marotten). Nach Popper begehen die meisten Ansätze den Hauptfehler darin, die Zweiteilung des Lernvorgangs zu übersehen und die Wiederholung als das eigentliche und wesentliche Element des Bewegungslernens zu verstehen, und laufen so auf eine irrige induktivistische Konditionierungstheorie hinaus.
Literatur
- Shelly A. Boudreau, Dario Farina, Deborah Falla: The role of motor learning and neuroplasticity in designing rehabilitation approaches for muscukloskeletal pain disorders. In: Manual Therapy, 2010, doi:10.1016/J.math.2010.05.008
- Zoubin Ghahramani, Daniel M. Wolpert: Modular Decomposition in visuomotor Learning. In: Nature. 386, März 1997, S. 392–395.
- Richard A. Schmidt, Timothy D. Lee: Motor Control and Learning. 4. Auflage. Champaign IL 2005.
- Anne Shumway-Cook, Marjorie H. Woollacot: Motor Control. 3. Auflage. Philadelphia 2007, S. 22.
- Richard A. Schmidt: A Schema Theory of discrete Motor Skill Learning. In: Psychological Review. 82 (1975), S. 225–260.
- Ronald G. Marteniuk: Information Processing in Motor Skills. Holt Rinehart and Winston, New York 1976.
- Eric R. Kandel, James H. Schwartz, Thomas M. Jessel: Principles of Neural Science. 4. Auflage. McGraw-Hill Companies, New York 2000, ISBN 0-8385-7701-6.
- Heidrun H. Schewe: Die Bewegung des Menschen. Thieme Verlag, Stuttgart 1988, ISBN 3-13-706901-7.
- Heidrun H. Schewe: Das human-performance-model – ein Erklärungsansatz für das Bewegungslerne. In: Krankengymnastik 31(1979) S. 541–554
- Heidrun H. Schewe: Wege zum Verständnis von Bewegung und Bewegungslernen. In: B. Lipp, W. Schlegel, K. Nielsen, M. Streubelt: Gefangen im eigenen Körper – Lösungswege / Neurorehabilitation. Neckar-Verlag, Villingen-Schwenningen 2000.
- Erich von Holst, Horst Mittelstaedt: Das Reafferenzprinzip. In: Naturwissenschaften. 37 (1950) S. 464–476.
- Peter H. Lindsay, Donald A. Norman: Human Information Processing, an Introduction to Psychology. Academic Press, New York / London 1977.
- Thorsten Kahnt, Marcus Grueschow, Oliver Speck, John Dylan Haynes: Perceptual Learning and Decision Making in Human Medial Frontal Cortex. In: Neuron. 70, 2011, S. 549–559.
- Christian Rosenmund, Carsten Stevens: Definition of the Readily Releasable Pool of Vesicles at Hippocampal Synapses. In: Neuron. 16, 1996, S. 1187–1207.
- Burrus Frederic Skinner: Was ist Behaviorismus. Rowohlt Verlag GmbH. Reinbek bei Hamburg. 1978.
- Thorsten Kahnt, Marcus Grueschow, Oliver Speck, John Dylan Haynes: Perceptual Learning and Decision Making in Human Medial Frontal Cortex. In: Neuron. 70, 2011, S. 549–559.
- David W. Franklin, Theodore E. Milner: Adaptive Control of Stiffness to Stabilize Hand Position with Large Loads. in: Experimental Brain Research 152 (2003) S. 211–220.
- Joseph Y. Nashed, Isaac L. Kurtzer and Stephen Scott: Context-dependent inhibition of unloaded muscles during long-latency epoch, in: Journal of Neurophysiology 113 (2015) S. 192–202.
- David W. Franklin Daniel M. Wolpert: Specificity of Reflex Adaptation for Task-Relevant Variability in: The Journal of Neuroscience (Behavioral/Systems/Cognitive) 24 (2008) S. 14165–14175
- Maurice A. Smith, Ali Ghazizadeh, Reza Shadmer: Interacting Adaptive Processes with different Timescales Underlie Short-Term Motor Learning. Plos Biology. Band 4, Juni 2006, Ausgabe 6, e179
- Dieter Kruber: Sport. In der Reihe: Erhard Schmidt, Herausgeber; Standardwerk des Lehrers Ferdinand Kamp Verlag Bochum 1973.
- Friedrich Fetz: Allgemeine Methodik der Leibesübungen. 4. Auflage Limpert Verlag Frankfurt 1971.
- Daniel M.Wolpert, Jörn Diedrichsen, J. Randall Flanagan: Principles of Sensorimotor Learning. In: Neuroscience. 12, 2011, S. 739–751.
- Michael J. Zigmond, Floyd E. Bloom, Story C. Landis, James L. Roberts, Larry R. Squire: Fundamental Neuroscience. Academic Press. San Diego / London 1999.
Einzelnachweise
- Daniel M. Wolpert, Jörn Diedrichsen, J. Randall Flanagan: Principles of Sensorimotor Learning. In: Neuroscience. 12, 2011, S. 747–748.
- Michael J.Zigmond, Floyd E. Bloom, Story C. Landis, James L. Roberts, Larry R. Squire: Fundamental Neuroscience. Academic Press. San Diego / London 1999. Kapitel 9. Neurotransmitter Receptors S. 235–268, und Kapitel 55 Learning and Memory. Basic mechanisms S. 1411–1453
- Michael J.Zigmond, Floyd E. Bloom, Story C. Landis, James L. Roberts, Larry R. Squire: Fundamental Neuroscience. Academic Press. San Diego / London 1999. Kapitel 10 Intracellular Signaling S. 269–315
- Shelly A. Boudreau, Dario Farina, Deborah Falla: The role of motor learning and neuroplasticity in designing rehabilitation approaches for muscukloskeletal pain disorders. In: Manual Therapy, 2010, doi:10.1016/J.math.2010.05.008, S. 2/3
- Erich von Holst, Horst Mittelstaedt: Das Reafferenzprinzip. In: Naturwissenschaften. 37 (1950) S. 464–476.
- Peter H. Lindsay, Donald A. Norman' Human Information Processing, an Introduction to Psychology. Academic Press, New York/ London 1977.
- Ronald G. Marteniuk. Information Processing in Motor Skills. Holt Rinehart and Winston New York 1976.
- Heidrun H. Schewe: Die Bewegung des Menschen. Thieme Verlag, Stuttgart 1988, ISBN 3-13-706901-7.
- siehe zum Beispiel: Zoubin Ghahramani, Daniel M. Wolpert: Modular Decomposition in visuomotor Learning. In: Nature. 386, März 1997, S. 392–395; und: Thorsten Kahnt, Marcus Grueschow, Oliver Speck, John Dylan Haynes: Perceptual Learning and Decision Making in Human Medial Frontal Cortex. In: Neuron. 70, 2011, S. 549–559.
- siehe zum Beispiel: Christian Rosenmund, Carsten Stevens: Definition of the Readily Releasable Pool of Vesicles at Hippocampal Synapses. In: Neuron. 16, 1996, S. 1187–1207.
- zum Beispiel: Daniel M. Wolpert, Jörn Diedrichsen, J. Randall Flanagan; Principles of sensorimotor learning. In: Nature Reviews 2011. S. 730–751.
- Eric R. Kandel, James H. Schwartz, Thomas M. Jessel: Principles of Neural Science. 4. Auflage. McGraw-Hill Companies, New York 2000,
- zum Beispiel: oder David W. Franklin, Theodore E. Milner; Adaptive Control of Stiffness to Stabilize Hand Position with Large Loads in: Experimental Brain Research 152 (2003) S. 211–220; oder: Daniel M. Wolpert, Jörn Diedrichsen and Randall Flanagan; Principles of Sensorimotor Learning in: Nature Reviews (Neuroscience) 12 (2011) S. 739–751
- zum Beispiel: Joseph Y. Nashed, Isaac L. Kurtzer und Stephen Scott; Context-dependent inhibition of unloaded muscles durin long-latency epoch. In: Journal of Neurophysiology 113 (2015) S. 192–202; oder: David W. Franklin and Daniel M. Wolpert; Specificity of Reflex Adaptation for Task-Relevant Variability in: The Journal of Neuroscience (Behavioral/Systems/Cognitive) 24 (2008) S. 14165–14175
- Maurice A. Smith, Ali Ghazizadeh, Reza Shadmer; Interacting Adaptive Processes with different Timescales Underlie Short-Term Motor Learning. Plos Biology. Band 4, Juni 2006, Ausgabe 6, e179
- Dieter Kruber. Sport in der Reihe: Erhard Schmidt Herausgeber; Standardwerk des Lehrers Ferdinand Kamp Verlag Bochum 1973. S. 66 Siehe auch: Frieder Fetz. Allgemeine Methodik der Leibesübungen. 4. Auflage Limpert Verlag Frankfurt 1971 S. 91 ff.
- Natürliches Turnen , Herausgegeben von Hans Groll, 3 Bände. Wien/München. 1930–1942
- John Broadus Watson: Behaviorismus. Deutsche Übersetzung. Köln 1968.
- zum Beispiel: Burrus Frederic Skinner; Was ist Behaviorismus. Rowohlt Verlag GmbH. Reinbek bei Hamburg. 1978. S. 9–10
- "nach: Robert Mills Gagné, Die Bedingungen des menschlichen Lernens. Schroedel Verlag, 1969, S. 103.
- C. Ferster, B. Skinner: Schedules of Reinforcement. New York 1957.
- Erich v. Holst: Zur Verhaltensphysiologie bei Tieren und Menschen. gesammelte Abhandlungen Band 1 und 2. Piper paperback, München 1970.
- siehe auch: Richard A. Schmidt, Timothy D. Lee: Motor Control and Learning. A behavioral Emphasis. 4. Auflage. Human Kinetics Publishers, Champaign IL 2005, S. 164.
- Richard A. Schmidt: A Schema Theory of discrete Motor Skill Learning. In: Psychological Review. 82 (1975), S. 225–260.
- G. A. Miller, E. Galanter, K. H. Pribram: Plans and Struktures of Behavior. London 1960. Deutsch: Strategien des Handelns. Pläne und Strukturen des Verhaltens. Stuttgart 1974.
- Erich. v. Holst, Horst Mittelstaedt: Das Reafferenzprinzip. In: Naturwissenschaften. 37 (1950) S. 464–476.
- Peter H. Lindsay, Donald A. Norman: Human Information Processing, an Introduction to Psychology. Academic Press, New York/ London 1977.
- Ronald G. Marteniuk: Information Processing in Motor Skills. Holt Rinehart and Winston, New York 1976.
- H. T. A. Whiting (Hrsg.): Readings in Human Performance. London 1975.
- A. M. Gentile: A Working Model of Skill Akquisition with Application to Teaching. In: Quest. 17. 1972, S. 42–45.
- Heidrun Schewe: Wege zum Verständnis von Bewegung und Bewegungslernen. In: B. Lipp, W. Schlegel, K. Nielsen, M. Streubelt: Gefangen im eigenen Körper – Lösungswege / Neurorehabilitation. Neckar-Verlag, Villingen-Schwenningen 2000.
- zum Beispiel: F. Crevecoeur, I. Kurtzer, S. H. Scott: Fast corrective responses are evoked by perturbations approaching the natural variability of posture and movement tasks. In: Journal of Neurophysiology. 107 (2012), S. 2821–2832. oder J. Andrew Pruszinski, Isaac Kurtzer, Stephen H. Scott: The long latency reflex is composed of at least two functionally independent processes. In: Journal of Neurophysiology. 106 (2011) oder Joseph Y. Nashed, Isaac L. Kurtzer and Stephen Scott: Context-dependent inhibition of unloaded muscles during long-latency epoch, in: Journal of Neurophysiology 113 (2015) S. 192–202.
- Gerald M. Edelmann: Neural Darwinism. The Theory of Neural Group Selection. Basic Books, New York 1987.
- Gerald M. Edelmann, Giulio Tononi: Gehirn und Geist, wie aus Materie Bewusstsein entsteht. C.H. Beck Verlag, München 2002, S. 68.
- Karl R. Popper, John C. Eccles: Das Ich und sein Gehirn. 1977.
- Karl R. Popper: Objektive Erkenntnis. 1973.
- Karl R. Popper: Wissen und das Leib-Seele-Problem. 1994.
- Karl R. Popper, Konrad Lorenz, Franz Kreuzer: Die Zukunft ist offen. Das Altenberger Gespräch. 1985