Pappaceras
Pappaceras ist eine ausgestorbene Gattung der Unpaarhufer aus der Familie der Indricotheriidae. Sie repräsentiert den bisher ältesten Vertreter der Indricotherien. Der überwiegende Teil des bekannten Fossilmaterials kam in der Inneren Mongolei zu Tage. Gefunden wurden bisher vor allem Schädel- und Gebissreste. Die Fossilreste lagerten in verschiedenen Gesteinsformationen, deren Altersspanne sich vom Übergang des Unteren zum Mittleren Eozän bis zum ausgehenden Mittleren Eozän erstreckt. Die absoluten Altersangaben betragen somit zwischen rund 50 und rund 37 Millionen Jahren. Die Tiere waren relativ klein für Indricotherien und ernährten sich wohl von weicher Pflanzenkost, die mit überwiegend senkrechten Kaubewegungen zerkleinert wurde. Die Gattung erhielt im Jahr 1963 ihre wissenschaftliche Erstbeschreibung, sie galt aber lange Zeit als identisch mit Forstercooperia. Erst weitere Neufunde führten im Jahr 2016 wieder zur Anerkennung von Pappaceras als eigenständige Gattung. Es sind mehrere Arten belegt.
| Pappaceras | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Schädel von Pappaceras, Holotyp von P. meiomenus in verschiedenen Ansichten | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unteres bis Mittleres Eozän (Ypresium bis Lutetium) | ||||||||||||
| 52 bis 37,7 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Pappaceras | ||||||||||||
| Wood, 1963 | ||||||||||||
Merkmale
Pappaceras ist ein kleiner Vertreter der Indricotherien. Er wurde bisher durch mehrere Schädel und Unterkiefer dokumentiert. Der Schädel wies eine Länge von rund 39 cm auf und war langgestreckt geformt. Am Rostrum stand der aufsteigende Abschnitt des Mittelkieferknochens mit dem Nasenbein in Kontakt. Dadurch entstand ein kleiner Naseninnenraum, der im hinteren Bereich etwa bis zur Position der Eckzähne reichte. Der horizontale Abschnitt besaß eine Bogenform. Das Nasenbein beanspruchte den gesamten Abschnitt der Voraugenregion. Es war dünn, im Querschnitt flach und endete vorn spitz. Wie bei allen Indricotherien wies es keine Ansatzstellen für eine Hornbildung auf. Die Knochennaht mit dem Stirnbein verlief bogenförmig eingezogen. Der Oberkiefer hatte in Seitenansicht eine Trapezgestalt und ragte hoch auf. Das Foramen infraorbitale lag oberhalb des vierten Prämolaren. Das Tränenbein reichte weit in den Gesichtsabschnitt hinein und war hier quadratisch geformt. Den Tränenkanal öffnete sich seitlich vor der Orbita. Der Jochbogen war grazil gebaut und kragte nur leicht aus. Sein vorderer Ansatz lag etwa 40 mm hinter dem vorderen Orbitarand. Auf dem Stirnbein waren zwei schwache Knochenrippeln ausgebildet, die nach hinten zogen und sich allmählich verdickten. Sie vereinten sich am Scheitelbein zu einem Scheitelkamm. Das Hinterhauptsbein war breit und nahezu senkrecht orientiert. Es wies nur einzelne schwache Knochenwülste auf. Der Processus paroccipitalis war schlank und nicht vollständig mit dem Processus posttympanicus verwachsen, zudem überragte er letzteren. Die Gelenkflächen zum Kontakt mit der Halswirbelsäule zeigten nach unten. An der Schädelbasis befand sich am Schuppenteil des Schläfenbeins die langovale Glenoidgrube zur Gelenkung mit dem Unterkiefer.[1]
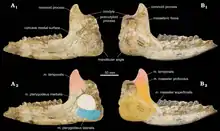
Der Unterkiefer war langgestreckt und verlief am unteren Rand nahezu gerade. Lediglich der Abschnitt der Symphyse besaß eine noch oben gerichtete Orientierung. Das Gebiss umfasste die vollständige Bezahnung der frühen Höheren Säugetiere mit folgender Zahnformel: . Insgesamt bestanden also 44 Zähne, allerdings ist es möglich, dass der jeweils erste Prämolar bei einigen Vertretern reduziert war und diese so nur über 40 Zähne verfügten. Die Schneidezähne wiesen mit Ausnahme des jeweils inneren Paares alle etwa die gleiche Größe auf. Allgemein waren sie klein und von konischer Gestalt. Somit fehlten die für späte Indricotherien typischen hypertrophierten Schneidezähne, was in etwa mit Forstercooperia übereinstimmt, bei diesem variierten sie aber stärker in der Größe. Zwischen den einzelnen Schneidezähnen waren kleine Diastemata ausgebildet, die im oberen Gebiss etwa 5 mm Breite besaßen, so dass das vordere Gebiss keine geschlossene Reihe bildete. Der ähnlich geformte Eckzahn überragte die Schneidezähne. Die vorderen Prämolaren ähnelten nicht den Molaren, waren also noch nicht molarisiert, die drei hinteren Prämolaren waren deutlich länger als der vorderste. Die Mahlzähne hatten eine quadratische bis trapezförmige Gestalt. Sie stimmten im Aufbau weitgehend mit denen der Nashörner überein mit einem auf den vorderen beiden Oberkiefermolaren charakteristischen π-förmigen Schmelzbandmuster (zusammengesetzt aus dem entlang der Zahnaußenkante verlaufenden Ectoloph und den rechtwinklig dazu stehenden Metaloph und Protoloph). Auf dem letzten oberen Molaren kamen einzelne markante topographische Marken vor, etwa der Metaconus und das Ectoloph, die bei Forstercooperia nicht ausgebildet sind. Die Länge der oberen und unteren Molaren variierte zwischen 25 und 34 mm, die Prämolaren waren entsprechend kürzer.[2][1][3]

Fossilfunde
Fast alle Fossilreste von Pappaceras kamen bisher in der Inneren Mongolei im Norden von China zu Tage. Bedeutend ist hier der östliche Teil des Erlian-Beckens nahe der Grenze zur Mongolei, das wichtigste Fundgebiet liegt südlich der Stadt Erlian. Es verteilt sich auf mehrere Fundlokalitäten im Umkreis von Iren Dabasu und Baron Sog. Den ältesten Fund barg hier die Arshanto-Formation, welche allgemein in das Untere Eozän datiert. Die Fundlage im oberen Abschnitt der Gesteinseinheit spricht eher für den Zeitraum des Übergangs vom Unteren zum Mittleren Eozän, was in einem Alterswert von rund 50 Millionen Jahren resultiert (Oberes Ypresium, lokalstratigraphisch Oberes Arshantum).[1] Bestätigt wird dies auch unter anderem durch paläomagnetische Untersuchungen. Dadurch repräsentiert der Fund den bisher ältesten eines Vertreters der Indricotherien. Es handelt sich um einen nahezu vollständigen, sehr gut erhaltenen Schädel. Die anderen Fossilreste aus dem Erlian-Becken sind der Irdin-Manha-Formation aus dem Mittleren Eozän zuzuweisen (Ypresium, lokalstratigraphisch Irdinmanhum). Es handelt sich um einzelne Teilschädel sowie zahlreiche Unterkiefer und Gebissreste.[2][4] Sowohl die Arshanto- als auch die Irdin-Manha-Formation bestehen aus sandigen Schluff- und feinkörnigen Sandsteinen, die teilweise von eingearbeiteten Kiesrinnen unterbrochen werden. Sie stellen Hinterlassenschaften ehemaliger Flüsse dar.[5][6] Ein nahezu vollständiger Unterkiefer wurde auch aus der Shara-Murun-Formation im Südosten der Fundregion berichtet. Er kann mit einer Alterstellung im ausgehenden Mitteleozän (lokalstratigraphisch Sharamurunium) als der bisher jüngste Fund der Gattung angesehen werden.[4] Als möglicherweise einer der wenigen Knochenreste von Pappaceras außerhalb der Inneren Mongolei wird ein Oberkiefer aus Lunan in der bezirksfreien Stadt Tangshan östlich von Peking eingestuft. Das Stück weist wohl ein obereozänes Alter auf.[7][3]
Paläobiologie

Die niederkronigen Zähne mit ihrer einfachen Struktur lassen eine auf weiche pflanzliche Kost basierte Ernährung annehmen. Das kann auch durch den generell schmalen Mittelkieferknochen angenommen werden. Der niedrige aufsteigende Ast des Unterkiefers und die dort ausgeprägte markante Fossa masseterica verweisen auf einen gut entwickelten Musculus masseter, der aber durch den eher grazilen Jochbogen nicht ganz so dominant war wie beim verwandten Juxia. Der Masseter-Muskel bewirkt hauptsächlich vertikale Kaubewegungen. Am Alisphenoid (Keilbeinflügelknochen) liegt eine Furche an, die die Ankerstellen für den Musculus pterygoideus medialis andeutet. Der Muskel ist gut bei heutigen Pferden und Tapiren entwickelt, bei Nashörnern spielt er dagegen keine Rolle. Er fungiert als Gegenspieler des Masseter-Muskels und lässt rotierende Kaubewegungen zu. Diese waren aber wahrscheinlich bei Pappaceras nur untergeordnet von Bedeutung, da etwa der flache Gaumen nur wenig Zwischenraum für die Zunge bei seitlichen Bewegungen ließ. Die spitzen unteren und oberen Schneidezähne griffen bei Gebissschluss ineinander und zeigen auffällige Abnutzungsfazetten. Möglicherweise konnte Pappaceras dadurch auch härtere Pflanzenteile verwerten, indem sie mit den vorderen Zähnen durchbohrt wurden. Die Schneidezähne eigneten sich aber nicht zum Zerschneiden von Nahrung wie die der Pferde mit ihren geraden Kanten.[8]
Systematik
Pappaceras ist eine Gattung aus der ausgestorbenen Familie der Indricotheriidae, die wiederum innerhalb der Überfamilie der Rhinocerotoidea steht und somit zur Nahverwandtschaft der heutigen Nashörner gehört. Die Indricotherien brachten große bis extrem große Formen hervor, darunter auch mit Paraceratherium eines der größten bekannten Landsäugetiere überhaupt. Generell wiesen die Vertreter der Familie abweichend von den Nashörnern keine Hornbildung auf der Nase auf. Des Weiteren können die Indricotheriidae anhand des vorderen Gebisses von den Nashörnern unterschieden werden. So besaßen erstere kurze, konische Schneidezähne, aus denen sich bei späteren Angehörigen je ein dolchartiges Paar im Ober- und Unterkiefer entwickelte. Die Nashörner verfügen dagegen nur über ein dolchartig geformtes Paar im Unterkiefer, während im Oberkiefer ein meißelartiges Schneidezahnpaar auftritt.[9][10] Innerhalb der Indricotherien bildet Pappaceras eine sehr frühe Form. Gemeinsam mit Forstercooperia wird es teilweise in die Unterfamilie der Forstercooperiinae gestellt.[1]
Innere Systematik der Familie der Indricotheriidae nach Wang et al. 2016[1]
|
Die Gattung enthält mehrere Arten:[1]
- Pappaceras confluens Wood, 1963
- Pappaceras meiomenus Wang, Bai, Meng & Wang, 2016
- Pappaceras minuta (Lucas, Schoch & Manning. 1981)
Von den drei Arten ist P. minuta mit einer Länge der oberen Molarenreihe von 74,5 bis 77,6 mm die kleinste Art, P. confluens bildet mit einer entsprechenden Länge von 87,1 bis 90,5 mm die größte. P. meiomenus liegt mit 77,6 bis 80,9 mm dazwischen. Bei letzterer sind zudem die vordersten Prämolaren reduziert. Sie ist außerdem mit einer Altersstellung im ausgehenden Unteren Eozän deutlich älter als die beiden anderen, die ins Mittlere Eozän datieren.[2][4][1]
Die wissenschaftliche Erstbeschreibung von Pappaceras erfolgte im Jahr 1963 durch Horace Elmer Wood. Sie basierte auf einem unvollständigen Schädel (Exemplarnummer AMNH 26660) zuzüglich eines Unterkiefers, beide Funde waren Mitte September 1930 während der Sixth American Museum Central Asiatic Expedition in der Irdin-Manha-Formation in der Mongolei gefunden worden. Der Gattungsname Pappaceras setzt sich aus den griechischen Wörtern παππος (pappos für „Großvater“), α (a oder alpha für „primitiv“ oder „ohne“) und κέρας (kéras für „Horn“) zusammen.[2] Nur wenige Jahre nach der Erstbeschreibung sah Leonard B. Radinsky die eigenständige Stellung von Pappaceras als problematisch an und synonymisierte die Gattung mit Forstercooperia. Dieses hatte Wood bereits 1938 anhand eines Teilschädels ebenfalls aus der Irdin-Manha-Formation eingeführt, zunächst als Cooperia und später in Forstercooperia umbenannt.[11] Die Gleichsetzung der beiden Gattungen fand weitgehend Zustimmung in der Fachwelt.[4][12] Die Entdeckung von weiterem Fundmaterial in der Arshanto-Formation, Innere Mongolei, führte aber im Jahr 2016 wieder zur Anerkennung von Pappaceras. Begründet wurde dies damit, dass sich Pappaceras und Forstercooperia in einzelnen Merkmalen der Zähne unterscheiden. Außerdem war bei der 2016 ebenfalls neu etablierten Art P. meiomenus der vorderste Prämolar im Oberkiefer bereits verschwunden, womit erste Ansätze zur Reduktion der Zahnanzahl des Gebisses erkennbar sind.[1] Spencer George Lucas und Forscherkollegen benannten im Jahr 1981 die neue Art Forstercooperia minuta. Sie stellten nicht nur zahlreiches Schädelmaterial aus der Irdin-Manha-Formation zu der Art, sondern auch einen linken Unterkiefer aus der Galisteo-Formation im Santa Fe County im US-Bundesstaat New Mexico.[4] Bei der Wiederanerkennung von Pappaceras 2016 überführten die Autoren der Studie die Art F. minuta in die Gattung, das Material aus Nordamerika wurde allerdings später aus den Indricotherien ausgegliedert.[3] Damit bleibt Pappaceras auf dem asiatischen Kontinent beschränkt. Eine vergleichbare Entwicklung gab es auch bei Forstercooperia, für das anfangs auch eine nordamerikanische Verbreitung angenommen wurde. Eine spätere Begutachtung des dortigen Fundmaterials resultierte aber in dem Ausschluss aus der Indricotherien und in der Beschreibung von Uintaceras als möglicherweise sehr frühen Vertreter der Nashörner.[13]
Literatur
- Haibing Wang, Bin Bai, Jin Meng und Yuanqing Wang: Earliest known unequivocal rhinocerotoid sheds new light on the origin of Giant Rhinos and phylogeny of early rhinocerotoids. Scientific Reports 6, 2016, S. 39607 doi:10.1038/srep39607
- Horace Elmer Wood: A primitive rhinoceros from the late eocene of Mongolia. American Museum Novitates 2146, 1963, S. 1–12
Einzelnachweise
- Haibing Wang, Bin Bai, Jin Meng und Yuanqing Wang: Earliest known unequivocal rhinocerotoid sheds new light on the origin of Giant Rhinos and phylogeny of early rhinocerotoids. Scientific Reports 6, 2016, S. 39607 doi:10.1038/srep39607
- Horace Elmer Wood: A primitive rhinoceros from the late eocene of Mongolia. American Museum Novitates 2146, 1963, S. 1–12
- Hai-Bing Wang, Bin Bai, Jin Meng und Yuan-Qing Wang: A New Species of Forstercooperia (Perissodactyla: Paraceratheriidae) from Northern China with a Systematic Revision of Forstercooperiines. American Museum Novitates 3897, 2018, S. 1–41
- Spencer G. Lucas, Robert M. Schoch und Earl Manning: The Systematics of Forstercooperia, a Middle to Late Eocene Hyracodontid (Perissodactyla: Rhinocerotoidea) from Asia and Western North America. Journal of Paleontology 55 (4), 1981, S. 826–841
- Jin Meng, Yuanqing Wang, Xijun Ni, Christopher Beard, Qian Li, Xun Jin und Bin Bai: New Stratigraphic Data from the Erlian Basin: Implications for the Division, Correlation, and Definition of Paleogene Lithological Units in Nei Mongol (Inner Mongolia). American Museum Novitates 3570, 2007, S. 1–31
- Yuanqing Wang, Jin Meng, Christopher K. Beard, Qian Li, Xijun Ni, Daniel L. Gebo, Bin Bai, Xun Jin und Ping Li: Early Paleogene stratigraphic sequences, mammalian evolution and its response to environmental changes in Erlian Basin, Inner Mongolia, China. Science China, Earth Sciences 53 (.12), 2010, S. 1918–1926
- Chow Minchen, Chang Yuping und Ting Suyin: Some early Tertiary Perissodactyla from Lunan Basin, E. Yunnan. Vertebrata Palasiatica 12 (4), 1974, S. 262–273
- Hai-Bing Wang, Bin Bai, Yan-Xin Gong, Jin Meng und Yuan-Qing Wang: Reconstruction of the cranial musculature of the paraceratheriid rhinocerotoid Pappa ceras meiomenus and inferences of its feeding and chewing habits. Acta Palaeontologica Polonica 62 (2), 2017, S. 259–271
- Leonard B. Radinsky: The families of the Rhinocerotoidea (Mammalia, Perissodactyla). Journal of Mammalogy 47 (4), 1966, S. 631–639
- Leonard B. Radinsky: A review of the Rhinocerotoid Family Hyracodontidae (Perissodactyla). Bulletin of the American Museum of Natural History 136 (1), 1967, S. 1–47
- Horace Elmer Wood: Cooperia totadentata, a remarkable rhinoceros from the eocene of Mongolia. American Museum Novitates 1012, 1938, S. 1–22
- Spencer George Lucas und Jay C. Sobus: The systematics of Indricotheres. In: Donald R. Prothero und R. Schoch (Hrsg.): The evolution of Perissodactyls. New York, Oxford University Press, 1989, S. 358–378
- Luke T. Holbrook und Spencer George Lucas: A new genus of rhinocerotoid from the Eocene of Utah and the status of North American "Forstercooperia". Journal of Vertebrate Paleontology 17 (2), 1997, S. 384–396