Phorusrhacidae
Die Phorusrhacidae, auch „Terrorvögel“, sind eine ausgestorbene Familie der Vögel. Es handelt sich um teils große, bodenbewohnende und meist flugunfähige Tiere, deren Fossilien vor allem in Südamerika, aber auch in Nordamerika gefunden wurden. Reste aus anderen Erdteilen sind in ihrer Zuweisung umstritten. Die „Terrorvögel“ nahmen während des Paläogens und des Neogens in Südamerika die Rolle der Topräuber ein. Die rezent nächstverwandten Vögel sind die südamerikanischen Seriemas (Cariamidae). Seit ihrer wissenschaftlichen Einführung im ausgehenden 19. Jahrhundert wurden mehr als ein Dutzend Gattungen mit über zwanzig Arten beschrieben. Neben den mitunter enormen Körperausmaßen der Phorusrhacidae – die größten Vertreter wurden 2,4 m, eventuell auch bis zu 3 m hoch – stellen der schmale Schädel und Brustkorb, die langen Hintergliedmaßen, das hakenförmig nach unten gebogene Schnabelende sowie die stark gekrümmten letzten Zehenglieder typische Merkmale dar. Letztere beiden Charakteristika können als Anpassung an die fleischhaltige Nahrung gewertet werden. Der gesamte Körperbau verweist auf schnelläufige Beutegreifer, die die damals offenen Landschaften Südamerikas bewohnten. Die ältesten sicheren Funde von „Terrorvögeln“ datieren in das Oligozän, die jüngsten Fossilreste stammen aus dem Oberpleistozän. Mit wenigen Ausnahmen ist das überwiegende Fundmaterial aber stärker fragmentiert.
| Phorusrhacidae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Paraphysornis | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oligozän bis Oberes Pleistozän | ||||||||||||
| 35 Mio. Jahre bis 18.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Phorusrhacidae | ||||||||||||
| Ameghino, 1889 | ||||||||||||
Merkmale

Die Angehörigen der Phorusrhacidae waren flugunfähige Vögel, die mittelgroße bis sehr große Arten umfassten. Sie erreichten ein Körpergewicht von etwa 5 bis zu 180 kg und wurden 0,8 bis 2,4 m, möglicherweise auch bis zu 3 m hoch. Ein generelles, die Gruppe definierendes Merkmal findet sich in dem kräftigen, allerdings seitlich verschmälerten Körperbau, der den Tieren in Anblick von vorn eine schlanke Gestalt verlieh. Dies betrifft vor allem den Schädel, den Brustkorb und das Becken. Auffallend war des Weiteren der hohe und schmale Oberkiefer, der am vorderen Ende spitz nach unten bog und so ein hakenförmiges Ende aufwies. Dies findet sich auch bei heutigen Greifvögeln und kann als eine Anpassung an fleischliche Nahrung interpretiert werden. Dem entsprach auch der Unterkiefer mit seiner fest verwachsenen Symphyse. Die Nasenlöcher waren groß und wurden nicht von einer Scheidewand getrennt. Weiterhin auffällig zeigten sich der lange Hals und die verkleinerten Flügel. Letzteren gegenüber waren die Hintergliedmaßen äußerst kräftig, am Tarsometatarsus trat ein im Querschnitt dreieckiger Hypotarsus auf, ein Wulst am oberen Ende. Das Becken wiederum war im vorderen Bereich stark zurückgebildet, was unter anderem auch von den Habichtartigen bekannt ist. Als zusätzlicher Hinweis auf eine beutegreifende Ernährungsweise wiesen die Füße spitz gebogene Endzehen auf, die kräftige Krallen trugen. Ergänzende Charakteristika betreffen spezielle Skelettmerkmale. Hierzu zählen etwa die extrem reduzierten Fortsätze am Rabenbein, das Fehlen der hakenartigen Fortsätze an den Rippen und der deutlich gebogene obere Schaftverlauf des Oberarmknochens.[1][2][3]
Paläobiologie

Generell waren die Phorusrhacidae an eine schnelle Fortbewegung am Boden angepasst, worauf neben dem hohen Becken auch die verlängerten unteren Gliedmaßenpartien der Laufbeine (Tibiotarsus und Tarsometatarsus) verweisen. Bei den meisten Formen mit bekanntem Beinskelett erreicht der Tarsometatarsus zwischen 60 und 85 % der Länge des Tibiotarsus, was für ausgesprochen gestreckte Füße spricht. In absoluten Zahlen ausgedrückt besitzt der Tibiotarsus bei Phorusrhacos eine Länge von rund 50 cm, der Tarsometatarsus von 38,5 cm. Entsprechende Maße bei Mesembriornis lauten 42 cm und 36 cm. Zudem sind die Knochen relativ grazil, der Schaftdurchmesser des Tarsometatarsus bei Phorusrhacos beträgt 3,7 cm, bei Mesembriornis 2,7 cm. Alle diese Merkmale verweisen auf gute Laufeigenschaften. Eine Ausnahme bildet hier aber Paraphysornis. Diese sehr große Form hat einen deutlich robusteren Skelettbau, bei dem der Tarsometatarsus weniger als 60 % der Länge des Tibiotarsus ausmacht. Demnach wurde ersterer etwa 31,5 cm lang, letzterer gut 55 cm. Dadurch sind die Füße bei Paraphysornis nicht nur relativ, sondern auch absolut kürzer als im Vergleich zum gleichgroßen Phorusrhacos. Auch der Schaftdurchmesser des Tarsometatarsus ist bei Paraphysornis mit 5,7 cm deutlich umfangreicher. Es wird daher angenommen, dass die Form weniger stark an einen schnellen Lauf angepasst war und eventuell als Lauerjäger oder Aasfresser in Erscheinung trat. Als weiterer Hinweis auf eine agile Fortbewegung bei den meisten Phorusrhaciden kann der Bau der Füße gewertet werden. Diese sind mit drei nach vorn zeigenden Zehen ausgestattet, während ein vierter Zeh verkleinert ist und nicht den Boden berührte. Eine reduzierte Anzahl der Zehen bei Kontakt mit dem Untergrund verringert wiederum den Reibungswiderstand beim Laufen. Das Merkmal ist vor allem bei den heutigen Straußen extrem ausgeprägt, die nur über zwei funktionale Zehen je Fuß verfügen.[1][4][5]
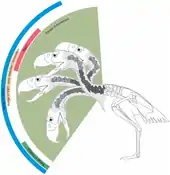
Die kräftigen und langgestreckten Beinknochen ermöglichten somit eine hohe Laufgeschwindigkeit. Schätzungen und Vergleiche mit heutigen Straußen befürworten für mehrere der größeren Formen der Phorusrhacidae Geschwindigkeiten von bis zu 50 Kilometer pro Stunde. Der außerordentlich massive Tibiotarsus vom Mesembriornis würde theoretisch sogar bis zu 97 Kilometer pro Stunde zulassen. Da diese Geschwindigkeit heute lediglich vom Gepard erreicht wird, nehmen Wissenschaftler an, dass die Vögel höchstwahrscheinlich mit kräftigen Tritten ihre Beute attackierten. Damit wären sie in der Lage, Reaktionskräfte von dem dreinhalbfachen Wert ihres eigenen Körpergewichts zu erzeugen, was umgerechnet etwa 2400 Newton entspricht. Die Kraft hätte ausgereicht, um bei heutigen mittelgroßen Huftieren wie der Saiga, den Thomson-Gazellen oder dem Alpensteinbock bedrohliche Knochenbrüche zu erzeugen.[6][5]
Bei einem Angriff gebrauchten die Vögel außerdem ihren Kopf mit dem großen Schnabel wie eine Axt, wie Untersuchungen am Schädel von Andalgalornis zeigten. Ausschlaggebend hierfür ist der stark verfestigte Bau des Schädels und der große und hohe sowie seitlich komprimierte Schnabel, die beide horizontalen Scherkräften nur wenig widerstehen konnten, dafür aber besser mit vertikalen Bewegungen zurechtkamen. Die an der Schnabelspitze erzeugten Kräfte waren dagegen mit rund 133 Newton eher gering. Dadurch kamen als Beute entweder nur kleinere Tiere in Betracht oder aber größere wurden mit schnellen und präzisen Hieben erlegt.[7] Der kräftig gebaute Hals ermöglichte dabei nicht nur den großen und massiven Schädel zu halten, er unterstützte vor allem auch schnelle Auf- und Abwärtsbewegungen. Dadurch waren im Zuge eines Angriffs einerseits Schläge gegen ein Beutetier möglich, andererseits konnte so der Kopf wiederum schnell in die Ausgangsposition zurückgebracht werden.[8]
Als mögliche Hinweise auf die Ernährungsweise der Phorusrhacidae werden einzelne fossilisierte Gewölle aus der Andalgala-Formation in der argentinischen Provinz Catamarca gewertet. Die aus dem Oberen Miozän stammenden ovaloiden Strukturen sind bis zu 4 cm lang und 1,9 cm breit sowie 1,0 cm hoch. Sie enthalten eine dichte Packung aus Gebissresten und Langknochen, die überwiegend zu Trugratten und Wühlern gehören. Die einzigen bekannten Nachweise räuberischer Vögel aus der Gesteinseinheit wurden als Procariama und Andalgalornis identifiziert, zwei eher kleine Vertreter der Phorusrhaciden.[9] Für eine genaue Beurteilung fehlen allerdings bisher Vergleichsfunde.[10]
Systematik
Äußere Systematik
Nähere Verwandtschaft der Phorusrhacidae nach Worthy et al. 2017[11]
|
Die Phorusrhacidae sind eine ausgestorbene Familie der Vögel (Aves). Die genaue systematische Stellung der Gruppe wurde im Laufe der Geschichte recht unterschiedlich beurteilt. Eine in der Anfangszeit angenommene Zuordnung zu den aus heutiger Sicht eher uneinheitlichen Laufvögeln[12] konnte bereits 1899 durch Charles W. Andrews vom British Museum in London widerlegt werden. Er wies nach, dass es sich um Verwandte der heute noch in südamerikanischen Savannen lebenden Seriemas (Cariamidae) handelt.[13] Teilweise wird dadurch eine engere Beziehung zu den ebenfalls ausgestorbenen Familien der Bathornithidae aus Nordamerika und der Idiornithidae aus Europa gesehen. Beide Gruppen lebten vom Eozän bis zum frühen Oligozän. Alle vier Familien gehören in einer eher traditionellen Auffassung zur Ordnung der Kranichvögel (Gruiformes) und werden darin in einer Überfamilie der Cariamoidea[14] oder einer Unterordnung der Cariamae[15] zusammengefasst. Da neuere, sich auf vergleichende DNA-Sequenzanalysen stützende Forschungen die Monophylie der Kranichartigen in Frage stellen und die Seriemas außerhalb der Ordnung sehen,[16] ist auch die Zugehörigkeit der Phorusrhacidae zu den Kranichartigen inzwischen zweifelhaft und in neueren Veröffentlichungen werden sie zusammen mit den Seriemas der Ordnung der Cariamiformes zugeordnet.[17]
Überblick über die Gattungen
Folgende Gattungen sind innerhalb der Phorusrhacidae anerkannt:[1][18][19][20][21]
- Familie: Phorusrhacidae Ameghino, 1889
- Andalgalornis Patterson & Kraglievich, 1960; Oberes Miozän bis Unteres Pliozän (Argentinien)
- Andrewsornis Patterson, 1941; Mittleres bis Oberes Oligozän (Argentinien)
- Devincenzia Kraglievich, 1932; Oberes Miozän bis Oberes Pliozän/Unteres Pleistozän (Argentinien und Uruguay)
- Kelenken Bartelli, Chiappe & Tambussi, 2007; Mittleres Miozän (Argentinien)
- Llallawavis Degrange, Tambussi, Taglioretti, Dondas & Scaglia, 2015; Oberes Pliozän (Argentinien)
- Mesembriornis Moreno, 1889; Oberes Miozän bis Oberes Pliozän (Argentinien)
- Paraphysornis Alvarenga, 1993; Oberes Oligozän bis Unteres Miozän (Brasilien)
- Patagorhacos Agnolin & Cafrat, 2015; Unteres Miozän (Argentinien)
- Patagornis Moreno & Mercerat, 1891; Unteres bis Mittleres Miozän (Argentinien)
- Phorusrhacos Ameghino, 1887; Unteres bis Mittleres Miozän (Argentinien)
- Physornis Ameghino, 1895; Mittleres bis Oberes Oligozän (Argentinien)
- Procariama Rovereto, 1914; Oberes Miozän bis Unteres Pliozän (Argentinien)
- Psilopterus Moreno & Mercerat, 1891; Mittleres Oligozän bis Oberes Pleistozän (Argentinien)
- Titanis Brodkorb, 1963; Pliozän bis Unteres Pleistozän (Nordamerika)
Eine fragliche Stellung innerhalb der Phorusrhacidae weist folgende Gattung auf:[3]
- Palaeopsilopterus Alvarenga, 1985; Mittleres Paläozän (Brasilien)
Darüber hinaus wurden drei weitere Gattung beschrieben:[22][23][24]
- Eleuterornis Schaub, 1940; Mittleres Eozän (Frankreich)
- Lavocatavis Mourer-Chauviré, Tabuce, Mahboubi, Adaci & Bensalah, 2011; Mittleres Eozän (Algerien)
- Macrornis Seeley, 1866; Oberes Eozän (England)
Aufgrund ihres Auftretens in Europa und Afrika sehen zahlreiche Autoren die Position der drei Gattungen innerhalb der Phorusrhacidae oder deren unmittelbaren äußeren Verwandtschaft kritisch. Das Fundmaterial ist zumeist spärlich. So liegt von dem im Jahr 2011 eingeführten Lavocatavis nur ein einzelner Oberschenkelknochen aus der Glib-Zegdou-Formation vor,[22] das zwei Jahre später zu den Phorusrhacidae gestellte Eleutherornis basiert auf einzelnen Bein- und Fußknochen aus Lissieu bei Lyon.[23] Gleiches kann zu Macrornis gesagt werden. Hierbei handelt es sich um einen großen Vogel mit ursprünglich weitgehend umbekannten Beziehungen, der anhand eines einzelnen Tibiotarsus aus der Totland-Bay-Formation in Hampshire bekannt ist.[24] Darüber hinaus wurde Ameghinornis aus dem aquitanischen Becken in Frankreich als weiterer Vertreter der Phorusrhacidae vorgestellt. Dieser rund 38 bis 35 Millionen Jahre alte und damit in das Mittlere Eozän datierende Vogelvertreter wurde nur so groß wie ein Seriema und konnte offenbar noch fliegen.[25] Die Form gilt aber nun als identisch mit Strigogyps aus der Gruppe der Ameghinornithidae.[26][27]
Neben diesen Funden sind von der antarktischen King-George-Insel zwei einzelne, dreizehige 18 cm lange Fußabdrücke aus der Zeit von vor 55 Millionen Jahren bekannt, die entweder einem Phorusrhaciden oder einem Laufvogel zugeordnet werden. Außerdem stammt aus 40 Millionen Jahre alten Schichten auf der Seymour-Insel der Vorderteil eines Schnabels, der als zu den Phorusrhacidae gehörig angesprochen wurde. Beide Funde gelten als problematisch, letzterer ist weiteren Untersuchungen zufolge eher unspezifisch und kann momentan keiner Gruppe genauer zugeordnet werden.[28] Somit existieren für alle Funde außerhalb von Nord- und Südamerika entgegenstehende Merkmale, alternative Deutungen oder kritische Ansichten, so dass ein Vorkommen der Phorusrhacidae in der Alten Welt und anderen Erdteilen bisher als äußerst zweifelhaft gilt.[29]
Gliederungsversuche
Für die süd- und nordamerikanischen Formen schlugen Herculano M. F. Alvarenga und Elizabeth Höfling im Jahr 2003 eine Unterteilung der Phorusrhacidae in fünf Unterfamilien vor:[1]
- Familie: Phorusrhacidae Ameghino, 1889
- Unterfamilie: Brontornithinae Moreno & Mercerat, 1891 (Brontornis, Paraphysornis, Physornis)
- Unterfamilie: Phorusrhacinae Ameghino, 1889 (Devincenzia, Phorusrhacos, Titanis, zuzüglich von Kelenken)
- Unterfamilie: Patagornithinae Mercerat, 1897 (Patagornis, Andrewsornis, Andalgalornis)
- Unterfamilie: Psilopterinae Dolgopol de Saez, 1927 (Psilopterus, Procariama, Palaeopsilopterus)
- Unterfamilie: Mesembriornithinae Kraglievich, 1932 (Mesembriornis, zuzüglich Llallawavis)
Die Brontornithinae enthielten nach Alvarenga und Höfling Brontornis. Dieser wurde im Jahr 2007 von Federico L. Agnolin ausgeschlossen und in eine nähere Verwandtschaft mit den Gänsevögeln verschoben. Die verbliebene Gruppe benannte er in die von ihm neu geschaffene Unterfamilie der Physornithinae um.[30] Hierbei umfassen die Physornithinae und die Phorusrhacinae große bis sehr große Formen, unter anderem mit Kelenken den größten bekannten Vertreter der Phorusrhacidae. Sie wiesen Körperhöhen von bis zu 2,4 oder sogar auch bis zu 3 m und ein Gewicht bis zu 180 kg auf. Ihr zeitliches Auftreten umfasst den Zeitraum von vor 27 bis 1,8 Millionen Jahren. Die anderen drei Unterfamilien schließen meist kleine bis mittelgroße Arten von etwas unter 1,0 bis 1,5 m Körperhöhe ein, wobei die Patagornithinae eine Mittelstellung einnehmen. Zu den kleinsten Formen der Phorusrhacidae gehören mehrere Angehörige von Psilopterus, die nur rund 0,6 bis 0,8 m hoch wurden und rund 5 kg schwer waren. Ein Teil der kleineren Formen hatte möglicherweise einen Rest Flugfähigkeit bewahrt. Fossilien aus diesen drei Gruppe sind 63 Millionen bis knapp 100.000 Jahre alt.[31][1]
Im Jahr 2009 überarbeitete Agnolin in einer umfassenden Revision die Gliederung der Phorusrhacidae erneut. Hierbei spaltete er die Gruppe in zwei Familien auf. Die kleineren Formen verwies er zu den Psilopteridae. Innerhalb der eigentlichen Phorusrhacidae behielt er die größeren Formen um Phorusrhacos, wobei er den robusten Vertretern einen eigenen Status als Tribus der Physornithini vorbehielt. Dadurch ergab sich folgende Gliederung:[3]
- Überfamilie: Phorusrhacoidea Patterson, 1941
- Familie: Psilopteridae Dolgopol de Saez, 1927
- Incertae sedis (Hermosiornis (einschließlich Mesembriornis))
- Unterfamilie: Psilopterinae Dolgopol de Saez, 1927 (Psilopterus, Procariama)
- Familie Phorusrhacidae Ameghino, 1889
- Incertae sedis (Andrewsornis, Patagornis)
- Unterfamilie: Phorusrhacinae Ameghino, 1889 (Andalgalornis, Devincenzia, Phorusrhacos, Titanis, Kelenken)
- Tribus: Physornithini Agnolin, 2007 (Paraphysornis, Physornis)
Brontornis stand in dieser Gliederung weiterhin außerhalb der Verwandtschaftsgruppe, während Palaeopsilopterus im Umfeld der Seriemas eingeordnet wurde.[3]
Innere Systematik der Phorusrhacidae nach Degrange et al. 2015[20]
|
Weitergehende phylogenetische Untersuchungen erbrachten wenig Unterstützung für die angenommene Unterteilung in mehrere Unterfamilien nach Alvarenga und Höfling 2003. Vielmehr stützten sie eine prinzipielle Zweiteilung, wie sie von Agnolin postuliert wurde. Auf der einen Seite bilden vor allem die großen Vertreter der Phorusrhacidae eine eher in sich geschlossene Gruppe, die sich aus den Vertretern der Phorusrhacinae, der Physornithinae und der Patagornithinae zusammensetzt. Die Tiere waren in der Regel schwerer als 70 kg. Für die Gruppe wurde die vorläufige Bezeichnung als „echte Terrorvögel“ vorgeschlagen. Die zweite Gruppe wird durch die eher kleinwüchsigen Angehörigen der Psilopterinae und zusätzlich der Mesembriornithinae gebildet, jedoch ergaben sich innerhalb der Unterfamilien einzelne Verschiebungen. Sie unterscheiden sich von den „echten Terrorvögeln“ durch ihre möglicherweise noch vorhandenen Flugeigenschaften bei einigen Formen. Mit Mesembriornis tritt hier eine Form auf, die allerdings ebenfalls relativ groß wurde und gut 70 kg wog. Vorläufig erhielt die zweite Gruppe den Terminus „Psilopterines“. Die Aufteilung in eine große und eine kleine Formengruppe, bei der letztere aber auch einzelne größere Mitglieder einschließt, führt zu der Annahmen, dass der enorme Körperzuwachs innerhalb der Phorusrhacidae mehrfach und möglicherweise unabhängig voneinander stattfand.[20]
Die Zweiteilung der Phorusrhacidae spiegelt sich im Schädelbau wieder. Jede Gruppe verfügt über einen eigenständigen Morphotyp. Der Schädel der „Psilopterines“ erinnert noch deutlich an dem der nahe verwandten Seriemas und anderer Vögel, bei denen durch flache und dünne Knochen eine sehr flexible Konstruktion entsteht. Dem Gegenüber haben die „echten Terrorvögel“ durch Knochenverdickungen einen extrem verfestigten und starren Schädel. Beiden gemeinsam ist allerdings der hohe, seitlich verschmälerte Schnabel. Dieses Merkmal ist sonst nur von anderen ausgestorbenen großen bodenlebenden Vögeln überliefert, beispielsweise Gastornis oder den Donnervögeln. Einzig die Phorusrhacidae bildeten ein hakenförmig nach unten gebogenes Schnabelende heraus, das auf die räuberische Ernährungsweise deutet. Dass dieses sowohl bei den „Psilopterines“ als auch bei den „echten Terrorvögel“ vorkommt, wird trotz des unterschiedlichen Schädelbaus als Hinweis auf eine ähnliche Ernährungsweise angesehen.[32]
Stammesgeschichte
Ursprünge und Verbreitung
Sofern Palaeopsilopterus als Mitglied der Phorusrhacidae anzusehen ist, stammen die ältesten Funde bisher mit einzelnen Beinknochen aus Itaboraí bei Rio de Janeiro in Brasilien und datieren in das Mittlere Paläozän. Einzelne Autoren gruppieren die Vogelgattung allerdings an die Basis der Cariamiformes.[3] Weitere Funde kamen dann erst aus dem Mittleren Oligozän zu Tage, sie können verschiedenen Formen zugeordnet werden. Hierbei handelt es sich einerseits im Physornis, andererseits um Andrewsornis und Psilopterus. Ersterer ist über einzelne fragmentierte Schädel- und Skelettknochen aus der Provinz Santa Cruz in Argentinien belegt. Von letzterem sind mehrere Arten beschrieben worden, die älteste ließ sich über einen bruchstückhaften Tarsometatarsus aus der Provinz Chubut, ebenfalls Argentinien, nachweisen. Andrewsornis, definiert anhand eines bruchstückhaften Schädels,[33] stammt aus räumlicher und zeitlicher Nähe. Die Dokumentation der drei Formen in Patagonien zeigt auf, dass zu dieser Zeit sowohl einer der größten als auch einer der kleinsten sowie ein mittelgroßer Vertreter der Phorusrhaciden im südlichen Südamerika präsent waren. Im Übergang vom Oligozän zum Miozän trat dann der gleichfalls große Paraphysornis auf, von dem ein nahezu vollständiges Skelett aus der Tremembé-Formation bei São Paulo in Brasilien vorliegt.[34][35] Anhand eines einzelnen Schädelknochens und eines Fragmentes der Vordergliedmaßen wurde der mittelgroße Patagorhacos aus der untermiozänen Chichinales-Formation im nördlichen Patagonien beschrieben.[21] Von Bedeutung ist vor allem das recht umfangreiche Material aus der Santa-Cruz-Formation wiederum im Patagonien, die im ausgehenden Unteren Miozän zur Ablagerung kam. Die bedeutende Fossillagerstätte und Gesteinseinheit brachte nicht nur die Charakterform Phorusrhacos hervor, hier wurden auch Psilopterus und Patagornis aufgefunden. Allen drei Angehörigen der „Terrorvögel“, die eine große, eine kleine und eine intermediäre Form repräsentieren, sind Schädelfunde und Teile des Körperskelettes zuordbar.[17][36][37][38]

Nachfolgend im Miozän sind unter anderem große Angehörige wie Kelenken und Devincenzia präsent. Von Kelenken ist bisher nur ein Schädel und ein Beinknochen aus Patagonien bekannt, die Funde lassen aber auf den möglicherweise größten Angehörigen der Phorusrhacidae schließen. Sie kamen in der Collón-Curá-Formation am Río Negro zu Tage.[39] Nur wenig kleiner wurde Devincenzia, der in der heutigen Pamparegion beheimatet war. Ähnlich zu seinem anderen großen Verwandten sind auch hier weitgehend nur Einzelfunde dokumentiert.[40][41] Ein Teilskelett und einzelne Gliedmaßenknochen aus der Andalgala-Formation im nördlichen Argentinien werden wiederum zu Andalgalornis gezählt. Der eher grazil gebaute „Terrorvogel“ trat im Übergang vom Miozän zum Pliozän auf. Etwa gleichalt ist Mesembriornis aus der Monte-Hermoso- und Chapadmalal-Formations im zentralen Argentinien. Neben Phorusrhacos gehört die Gattung zu den wissenschaftlich am frühesten bekannten Vertretern der Phorusrhacidae, erreichte aber nicht die Ausmaße der Charakterform. Ein fast vollständiges Skelett aus der Playa-Los-Lobos-Allo-Formation bei Mar del Plata in Argentinien verweist auf den kleinen, nur 20 kg schweren Llallawavis. Die Gesteinseinheit entstand im Mittleren Pliozän. Die gut erhaltene Ohrregion ermöglichte die Rekonstruktion des Hörvermögens der Tiere, die niedrigere Frequenzen verarbeiten konnten als es bei rezenten Vögeln der Fall ist.[20] Mit der Entstehung des Isthmus von Panama, das heißt der Bildung einer Landbrücke zwischen Nord- und Südamerika im Verlauf des Pliozäns, breiteten sich die Phorusrhacidae auch nach Nordamerika aus. Als stratigraphisch jüngster Fund galt lange Zeit der rund 2,5 bis 1,5 Millionen Jahre alte Titanis aus dem südlichen Teil der heutigen USA. Das nur fragmentiert überlieferte Fossilmaterial lässt einen großen Vertreter annehmen.[42][1]
Im Laufe ihrer Entwicklung nahmen die „Terrorvögel“ in Südamerika zusammen mit den fleischfressenden Beutelsäugern der Ordnung der Sparassodonta und den terrestrischen Krokodilen der Familie der Sebecidae die Rolle der hier fehlenden Raubtiere (Carnivora) ein. Während die beiden anderen Gruppen aber eher langsam waren, spezialisierten sich die meisten der Phorusrhaciden auf schnelle Beute in den im Oberen Oligozän vor 27 Millionen Jahren zunehmend offener werdenden Trockenwäldern und Savannen Südamerikas. Bis zum Beginn des Pliozäns verschwanden die räuberischen Beutelsäuger zum Großteil aus den offenen Landschaften Südamerikas, eventuell durch Konkurrenz mit den Phorusrhaciden oder bedingt durch klimatische Veränderungen.[43][44][45]
Aussterben
Die großen Phorusrhaciden waren in Südamerika und Nordamerika weitgehend im ausgehenden Pliozän bis beginnenden Pleistozän verschwunden. Es verblieben lediglich die kleineren Formen in Südamerika.[46] Im Jahr 2010 publizierte eine Forschergruppe um Herculano M. F. Alvarenga nicht genauer bestimmte Reste eines Vertreters der Phorusrhacidae aus Uruguay, die ein absolutes Alter von nur 17.620 ± 100 Jahre haben.[47] Dies entspricht chronostratigraphisch dem obersten Pleistozän. Teilweise wird der Rest Procariama zugeordnet,[48] doch sehen einige Autoren einen Verweis zu den Phorusrhaciden auch kritisch.[49][50] Ein weiterer Fund von Psilopterus aus Uruguay mit einem OSL-Alter von 96.040 ± 6300 Jahren, der 2018 von Washington Jones und Kollegen veröffentlicht wurde, belegt zusätzlich das Überleben der Gruppe bis ins Obere Pleistozän.[31] Demnach starben in Südamerika die Phorusrhacidae relativ spät aus. Über die Ursache ihres Verschwindens ist nur sehr wenig bekannt. Die Bildung des Isthmus von Panama ermöglichte es nicht nur den „Terrorvögeln“ nach Nordamerika zu wandern, sie bot auch die Chance für die Einwanderung von Raubtieren wie den Säbelzahnkatzen (Machairodontinae) und den Hunden (Canidae) nach Südamerika.[43] Es ist zu vermuten, dass es dadurch zu einer stärkeren Konkurrenz zwischen diesen beiden Gruppen von Beutegreifern kam.[46]
Forschungsgeschichte
Der erste Phorusrhacide wurde 1887 durch den argentinischen Naturforscher Florentino Ameghino nach einem Unterkiefer beschrieben, gefunden in der Santa-Cruz-Formation in der argentinischen Provinz Santa Cruz. Hierbei verwendete er erstmals die wissenschaftliche Bezeichnung Phorusrhacos.[51] Der Name leitet sich höchstwahrscheinlich von den griechischen Wörtern φορός (phoros) für „tragend“ oder „bringend“ und ῥάκος (rakos) für „zerrissen“ oder „zerlumpt“ ab, er bezieht sich möglicherweise auf die bruchstückhafte Erhaltung des Fundes.[52] Ameghino hielt den Unterkiefer anfangs für den Rest eines zahnlosen Säugetiers, das er in die Nähe der Faultiere und Gürteltiere einordnete. Zwei Jahre später wandelte er den Gattungsnamen in einem umfangreichen Katalogwerk zur Fauna Argentiniens in Phororhacos ab und begründete gleichzeitig die darauf basierende Familienbezeichnung Phororhacosidae.[53] Die Erkenntnis, dass es sich hierbei um den Rest eines riesigen Vogels handelte, brachte er wiederum im Jahr 1891 nach der Entdeckung eines weitgehend vollständigen Schädels zu Papier, wobei er sowohl die von ihm neu gewählte Schreibweise der Gattung als auch den Namen der Familie beibehielt.[54][12] Richard Lydekker korrigierte im Jahr 1893 die Bezeichnung der Familie in Phororhacidae.[55] In der nachfolgenden Zeit blieben beide Namen, Phororhacos und Phororhacidae, weitgehend in Gebrauch.[56] Erst im Jahr 1963 wies Pierce Brodkorb darauf hin, dass aufgrund der Namenspriorität in der zoologischen Nomenklatur Phorusrhacos vorzuziehen ist. Der korrekte Familienname lautet demnach Phorusrhacidae.[42] Dies wurde im Jahr 1992 in einer Sitzung der International Commission on Zoological Nomenclature offiziell bestätigt (Opinion 1687).[57][1] Unabhängig davon wird die heute noch verwendete Bezeichnung der Überfamilie der Phororhacoidea, basierend auf Ameghinos korrigierter Schreibweise, in der Regel auf Bryan Patterson aus dem Jahr 1941 zurückgeführt.[33] Analog zur Familie ist allerdings die nomenklatorisch genauere Version mit Phorusrhacoidea zu benennen, was bereits Brodkorb in den 1960er Jahren anmerkte.[58][59]
Vor allem die Frühphase der Erforschung der Phorusrhacidae war bestimmt von einer Fehde, die in etwa an die sogenannten „Bone Wars“ in Nordamerika erinnerte, welche zwischen den Forschern Othniel Charles Marsh und Edward Drinker Cope in den 1870er und 1880er Jahren ausgetragen wurde. In Südamerika konkurrierte Ameghino mit Francisco Pascasio Moreno, dem damaligen Direktor des La-Plata-Museums in der gleichnamigen Stadt in Argentinien. Neben wichtigen Fossilfunden beinhaltete die Fehde auch die Deutungshoheit über die wissenschaftliche Namensgebung.[60] Kurz nach Ameghinos Benennung der Familie der Phorusrhacidae erstellte Moreno zusammen mit Alcide Mercerat im Jahr 1891 ein eigenes Konzept der systematischen Zuordnung der „Terrorvögel“. Beide etablierten die Gruppe der Stereornithes, die sie als übergeordnete Einheit auffassten. In dieser vereinten sie verschiedene Familien wie die Brontornithidae und die Stereornithidae. In letzterer gliederten Moreno und Mercerat Phorusrhacos, Mesembriornis und Patagornis ein.[61] Das Stereornithes-Konzept wurde teilweise noch bis in das 20. Jahrhundert übernommen und abgewandelt. So unter anderem führte Lucas Kraglievich im Jahr 1932 innerhalb der Stereornithes die Familien der Phororhacidae sowie der Psilopteridae und erweiterte sie um die Devincenziidae.[41] Mit dem Aufkommen moderner phylogenetischer Untersuchungsmethoden verlor das Konzept an Bedeutung, zumal die Typusform Stereornis lediglich ein Synonym zu Phorusrhacos ist.[1]
Rezeption
In der Popkultur wird unter anderem im Videospiel ARK: Survival Evolved Bezug auf die „Terrorvögel“ genommen.[62] Auch die fünfte Episode der erstmals im Jahr 2001 ausgestrahlten BBC-Serie Walking with Beasts thematisiert die Tiere. Namentlich erwähnt wird hierbei Phorusrhacos, der laut der Serie mit der Säbelzahnkatze Smilodon konkurrierte. Beide waren jedoch keine Zeitgenossen.[63]
Literatur
- Federico Agnolín: Sistemática y filogenia de las aves fororracoideas (Gruiformes, Cariamae). Buenos Aires, Fundación de Historia Natural Félix de Azara, 2009, S. 1–78.
- Herculano M. F. Alvarenga und Elizabeth Höfling: Systematic revision of the Phorusrhacidae (Aves: Ralliformes). Papéis Avulsos de Zoologia 43 (4), 2003, S. 55–91, PDF.
- Herculano Alvarenga, Luis Chiappe und Sara Bertelli: Phorusracids: The Terror birds. In: Gareth Dyke und Gary Kaiser (Hrsg.): Living Dinosaurs: The History and evolution of modern birds. John Wilex & Sons Ltd., 2011, S. 187–203 (Onlinepublikation), ISBN 978-0-470-65666-2.
- Delphine Angst und Eric Buffetaut: Palaeobiology of Giant Flightless Birds. Oxford, 2017, S. 1–282 (S. 133–160).
Einzelnachweise
- Herculano M. F. Alvarenga und Elizabeth Höfling: Systematic revision of the Phorusrhacidae (Aves: Ralliformes). Papéis Avulsos de Zoologia 43 (4), 2003, S. 55–91, PDF.
- Luis M. Chiappe und Sara Bertelli: Skull morphology of giant terror birds. These monstrous birds were probably more agile and less portly than previously thought. Nature 443, 2006, S. 929.
- Federico Agnolín: Sistemática y filogenia de las aves fororracoideas (Gruiformes, Cariamae). Buenos Aires, Fundación de Historia Natural Félix de Azara, 2009, S. 1–78.
- Delphine Angst, Eric Buffetaut, Christophe Lécuyer und Romain Amiot: A new method for estimating locomotion type in large ground birds. Palaeontology 59 (2), 2016, S. 217–223, doi:10.5061/dryad.609j4.
- Federico J. Degrange: Hind limb morphometry of terror birds (Aves, Cariamiformes, Phorusrhacidae): functional implications for substrate preferences and locomotor lifestyle. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 106, 2017, S. 257–276, doi:10.1017/S1755691016000256.
- R. Ernesto Blanco und Washington W. Jones: Terror birds on the run: a mechanical model to estimate its maximum running speed. Proceedings of the Royal Society B 272, 2005, S. 1769–1773, doi:10.1098/rspb.2005.3133.
- Federico J. Degrange, Claudia P. Tambussi, Karen Moreno, Lawrence M. Witmer und Stephen Wroe: Mechanical Analysis of Feeding Behavior in the Extinct „Terror Bird“ Andalgalornis steulleti (Gruiformes: Phorusrhacidae). PLoS ONE 5 (8), 2010, S. e11856, doi:10.1371/journal.pone.0011856.
- Claudia P. Tambussi, Ricardo de Mendoza, Federico J. Degrange und Mariana B. Picasso: Flexibility along the Neck of the Neogene Terror Bird Andalgalornis steulleti (Aves Phorusrhacidae). PLoS ONE 7 (5), 2012, S. e37701, doi:10.1371/journal.pone.0037701.
- Norma L. Nasif, Graciela Esteban, und Pablo E. Ortiz: Novedoso hallazgo de egagrópilas en el Mioceno tardío, Formación Andalhuala, provincia de Catamarca, Argentina. Serie Correlación Geológica 25, 2009, S. 105–114.
- Claudia I. Montalvo und Fernando J. Fernández: Review of the actualistic taphonomy of small mammals ingested by South American predators. Its importance in the interpretation of the fossil record. Publicación Electrónica de la Asociación Paleontológica Argentina 19 (1), 2019, S. 18–46, doi:10.5710/PEAPA.11.03.2019.275.
- Trevor H. Worthy, Federico J. Degrange, Warren D. Handley und Michael S. Y. Lee: The evolution of giant flightless birds and novel phylogenetic relationships for extinct fowl (Aves, Galloanseres). Royal Society open science 4, 2017, S. 170975, doi:10.1098/rsos.170975.
- Florentino Ameghino: Enumeración de las aves fósiles de la República Argentina. Revista Argentina Historia Natural 1, 1891, S. 441–453 ().
- Charles W. Andrews: On the extinct birds of Patagonia. – I. The skull and skeleton f Phororhacos inflatus Ameghino.. Transactions of the Zoological Society of London 15 (3), 1899, S. 55–86 ().
- B. C. Livezey und R. L. Zusi: Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. Zoological Journal of the Linnean Society 149, 2007, S. 1–95.
- Robert L. Carroll: Paläontologie und Evolution der Wirbeltiere, Thieme, Stuttgart, 1993, ISBN 3-13-774401-6.
- Shannon J. Hackett, Rebecca T. Kimball, Sushma Reddy, Rauri C. K. Bowie, Edward L. Braun, Michael J. Braun, Jena L. Chojnowski, W. Andrew Cox, Kin-Lan Han, John Harshman,Christopher J. Huddleston, Ben D. Marks, Kathleen J. Miglia, William S. Moore, Frederick H. Sheldon, David W. Steadman, Christopher C. Witt und Tamaki Yuri: A Phylogenomic Study of Birds Reveals Their Evolutionary History. Science 320 (5884), 2008, S. 1763–1768, doi:10.1126/science.1157704.
- Federico J. Degrange und Claudia P. Tambussi: Re-Examination of Psilopterus lemoinei (Aves, Phorusrhacidae), a Late Early Miocene Little Terror Bird from Patagonia (Argentina). Journal of Vertebrate Paleontology 31 (5), 2011, S. 1080–1092, doi:10.1080/02724634.2011.595466.
- Sara Bertelli, Luis M. Chiappe und Claudia Tambussi: A new Phorusrhacid (Aves: Cariamae) from the Middle Miocene of Patagonia, Argentina. Journal of Vertebrate Paleontology 27 (2), 2007, S. 409–419.
- Herculano Alvarenga, Luis Chiappe und Sara Bertelli: Phorusracids: The Terror birds. In: Gareth Dyke und Gary Kaiser (Hrsg.): Living Dinosaurs: The History and evolution of modern birds. John Wilex & Sons Ltd., 2011, S. 187–203.
- Federico J. Degrange, Claudia P. Tambussi, Matías L. Taglioretti, Alejandro Dondas, Fernando Scaglia. A new Mesembriornithinae (Aves, Phorusrhacidae) provides new insights into the phylogeny and sensory capabilities of terror birds. Journal of Vertebrate Paleontology 35 (2), 2015, S. e912656, doi:10.1080/02724634.2014.912656.
- Federico L. Agnolin und Pablo Chafrat: New fossil bird remains from the Chichinales Formation (Early Miocene) of northern Patagonia, Argentina. Annales du Paléontologie 101, 2015, S. 87–94, doi:10.1016/j.annpal.2015.02.001.
- Cécile Mourer-Chauviré, Rodolphe Tabuce, M’hammed Mahboubi, Mohammed Adaci und Mustapha Bensalah: A Phororhacoid bird from the Eocene of Africa. Naturwissenschaften 98, 2011, S. 815–823, doi:10.1007/s00114-011-0829-5.
- Delphine Angst, Eric Buffetaut, Christophe Lécuyer und Romain Amiot: „Terror Birds“ (Phorusrhacidae) from the Eocene of Europe Imply Trans-Tethys Dispersal. PLoS ONE 8 (11), 2013, S. e80357, doi:10.1371/journal.pone.0080357.
- Eric Buffetaut und Delphine Angst: Macrornis tanaupus Seeley, 1866: an enigmatic giant bird from the upper Eocene of England. Geological Magazine 158 (6), 2021, S. 1129–1134, doi:10.1017/S0016756820001466.
- Cécile Mourer-Chauviré: Première indication de la présence de phorusrhacidés, famille d'oiseaux géants d'Amérique du Sud, dans la Tertiaire Européens: Ameghinornis nov. gen. (Aves, Ralliformes) des phosphorites du Quercy, France. Géobio 14 (5), 1981, S. 637–647.
- Gerald Mayr: „Old World phorusrhacids“ (Aves, Phorusrhacidae): a new look at Strigogyps („Aenigmavis“) sapea (Peters 1987). PaleoBios 25, 2005, S. 11–16.
- Dieter S. Peters: The fossil family Ameghinornithidae (Mourer-Chauviré 1981): a short synopsis. Journal of Ornithology 148 (1), 2007, S. 25–28, doi:10.1007/s10336-006-0095-z.
- Marcos M. Cenizo: Review of the putative Phorusrhacidae from the Cretaceous and Paleogene of Antarctica: new records of ratites and pelagornithid birds. Polish Polar Research 33 (3), 2012, S. 225–244, doi:10.2478/v10183-012-0014-3.
- Gerald Mayr: Avian Evolution: The Fossil Record of Birds and its Paleobiological Significance. John Wiley & Sons, 2016. ISBN 978-1-119-02073-8 (Abschnitt „Were there phorusrhacids outside the Americas?“).
- Federico L. Agnolin: Brontornis burmeisteri Moreno & Mercerat, un Anseriformes (Aves) gigante del Mioceno Medio de Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales Nueva Serie 9, 2007, S. 15–25.
- Washington Jones, Andrés Rinderknecht, Herculano Alvarenga, Felipe Montenegro und Martín Ubilla: The last terror birds (Aves, Phorusrhacidae): new evidence from the late Pleistocene of Uruguay. Paläontologische Zeitschrift 92, 2018, S. 365–372, doi:10.1007/s12542-017-0388-y.
- Federico J. Degrange: A Revision of Skull Morphology In Phorusrhacidae (Aves, Cariamiformes). Journal of Vertebrate Paleontology 40 (6), 2020, S. e1848855, doi:10.1080/02724634.2020.1848855.
- Bryan Patterson: A new phororhacoid bird from the Deseado Formation of Patagonia. Geological Series of Field Museum of Natural History 8 (8), 1941, S. 49–54 ().
- Herculano Marcos Ferraz de Alvarenga: Uma gigantesca ave fóssil do Cenozóico brasileiro: Physornis brasiliensis sp. n. Anais da Academia Brasileira de Ciências 54, 1982, S. 697–712.
- Herculano Marcos Ferraz de Alvarenga: Paraphysornis. novo gênero para Physornis brasiliensis Alvarenga, 1982 (Aves: Phorusrhacidae). Anais da Academia Brasileira de Ciências 65, 1993, S. 403–406.
- Federico J. Degrange, Jorge I. Noriega und Juan I. Areta: Diversity and paleobiology of the Santacrucian birds. In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene paleobiology in Patagonia: High-latitude paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 138–155.
- Federico J. Degrange, Drew Eddy, Pablo Puerta und Julia Clarke: New skull remains of Phorusrhacos longissimus (Aves, Cariamiformes) from the Miocene of Argentina: implications for the morphology of Phorusrhacidae. Journal of Paleontology 93 (6), 2019, S. 1221–1233, doi: 10.1017/jpa.2019.53.
- Juan M. Diederle und Jorge I. Noriega: New records of birds from the Santa Cruz Formation (Early–Middle Miocene) at the Río Santa Cruz valley, Patagonia, Argentina. Publicación Electrónica de la Asociación Paleontológica Argentina 19 (2), 2019, S. 55–61.
- Sara Bertelli, Luis M. Chiappe und Claudia Tambussi: A new Phorusrhacid (Aves: Cariamae) from the Middle Miocene of Patagonia, Argentina. Journal of Vertebrate Paleontology 27 (2), 2007, S. 409–419, [[doi:10.1671/0272-4634(2007)27[409:ANPACF]2.0.CO;2]].
- Lucas Kraglievich: Contribución al conocimiento de las aves fósiles de la época araucoentrerriana. Physis: Revista de la Sociedad Argentina de Ciencias Naturales 10, 1931, S. 304–315.
- Lucas Kraglievich: Una gigantesca ave fósil del Uruguay, Devincenzia gallinali n. gen. n. sp., tipo de una nueva familia, Devincenziidae, del Orden Stereornithes. Anales del Museo de Historia Natural de Montevideo Serie II 3, 1932, S. 323–353.
- Pierce Brodkorb: A giant flightless bird from the Pleistocene of Florida. The Auk 80 (2), 1963, S. 111–115.
- Francisco J. Prevosti, Analía Forasiepi und Natalia Zimicz: The Evolution of the Cenozoic Terrestrial Mammalian Predator Guild in South America: Competition or Replacement? Journal of Mammalian Evolution 20, 2013, S. 3–21, doi:10.1007/s10914-011-9175-9.
- Sergio Daniel Tarquini, Sandrine Ladevèze und Francisco Juan Prevosti: The multicausal twilight of South American native mammalian predators (Metatheria, Sparassodonta). Scientific Reports 12, 2022, S. 1224, doi:10.1038/s41598-022-05266-z.
- Kateryn Pino, Paulo Vallejos-Garrido, Nicolás Espinoza-Aravena, Rebecca B. Cooper, Daniele Silvestro, Cristián E. Hernández und Enrique Rodríguez-Serrano: Regional landscape change triggered by Andean uplift: The extinction of Sparassodonta (Mammalia, Metatheria) in South America. Global and Planetary Change, 2022, S. 103758, doi:10.1016/j.gloplacha.2022.103758.
- Claudia P. Tambussi, M. Ubilla und D. Perea: The youngest large carnassial bird (Phorusrhacidae, Phorusrhacinae) from South America (Pliocene–Early Pleistocene of Uruguay). Journal of Vertebrate Paleontology 19 (2), 1999, S. 404–406.
- Herculano Alvarenga, Washington Jones und Andrés Rinderknecht: The youngest record of phorusrhacid birds (Aves, Phorusrhacidae) from the late Pleistocene of Uruguay. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 256/2, 2010, S. 229–234.
- Raúl Ignacio Vezzosi: First record of Procariama simplex Rovereto, 1914 (Phorusrhacidae, Psilopterinae) in the Cerro Azul Formation (upper Miocene) of La Pampa Province; remarks on its anatomy, palaeogeography and chronological range. Alcheringa: An Australasian Journal of Palaeontology 36 (2). 2012, S. 157–169, doi:10.1080/03115518.2011.597657.
- Claudia p: Tambussi. Palaeoenvironmental and faunal inferences based on the avian fossil record of Patagonia and Pampa: what works and what does not. Biological Journal of the Linnean Society 103, 2011, S. 458–474.
- Federico L. Agnolin: La posición sistemática de Hermosiornis (Aves, Phororhacoidea) y sus implicancias filogenéticas. Revista del Museo Argentino de Ciencias Naturales Nueva Serie 15 (1), 2013, S. 39–60.
- Florentino Ameghino: Enumeración sistemática de las espécies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos Eocenos de la Patagonia austral y depositados en el Museo de La Plata. Boletin del Museo de La Plata 1, 1887, S. 1–26 ().
- T. S. Palmer: Index Generum Mammalium: A List of the Genera and Families of Mammals. North American Fauna 23, 1904, S. 620–621 ().
- Florentino Ameghino: Contribuición al conocimiento de los mamíferos fósiles de la República Argentina. Actas Academia Nacional Ciencias de Cordoba 6, 1889, S. 1–1028 (S. 659) ().
- Florentino Ameghino: Mamíferos y aves fósiles Argentinos: espécies nuevas: adiciones y correciones. Revista Argentina Historia Natural 1, 1891, S. 240–259 ().
- Richard Lydekker: On the extinct giant birds of Argentina. Ibis 5, 1893, S. 40–47 ().
- Luis M. Chiappe und Miguel F. Soria: Phororhacos Ameghino, 1889 (Aves, Gruiformes): proposed conservation. Bulletin of Zoological Nomenclature 47 (3), 1990, S. 198–201 ().
- International Commission on Zoological Nomenclature: Opinion 1687. Phorusrhacos Ameghino, 1887 (Aves, Gruiformes): not suppressed. Bulletin of Zoological Nomenclature 49 (2), 1992, S. 176–177 ().
- P. Brodkorb: Catalogue of fossil birds, Part III (Ralliformes, Ichthyornithiformes, Charadriiformes). Bulletin of Florida State Museum 2, 1967, S. 99–220 ().
- Eric Buffetaut: Phororhacoidea or Phorusrhacoidea? A note on the nomenclature of the “terror birds”. Annales de Paléontologie 99 (2), 2013, S. 157–161, doi:10.1016/j.annpal.2013.02.004.
- Eric Buffetaut: Who discovered the Phorusrhacidae? An episode in the history of avian palaeontology. In: Ursula B. Göhlich, Andreas Kroh (Hrsg.): Paleornithological Research 2013. Proceedings of the 8th International Meeting of the Society of Avian Paleontology and Evolution. Verlag Naturhistorisches Museum Wien, 2013. S. 122–132.
- Francisco P. Moreno und Alcide Mercerat: Catálogo de los pájaros fósiles de la República Argentina conservados en el Museo de La Plata. Anales del Museo de La Plata, Paleontología Argentina 1, 1891, S. 7–71 ().
- Terror Bird auf ARK Survival Evolved Wiki (abgerufen am 7. Februar 2021).
- Tim Haines: Die Erben der Saurier. Im Reich der Urzeit. Köln, 2002, S. 182–219 ISBN 9783802514616.