Strauße (Gattung)
Die Strauße (Struthio) sind eine Gattung großer, flugunfähiger Vögel. Sie stehen innerhalb der im Deutschen gleichnamigen Familie der Strauße (Struthionidae) und der Ordnung der Struthioniformes. Sowohl Familie als auch Ordnung gelten als rezent monotypisch. Die Vögel kommen in den relativ ariden Gebieten in Afrika vor. Es werden zwei heute lebende Arten unterschieden, der Afrikanische Strauß (Struthio camelus) und der Somalistrauß (Struthio molybdophanes). Daneben existiert eine Reihe fossiler und subfossiler Funde aus Asien und Europa, die der Gattung zugerechnet werden.
| Strauße | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Afrikanische Strauße (Struthio camelus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Ordnung | ||||||||||||
| Struthioniformes | ||||||||||||
| Latham, 1790 | ||||||||||||
| Wissenschaftlicher Name der Familie | ||||||||||||
| Struthionidae | ||||||||||||
| Vigors, 1825 | ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Struthio | ||||||||||||
| Linnaeus, 1758 |
Merkmale

Die Männchen der Strauße sind bis zu 275 Zentimeter hoch und haben ein Gewicht bis zu 156 Kilogramm. Weibchen sind kleiner: Sie sind 175 bis 190 Zentimeter hoch und 90 bis 110 Kilogramm schwer. Strauße sind damit die größten Vögel der Welt. Sie haben einen ovalen, horizontal ausgerichteten, schweren Rumpf und lange, unbefiederte Beine mit zwei nach vorn gerichteten, dicken Zehen an den Füßen. Der Kopf ist klein und unbefiedert, der Hals lang und nur minimal befiedert. Die Augen sind relativ groß; der Schnabel ist kurz, flach und breit. Das Gefieder der Männchen ist größtenteils schwarz, das der Weibchen ist graubraun.[1]
Verbreitung
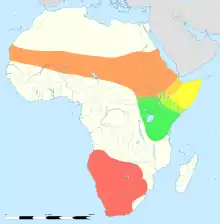
Die beiden heutigen Arten der Strauße kommen in Afrika vor und beschränken sich dort auf die Regionen südlich der Sahara. Der Afrikanische Strauß (Struthio camelus) besiedelt mit mehreren Unterarten ein größeres Gebiet im südlichen, östlichen und nördlichen Teil des Kontinentes. Der Somalistrauß (Struthio molybdophanes) ist lediglich im nordöstlichen Afrika präsent. Noch in historischer Zeit bewohnten die Vögel auch den westlichen Teil Asiens, enger genommen die Arabische Halbinsel. Hier war der Arabische Strauß verbreitet, eine Unterart des Afrikanischen Straußes, der in den 1960er Jahren aber ausgerottet wurde. Als bevorzugte Lebensräume dienen offene savannen- bis teils wüstenartige Landschaften. Die nördlichen und südlichen Populationen werden durch den zentral- und ostafrikanischen Regen- und Miombo-Waldgürtel voneinander getrennt.[2][3]
Lebensweise
Strauße kommen in Steppen, Savannen, offenen, trockenen Wäldern und Halbwüsten in Afrika vor. Sie haben sich stark an ein Leben in den offenen Landschaften angepasst und erreichen Laufgeschwindigkeiten von bis zu 70 km/h.[3] Prinzipiell sind sie Allesfresser, bevorzugen aber Grünpflanzen. Kleine Pflanzen werden aus dem Boden gezogen und mitsamt ihren Wurzeln gefressen. Strauße verbringen einen großen Teil des Tages mit der Nahrungssuche und beziehen über ihre Nahrung auch einen großen Teil der benötigten Feuchtigkeit.[1]
Fortpflanzung
Strauße sind polygam. Sowohl Männchen als auch Weibchen paaren sich mit mehreren Sexualpartnern. Als „Nest“ kratzt das Männchen eine Vertiefung in den Erdboden, in die mehrere (bis zu 18) Weibchen ihre Eier legen. Zuerst legt das Hauptweibchen in der Regel sieben bis zehn Eier in das Nest, darauf mehrere Nebenweibchen weitere Eier, so dass die gesamte Gelegegröße 20 bis 25 Eier umfassen kann. Als Standort des Nestes wird ein Ort gewählt, der dem brütenden Strauß einen guten Überblick über die Umgebung ermöglicht. Sowohl das Männchen als auch das Hauptweibchen bebrüten über einen Zeitraum von etwa sechs Wochen das Gelege. Drei Tage nach dem Schlüpfen verlassen die Jungstrauße das Nest und werden anschließend vom Männchen und vom Hauptweibchen geführt und beschützt. Selbständig sind sie mit einem Alter von etwa einem Jahr.[1]
Systematik
Allgemein
Innere Systematik der Urkiefervögel basierend auf genetischen Daten nach Urantówka et al. 2020[4]
|
Die Strauße sind eine Gattung aus der im Deutschen gleichnamigen Familie der Strauße (Struthionidae) innerhalb der Ordnung der Struthioniformes. Sowohl Familie als auch Ordnung gelten als rezent monotypisch. Der Gattung werden zwei Arten zugewiesen, deren Verbreitung heute auf Afrika beschränkt bleibt. Die Strutioniformes als höhere taxonomische Einheit werden den Urkiefervögeln (Palaeognathae) zugerechnet. Dadurch besteht eine engere Verwandtschaft mit anderen großen Laufvögeln wie den Emus (Dromaius) und Kasuaren (Casuarius) aus der Gruppe der Casuariiformes und den Nandus (Rhea) aus der Gruppe der Rheiformes. Als in jüngerer Zeit ausgestorbene Vertreter werden zudem die Elefantenvögel (Aepyornithiformes) und die Moas (Dinornithiformes) hinzugerechnet.[4][5][6] Darüber hinaus bestehen aus anatomischer Sicht engere Beziehungen zu einzelnen Formengruppen des Paläogens wie etwa den Lithornithiformes.[3]
Übersicht über die Gattungen und Arten
Die Gattung der Strauße wird heute in zwei Arten aufgegliedert. Diese umfassen den Afrikanischen Strauß (Struthio camelus) und den Somalistrauß (Struthio molybdophanes). Beide werden seit 2014 als eigenständig geführt, ihre genetische Trennung erfolgte im Pliozän vor 4 1 bis 3,6 Millionen Jahren.[2] Teilweise wird aufgrund molekulargenetischer Befunde diskutiert, ob die nördlichen und östlich-südlichen Unterarten des Afrikanischen Straußes ebenfalls einen Artstatus erhalten sollen.[7]
Daneben sind mehrere Fossilformen bekannt. Unter Hinzuziehung dieser gliedert sich die Familie der Strauße folgendermaßen:[8][3]
- Familie: Struthionidae Vigors, 1825
- Gattung: † Palaeostruthio Burchak-Abramovich, 1953
- † Palaeostruthio karatheodoris (Forsyth-Major, 1888)
- Gattung: † Pachystruthio Kretzoi, 1954
- † Pachystruthio dmanisensis (Вurchak-Abramovich & Vekua, 1990)
- † Pachystruthio pannonicus Kretzoi, 1954
- † Pachystruthio transcaucasicus (Burchak-Abramovich & Vekua, 1971)
- Gattung: Struthio Linnaeus, 1758
- † Struthio anderssoni Lowe, 1931
- † Struthio asiaticus Milne-Edwards, 1871 (Asiatischer Strauß)
- † Struthio barbarus Arambourg, 1979
- † Struthio brachydactylus Burchak-Abramovich, 1949
- Struthio camelus Linnaeus, 1758 (Afrikanischer Strauß)
- † Struthio chersonensis (Brant, 1873)
- † Struthio coppensi Mourer-Chauviré, Senut, Pickford & Mein, 1996
- † Struthio linxiaensis Hou, Zhou, Zhang & Wang, 2005
- Struthio molybdophanes Rothschild, 1919 (Arabischer Strauß)
- † Struthio novorossicus Alexejev, 1916
- † Struthio oldawayi Lowe, 1933
- † Struthio orlovi Kurochkin & Lungu, 1970
- † Struthio wimani Lowe, 1931
Die Eigenständigkeit einiger Arten ist momentan in Diskussion. Dadurch könnte etwa Struthio novorossicus identisch sein mit Struthio asiaticus, ebenso wäre Pachystruthio dmanisensis synonym zu Pachystruthio pannonicus. Einige Formen wie Pachystruthio dmanisensis oder Palaeostruthio karatheodoris wurden ursprünglich ebenfalls innerhalb der Gattung Struthio geführt, später aber ausgelagert. Struthio linxiaensis steht teilweise innerhalb der Untergattung Orientornis, die mitunter auch als eigenständige Gattung angesehen wird.[3]
Neben diesen bekannten, auf Skelettmaterial begründeten ausgestorbenen Vertretern wurden noch einige Formengruppen benannt, die auf fossilen Eierschalen basieren und als „Eiergattungen“ (oogenus) bezeichnet werden. Hierunter fallen beispielsweise die aus dem Miozän des südwestlichen Afrikas beschriebenen Gattungen Diamantornis, Namaornis und Tsondabornis, des Weiteren Psammornis, der über einen längeren Zeitraum vom Eozän bis zum Holozän im nördlichen Afrika und angrenzenden Gebieten belegbar ist, oder der im Pliozän und Pleistozän nachweisbare Struthiolithus, wahrscheinlich identisch zu Struthio.[9][10][11] Die Unterscheidung der einzelnen Formen erfolgt anhand der Gestaltung der Eierschalen, vornehmlich der Art der Poren. Voneinander abtrennbar sind hier zwei größere Einheiten, der „struthionide Typ“ und der „aepyornithoide Typ“. Letzterer, benannt nach den nur auf Madagaskar vorkommenden Elefantenvögeln, weist Poren auf, die in linear gerichteten Riefen auftreten, ersterer zeigt in fleckenartigen Eintiefungen angeordnete Poren. Es sind allerdings auch Übergangstypen und einzelne Sonderformen dokumentiert.[12][13]
In der höheren Gliederung werden die Struthionidae innerhalb der Struthioniformes eingeordnet. Diesen gehören folgende Familien an:
- Ordnung: Struthioniformes Latham, 1790
- Familie: † Eogruidae Wetmore, 1934
- Familie: † Ergilornithidae Kozlova, 1960
- Familie: † Palaeotididae Houde & Haubold, 1987
- Familie: Struthionidae Vigors, 1825
Die Palaeotididae mit Palaeotis als Typusform galten lange Zeit als mit den Straußen eng verbunden und gehörten somit zur Familie der Struthionidae. Hier bildeten sie die eigene Unterfamilie der Palaeotidinae.[14] Neuere Bearbeitungen sehen diese nun als eigenständige Familie an, deren genaue systematische Zuweisung innerhalb der Urkiefervögel aber unklar blieb.[15][16][17] Des Weiteren ordnete man sowohl die Eogruidae als auch die Ergilornithidae ursprünglich als Angehörige der Kranichvögel (Gruiformes) ein. Alle drei Gruppen wurden in einer Studie aus dem Jahr 2021 als Stammformen der Struthioniformes ausgewiesen. Allerdings formen die Eogruidae vermutlich eine paraphyletische Gruppe, welche die Ergilornithidae mit einschließt. Unklar bleibt die Position der Geranoididae, eine ausgestorbene Gruppe aus dem Paläogen Nordamerikas, die ebenfalls als urtümliche Kranichvögel angesehen wurden.[18] Auch hier besteht die Möglichkeit einer Zuordnung zu den Struthioniformes.[19][3]
Stammesgeschichte

Der Fossilbericht der Strauße verteilt sich auf Afrika und Eurasien. Die Funde straußenartiger Vögel reichen bis in das Paläogen zurück. Unter der Voraussetzung einer Zuordnung von Remiornis zu den Straußenartigen könnte ihr frühestes Auftreten bereits im ausgehenden Paläozän erfolgt sein. Das überwiegend stark fragmentierte Fundmaterial, das etwa von Rivecourt im nördlichen Frankreich dokumentiert ist,[20] erschwert aber eine genaue systematische Zuweisung. Als ältester eindeutiger Vertreter wird gegenwärtig Galligeranoides betrachtet, ein Angehöriger der Palaeotididae. Von dieser Form sind einzelne Beinknochen aus Saint-Papoul im südwestlichen Frankreich belegt, wo sie zusammen mit dem riesenhaften Gastornis auftrat. Die Reste dürften zwischen 56 und 51 Millionen Jahre alt sein und gehören damit dem Unteren Eozän an.[21] Deutlich besser dokumentiert ist der nahe verwandte Palaeotis. Mehrere nahezu vollständige Skelette liegen unter anderem aus den Faunengemeinschaften des Geiseltals und der Grube Messel vor. Beide Lokalitäten datieren in das Mittlere Eozän. Die nur rund 1 m großen Tiere lebten hier in tropischen Regenwäldern und waren wohl noch nicht an hohe Laufgeschwindigkeiten angepasst, worauf auch die dreizehigen Füße verweisen.[14][15][3]
In etwa dem gleichen Zeitraum erscheinen erstmals die Eogruidae und die Ergilornithidae. Beide Gruppe sind überwiegend aus dem östlichen und zentralen Asien dokumentiert. Ergilornis als Charakterform letzterer ist in großer Anzahl aus der Ergilin-Dzo-Formation in der Mongolei nachgewiesen.[22] Hier trat auch Eogrus als Nominatgattung ersterer auf, weitere Fundpunkte umfassen etwa die Irdin-Manha-Formation in der Mongolei oder die Obayla- und die Kustovskaya-Formation im östlichen Kasachstan. Diese frühen Forman aus dem Eozän wiesen noch einen dreizehigen Fuß auf. Nach einer Überlieferungslücke im Oligozän erwarben die Eogruidae und Ergilornithidae im Miozän eine starke Anpassung an eine schnelle Fortbewegung in offenen Landschaften. Erkennbar ist dies unter anderem an der Reduktion auf zwei funktionale Zehen je Fuß, analog zu den heutigen Straußen. Das betrifft beispielsweise Sinoergilornis aus dem Linxia-Becken in der chinesischen Provinz Gansu und Urmiornis, der mit Fundpunkten im zentralen und südlichen Asien sowie im östlichen Europa recht weit verbreitet war.[23][19][3]

Die Struthionidae sind erstmals im Unteren Miozän nachweisbar. Einer der frühesten Nachweise bildet Struthio coppensi von Elisabethfeld in Namibia. Es sind aber bisher nur einzelne Beinknochen bekannt, die auf einen zweizehigen Fuß hindeuten. Insgesamt war die Art noch kleiner und graziler gebaut als heutige Strauße. Die frühen Funde aus dem südwestlichen Afrika lassen einen Ursprung der Gattung auf dem Kontinent annehmen.[24] Weitere Fossilreste von Struthio sind aus dem Mittleren Miozän von Fort Ternan in Kenia aufgefunden wurden.[25] Etwa zeitgleich tritt die Form in Anatolien und damit erstmals außerhalb von Afrika auf. Die Reste können wohl Struthio brachydactylus zugeordnet werden.[26] In der Folgezeit erreichte Struthio eine sehr weite Verbreitung. Neben Afrika als Ursprungsregion sind Funde sowohl im östlichen Asien als auch in Europa aufgedeckt worden. Einige Arten des Pleistozäns wiesen dabei riesige Ausmaße auf, die die der heutigen Formen weit übertrafen. Dies trifft etwa auf Struthio oldawayi aus der Olduvai-Schlucht in Tansania zu, dessen Knochenmaße um rund 33 % größer sind als jene eines rezenten männlichen Individuums. Bei der nordafrikanischen Form Struthio barbarus liegen die Werte immerhin noch um 20 % über denen der gegenwärtigen Vertreter. Für den aus Choukoutien in China beschriebenen Struthio anderssoni werden wiederum Gewichtsschätzungen von rund 270 kg angegeben, basierend auf der Größe des Oberschenkelknochens und einiger Eierfunde.[27][28] In Eurasien starb Struthio vermutlich erst im Oberen Pleistozän aus.[29][3]
Eurasische Funde von Straußen umfassen darüber hinaus mit Palaeostruthio und Pachystruthio zwei weitere Gattungen mit teils gewaltigen Ausmaßen. Palaeostruthio trat überwiegend im Oberen Miozän im südöstlichen und östlichen Europa auf, wie es etwa einzelne Fußknochen aus Kalimantsi und Hadzhidimovo in Bulgarien oder Halswirbel aus Kerassia in Griechenland zeigen.[30][31] Pachystruthio wiederum ist weitgehend aus dem Altpleistozän überliefert. Aus der Fundstelle Dmanissi in Georgien erhaltene Knochenreste werden Pachystruthio dmanisensis zugeordnet, einem bis zu 450 kg schweren Tier, das einen der größten bekannten Vögeln überhaupt repräsentiert.[32] Des Weiteren dienten ein Zehenknochen und beinahe einhundert Eierschalen aus Kisláng in Ungarn zur Etablierung der Art Pachystruthio pannonicus.[33] Der gleichen Gattung eingegliedert werden auch Beinknochen aus dem Nihewan-Becken in der chinesischen Provinz Hebei, die zu einem vermutlich rund 300 kg schweren Individuum gehören. Die Altersstellung entspricht weitgehend der von Dmanissi.[8][3]
Quellen
- David W. Winkler, Shawn M. Billerman, Irby J. Lovette: Bird Families of the World: A Guide to the Spectacular Diversity of Birds. Lynx Edicions (2015), ISBN 978-8494189203. Seite 36.
- Stephanie Freitag, Terence J. Robinson: Phylogeographic patterns in mitochondrial DNA of the Ostrich (Struthio camelus). In: The Auk Band 110 (3), 1993, S. 614–622.
- Klara Widrig, Daniel J. Field: The Evolution and Fossil Record of Palaeognathous Birds (Neornithes: Palaeognathae). In: Diversity Band 14, 2022, S. 105, doi:10.3390/d14020105.
- Adam Dawid Urantówka, Aleksandra Kroczak, Paweł Mackiewicz: New view on the organization and evolution of Palaeognathae mitogenomes poses the question on the ancestral gene rearrangement in Aves. In: BMC Genomics Band 21, 2020, S. 874, doi:10.1186/s12864-020-07284-5.
- Kieren J. Mitchell, Bastien Llamas, Julien Soubrier, Nicolas J. Rawlence, Trevor H. Worthy, Jamie Wood, Michael S. Y. Lee, Alan Cooper: Ancient DNA reveals elephant birds and kiwi are sister taxa and clarifies ratite bird evolution. Science 344 (6186), 2014, S. 898–900, doi:10.1126/science.1251981.
- Takahiro Yonezawa, Takahiro Segawa, Hiroshi Mori, Paula F. Campos, Yuichi Hongoh, Hideki Endo, Ayumi Akiyoshi, Naoki Kohno, Shin Nishida, Jiaqi Wu, Haofei Jin, Jun Adachi, Hirohisa Kishino, Ken Kurokawa, Yoshifumi Nogi, Hideyuki Tanabe, Harutaka Mukoyama, Kunio Yoshida, Armand Rasoamiaramanana, Satoshi Yamagishi, Yoshihiro Hayashi, Akira Yoshida, Hiroko Koike, Fumihito Akishinonomiya, Eske Willerslev, Masami Hasegawa: Phylogenomics and Morphology of Extinct Paleognaths Reveal the Origin and Evolution of the Ratites. Current Biology 27 (1), 2017, S. P68–77, doi:10.1016/j.cub.2016.10.029.
- Joshua M. Miller, Sara Hallager, Steven L. Monfort, John Newby, Kelley Bishop, Scott A. Tidmus, Peter Black, Bill Houston, Conrad A. Matthee, Robert C. Fleischer: Phylogeographis analysis of nuclear and mtDNA support subspecies designation in ostrich (Struthio camelus). In: Conservation Genetics Band 12, 2011, S. 423–431, doi:10.1007/s10592-010-0149-x.
- Eric Buffetaut, Delphine Angst: A Giant Ostrich from the Lower Pleistocene Nihewan Formation of North China, with a Review of the Fossil Ostriches of China. In: Diversity Band 13, 2021, S. 47, doi:10.3390/d13020047.
- Martin Pickford: New Ratite Eggshells from the Miocene of Namibia. In: Communications of the Geological Survey of Namibia Band 15, 2014, S. 70–90.
- Faysal Bibi, Alan B. Shabel, Brian P. Kraatz, Thomas A. Stidham: New Fossil Ratite (Aves: Palaeognathae) Eggshell Discoveries from the Late Miocene Baynunah Formation of the United Arab Emirates, Arabian Peninsula. In: Palaeontologia Electronica Band 9 (1), 2006, S. 2A ().
- Eric Buffetaut: The Enigmatic Avian Oogenus Psammornis: A Review of Stratigraphic Evidence. In: Diversity Band 14 (2), 2022, S. 123, doi:10.3390/d14020123.
- E. G. Franz Sauer: Ratite eggshells and phylogenetic questions. In: Bonner Zoologische Beiträge Band 23, 1972, S. 3–48.
- Cécile Mourer-Chauviré, Brigitte Senut, Martin Pickford, Pierre Mein, Yannicke Dauphin: Ostrich legs, eggs and phylogenies. In: South African Joumal of Science Band 92, 1996, S. 492–495.
- Peter Houde, Hartmut Haubold: Palaeotis weigelti restudied: a small Middle Eocene ostrich (Aves: Struthioniformes). In: Palaeovertebrata. Band 17, 1987, S. 27–42.
- Dieter Stefan Peters: Ein vollständiges Skelett von Palaeotis weigelti (Aves, Palaeognathae). In: Courier Forschungsinstitut Senckenberg Band 107, 1988, S. 223–233.
- Gareth J. Dyke, Marcel van Tuinen: The evolutionary radiation of modern birds (Neornithes): reconciling molecules, morphology and the fossil record. In: Zoological Journal of the Linnean Society Band 141, 2004, S. 153–177.
- Gerald Mayr: Hindlimb morphology of Palaeotis suggests palaeognathous affinities of the Geranoididae and other “crane-like” birds from the Eocene of the Northern Hemisphere. In: Acta Palaeontologica Polonica Band 64 (4), 2019, S. 669–678, doi:10.4202/app.00650.2019.
- Gerald Mayr: On the taxonomy and osteology of the Early Eocene North American Geranoididae (Aves, Gruoidea). In: Swiss Journal of Palaeontology Band 135, 2016, S. 315–325, doi:10.1007/s13358-016-0117-2.
- Gerald Mayr, Nikita Zelenkov: Extinct crane-like birds (Eogruidae and Ergilornithidae) from the Cenozoic of Central Asia are indeed ostrich precursors. In: Ornithology Band 138, 2021, S. 1–15, doi:10.1093/ornithology/ukab048.
- Eric Buffetaut, Gaël de Ploëg: Giant Birds from the Uppermost Paleocene of Rivecourt (Oise, Northern France). In: Boletim do Centro Português de Geo-História e Pré-História Band 2 (1), 2020, S. 29–33.
- Estelle Bourdon, Cecilie Mourer-Chauvire und Yves Laurent: Early Eocene birds from La Borie, southern France. In: Acta Palaeontologia Polonica Band 61 (1), 2016, S. 175–190, doi:10.4202/app.00083.2014.
- Julia A. Clarke, Mark A. Norell, Demberelyin Dashzeveg: New Avian Remains from the Eocene of Mongolia and the Phylogenetic Position of the Eogruidae (Aves, Gruoidea). In: American Museum Novitates Band 3494, 2005, S. 1–17.
- Grace Musser, Zhiheng Li, Julia A. Clarke: A new species of Eogruidae (Aves: Gruiformes) from the Miocene of the Linxia Basin, Gansu, China: Evolutionary and climatic implications. In: The Auk Band 137, 2019, S. 1–1, doi:10.1093/auk/ukz067.
- Cécile Mourer-Chauviré, Brigitte Senut, Martin Pickford, Pierre Mein: Le plus ancien représentant du genre Struthio (Aves, Struthionidae), Struthio coppensi n. sp., du Miocène inférieur de Namibie. In: Comptes Rendus de l'Académie des Sciences Band 322, 1996, S. 325–332.
- Leona M. Leonard, Gareth J. Dyke, Cyril A.Walker: New specimens of a fossil ostrich from the Miocene of Kenya. In: Journal of African Earth Sciences Band 45 (4–5), 2006, S. 391–394.
- E. G. Franz Sauer: A Miocene ostrich from Anatolia. In: Ibisd Band 121 (4), 1979, S. 494–501.
- Cécile Mourer-Chauviré, Denis Geraads: The Struthionidae and Pelagornithidae (Aves: Struthioniformes, Odontopterygiformes) from the late Pliocene of Ahl Al Oughlam, Morocco. In: Oryctos Band 7, 2008, S. 169–194.
- Eric Buffetaut 1 and Delphine Angst: How large was the giant ostrich of China? In: EVOLUÇÃO - Revista de Geistória e Pré-História Band 2 (1), 2017, S. 2, 6–8.
- Sonal Jain, Niraj Rai, Giriraj Kumar, Parul Aggarwal Pruthi, Kumarasamy Thangaraj, Sunil Bajpai, Vikas Pruthi: Ancient DNA Reveals Late Pleistocene Existence of Ostriches in Indian Sub-Continent. In: PLoS ONE Band 12 (3), 2017, S. e0164823, doi:10.1371/journal.pone.0164823.
- Zlatozar Boev, Nikolai Spassov: First record of ostriches (Aves, Struthioniformes, Struthionidae) from the late Miocene of Bulgaria with taxonomic and zoogeographic discussion. In: Geodiversitas. Band 31 (3), 2009, S. 493–507.
- Panagiotis Kampouridis, Dimitrios Michailidis, Nikolaos Kargopoulos, Socrates Roussiakis, Georgios Theodorou: First description of an ostrich from the late Miocene of Kerassia (Euboea, Greece): remarks on its cervical anatomy. In: Historical Biology. An International Journal of Paleobiology Band 33 (10), 2021, S. 2228–2235, doi:10.1080/08912963.2020.1779252.
- Nikita V. Zelenkov, Alexander V. Lavrov, Dmitry B. Startsev, Innessa A. Vislobokova, Alexey V. Lopatin: A giant early Pleistocene bird from eastern Europe: unexpected component of terrestrial faunas at the time of early Homo arrival. In: Journal of Vertebrate Paleontology Band 39 (2), 2019, S. e1605521, doi:10.1080/02724634.2019.1605521.
- Miklós Kretzoi: Bericht über die Calabrische (Villafranchische) Fauna von Kisláng, Kom. Fejér. Jahresberichte der Ungarischen Geologischen Anstalt, 1954, S. 239–264 ().